
The Influence of Red Cabbage Extract Nanoencapsulated with Brassica Plasma Membrane Vesicles on the Gut Microbiome of Obese Volunteers

 , ,
, ,  ,
,  ,
,  and
and
Abstract
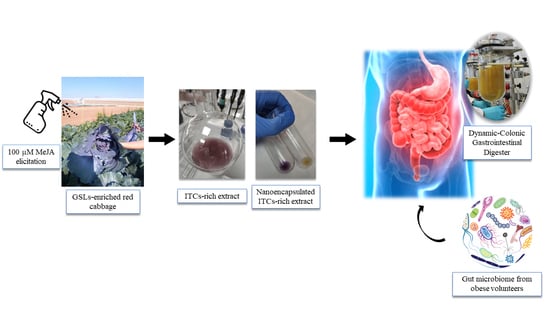
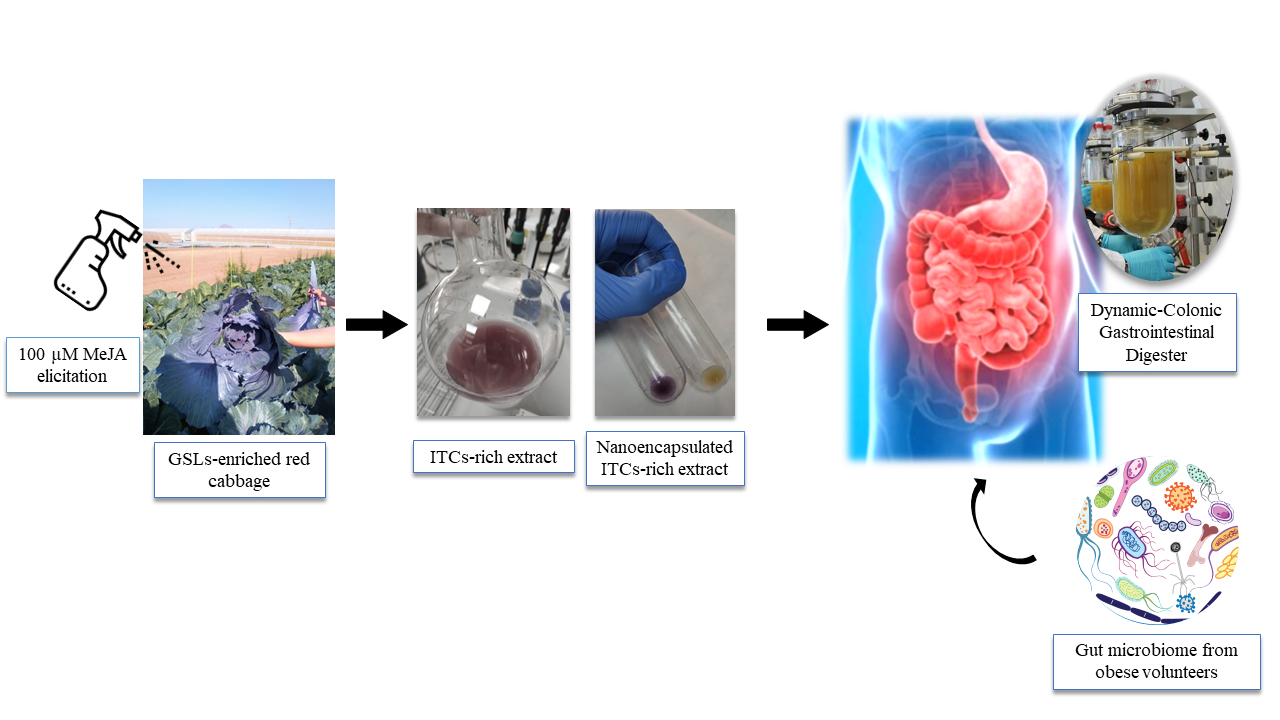
:
1. Introduction
2. Materials and Methods
2.1. Plant Material and Treatments
2.2. Extraction of Intact GSLs
2.3. Elaboration of the ITC-Rich Ingredient Prototype
2.4. Microsomal Fraction Extraction
2.5. Plasma Membrane Isolation
2.6. Dynamic Gastrointestinal and Colonic Fermentation Model
2.7. Microbiota Composition and 16S rRNA Analysis
2.8. Bioinformatic Analysis
2.9. Glucosinolates and Isothiocyanates Quantitative Determination
2.10. Data Analysis
3. Results
3.1. Field Elicitation of Red Cabbage
3.2. Characterization of Extracts
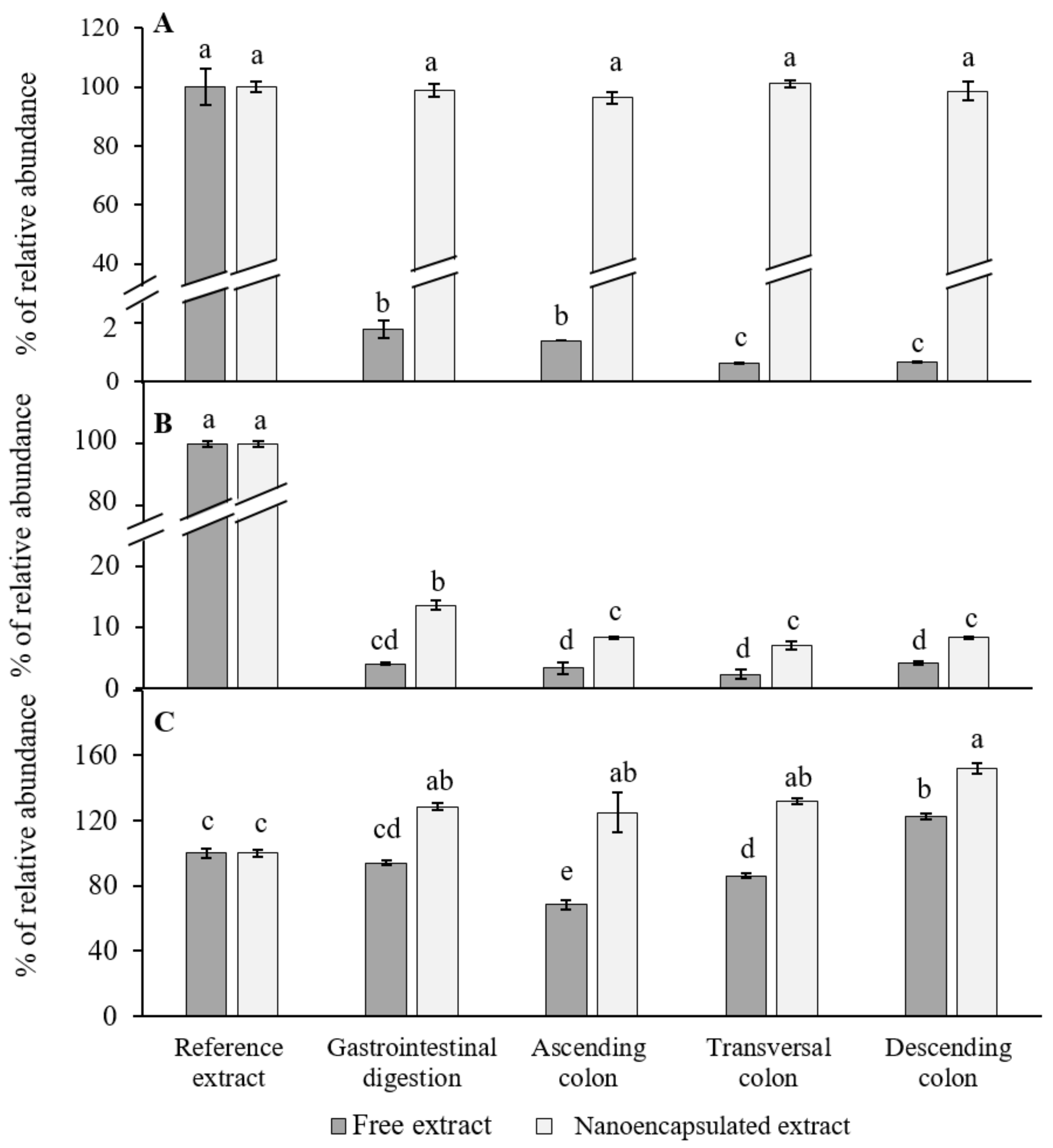
3.3. Dynamic Gastrointestinal and Colonic Fermentation Model
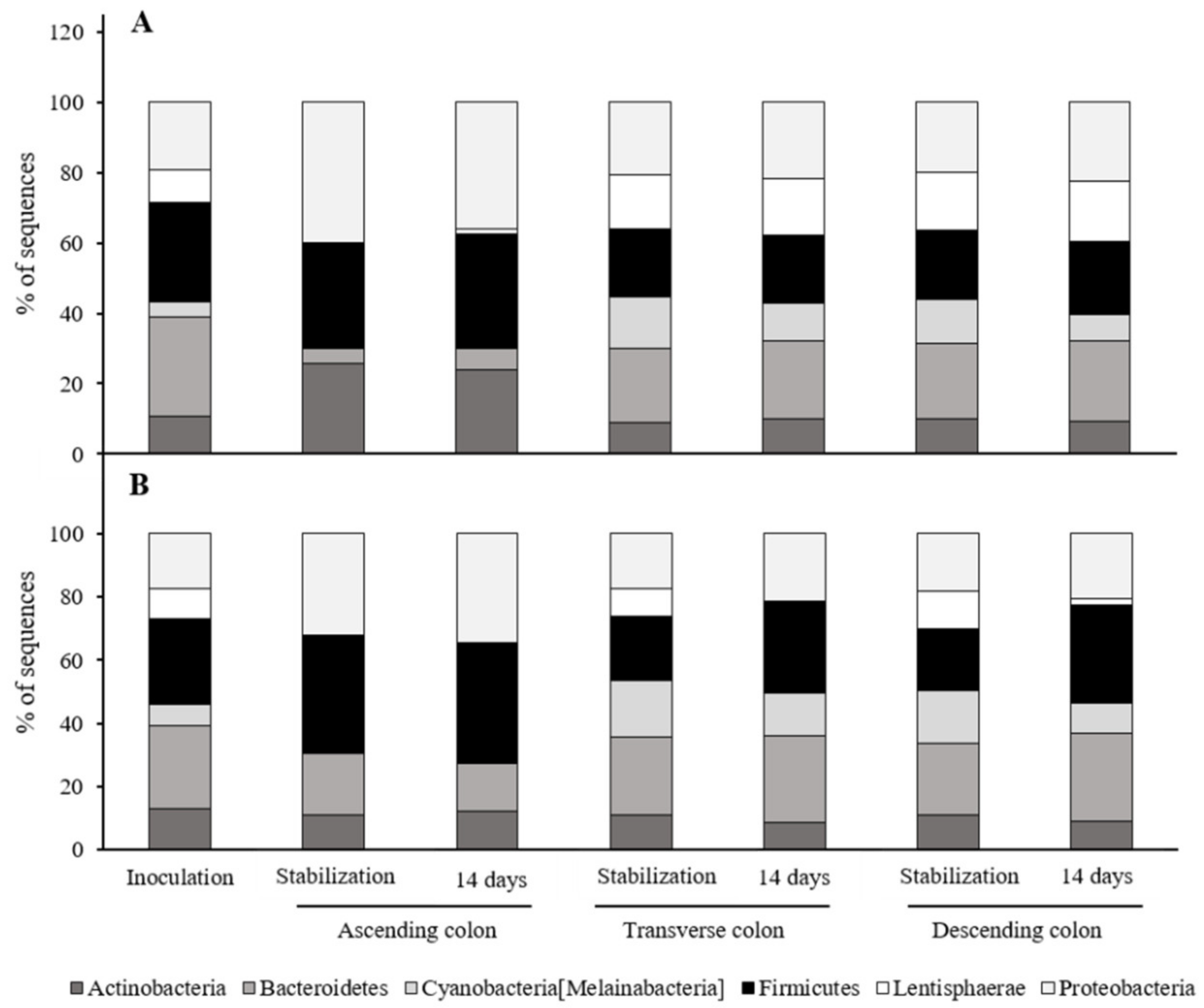
3.4. Effects of the Red Cabbage Extracts on the Microbiome
3.5. Butyric Acid Production by the Microbiota
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martins, T.; Colaço, B.; Venâncio, C.; Pires, M.J.; Oliveira, P.A.; Rosa, E.; Antunes, L.M. Potential Effects of Sulforaphane to Fight Obesity. J. Sci. Food Agric. 2018, 98, 2837–2844. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.; Poudyal, H.; Panchal, S.K. Functional Foods as Potential Therapeutic Options for Metabolic Syndrome. Obes. Rev. 2015, 16, 914–941. [Google Scholar] [CrossRef] [PubMed]
- Houghton, C.A. The Emergence of Sulforaphane as a Clinically-Relevant Nutraceutical in the Prevention and Treatment of Disease. In Current Topics in Medicine and Medical Research Vol. 2; Book Publisher International: London, UK, 2020; p. 123. [Google Scholar] [CrossRef]
- Radziejewska-Kubzdela, E.; Biegańska-Marecik, R. A Comparison of the Composition and Antioxidant Capacity of Novel Beverages with an Addition of Red Cabbage in the Frozen, Purée and Freeze-Dried Forms. LWT Food Sci. Technol. 2015, 62, 821–829. [Google Scholar] [CrossRef]
