
Enhancement of Anti-Proliferative Activity of the Extracts from Dehulled Adlay by Fermentation with Bacillus subtilis




Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials and Chemicals
2.2. Preparation of Different Extracts from Samples
2.3. Cell Culture
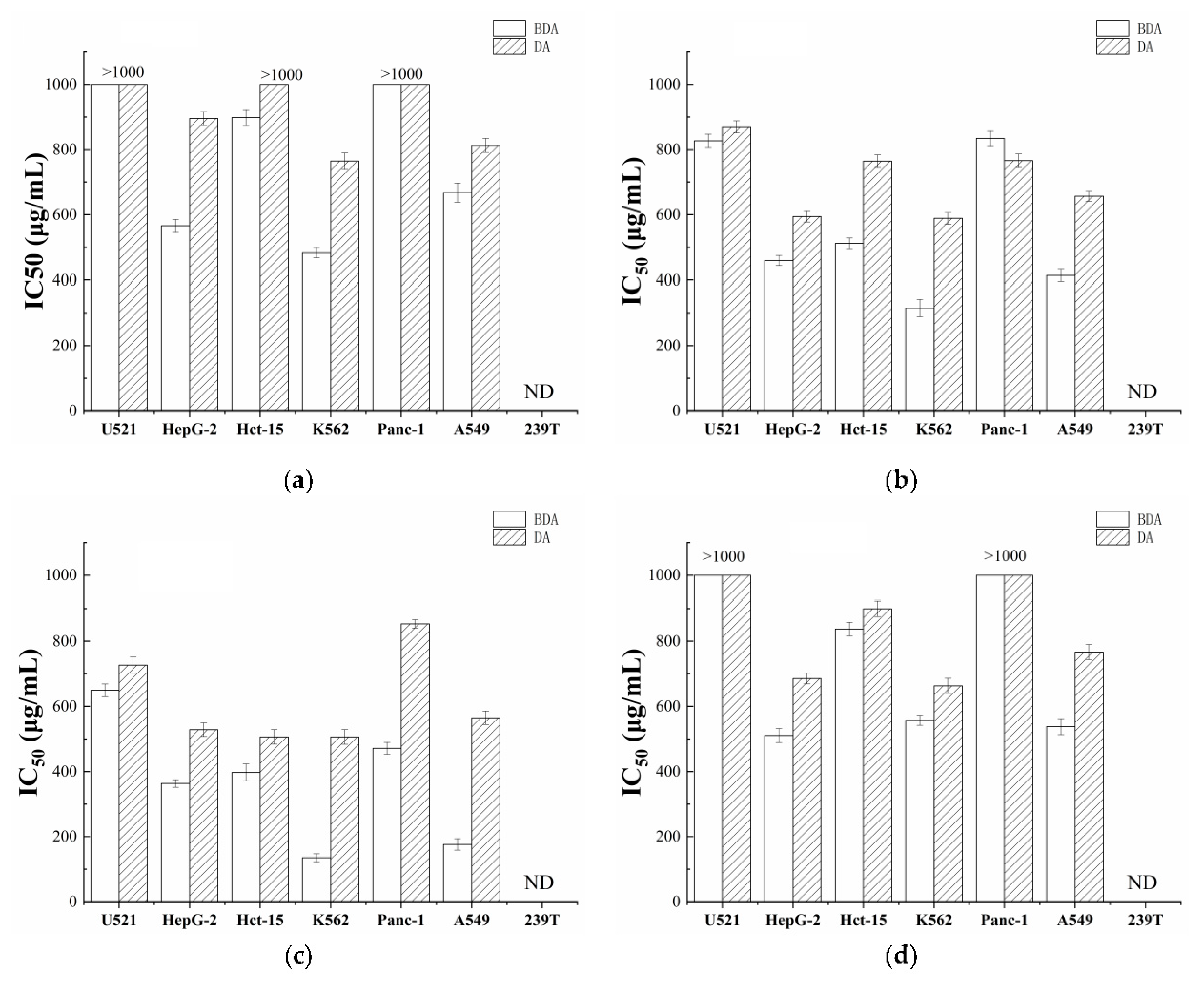
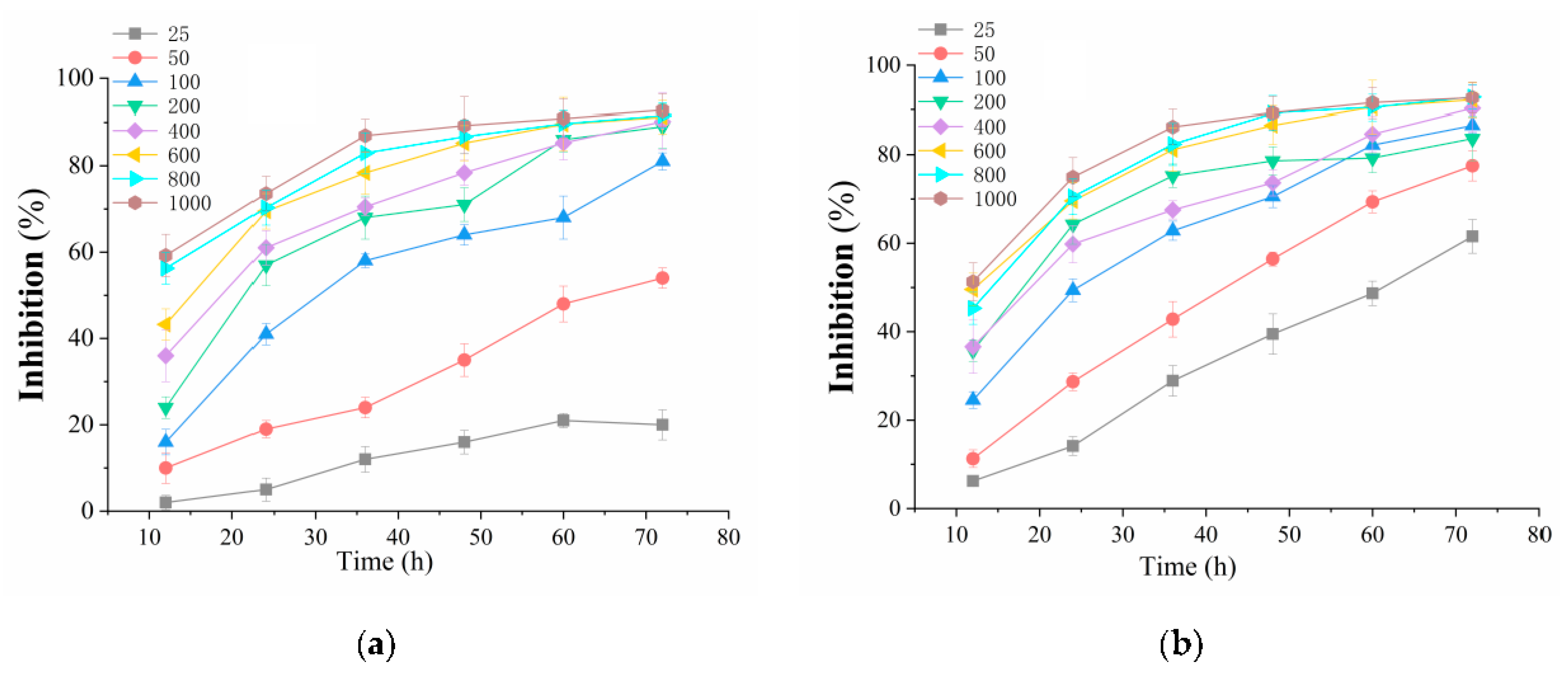
2.4. Cell Viability Assay
2.5. Determination of Bioactive Compositions
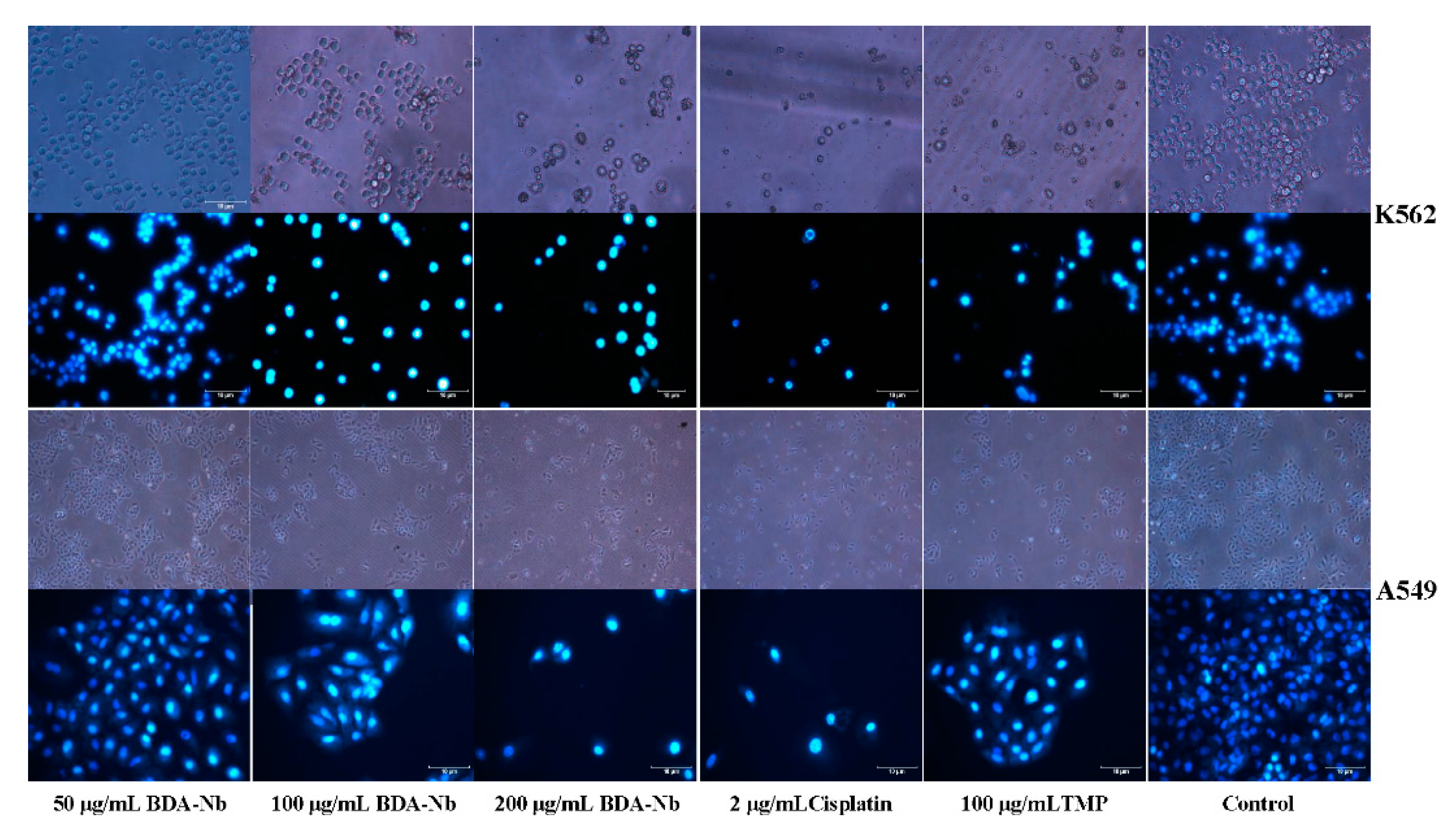
2.6. Morphological Observations
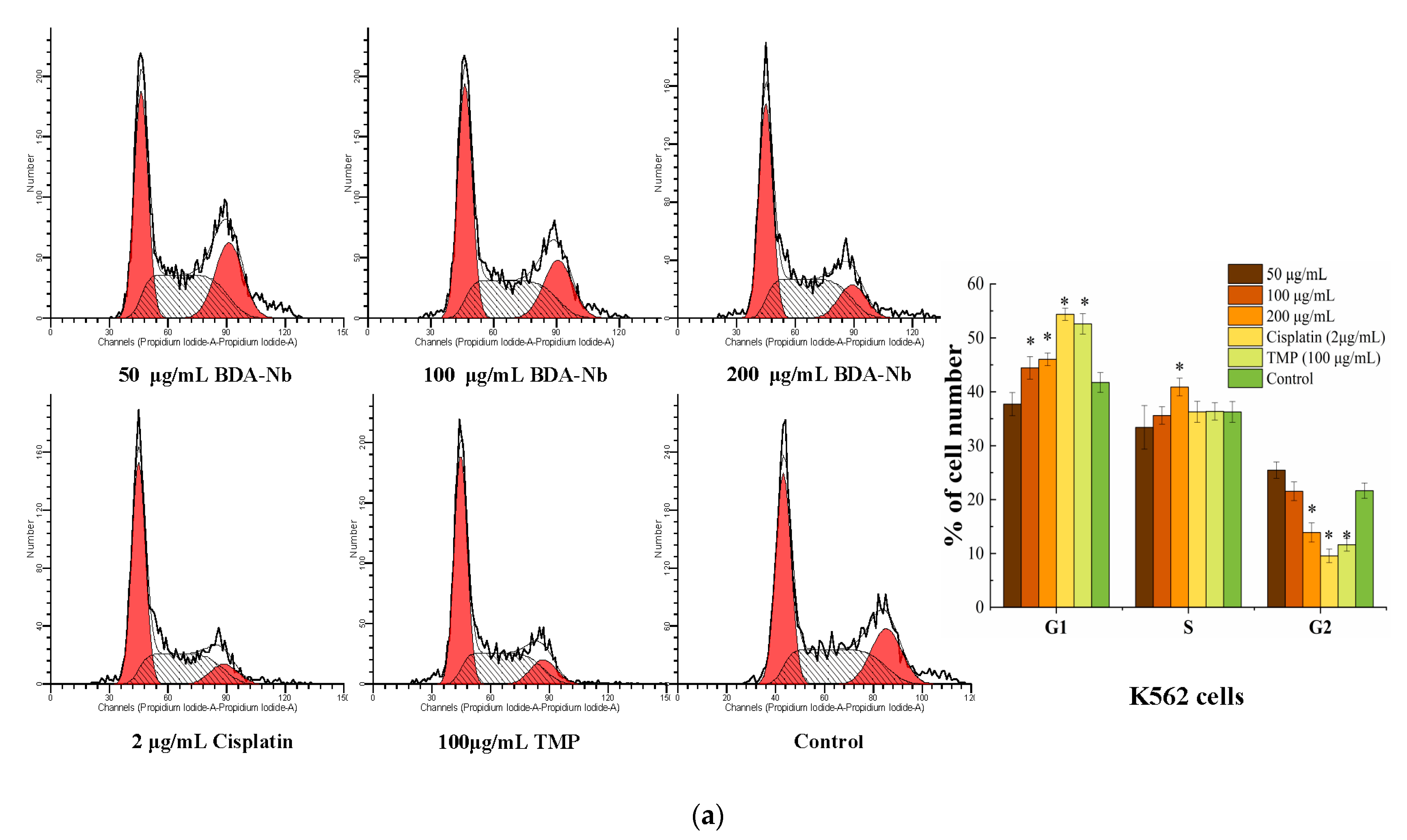
2.7. Flow Cytometry Analysis
2.8. Western Blot Analysis
2.9. Statistical Analysis
3. Results
3.1. Fermentation Improved the Anti-Proliferative Activity of Dehulled Adlay
3.2. Fermentation Increased the Bioactive Component Content of Dehulled Adlay
3.3. BDA-Nb Inhibited the Growth of K562 and A549 Cells
3.4. BDA-Nb Induced Morphological Changes in K562 and A549 Cells
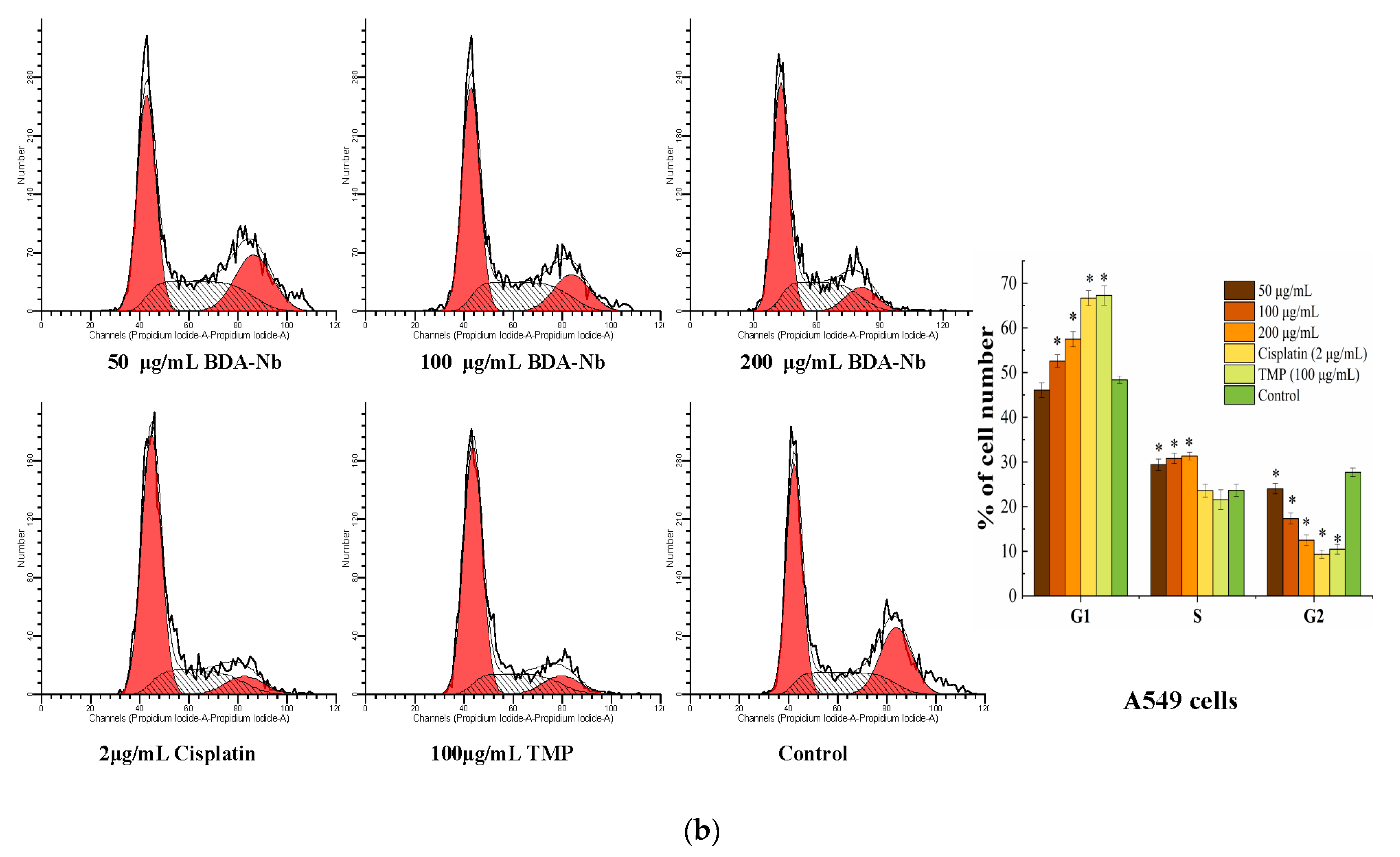
3.5. BDA-Nb Disturbed Cell Cycle Progression in K562 and A549 Cells
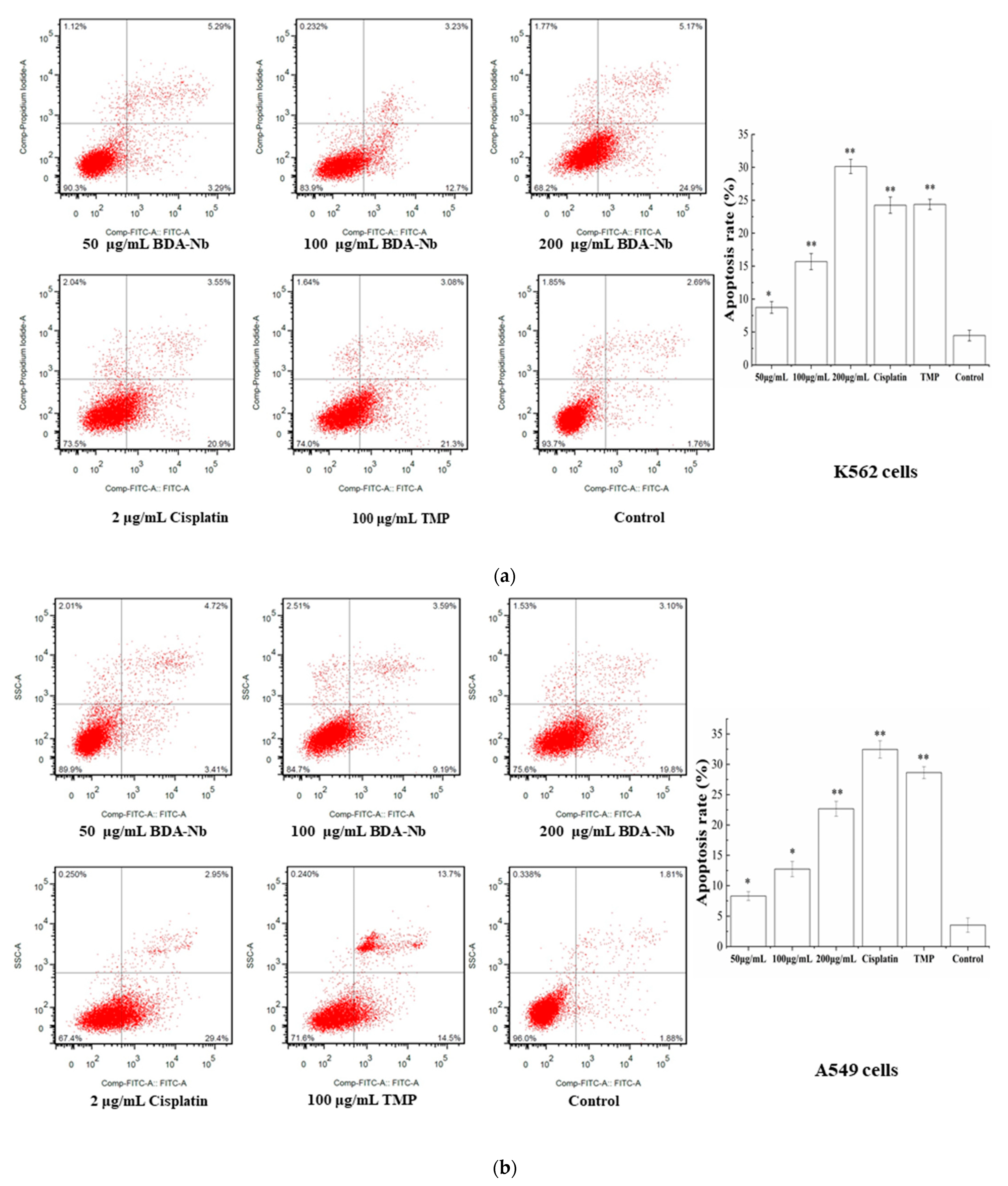
3.6. BDA-Nb Induced Apoptosis in K562 and A549 Cells
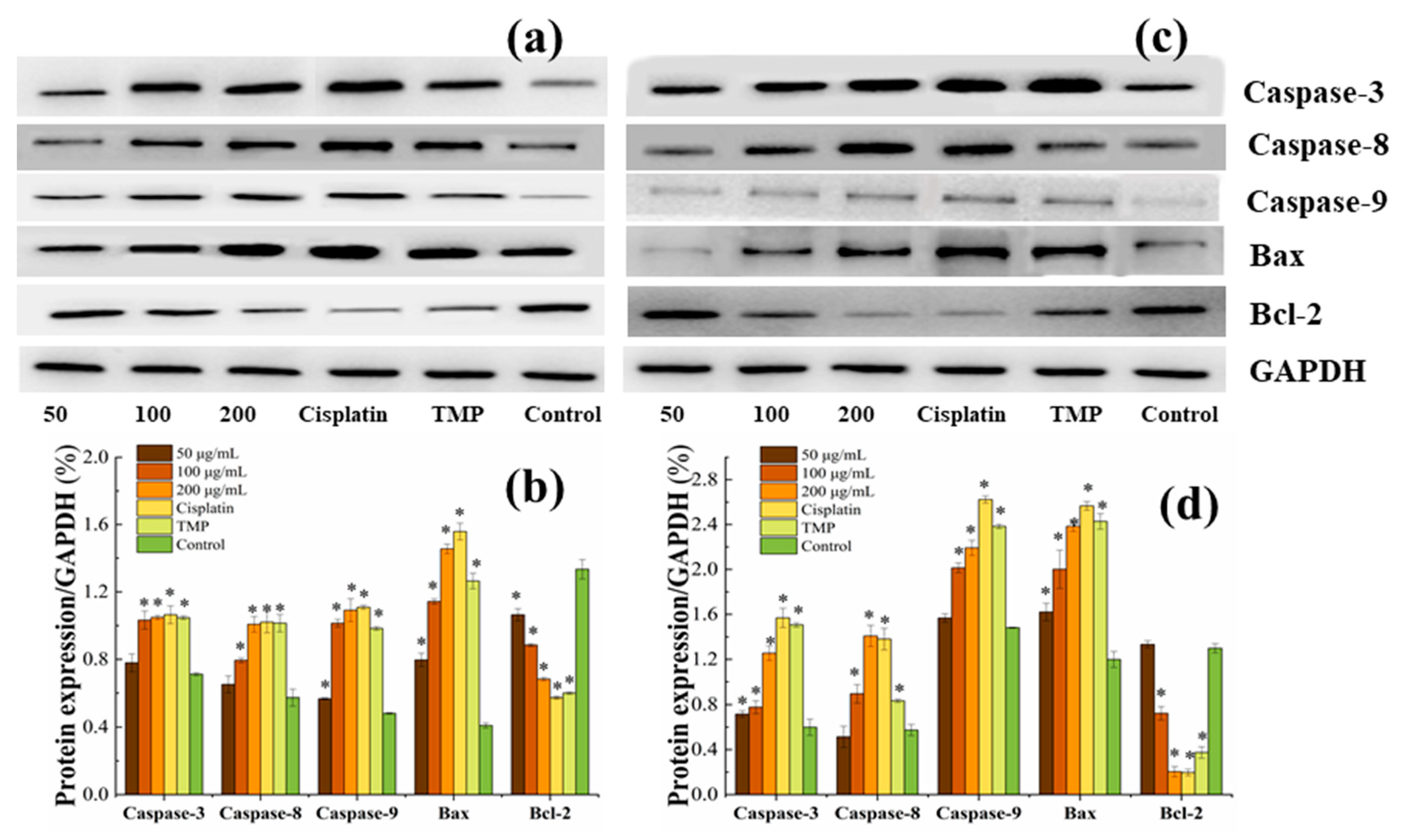
3.7. BDA-Nb Induced Protein Expression in K562 and A549 Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lv, P.H.; Song, Y.Y.; Liu, C.; Yu, L.P.; Shang, Y.L.; Tang, H.; Sun, S.H.; Wang, F.K. Application of Bacillus subtilis as a live vaccine vector: A review. J. Vet. Med. Sci. 2020, 82, 1693–1699. [Google Scholar] [CrossRef]
- Chen, G.; Liu, Y.; Zeng, J.R.; Tian, X.F.; Bei, Q.; Wu, Z.Q. Enhancing three phenolic fractions of oats (Avena sativa L.) and their antioxidant activities by solid-state fermentation with Monascus anka and Bacillus subtilis. J. Cereal Sci. 2020, 93, 102940. [Google Scholar] [CrossRef]
