
Antibiotic Resistant Enterobacteriaceae in Milk Alternatives
by
 , and
, and
Winnie Mukuna
1, 
Abdullah Ibn Mafiz
1,
Bharat Pokharel
1,
Aniume Tobenna
1 and
Agnes Kilonzo-Nthenge
2,* 1
Department of Agriculture and Environmental Sciences, Tennessee State University, 3500 John A. Merritt Boulevard, Nashville, TN 37209, USA
2
Department of Human Sciences, Tennessee State University, 3500 John A. Merritt Boulevard, Nashville, TN 37209, USA
*
Author to whom correspondence should be addressed.
Foods 2021, 10(12), 3070; https://0-doi-org.brum.beds.ac.uk/10.3390/foods10123070
Submission received: 29 September 2021
/
Revised: 2 December 2021
/
Accepted: 8 December 2021
/
Published: 10 December 2021
(This article belongs to the Special Issue Foodborne Pathogens and Food Safety)
Abstract
:The consumption of non-dairy milk is on the rise due to health benefits. Although there is increasing inclination towards milk alternatives (MA), there is limited data on antibiotic resistant bacteria in these substitutes. The aim of this study was to investigate antimicrobial resistance of bacteria isolated from MA. A total of 138 extracts from almonds (n = 63), cashew nuts (n = 36), and soybeans (n = 39) were analyzed for Enterobacteriaceae. The identification of the bacteria was based on biochemical and PCR methods. Antibiotic sensitivity was determined by using the Kirby-Bauer disk diffusion technique. Overall, 31% (43 of 138) of extracts were positive for Enterobacteriaceae. Ten bacterial species were identified, of which Enterobacter cloacae (42.7%) and Enterobacter cancerogenus (35.4%) were the most predominant species (p < 0.05). Antibiotic resistance was exhibited to vancomycin (88.3%), novobiocin (83.8%), erythromycin (81.1%), which was significantly higher (p < 0.05) than in tetracycline (59.5%), cefpodoxime (30.6%), and nalidixic acid (6.3%). There was no resistance displayed to kanamycin and imipenem. ERY-NOV-VAN-TET and ERY-NOV-CEP-VAN-TET were the most common resistant patterns displayed by Enterobacter cloacae. The findings of this study suggest that MAs, though considered healthy, may be a reservoir of multidrug resistant opportunist pathogens.
1. Introduction
Milk is considered a superior source of micro- and macro-nutrients compared to milk alternatives (MA) [1]. However, its association with increased risks of cardiovascular diseases, diabetes, cancer, and as a principal vehicle for transmission of foodborne pathogens continues to make it unfavorable. Generally, cow milk is frequently consumed and dominates global milk production [2], accounting for 85% of the world’s production, followed by buffalo milk at 11%, goat (2.3%), sheep (1.4%), and camel (0.2%) [3]. However, due to the current changes in lifestyles towards a healthier diet, there has been an increasing trend in the consumption of MA [4]. The U.S. market for MA is increasing and has reached an annual sales volume of $1.8 billion [4]. The increased market growth is attributed to the consumers’ preference for vegan diets, increasing instances of lactose intolerance, and a growing demand for fortified non-dairy food and beverages [5,6,7]. Generally, consumers’ perception is that MA are healthier than milk [8]. Milk alternatives are becoming increasingly popular; however, they are characterized by low protein content, and poor bioavailability of minerals and vitamins [9]. With the increasing demand for these MA, different plants with varying functional attributes are being explored as bases for primary materials for processing [10]. Soymilk, which originated from Asia [11], is the most globally consumed MA while almond milk is the most prevalently used, solely based on sales volume [12]. Other available MA are sourced from cashew nuts, hemp, coconut, rice, etc. [10]. The majority of non-dairy consumers purchase their MA from grocery stores, though a sector of the population make these milk substitutes at home by using raw nuts or seeds. Hence, home-made milk alternative might potentially be contaminated if food safety is not practiced during preparation and storage. Although MA are an intensifying trend, the usage of the term “milk” to mean plant-based substitutes to milk is debatable and is protected by legislation in several countries [8].
There are abundant MA in the market, such as almond, cashew, soy, rice, hazelnut, and oat milk [13,14]. Nuts and seeds, the primary raw materials for milk alternatives, may come into direct contact with soil and be contaminated with pathogenic bacteria at pre- or post-harvest period [15]. It is usually thought that, due to less moisture content, nuts, seeds, and grains are less susceptible to microbial contamination [16]. Regrettably, this attribute does not exempt nuts and seeds from contamination with foodborne pathogens. For instance, Salmonella serovar has been detected in almonds, pecans, and peanuts [15,17,18], E. coli O157:H7 in walnuts [19], and Listeria spp. in peanuts, almonds, cashews, and hazelnuts [20]. Moreover, Pseudomonas spp., Clostridium spp., and Klebsiella spp. have been detected in other nuts [21]. These bacteria and others that are prevalent in raw nuts and seeds belong to the family Enterobacteriaceae, the most prevalent human opportunistic pathogens [22].
The increasing frequency of antimicrobial resistant bacteria is a global threat [23]. Accordingly, it is important to study the presence of antimicrobial resistant Enterobacteriaceae in MA, especially with the continuous increase trend in consumption. Antimicrobial resistant bacteria cause illnesses that have high morbidity and mortality [24], one of the greatest health challenges in the 21st century [25]. Around 99,000 individuals die every year in the USA owing to drug-resistant infections [26]. Antimicrobial-resistant Enterobacteriaceae in milk and milk products has been reported in numerous studies [27]. Just like milk, milk substitutes can also be potential vehicles for transmission of antimicrobial resistant foodborne pathogens to consumers. Antimicrobial resistant pathogens originating from raw nuts or seeds might be transferred to MA during preparation at processing facilities or at home. To the best of our knowledge, there has been limited enquiry of the possible occurrence of antimicrobial resistant bacteria in raw MA. Consequently, this study aims to investigate the presence of opportunist Enterobacteriaceae in MA and their resistance to antibiotics used both in human and animal medicine.