- Blažević, I.; Montaut, S.; Burčul, F.; Olsen, C.E.; Burow, M.; Rollin, P.; Agerbirk, N. Glucosinolate Structural Diversity, Identification, Chemical Synthesis and Metabolism in Plants. Phytochemistry 2020, 169, e112100. [Google Scholar] [CrossRef] [PubMed]
- Bhat, R.; Vyas, D. Myrosinase: Insights on Structural, Catalytic, Regulatory, and Environmental Interactions. Crit. Rev. Biotechnol. 2019, 39, 508–523. [Google Scholar] [CrossRef] [PubMed]
- Soundararajan, P.; Kim, J.S. Anti-Carcinogenic Glucosinolates in Cruciferous Vegetables and Their Antagonistic Effects on Prevention of Cancers. Molecules 2018, 23, 2983. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaczmarek, J.L.; Liu, X.; Charron, C.S.; Novotny, J.A.; Jeffery, E.H.; Seifried, H.E.; Ross, S.A.; Miller, M.J.; Swanson, K.S.; Holscher, H.D. Broccoli Consumption Affects the Human Gastrointestinal Microbiota. J. Nutr. Biochem. 2019, 63, 27–34. [Google Scholar] [CrossRef]
- Kellingray, L.; Tapp, H.S.; Saha, S.; Doleman, J.F.; Narbad, A.; Mithen, R.F. Consumption of a Diet Rich in Brassica Vegetables Is Associated with a Reduced Abundance of Sulphate-Reducing Bacteria: A Randomised Crossover Study. Mol. Nutr. Food Res. 2017, 61, e1600992. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.K.; Kang, Y.F.; Zhao, X.Y.; Liu, Y.P.; Zhang, X.W.; Zhang, S.J. Effects of Elicitation on Bioactive Compounds and Biological Activities of Sprouts. J. Funct. Foods 2019, 53, 136–145. [Google Scholar] [CrossRef]
- Moreno, D.A.; López-Berenguer, C.; Martínez-Ballesta, M.C.; Carvajal, M.; García-Viguera, C. Basis for the New Challenges of Growing Broccoli for Health in Hydroponics. J. Sci. Food Agric. 2008, 88, 904–910. [Google Scholar] [CrossRef]
- Baenas, N.; García-Viguera, C.; Moreno, D.A. Elicitation: A Tool for Enriching the Bioactive Composition of Foods. Molecules 2014, 19, 13541–13563. [Google Scholar] [CrossRef] [Green Version]