- Li, W.; Wang, T. Effect of solid-state fermentation with Bacillus subtilis lwo on the proteolysis and the antioxidative properties of chickpeas. Int. J. Food Microbiol. 2021, 338, 108988. [Google Scholar] [CrossRef]
- Dai, C.H.; Ma, H.L.; He, R.H.; Huang, L.R.; Zhu, S.; Ding, Q.Z.; Luo, L. Improvement of nutritional value and bioactivity of soybean meal by solid-state fermentation with Bacillus subtilis. LWT-Food Sci. Technol. 2017, 86, 1–7. [Google Scholar] [CrossRef]
- Chen, L.C.; Fan, Z.Y.; Wang, H.Y.; Wen, D.C.; Zhang, S.Y. Effect of polysaccharides from adlay seed on anti-diabetic and gut microbiota. Food Funct. 2019, 1, 4372–4438. [Google Scholar] [CrossRef]
- Zhu, F. Coix: Chemical composition and health effects. Trends Food Sci. Technol. 2017, 61, 160–175. [Google Scholar] [CrossRef]
- Hong, I.H.; Choi, J.Y.; Kim, A.Y.; Lee, E.M.; Kim, J.H.; Park, J.H.; Choi, S.W.; Jeong, K.S. Anti-rheumatoid arthritic effect of fermented Adlay and Achyranthes japonica Nakai on collagen-induced arthritis in mice. Food Agric. Immunol. 2017, 28, 14–26. [Google Scholar] [CrossRef] [Green Version]
- Zeng, H.Y.; Qin, L.K.; Liu, X.Y.; Miao, S. Increases of Lipophilic Antioxidants and Anticancer Activity of Coix Seed Fermented by Monascus purpureus. Foods 2021, 10, 566. [Google Scholar] [CrossRef]
- Wu, S.J.; Fang, J.Y.; Ng, C.C.; Wang, C.Y.; Shyu, Y.T. Anti-inflammatory activity of Lactobacillus-fermented adlay-soymilk in LPS-induced macrophages through suppression of NF-κB pathways. Food Res. Int. 2013, 52, 262–268. [Google Scholar] [CrossRef]
- Wang, C.Y.; Lin, H.T.; Wu, S.C. Influence of dietary supplementation with Bacillus-fermented adlay on lipid metabolism, antioxidant status and intestinal microflora in hamsters. J. Sci. Food Agric. 2011, 91, 2271–2276. [Google Scholar] [CrossRef]