2. Materials and Methods
2.1. Sample Collection and Preparation
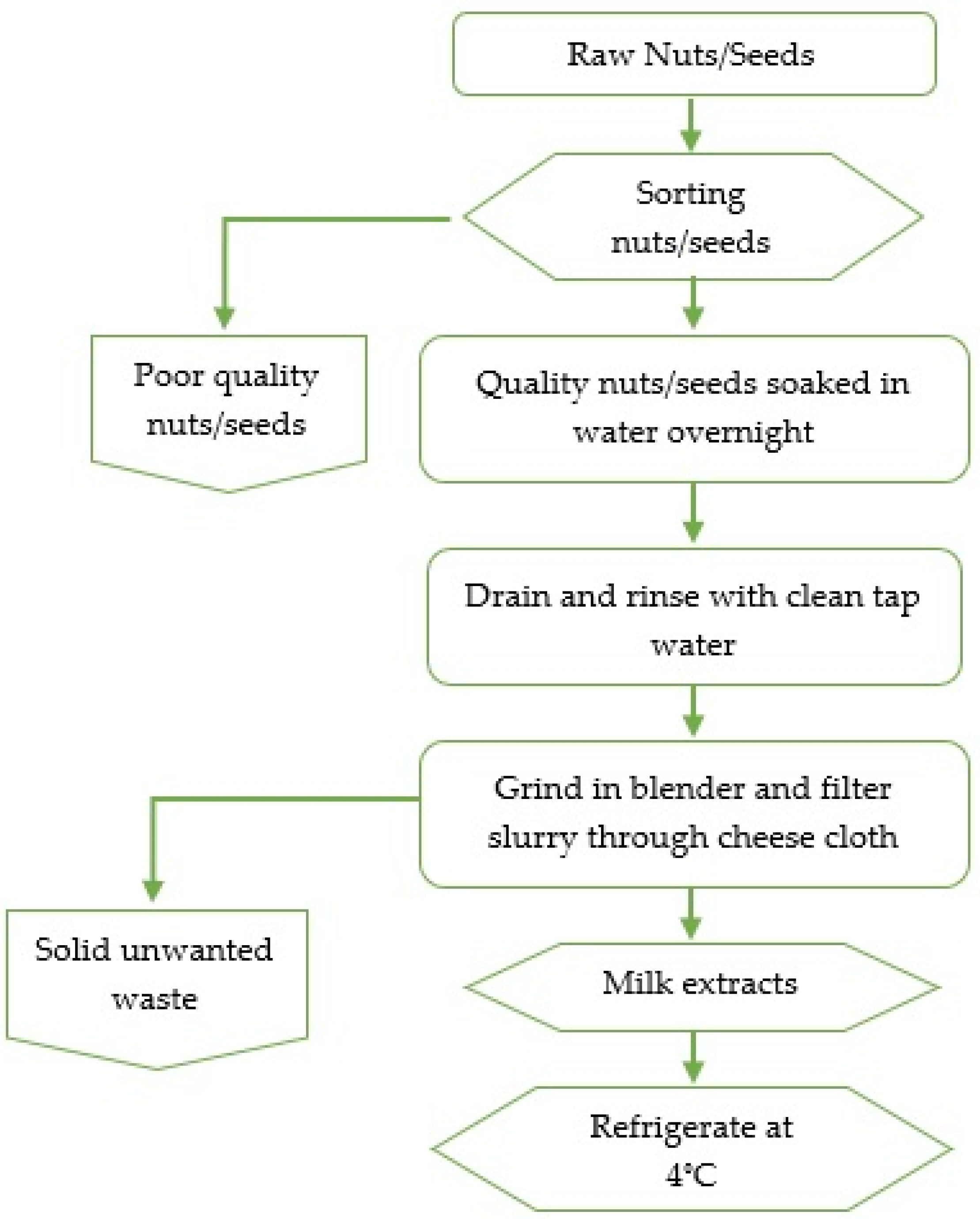
Raw nuts (almond, cashew) and soybeans were randomly purchased from 3 local stores in Davidson County, Tennessee, depending on availability. The preparation of almonds, cashew nuts, and soybean extracts involved schematic steps as displayed in the flowchart (Figure 1). Briefly, in duplicates, 5 g of each sample (almonds, cashews, and soybeans) were sorted from unwanted materials (damaged, split seeds or nuts), followed by soaking separately overnight in 45 mL sterile distilled water at room temperature. Next, in duplicates, each sample was disintegrated in a laboratory blender (Waring Division, Dynamics Corporation, New Hartford, CT, USA) for 3 min at high speed. The resulting slurry was filtered through a cheesecloth (Farberware, Fairfield, CA, USA) to attain milk extracts which were then placed in sterile capped containers. A total of 138 extracts (almond nuts = 63, cashew nuts = 36, and soybeans = 39) were analyzed for Enterobacteriaceae and AMR by using biochemical and molecular tests.
Enrichment of Milk and Bacterial Identification
One ml of nuts and seed extracts was enriched in 9 mL Enterobacteriaceae enrichment (EE) broth Mossel enrichment (BD, Sparks, MD) and incubated at 37 °C for 24 h. From each enriched sample, 1µL was streaked onto violet red bile agar (Oxoid, Basingstoke, and Hants, UK) and incubated for 18–24 h at 37 °C. Red to dark purple colonies surrounded by red-purple halos were identified as presumptive Enterobacteriacea. Enterobacteriacea isolates and further characterized by using oxidase and API 20E (bioMerieux, Hazelwood, MO, USA) tests. Three colonies per plate were selected for API biochemical testing. Due to the role played by Klebsiella pneumoniae and ronobacter sakazakii as opportunist pathogens in clinical settings, isolates above the 90% confidence interval were stored at −80 °C for further testing.
2.2. DNA Extraction and Confirmation of Klebsiella and Cronobacter Sakazakii
Biochemically identified K. pneumoniae and C. sakazakii isolates from almond and cashew extracts, respectively, were further confirmed by PCR. DNA was extracted from overnight cultures (≤2 × 109 cells) using the PureLink Genomic DNA Mini Kit (Invitrogen, Carlsbad, CA, USA). DNA concentrations and integrity were determined using a NanoDrop 2000 (Thermo Scientific, Pittsburgh, PA, USA) and agarose gel electrophoresis, respectively. Oligonucleotide primer pairs were synthesized (Operon Technologies, Huntsville, AL, USA) and used to amplify genes of interest. The sequences of the primer pair used for targeting C. sakazakii target gene ompA (469 bp) was 3′-GGATTTAACCGTGAACTTTTCC-5′ and 5′-CGCCAGCGATGTTAGAAGA-3′ [28,29]. Each reaction mixture (20 μL) contained 4 μL DNA template, 1 μL of each primer (×2), 10 μL master mix, 2 μL RNase free water and, 2 μL coral load (supplied with the kit). C. muytjensii (ATCC 51329) was used as a positive control for the detection and identification methods. Reaction conditions for PCR were initial denaturation at 95 °C for 5 min, 30 cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 1 min, extension at 72 °C for 10 min, and final extension at 72 °C for 10 min.