- Baenas, N.; Villaño, D.; García-Viguera, C.; Moreno, D.A. Optimizing Elicitation and Seed Priming to Enrich Broccoli and Radish Sprouts in Glucosinolates. Food Chem. 2016, 204, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Ibañez, P.; Moreno, D.A.; Nuñez-Gomez, V.; Agudelo, A.; Carvajal, M. Use of Elicitation in the Cultivation of Bimi® for Food and Ingredients. J. Sci. Food Agric. 2020, 100, 2099–2109. [Google Scholar] [CrossRef] [PubMed]
- Baenas, N.; García-Viguera, C.; Moreno, D.A. Biotic Elicitors Effectively Increase the Glucosinolates Content in Brassicaceae Sprouts. J. Agric. Food Chem. 2014, 62, 1881–1889. [Google Scholar] [CrossRef] [PubMed]
- Hassini, I.; Rios, J.J.; Garcia-Ibañez, P.; Baenas, N.; Carvajal, M.; Moreno, D.A. Comparative Effect of Elicitors on the Physiology and Secondary Metabolites in Broccoli Plants. J. Plant Physiol. 2019, 239, 1–9. [Google Scholar] [CrossRef]
- Ku, K.M.; Jeffery, E.H.; Juvik, J.A. Optimization of Methyl Jasmonate Application to Broccoli Florets to Enhance Health-Promoting Phytochemical Content. J. Sci. Food Agric. 2014, 94, 2090–2096. [Google Scholar] [CrossRef]
- Corbo, M.R.; Bevilacqua, A.; Petruzzi, L.; Casanova, F.P.; Sinigaglia, M. Functional Beverages: The Emerging Side of Functional Foods. Compr. Rev. Food Sci. Food Saf. 2014, 13, 1192–1206. [Google Scholar] [CrossRef]
- Radünz, M.; Hackbart, H.C.D.S.; Bona, N.P.; Pedra, N.S.; Hoffmann, J.F.; Stefanello, F.M.; Da Rosa Zavareze, E. Glucosinolates and Phenolic Compounds Rich Broccoli Extract: Encapsulation by Electrospraying and Antitumor Activity against Glial Tumor Cells. Colloids Surf. B Biointerfaces 2020, 192, 111020. [Google Scholar] [CrossRef] [PubMed]
- Aranaz, P.; Navarro-Herrera, D.; Romo-Hualde, A.; Zabala, M.; López-Yoldi, M.; González-Ferrero, C.; Gil, A.G.; Alfredo Martinez, J.; Vizmanos, J.L.; Milagro, F.I.; et al. Broccoli Extract Improves High Fat Diet-Induced Obesity, Hepatic Steatosis and Glucose Intolerance in Wistar Rats. J. Funct. Foods 2019, 59, 319–328. [Google Scholar] [CrossRef]