- Wen, A.Y.; Xie, C.Z.; Mazhar, M.; Zhu, Y.; Zeng, H.Y.; Qin, L.K.; Zhu, Y. Comparative evaluation of drying methods on kinetics, biocompounds and antioxidant activity of Bacillus subtilis-fermented dehulled adlay. Dry Technol. 2020, 38, 1505–1515. [Google Scholar] [CrossRef]
- Wen, A.Y.; Qin, L.K.; Zeng, H.Y.; Zhu, Y. Comprehensive evaluation of physicochemical properties and antioxidant activity of B. subtilis-fermented polished adlay subjected to different drying methods. Food Sci. Nutr. 2020, 8, 2124–2133. [Google Scholar] [CrossRef] [Green Version]
- Wen, A.; Xie, C.; Mazhar, M.; Wang, C.; Zeng, H.; Qin, L.; Zhu, Y. Tetramethylpyrazine from adlay (Coix lacryma-jobi) biotransformation by Bacillus subtilis and its quality characteristics. J. Food Sci. Technol. 2020, 57, 4092–4102. [Google Scholar] [CrossRef]
- Hu, H.G.; Zhao, Q.L.; Pang, Z.C.; Xie, J.H.; Lin, L.J.; Yao, Q.S. Optimization extraction, characterization and anticancer activities of polysaccharides from mango pomace. Int. J. Biol. Macromol. 2018, 117, 1314–1325. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, Y.J.; Huang, H.S.; Leu, Y.L.; Peng, K.C.; Chang, C.J.; Chang, M.Y. Anticancer activity of Kalanchoe tubiflora extract against human lung cancer cellsin vitro andin vivo. Environ. Toxicol. 2016, 31, 1663–1673. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.C.; Huang, L.H.; Chiang, W.C.; Hsia, S.M. Hexane fraction of adlay (Coix lachryma-jobi L.) testa ethanolic extract inhibits human uterine sarcoma cancer cells growth and chemosensitizes human uterine sarcoma cells to doxorubicin. Phytomedicine 2018, 47, 69–80. [Google Scholar] [CrossRef]
- Abdelrahman, M.; Mahmoud, H.Y.; El-Sayed, M.; Tanaka, S.; Tran, L.S. Isolation and characterization of Cepa2, a natural alliospiroside A, from shallot (Allium cepa L. Aggregatum group) with anticancer activity. Plant Physiol. Biochem. 2019, 116, 167–173. [Google Scholar] [CrossRef]