A Multiplex PCR plus kit (Qiagen, Hillden, Germany) was used to amplify K. pneumoniae and Klebsiella spp primers in a single reaction [30]. Primer pair 5′-CAA CGG TGT GGT TAC TGA CG-3′ and 5′-TCT ACG AAG TGG CCG TTT TC-3′ targeted gene rpoB (108 bp) in K. pneumoniae isolates as described by [30], and 5′- CGC GTA CTA TAC GCC ATG AAC GTA-3′ and 5′-ACC GTT GAT CAC TTC GGT CAG G-3′ targeted gene gyrA (441bp) in Klebsiella spp. [31]. Each 50 μL reaction mixture contained 25 μL of master mix, 5 μL of 10 × primer mix (2.5 μM each primer), 100 ng DNA template, 5 μL Q-solution, 5 μL Coral Load dye, and 10 μL RNase free water. Reaction conditions for PCR were: initial denaturation at 95 °C for 5 min, 35 cycles of denaturation at 95 °C for 30 s, annealing at 60 °C for 90 s, extension at 72 °C for 90 s, and final extension at 68 °C for 10 min. K. pneumoniae (ATCC 49131) and Salmonella typhimurium (ATCC 13311) were used as positive and negative control, respectively. A nexus gradient Thermal Cycler (Eppendorf, Hauppauge, New York) was used for all amplifications. PCR products were electrophoresed in agarose gel stained with 0.1 µg/mL of ethidium bromide (Sigma-Aldrich, Madrid, Spain) and photographed under UV light.
2.3. Antibiotic Resistant Profiles
For all identified Enterobacteriaceae isolates (n = 110), the characterization of the strain resistance/susceptibility profiles was carried out as recommended by the Clinical and Laboratory Standards Institute guidelines [32]. The antimicrobial susceptibility test was conducted on isolates that were identified at ≤90 confidence interval by API 20E system. Antimicrobial disks (n = 8), with strength in parentheses were: vancomycin (VAN; 30 μg), novobiocin (NOVO; 30 μg), erythromycin (ERY; 15 μg), tetracycline (TET; 30 μg), cefpodoxime (CEF; 10 μg), kanamycin (KAN; 10 μg), nalidixic acid (NAL; 30 μg), and imipenem (IPM; 30 μg). The results were interpreted as susceptible, intermediate, and resistant based on the Clinical and Laboratory Standards Institute recommendations [32]. Escherichia coli ATCC 25922 and Staphylococcus aureus ATCC 25923 were used as control strains. Reference standard bacterial strains were verified simultaneously with controls.
2.4. Statistical Analysis
The bacterial data were expressed as percentages and analyzed using Microsoft Excel 2016 (Microsoft Corp., Redmond, WA, USA). Chi-square tests were used to measure the significance of difference in the incidence of Enterobacteriaceae and antimicrobial resistance. Data were analyzed using SPSS v. 25.0 (IBM SPSS, Chicago, IL, USA). p values of less than 0.05 were considered statistically significant.
3. Results and Discussion
3.1. Enterobacteriaceae in Nuts and Seeds Extract
Overall, 31% (43 of 138) of extracts were positive for Enterobacteriaceae. Specifically, Enterobacteriaceae isolation rates were 33.3% (21/63), 30.5% (11/36), and 28.2% (11/39) of almond, cashew, and soybean extracts, respectively (data not shown). Enterobacteriaceae offers valuable information on the hygienic conditions during food preparation or post-process contamination [33]. Overall, 79.7% (110 out of 138) Enterobacteriaceae isolates were identified from almond, cashew, and soybean milk extracts (Table 1).
In 2020, almond milk (MILKLAB and Blue Dimond Almond Breeze Chocolate Almond Milk) recalls were reported in Australia due to contamination with Pseudomonas [34]. These recalls support our results that MA can be contaminated with pathogenic bacteria. Our findings also suggest that MA may be contaminated with harmful microorganism. Pathogens such as Salmonella serovar., Listeria spp., E. coli spp., Campylobacter spp., Brucella spp. or Shigella spp. [35,36,37] have been associated with milk.
According to our findings, a total of 10 different commensal and pathogenic genera of Enterobacteriaceae were identified with the most common strain being Enterobacter cloacae at 42.7% (47 of 110), which was not significantly different (p > 0.05) from Enterobacter cancerogenus at 35.4% (39 of 110). E. cloacae is a commensal microorganism found in human and animal guts and widely found in food, soil, and water [38]. Although E. cloacae is not a common foodborne pathogen, its presence in MA is a concern as it is a widely known nosocomial pathogen and the third most prevalent acquired bacteria causing illness in hospital after E. coli and K. pneumoniae [39].
Our results indicate that clinically significant C. sakazakii accounted for 2.7% (3 out of 110) of the identified isolates. C. sakazakii was only detected in cashew extracts and was confirmed through amplification of the OmpA gene (469 bp) (Figure 2).
Earlier findings showed that OmpA is a determinant that causes C. sakazakii invasion of brain microvascular endothelial cells in vitro, and possibly contributes to pathogenesis of neonatal meningitis [40]. Cronobacter spp. is an emerging pathogen and a major concern, especially to hypersensitive clusters of the population including children and the elderly [41,42]. C. sakazakii is also considered as an evolving opportunistic pathogen [43] that has been detected in milk, and powdered infant milk among other sources [44]. Although there is no data on the incidence of C. sakazakii in MA, nuts and seeds are important raw materials in these substitutes which might be contaminated with pathogenic bacteria at any point during production, harvest, storage, and transportation [45]. At production and harvesting stages, pathogenic bacteria might transfer from the soils onto the nuts/seeds when they are in contact with the ground. One possible scenario is during almond harvesting as was the case of Salmonella in almonds grown in California [15].
K. pneumoniae spp. ozaenae (4.5%) and K. pneumoniae spp. pneumoniae (2.7%) in the current study were also isolated from almond and cashew extracts, respectively. As these two bacteria are emerging pathogens of concern, they were confirmed by multiplex PCR through amplification of rpoB (108 bp) and gyrA (441 bp) genes for K. pneumoniae and Klebsiella spp., respectively (Figure 3).