- Baenas, N.; Silván, J.M.; Medina, S.; de Pascual-Teresa, S.; García-Viguera, C.; Moreno, D.A. Metabolism and Antiproliferative Effects of Sulforaphane and Broccoli Sprouts in Human Intestinal (Caco-2) and Hepatic (HepG2) Cells. Phytochem. Rev. 2015, 14, 1035–1044. [Google Scholar] [CrossRef]
- Danafar, H.; Sharafi, A.; Kheiri Manjili, H.; Andalib, S. Sulforaphane Delivery Using MPEG–PCL Co-Polymer Nanoparticles to Breast Cancer Cells. Pharm. Dev. Technol. 2017, 22, 642–651. [Google Scholar] [CrossRef] [PubMed]
- Song, D.; Liang, H.; Kuang, P.; Tang, P.; Hu, G.; Yuan, Q. Instability and Structural Change of 4-Methylsulfinyl-3-Butenyl Isothiocyanate in the Hydrolytic Process. J. Agric. Food Chem. 2013, 61, 5097–5102. [Google Scholar] [CrossRef]
- Fahey, J.W.; Wade, K.L.; Stephenson, K.K.; Panjwani, A.A.; Liu, H.; Cornblatt, G.; Cornblatt, B.S.; Ownby, S.L.; Fuchs, E.; Holtzclaw, W.D.; et al. Bioavailability of Sulforaphane Following Ingestion of Glucoraphanin-Rich Broccoli Sprout and Seed Extracts with Active Myrosinase: A Pilot Study of the Effects of Proton Pump Inhibitor Administration. Nutrients 2019, 11, 1489. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zambrano, V.; Bustos, R.; Mahn, A. Insights about Stabilization of Sulforaphane through Microencapsulation. Heliyon 2019, e02951. [Google Scholar] [CrossRef]
- Danafar, H.; Sharaïfi, A.; Askarlou, S.; Manjili, H.K. Preparation and Characterization of PEGylated Iron Oxide-Gold Nanoparticles for Delivery of Sulforaphane and Curcumin. Drug Res. Stuttg. 2017, 67, 698–704. [Google Scholar] [CrossRef]
- Chalbi, N.; Martínez-Ballesta, M.C.; Youssef, N.B.; Carvajal, M. Intrinsic Stability of Brassicaceae Plasma Membrane in Relation to Changes in Proteins and Lipids as a Response to Salinity. J. Plant Physiol. 2015, 175, 148–156. [Google Scholar] [CrossRef]