- Chen, J.C.; Chen, Q.H.; Guo, Q.; Ruan, S.; Ruan, H.; He, G.Q.; Gu, Q. Simultaneous determination of acetoin and tetramethylpyrazine in traditional vinegars by HPLC method. Food Chem. 2010, 122, 1247–1252. [Google Scholar] [CrossRef]
- Park, N.; Lee, T.K.; Nguyen, T.T.H.; An, E.B.; Kim, N.M.; You, Y.H.; Park, T.S.; Kim, D. The effect of fermented buckwheat on producingl-carnitine- and γ-aminobutyric acid (GABA)-enriched designer eggs. J. Sci. Food Agric. 2017, 97, 2891–2897. [Google Scholar] [CrossRef]
- Xu, L.; Wang, P.; Ali, B.; Yang, N.; Chen, Y.S.; Wu, F.F.; Xu, X.M. Changes of the phenolic compounds and antioxidant activities in germinated adlay seeds. J. Sci. Food Agric. 2017, 97, 4227–4234. [Google Scholar] [CrossRef] [PubMed]
- Rajan, I.; Jayasree, P.R.; Kumar, P.R. Zerumbone induces mitochondria-mediated apoptosis via increased calcium, generation of reactive oxygen species and upregulation of soluble histone H2AX in K562 chronic myelogenous leukemia cells. Tumor Biol. 2015, 36, 8479–8489. [Google Scholar] [CrossRef] [PubMed]
- Tseng, Y.H.; Yang, J.Y.; Chang, H.L.; Lee, Y.L.; Mau, J.L. Antioxidant properties of methanolic extracts from monascal adlay. Food Chem. 2006, 97, 375–381. [Google Scholar] [CrossRef]
- Ayyash, M.; Johnson, S.K.; Liu, S.Q.; Mesmari, N.; Dahmani, S.; Dhaheri, A.S.; Kizhakkayil, J. In vitro investigation of bioactivities of solid-state fermented lupin, quinoa and wheat using Lactobacillus spp. Food Chem. 2019, 275, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Zhang, Y.Y.; Gao, Y.L.; Xu, Q.; Ju, X.R.; Wang, L.F. Identification and anti-tumour activities of phenolic compounds isolated from defatted adlay (Coix lachryma-jobi L. var. ma-yuen Stapf) seed meal. J. Funct. Foods 2016, 26, 394–405. [Google Scholar] [CrossRef]