During harvesting, almond trees are shaken to release the nuts and might stay on the ground for up to 2 weeks before collection [46]. Through this period, bacteria in the soil may be transferred to the hulls which might infiltrate to the kernel as has been demonstrated in Salmonella on almonds and pecans [18]. Klebsiella spp. have recently become significant pathogens in nosocomial infections [47] such as urinary tract infection, bacteremia, pneumonia, sepsis, and meningitis [48]. With the increased trend in adoption of MA and with some consumers making their nut and seed extracts at home, they may also be at risk of nosocomial infections originating from contaminated and unpasteurized extracts. To avoid potential infections from Klebsiella spp., consumers should be encouraged to adhere to food safety practices or drink pasteurized MA.
Other bacteria in the Enterobacteriaceae family were also identified in the current study (Table 1). Our findings present E. vulneris (1.8%) in soybean extracts. It is possible that the soybeans used in this study were contaminated with E. vulneris through soil or water that was used at preharvest or post-harvest. Jain et al. [49] hypothesized that an infant infected with gastroenteritis may have been infected by contaminated formula or water that was used to reconstitute the formula. Escherichia vulneris has previously been recovered from water, soil, human beings, and animals [50]. Rahnella aquatilis (0.9%) was another Enterobacteriaceae isolated from soybean milk extract in our study. Rahnella aquatilis is considered a primary and opportunistic pathogen that has been associated with diarrhea and endocarditis [51]. Milk alternatives may be extracted from nuts and seeds that may directly touch the soil during pre- or post-harvest [15]. Hence, restricted precautions must be taken during planting and harvesting of nuts and seeds and processing of MA. Additionally, nuts and seeds should be stored in dry facilities that are protected from rain and ground water, insects and pests, and that have optimal temperatures that avert microbial growth [45].
3.2. Antimicrobial Drug Resistance in Enterobacteriaceae
Detailed presentation of antimicrobial resistant Enterobacteriaceae species from MA extracts is shown in Table 2. In the present study, Enterobacteriaceae resistance in isolated bacteria was higher (p < 0.05) in vancomycin (90.0%), novobiocin (83.7%), and erythromycin (80.9%) than in tetracycline (60.0%), cefpodoxime (31.8%), and nalidixic acid (6.4%). The majority of Enterobacteriaceae in our study are opportunistic pathogens that cause nosocomial infections; hence their antimicrobial resistance might lead to impediments in treating infected individuals [52]. Our findings agree with a previous study that documented C. sakazakii resistance to both erythromycin and tetracycline [53]. Occurrence of antibiotic resistant C. sakazakii in nut and seed extracts is a concern because antibiotic therapy is a chosen path to avert Cronobacter infection in humans [54]. Resistance to erythromycin, tetracycline, vancomycin, and novobiocin was also exhibited by K. pneumoniae isolates in nut and seed extracts in our study, hence a concern, since K. pneumoniae is a significant multidrug-resistant (MDR) pathogen that causes hospital infections leading to high morbidity and death [55], one of the most severe challenges in clinical practice.
According to Zhou et al. [56], Klebsiella-resistant strains have increased more quickly than those of any other bacteria in the past decade. The consumption of both MA and milk may result in foodborne illnesses if not controlled [57]. According to our study, antibiotic resistant E. vulneris was detected in MA. Our data is supported by previous studies [58] which displayed multiple antibiotic resistant E. coli strains in milk.
The absence of resistance among all Enterobacteriaceae strains to kanamycin was also noted in the current study. Additionally, Enterobacteriaceae strains in this study did not display resistance to imipenem which agrees with previous findings [59]. Although no imipenem resistance was indicated in our findings, carbapenems have been used to treat numerous Enterobacteriaceae infections, hence there has been a rapid development in their resistance to the same. The rapid spread of carbapenem resistant Enterobacteriaceae (CRE) in the community is a national epidemiologic concern, since Enterobacteriaceae are common causes of nosocomial and community infections.
A total of seven multidrug-resistance patterns were observed in Enterobacteriaceae in this study (Table 3). Out of 110 Enterobacteriaceae isolates, 87 (79.1%) from nuts and seed extracts were multidrug-resistant. According to Nguyen et al. [60], an MDR isolate displays resistance to three or more classes of antibiotic. Overall, the most common resistance pattern (ERY-NOV-VAN-TET) in our study was exhibited in by Citrobacter youngae (1), E. cancerogenus (7), E. cloacae (18), E. vulneris (1) Pantoea spp. 3 (3), and Rahnella aquatilis (1) (number of isolates in parenthesis). Forty-four (44) E. cloacae and 28 E. cancerogenus isolates recovered from nuts and seed extracts were multidrug-resistant (MDR). ERY-NOV-VAN-TET was the most significant (p < 0.05) multidrug resistance pattern among E. cloacae isolates. E. cloacae and E. cancerogenus presented a common resistance pattern: ERY-NOV-CEP-NAL-VAN-TET, which was resistant to six out of eight antibiotics.
4. Conclusions
Processed MA and milk food safety can be improved by implementation of high sanitary standards that reduce risk of contamination. Milk contamination with micro-organisms can occur before harvest, during milking or postharvest, and in storage. Similarly, MA may be contaminated by use of pathogen tinted nuts or seeds before harvest, during collection, and processing, and in storage. With the increased trend in adoption of MA, consumers may also be at risk of infection with AMR bacteria from ingestion of unpasteurized MA. Therefore, it is imperative that consumers should be educated on safe milk handling practices. Although many consumers are aware of foodborne illnesses, they have limited knowledge of food storage, time, and temperature abuse that may increase bacterial growth.
Although MA are considered healthy, our data suggest that they are reservoirs of antibiotic resistant Enterobacteriaceae. Consumers should be aware of the impending risks of ingesting unpasteurized milk substitutes in their homes, which can harbor AMR bacteria that can pose serious health risks.
Author Contributions
Conceptualization, W.M. and A.K.-N.; methodology, W.M., A.K.-N. and B.P.; validation, W.M.; formal analysis, W.M. and A.K.-N.; investigation, W.M. and A.K.-N.; resources, A.K.-N.; writing—original draft preparation, W.M.; writing—review and editing, W.M., A.K.-N., A.I.M. and A.T.; visualization, W.M. and A.K.-N.; supervision, A.K.-N.; project administration, A.K.-N.; and funding acquisition, A.K.-N. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by United States Department of Agriculture/National Institute of Food and Agriculture. Grant No. Accession No: 1014615; Project: TENX-1813-FS.