- Maas, S.L.N.; Breakefield, X.O.; Weaver, A.M. Extracellular Vesicles: Unique Intercellular Delivery Vehicles. Trends Cell Biol. 2017, 27, 172–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Ibañez, P.; Nicolas-Espinosa, J.; Carvajal, M. Plasma Membrane Vesicles from Cauliflower Meristematic Tissue and Their Role in Water Passage. BMC Plant Biol. 2021, 21, 30. [Google Scholar] [CrossRef]
- Yepes-Molina, L.; Martínez-Ballesta, M.C.; Carvajal, M. Plant Plasma Membrane Vesicles Interaction with Keratinocytes Reveals Their Potential as Carriers. J. Adv. Res. 2020, 23, 101–111. [Google Scholar] [CrossRef] [PubMed]
- Patravale, V.B.; Naik, S.V.; Dhage, S.N. Role of Diet, Functional Foods, and Nutraceuticals in Brain Disorders. In Nutraceutical and Functional Foods in Disease Prevention; IGI Global: Hershey, PA, USA, 2018; pp. 256–287. [Google Scholar] [CrossRef]
- Gironés-Vilaplana, A.; Moreno, D.A.; García-Viguera, C. Phytochemistry and Biological Activity of Spanish Citrus Fruits. Food Funct. 2014. [Google Scholar] [CrossRef] [PubMed]
- Mazarakis, N.; Snibson, K.; Licciardi, P.V.; Karagiannis, T.C. The Potential Use of L-Sulforaphane for the Treatment of Chronic Inflammatory Diseases: A Review of the Clinical Evidence. Clin. Nutr. 2019, 39, 664–675. [Google Scholar] [CrossRef] [PubMed]
- Morales-Gurrola, G.; Simental-Mendía, L.E.; Castellanos-Juárez, F.X.; Salas-Pacheco, J.M.; Guerrero-Romero, F. The Triglycerides and Glucose Index Is Associated with Cardiovascular Risk Factors in Metabolically Obese Normal-Weight Subjects. J. Endocrinol. Investig. 2020, 43, 995–1000. [Google Scholar] [CrossRef]
- Lopez-Bastida, J.; Boronat, M.; Moreno, J.O.; Schurer, W. Costs, Outcomes and Challenges for Diabetes Care in Spain. Global. Health 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Björk, S. The Cost of Diabetes and Diabetes Care. Diabetes Res. Clin. Pract. 2001, 54, 13–18. [Google Scholar] [CrossRef]