- Lu, X.Y.; Liu, W.; Wu, J.H.; Li, M.X.; Wang, J.C.; Wu, J.H.; Luo, C. A polysaccharide fraction of adlay seed (Coix lachryma-jobi L.) induces apoptosis in human non-small cell lung cancer A549 cells. Biochem. Biophys. Res. Commun. 2013, 430, 846–851. [Google Scholar] [CrossRef]
- Lee, M.Y.; Lin, H.Y.; Cheng, F.W.; Chiang, W.C.; Kuo, Y.H. Isolation and characterization of new lactam compounds that inhibit lung and colon cancer cells from adlay (Coix lachryma-jobi L. var. ma-yuen Stapf) bran. Food Chem. Toxicol. 2008, 46, 1933–1939. [Google Scholar] [CrossRef]
- Liu, X.H.; Yang, Q.H.; Xi, Y.L.; Yu, K.W.; Wang, W.Z.; Zhao, X.M.; Kuo, X.G. Kanglaite injection combined with chemotherapy versus chemotherapy alone in the treatment of advanced non-small cell lung carcinoma. J. Cancer Res. Ther. 2014, 10, 46–51. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.C.; Wang, Z.G.; Zang, A.M.; Jiao, S.C.; Chen, S.M.; Fu, Y. Tetramethylpyrazine inhibits tumor growth of lung cancer through disrupting angiogenesis via BMP/Smad/Id-1 signaling. Int. J. Oncol. 2016, 48, 2079–2086. [Google Scholar] [CrossRef]
- Jia, X.H.; Xiao, F.F.; Li, J.C. Effect of Apollon siRNA combined with tetramethylpyrazine on proliferation and apoptosis of leukemia K562 cells. Chin. J. Contemp. Pediatrics 2014, 16, 135–140. [Google Scholar]
- Hai, L.; Lin, Z.; Qiang, H.; Fang, X. Inhibition effect of GABA on the growth of cholangiocarcinoma cell QBC 939 and its methanism. Chin. J. Cancer Prev. Treat. 2013, 20, 1585–1588. [Google Scholar]
- Al-Wadei, H.A.; Ullah, M.F.; Al-Wadei, M. GABA (γ-aminobutyric acid), a non-protein amino acid counters the β-adrenergic cascade-activated oncogenic signaling in pancreatic cancer: A review of experimental evidence. Mol. Nutr. Food Res. 2011, 55, 1745–1758. [Google Scholar] [CrossRef]