Acknowledgments
We are grateful for the PCR technical suggestions by Boniface Kimathi and Collins Khwatenge.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Rice, B.H.; Quann, E.E.; Miller, G.D. Meeting and exceeding dairy recommendations: Effects of dairy consumption on nutrient intakes and risk of chronic disease. Nutr. Rev. 2013, 71, 209–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Claeys, W.; Verraes, C.; Cardoen, S.; De Block, J.; Huyghebaert, A.; Raes, K.; Dewettinck, K.; Herman, L. Consumption of raw or heated milk from different species: An evaluation of the nutritional and potential health benefits. Food Control 2014, 42, 188–201. [Google Scholar] [CrossRef]
- Gerosa, S.; Skoet, J. Milk Availability: Trends in Production and Demand and Medium-Term Outlook; FAO, Agricultural Development Economics Division: Rome, Italy, 2012. [Google Scholar]
- Mintel. Added Value in Dairy Drinks, Milk and Cream. Available online: https://0-reports-mintel-com.brum.beds.ac.uk/display/792665/ (accessed on 2 September 2021).
- Mäkinen, O.E.; Wanhalinna, V.; Zannini, E.; Arendt, E.K. Foods for special dietary needs: Non-dairy plant-based milk substitutes and fermented dairy-type products. Crit. Rev. Food Sci. Nutr. 2016, 56, 339–349. [Google Scholar] [CrossRef]
- Stall, S.; Adams, G. Can almond milk be called milk? J. Ren. Nutr. 2017, 27, e15–e17. [Google Scholar] [CrossRef] [Green Version]
- Fiocchi, A.; Schünemann, H.J.; Brozek, J.; Restani, P.; Beyer, K.; Troncone, R.; Martelli, A.; Terracciano, L.; Bahna, S.L.; Rancé, F. Diagnosis and rationale for action against cow’s milk allergy (DRACMA): A summary report. J. Allergy Clin. Immunol. 2010, 126, 1119–1128.e1112. [Google Scholar] [CrossRef]
- McCarthy, K.; Parker, M.; Ameerally, A.; Drake, S.; Drake, M. Drivers of choice for fluid milk versus plant-based alternatives: What are consumer perceptions of fluid milk? J. Dairy Sci. 2017, 100, 6125–6138. [Google Scholar] [CrossRef] [PubMed]
- Aydar, E.F.; Tutuncu, S.; Ozcelik, B. Plant-based milk substitutes: Bioactive compounds, conventional and novel processes, bioavailability studies, and health effects. J. Funct. Foods 2020, 70, 103975. [Google Scholar]
- Sethi, S. Bread, Wine, Chocolate: The Slow Loss of Foods We Love; HarperCollins: New York, NY, USA, 2015. [Google Scholar]
- Afroz, M.; Anjum, W.; Islam, N.; Kobir, A.; Hossain, K.; Sayed, A. Preparation of soymilk using different methods. J. Food Nutr. Sci. 2016, 4, 11–17. [Google Scholar] [CrossRef] [Green Version]
- Watson, E. Almond Breeze Almond Milk only Contains 2% Almonds, Claims False Advertising Lawsuit. 2015. Available online: https://www.foodnavigator-usa.com/Article/2015/07/22/ (accessed on 20 October 2021).
- Vanga, S.K.; Raghavan, V. How well do plant based alternatives fare nutritionally compared to cow’s milk? J. Food Sci. Technol. 2018, 55, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Ziarati, P.; Shirkhan, F.; Mostafidi, M.; Zahedi, M.T. An overview of the heavy metal contamination in milk and dairy products. Acta Sci. Pharm. Sci. 2018, 2, 1–14. [Google Scholar]
- Danyluk, M.D.; Jones, T.M.; Abd, S.J.; Schlitt-Dittrich, F.; Jacobs, M.; Harris, L.J. Prevalence and amounts of Salmonella found on raw California almonds. J. Food Prot. 2007, 70, 820–827. [Google Scholar] [CrossRef]
- Willis, C.; Little, C.L.; Sagoo, S.; de Pinna, E.; Threlfall, J. Assessment of the microbiological safety of edible dried seeds from retail premises in the United Kingdom with a focus on Salmonella spp. Food Microbiol. 2009, 26, 847–852. [Google Scholar] [CrossRef]
- Calhoun, S.; Post, L.; Warren, B.; Thompson, S.; Bontempo, A.R. Prevalence and concentration of Salmonella on raw shelled peanuts in the United States. J. Food Prot. 2013, 76, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Brar, P.K.; Strawn, L.K.; Danyluk, M.D. Prevalence, level, and types of Salmonella isolated from North American in-shell pecans over four harvest years. J. Food Prot. 2016, 79, 352–360. [Google Scholar] [CrossRef] [PubMed]
- Davidson, G.R.; Frelka, J.C.; Yang, M.; Jones, T.M.; Harris, L.J. Prevalence of Escherichia coli O157: H7 and Salmonella on inshell California walnuts. J. Food Prot. 2015, 78, 1547–1553. [Google Scholar] [CrossRef] [PubMed]
- Eglezos, S. The bacteriological quality of retail-level peanut, almond, cashew, hazelnut, Brazil, and mixed nut kernels produced in two Australian nut-processing facilities over a period of 3 years. Foodborne Pathog. Dis. 2010, 7, 863–866. [Google Scholar] [CrossRef] [PubMed]
- Al-Moghazy, M.; Boveri, S.; Pulvirenti, A. Microbiological safety in pistachios and pistachio containing products. Food Control 2014, 36, 88–93. [Google Scholar] [CrossRef]
- Liu, S.; Kilonzo-Nthenge, A. Prevalence of multidrug-resistant bacteria from US-grown and imported fresh produce retailed in chain supermarkets and ethnic stores of Davidson County, Tennessee. J. Food Prot. 2017, 80, 506–514. [Google Scholar] [CrossRef]
- Jee, Y.; Carlson, J.; Rafai, E.; Musonda, K.; Huong, T.T.G.; Daza, P.; Sattayawuthipong, W.; Yoon, T. Antimicrobial resistance: A threat to global health. Lancet Infect. Dis. 2018, 18, 939–940. [Google Scholar] [CrossRef]
- Ventola, C.L. The antibiotic resistance crisis: Part 1: Causes and threats. Pharm. Ther. 2015, 40, 277. [Google Scholar]
- Centers for Disease Control and Prevention, Office of Infectious Disease Antibiotic Resistance Threats in the United States. 2013. Available online: http://www.cdc.gov/drugresistance/threat-report-2013 (accessed on 5 September 2021).