- Song, L.; Du, A.; Xiong, Y.; Jiang, J.; Zhang, Y.; Tian, Z.; Yan, H. γ-Aminobutyric acid inhibits the proliferation and increases oxaliplatin sensitivity in human colon cancer cells. Tumor Biol. 2016, 37, 14885–14894. [Google Scholar] [CrossRef]
- Li, F.H.; Zhang, X.L.; Li, Y.; Lu, K.K.; Yin, R.; Ming, J. Phenolics extracted from tartary (Fagopyrum tartaricum L. Gaerth) buckwheat bran exhibit antioxidant activity, and an antiproliferative effect on human breast cancer MDA-MB-231 cells through the p38/MAP kinase pathway. Food Funct. 2017, 8, 177–188. [Google Scholar] [CrossRef]
- Chai, T.T.; Quah, Y.; Ooh, K.F.; Ismail, N.I.M.; Ang, Y.V.; Elamparuthi, S.; Yeoh, L.Y.; Ong, H.C.; Wong, F.C. Anti-Proliferative, Antioxidant and Iron-Chelating Properties of the Tropical Highland Fern, Phymatopteris triloba (Houtt) Pichi Serm (Family Polypodiaceae). Trop. J. Pharm. Res. 2013, 12, 747–753. [Google Scholar] [CrossRef] [Green Version]
- Son, E.S.; Kim, S.H.; Kim, Y.O.; Lee, Y.E.; Kyung, S.Y.; Jeong, S.H.; Kim, Y.J.; Park, J.W. Coix lacryma-jobi var. ma-yuen Stapf sprout extract induces cell cycle arrest and apoptosis in human cervical carcinoma cells. BMC Complementary Altern. Med. 2019, 19, 312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu, Z.M.; Wan, H.Y.; Isha, A.; Armania, N. Antioxidant, antimicrobial and cytotoxic potential of condensed tannins from Leucaena leucocephala hybrid-Rendang. Food Sci. Hum. Well. 2016, 5, 65–75. [Google Scholar] [CrossRef] [Green Version]
- Czerwonka, A.; Kaławaj, K.; Sławińska-Brych, A.; Lemieszek, M.K.; Bartnik, M.; Wojtanowski, K.K.; Zdzisińska, B.; Rzeski, W. Anticancer effect of the water extract of a commercial Spirulina (Arthrospira platensis) product on the human lung cancer A549 cell line. Biomed. Pharmacother. 2018, 106, 292–302. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.S.; Lei, T.; Zhang, M. The Reversal Effect and Its Mechanisms of Tetramethylpyrazine on Multidrug Resistance in Human Bladder Cancer. PLoS ONE 2016, 11, e157759. [Google Scholar] [CrossRef]
- Espino, J.; González-Gómez, D.; Moreno, D.; Fernández-León, M.F.; Rodríguez, A.B.; Pariente, J.A.; Delgado-Adámez, J. Tempranillo-derived grape seed extract induces apoptotic cell death and cell growth arrest in human promyelocytic leukemia HL-60 cells. Food Funct. 2013, 4, 1759–1766. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.T.; Korivi, M.; Huang, H.C.; Thiyagarajan, V.; Lin, K.Y.; Huang, P.J.; Liu, J.Y.; Hseu, Y.C.; Yang, H.L. Inhibition of ROS production, autophagy or apoptosis signaling reversed the anticancer properties of Antrodia salmonea in triple-negative breast cancer (MDA-MB-231) cells. Food Chem. Toxicol. 2017, 103, 1–17. [Google Scholar] [CrossRef]
- Chen, Z.G.; Bishop, K.S.; Tanambell, H.; Buchanan, P.; Smith, C.; Quek, S.Y. Characterization of the bioactivities of an ethanol extract and some of its constituents from the New Zealand native mushroom Hericium novae-zealandiae. Food Funct. 2019, 1, 6633–6643. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.T.; Kao, C.J.; Huang, H.Y.; Huang, S.Y.; Chen, C.Y.; Lin, Y.S.; Wen, Z.H.; Wang, H.M.D. Astaxanthin reduces MMP expressions, suppresses cancer cell migrations, and triggers apoptotic caspases of in vitro and in vivo models in melanoma. J. Funct. Foods 2017, 31, 20–31. [Google Scholar] [CrossRef]
- Sasipawan, M.; Natthida, W.; Sahapat, B.; Kanjana, T.; Waraporn, T. Synergistic anticancer effect of the extracts from Polyalthia evecta caused apoptosis in human hepatoma (HepG2) cells. Asian Pac. J. Trop. Biol. 2012, 2, 589–596. [Google Scholar] [CrossRef] [Green Version]