- Golkar, Z.; Bagasra, O.; Pace, D.G. Bacteriophage therapy: A potential solution for the antibiotic resistance crisis. J. Infect. Dev. Ctries 2014, 8, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Ababu, A.; Endashaw, D.; Fesseha, H. Isolation and Antimicrobial Susceptibility Profile of Escherichia coli O157: H7 from Raw Milk of Dairy Cattle in Holeta District, Central Ethiopia. Int. J. Microbiol. 2020, 2020, 1–8. [Google Scholar] [CrossRef]
- Zhou, Y.; Wu, Q.; Xu, X.; Yang, X.; Ye, Y.; Zhang, J. Development of an immobilization and detection method of Enterobacter sakazakii from powdered infant formula. Food Microbiol. 2008, 25, 648–652. [Google Scholar] [CrossRef]
- Kilonzo-Nthenge, A.; Rotich, E.; Godwin, S.; Nahashon, S.; Chen, F. Prevalence and antimicrobial resistance of Cronobacter sakazakii isolated from domestic kitchens in middle Tennessee, United States. J. Food Prot. 2012, 75, 1512–1517. [Google Scholar] [CrossRef] [PubMed]
- Chander, Y.; Ramakrishnan, M.; Jindal, N.; Hanson, K.; Goyal, S.M. Differentiation of Klebsiella pneumoniae and K. oxytoca by multiplex polymerase chain reaction. Int. J. Appl. Res. Vet. Med. 2011, 9, 138. [Google Scholar]
- Brisse, S.; Verhoef, J. Phylogenetic diversity of Klebsiella pneumoniae and Klebsiella oxytoca clinical isolates revealed by randomly amplified polymorphic DNA, gyrA and parC genes sequencing and automated ribotyping. Int. J. Syst. Evol. Microbiol. 2001, 51, 915–924. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute (CLSI). Performance Standards for Antimicrobial Susceptibility Testing, 31st ed.; CLSI Supplement M100S; CLSI: Wayne, PA, USA, 2018. [Google Scholar]
- Heperkan, D.; Dalkilic-Kaya, G.; Juneja, V.K. Cronobacter sakazakii in baby foods and baby food ingredients of dairy origin and microbiological profile of positive samples. LWT 2017, 75, 402–407. [Google Scholar] [CrossRef]
- Food Authority. MILKLAB and Blue Diamond Almond Milk. 2020. Available online: https://www.foodauthority.nsw.gov.au/news/recalls/milklab-and-blue-diamond-almond-milk. (accessed on 20 October 2021).
- Langer, A.J.; Ayers, T.; Grass, J.; Lynch, M.; Angulo, F.J.; Mahon, B.E. Nonpasteurized dairy products, disease outbreaks, and state laws—United States, 1993–2006. Emerg. Infect. Dis. 2012, 18, 385. [Google Scholar] [CrossRef]
- Allocati, N.; Masulli, M.; Alexeyev, M.F.; Di Ilio, C. Escherichia coli in Europe: An overview. Int. J. Environ. Res. Public Health 2013, 10, 6235–6254. [Google Scholar] [CrossRef]
- Vandera, E.; Lianou, A.; Kakouri, A.; Feng, J.; Koukkou, A.-I.; Samelis, J. Enhanced control of Listeria monocytogenes by Enterococcus faecium KE82, a multiple enterocin–producing strain, in different milk environments. J. Food Prot. 2017, 80, 74–85. [Google Scholar] [CrossRef]
- Davin-Regli, A. Enterobacter aerogenes and Enterobacter cloacae; versatile bacterial pathogens confronting antibiotic treatment. Front. Microbiol. 2015, 6, 392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jarlier, V.; INVS. Surveillance of Multidrug Resistant Bacteria in French Healthcare Facilities BMR-Raisin Network Données. Saint-Maurice: Institut de Veille Sanitaire. 2014. Available online: http://www.invs.sante.fr (accessed on 3 September 2021).
- Mohan Nair, M.K.; Venkitanarayanan, K.; Silbart, L.K.; Kim, K.S. Outer membrane protein A (OmpA) of Cronobacter sakazakii binds fibronectin and contributes to invasion of human brain microvascular endothelial cells. Foodborne Pathog. Dis. 2009, 6, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organization (FAO); World Health Organization (WHO). Enterobacter sa-kazakii (Cronobacter spp.) in Powdered Follow-Up Formulae. In Microbiological Risk Assessment; Series No. 15; FAO: Rome, Italy, 2008. [Google Scholar]
- Hunter, C.J.; Bean, J. Cronobacter: An emerging opportunistic pathogen associated with neonatal meningitis, sepsis and necrotizing enterocolitis. J. Perinatol. 2013, 33, 581–585. [Google Scholar] [CrossRef] [Green Version]
- Yan, Q.; Condell, O.; Power, K.; Butler, F.; Tall, B.; Fanning, S. Cronobacter species (formerly known as Enterobacter sakazakii) in powdered infant formula: A review of our current understanding of the biology of this bacterium. J. Appl. Microbiol. 2012, 113, 1–15. [Google Scholar] [CrossRef]
- Mullane, N.; Iversen, C.; Healy, B.; Walsh, C.; Whyte, P.; Wall, P.; Quinn, T.; Fanning, S. Enterobacter sakazakii an emerging bacterial pathogen with implications for infant health. Minerva Pediatrica 2007, 59, 137–148. [Google Scholar]
- Brar, P.K.; Danyluk, M.D. Nuts and grains: Microbiology and preharvest contamination risks. Preharvest Food Saf. 2018, 105–121. [Google Scholar] [CrossRef]
- Kader, A.A.; Thompson, J.F. Postharvest Handling Systems: Tree Nuts; University California Publication: Davis, CA, USA, 2002; pp. 399–406. [Google Scholar]
- Nordmann, P.; Cuzon, G.; Naas, T. The real threat of Klebsiella pneumoniae carbapenemase-producing bacteria. Lancet Infect. Dis. 2009, 9, 228–236. [Google Scholar] [CrossRef]
- Zarkotou, O.; Pournaras, S.; Tselioti, P.; Dragoumanos, V.; Pitiriga, V.; Ranellou, K.; Prekates, A.; Themeli-Digalaki, K.; Tsakris, A. Predictors of mortality in patients with bloodstream infections caused by KPC-producing Klebsiella pneumoniae and impact of appropriate antimicrobial treatment. Clin. Microbiol. Infect. 2011, 17, 1798–1803. [Google Scholar] [CrossRef] [Green Version]