- Dai, J.; Gupte, A.; Gates, L.; Mumper, R.J. A comprehensive study of anthocyanin-containing extracts from selected blackberry cultivars Extraction methods, stability, anticancer properties and mechanisms. Food Chem. Toxicol. 2009, 47, 837–847. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bioactive Components | BDA-Nb | DA-Nb |
|---|---|---|
| Total phenolics (mg GAE/g dry basis) | 36.49 ± 1.37 a | 28.46 ± 1.67 b |
| Total flavonoids (mg RE/g dry basis) | 45.60 ± 2.09 a | 32.16 ± 1.87 b |
| TMP | 4.62 ± 0.25 a | ND |
| GABA | 13.90 ± 1.05 a | 5.29 ± 1.16 b |
| Protocatechuic acid | 1.11 ± 0.09 a | 0.51 ± 0.08 b |
| 2,3,4-Trihydroxybenzoic acid | 2.08 ± 0.07 a | 0.67 ± 0.12 b |
| Chlorogenic acids | 0.45 ± 0.12 a | 0.31 ± 0.07 a |
| p-Hydroxybenzoic acid | 0.32 ± 0.05 a | 0.22 ± 0.08 a |
| Caffeic acid | 0.84 ± 0.08 a | 0.24 ± 0.06 b |
| Trans-Cinnamic acid | 0.95 ± 0.11 a | 0.35 ± 0.07 b |
| Pcoumaric acid | 0.21 ± 0.07 b | 0.35 ± 0.05 a |
| Rutin | 0.56 ± 0.06 a | 0.34 ± 0.09 b |
| Ferulic Acid | 0.63 ± 0.07 a | 0.31 ± 0.13 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wen, A.; Zhu, Y.; Mazhar, M.; Qin, L.; Zeng, H.; Zhu, Y. Enhancement of Anti-Proliferative Activity of the Extracts from Dehulled Adlay by Fermentation with Bacillus subtilis. Foods 2021, 10, 2959. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10122959
Wen A, Zhu Y, Mazhar M, Qin L, Zeng H, Zhu Y. Enhancement of Anti-Proliferative Activity of the Extracts from Dehulled Adlay by Fermentation with Bacillus subtilis. Foods. 2021; 10(12):2959. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10122959
Chicago/Turabian StyleWen, Anyan, Yong Zhu, Muhammad Mazhar, Likang Qin, Haiying Zeng, and Yi Zhu. 2021. "Enhancement of Anti-Proliferative Activity of the Extracts from Dehulled Adlay by Fermentation with Bacillus subtilis" Foods 10, no. 12: 2959. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10122959