- Jain, S.; Nagarjuna, D.; Gaind, R.; Chopra, S.; Debata, P.; Dawar, R.; Sardana, R.; Yadav, M. Escherichia vulneris: An unusual cause of complicated diarrhoea and sepsis in an infant. A case report and review of literature. Microbe Infect. 2016, 13, 83–86. [Google Scholar] [CrossRef] [Green Version]
- Le Querler, L.; Donnio, P.; Poisson, M.; Rouzet-Gras, S.; Avril, J. Isolation of Escherichia vulneris from drinking water. Ann. Biol. Clin. 1997, 55, 33–35. [Google Scholar]
- Matsukura, H.; Katayama, K.; Kitano, N.; Kobayashi, K.; Kanegane, C.; Higuchi, A.; Kyotani, S. Infective endocarditis caused by an unusual gram-negative rod, Rahnella aquatilis. Pediatrica Cardiol. 1996, 17, 108–111. [Google Scholar] [CrossRef]
- Aghapour, Z.; Gholizadeh, P.; Ganbarov, K.; Bialvaei, A.Z.; Mahmood, S.S.; Tanomand, A.; Yousefi, M.; Asgharzadeh, M.; Yousefi, B.; Kafil, H.S. Molecular mechanisms related to colistin resistance in Enterobacteriaceae. Infect. Drug Resist. 2019, 12, 965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hochel, I.; Růžičková, H.; Krásný, L.; Demnerová, K. Occurrence of Cronobacter spp. in retail foods. J. Appl. Microbiol. 2012, 112, 1257–1265. [Google Scholar] [CrossRef]
- Depardieu, F.; Podglajen, I.; Leclercq, R.; Collatz, E.; Courvalin, P. Modes and modulations of antibiotic resistance gene expression. Clin. Microbiol. Rev. 2007, 20, 79–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navon-Venezia, S.; Kondratyeva, K.; Carattoli, A. Klebsiella pneumoniae: A major worldwide source and shuttle for antibiotic resistance. FEMS Microbiol. Rev. 2017, 41, 252–275. [Google Scholar] [CrossRef]
- Zhou, X.-Y.; Ye, X.-G.; He, L.-T.; Zhang, S.-R.; Wang, R.-L.; Zhou, J.; He, Z.-S. In vitro characterization and inhibition of the interaction between ciprofloxacin and berberine against multidrug-resistant Klebsiella pneumoniae. J. Antibiot. 2016, 69, 741–746. [Google Scholar] [CrossRef] [Green Version]
- Melini, F.; Melini, V.; Luziatelli, F.; Ruzzi, M. Raw and heat-treated milk: From public health risks to nutritional quality. Beverages 2017, 3, 54. [Google Scholar] [CrossRef] [Green Version]
- Sudda, M.M.; Mtenga, A.B.; Kusiluka, L.J.; Kassim, N. Prevalence and Antibiotic Susceptibility of Escherichia coli and Salmonella spp. isolated from milk of zero grazed cows in Arusha City. Afr. J. Microbiol. Res. 2016, 10, 1944–1951. [Google Scholar]
- Ombarak, R.A.; Hinenoya, A.; Elbagory, A.-R.M.; Yamasaki, S. Prevalence and molecular characterization of antimicrobial resistance in Escherichia coli isolated from raw milk and raw milk cheese in Egypt. J. Food Prot. 2018, 81, 226–232. [Google Scholar] [CrossRef]
- Nguyen, D.P.; Nguyen, T.A.D.; Le, T.H.; Tran, N.M.D.; Ngo, T.P.; Dang, V.C.; Kawai, T.; Kanki, M.; Kawahara, R.; Jinnai, M. Dissemination of extended-spectrum β-lactamase-and AmpC β-lactamase-producing Escherichia coli within the food distribution system of Ho Chi Minh City, Vietnam. Biomed. Res. Int. 2016, 2016, 8182096. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
A schematic diagram for extracting milk from almonds, cashews, and soybeans.

Figure 2.
Represents PCR amplification of the ompA gene in Cronobacter sakazakii, Lane 1: 1 kb ladder; lane 2: negative control; lane 3: positive control; lane 4–5: C. sakazakii isolates.
Figure 2.
Represents PCR amplification of the ompA gene in Cronobacter sakazakii, Lane 1: 1 kb ladder; lane 2: negative control; lane 3: positive control; lane 4–5: C. sakazakii isolates.

Figure 3.
Multiplex PCR amplification of gyrA and rpoB genes in K. pneumoniae and Klebsiella spp. Lane 1: 1 kb ladder; lane 2 & 11: positive control; lane 3–10: K. pneumoniae and Klebsiella spp.; lane 12: negative control.
Figure 3.
Multiplex PCR amplification of gyrA and rpoB genes in K. pneumoniae and Klebsiella spp. Lane 1: 1 kb ladder; lane 2 & 11: positive control; lane 3–10: K. pneumoniae and Klebsiella spp.; lane 12: negative control.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Presence (%) of Enterobacteriaceae in Nut and Seed Extracts.
| Bacterial Species | Total Isolates (N = 110) | No. (%) of ENT Isolates in Extracts | ||
|---|---|---|---|---|
| Almond Milk (n = 56) | Cashew Milk (n = 28) | Soy Milk (n = 26) | ||
| Enterobacter cancerogenus | 39 (35.4) a | 22 (39.28) a | 5 (17.9) b | 12 (46.2) a |
| Enterobacter cloacae | 47 (42.7) a | 21 (37.5) a | 15 (17.9) a | 11 (42.3) a |
| Klebsiella pneumoniae spp. ozaenae | 5 (4.5) bc | 5 (8.9) b | 0 (0) c | 0 (0) c |
| Pantoea spp. 3 | 8 (7.3) b | 8 (14.2) b | 0 (0) c | 0 (0) c |
| Chryseomonas luteola | 1 (0.9) c | 0 (0) c | 1 (3.6) b,c | 0 (0) bc |
| Citrobacter youngae | 1 (0.9) c | 0 (0) c | 1 (3.6) b,c | 0 (0) bc |
| Cronobacter sakazakii | 3 (2.7) b,c | 0 (0) c | 3 (10.7) b | 0 (0) bc |
| Klebsiella pneumoniae spp. pneumoniae | 3 (2.7) b,c | 0 (0) c | 3 (10.7) b | 0 (0) bc |
| Escherichia Vulneris | 2 (1.8) b,c | 0 (0) c | 0 (0) c | 2 (7.7) c |
| Rahnella aquatilis | 1 (0.9) c | 0 (0) c | 0 (0) c | 1 (3.8) c |
N: Total number of Enterobacteriaceae isolates. n: Total number of Enterobacteriaceae isolates from various extracts. a–c Mean percentages in the same column followed by different letters are significantly different (p < 0.05).
Table 2.
Resistant Antibiotics Profile and Enterobacteriaceae Nut and Seed Extracts.
| Antibiotics (µg) | No. (%) of Enterobacteriaceae Resistant to Antimicrobial Agents | No. (%) of Total Resistant (* R) Isolates | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Almond milk (n = 56) | Cashew Milk (n = 28) | Soy Milk (n = 26) | ||||||||
| R | I | S | R | I | S | R | I | S | ||
| Erythromycin (15) | 48 (85.7) b | 8 (14.3) b | 0 (0) e | 28 (100) a | 0 (0)c | 0 (0) d | 13 (50) b | 2 (7.7) bc | 11 (42.3) b | 89 (80.9) a |
| Novobiocin (30) | 51 (91.1) b | 1 (1.8) d | 4 (7.1) d | 27 (96.4) a | 13.6) a | 0 (0) d | 14 (53.9) b | 12 (46.1) a | 0 (0) d | 92 (83.7) a |
| Cefpodoxime (10) | 13 (23.2) d | 24 (42.9) a | 19 (33.9) c | 5 (17.9) c | 8 (28.6) b | 15 (53.6) b | 17 (65.4) ab | 5 (19.2) b | 4 (15.4) c | 35 (31.8) c |
| NalidixicAcid (30) | 5 (8.9) e | 2 (3.6) cd | 49 (87.5) b | 2 (7.1) cd | 0 (0) c | 26 (92.9) a | 0 (0) c | 2 (7.7) b | 24 (92.3) a | 7 (6.4) d |
| Imipenem (30) | 0 (0) f | 0 (0) d | 56 (100) a | 0 (0) d | 0 (0) c | 28 (100) a | 0 (0) c | 0 (0) c | 26 (100) a | 0 (0) e |
| Kanamycin (10) | 0 (0) f | 6 (10.7) c | 50 (89.3) b | 0 (0) c | 2 (7.1) c | 26 (92.9) a | 0 (0) c | 2 (7.7) bc | 24 (92.3) a | 0 (0) e |
| Vancomycin (30) | 56 (100) a | 0 (0) d | 0 (0) e | 28 (100) a | 0 (0) c | 0 (0) d | 15 (57.7) ab | 1 (3.8) c | 10 (38.5) bc | 99 (90.0) a |
| Tetracycline (30) | 31 (55.4) c | 17 (30.4) a | 8 (14.3) d | 14 (50) b | 8 (28.6) a | 6 (21.4) c | 21 (80.8) a | 5 (19.2) b | 0 (0) d | 66 (60.0) b |
R = Resistant, I = Intermediate, S = Susceptible (CLSI, 2018). * R = Total number of resistant isolates from all extracts (µg). n: Total number of Enterobacteriacea isolates from various extracts. a–f Mean percentages in the same column followed by different letters are significantly different (p < 0.05).
Table 3.
Antibiotic Resistance Patterns of Enterobacteriaceae in Nuts and Seed Extracts.
| Bacterial Species A | No of Isolates | Resistance Profile B |
|---|---|---|
| Chryseomonas luteola | 1 d | ERY-NOV-CEP-VAN-TET |
| Citrobacter youngae | 1 d | ERY-NOV-VAN-TET |
| Enterobacter Cancerogenus | 8 b,c | CEP-TET |
| 1 d | CEP-VAN-TET | |
| 1 d | ERY-CEP-TET | |
| 4 c,d | ERY-NOV-CEP-NAL-VAN-TET | |
| 3 c,d | ERY-NOV-CEP-VAN-TET | |
| 10 b | ERY-NOV-VAN | |
| 7 b,c | ERY-NOV-VAN-TET | |
| 3 c,d | ERY-VAN | |
| 2 c,d | NOV-CEP-TET | |
| Enterobacter cloacae | 3 c,d | ERY-NOV-CEP-NAL-VAN-TET |
| 12 a,b | ERY-NOV-CEP-VAN-TET | |
| 11 a,b | ERY-NOV-VAN | |
| 18 a | ERY-NOV-VAN-TET | |
| 2 c,d | NOV-VAN | |
| 1 d | VAN | |
| Cronobacter sakazakii | 2 c,d | ERY-NOV-VAN |
| 1 d | ERY-VAN | |
| Escherichia vulneris | 1 d | ERY-NOV-VAN-TET |
| 1 d | VAN | |
| Klebsiella pneumoniae spp. ozaenae | 5 c,d | NOV-VAN |
| Klebsiella pneumoniae spp. pneumoniae | 3 c,d | ERY-NOV-VAN |
| Pantoea spp. 3 | 3 c,d | ERY-NOV-VAN |
| 3 c,d | ERY-NOV-VAN-TET | |
| 1 d | ERY-VAN | |
| 1 d | VAN | |
| Rahnella aquatilis | 1 d | ERY-NOV-VAN-TET |
A Bacterial species isolated from milk extracts (MA). B Antibiotic resistance patterns against eight antibiotics: vancomycin (VAN), novobiocin (NOVO), erythromycin (ERY), tetracycline (TET), cefpodoxime (CEF), kanamycin (KAN), nalidixic acid (NAL), and imipenem (IPM). a–d Number of isolates in the same column followed by different letters are significantly different (p < 0.05).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Mukuna, W.; Mafiz, A.I.; Pokharel, B.; Tobenna, A.; Kilonzo-Nthenge, A. Antibiotic Resistant Enterobacteriaceae in Milk Alternatives. Foods 2021, 10, 3070. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10123070
AMA Style
Mukuna W, Mafiz AI, Pokharel B, Tobenna A, Kilonzo-Nthenge A. Antibiotic Resistant Enterobacteriaceae in Milk Alternatives. Foods. 2021; 10(12):3070. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10123070
Chicago/Turabian StyleMukuna, Winnie, Abdullah Ibn Mafiz, Bharat Pokharel, Aniume Tobenna, and Agnes Kilonzo-Nthenge. 2021. "Antibiotic Resistant Enterobacteriaceae in Milk Alternatives" Foods 10, no. 12: 3070. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10123070
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.