- WHO. Reducing Sodium Intake to Reduce Blood Pressure and Risk of Cardiovascular Diseases in Adults; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Khan, M.J.; Gerasimidis, K.; Edwards, C.A.; Shaikh, M.G. Role of Gut Microbiota in the Aetiology of Obesity: Proposed Mechanisms and Review of the Literature. J. Obes. 2016, 2016, e7353642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An Obesity-Associated Gut Microbiome with Increased Capacity for Energy Harvest. Nature 2006, 444, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Green, M.; Arora, K.; Prakash, S. Microbial Medicine: Prebiotic and Probiotic Functional Foods to Target Obesity and Metabolic Syndrome. Int. J. Mol. Sci. 2020, 21, 2890. [Google Scholar] [CrossRef]
- Viadel, B.; Rivera, J.D.; Navarro, M.T.; Tenllado, I.; Carreres, J.E.; García, J.; Blasco, M.; Subirats, S. Equipo Modular de Digestión In Vitro. Patent N# ES2361983B1 19 April 2012. Banco de Patentes de la Generalitat Valenciana. Available online: http://bancodepatentes.gva.es/es/listado-de-patentes/-/asset_publisher/xoxK0ZQPxN2u/content/equipo-modular-de-digestion-in-vitro (accessed on 10 May 2021).
- Molly, K.; Woestyne, M.V.; de Smet, I.; Verstraete, W. Validation of the Simulator of the Human Intestinal Microbial Ecosystem (SHIME) Reactor Using Microorganism-Associated Activities. Microb. Ecol. Health Dis. 1994, 7, 191–200. [Google Scholar] [CrossRef]
- Molly, K.; Vande Woestyne, M.; Verstraete, W. Development of a 5-Step Multi-Chamber Reactor as a Simulation of the Human Intestinal Microbial Ecosystem. Appl. Microbiol. Biotechnol. 1993, 39, 254–258. [Google Scholar] [CrossRef]
- Cuevas-Sierra, A.; Riezu-Boj, J.I.; Guruceaga, E.; Milagro, F.I.; Martínez, J.A. Sex-Specific Associations between Gut Prevotellaceae and Host Genetics on Adiposity. Microorganisms 2020, 8, 938. [Google Scholar] [CrossRef]
- Van de Wiele, T.; van den Abbeele, P.; Ossieur, W.; Possemiers, S.; Marzorati, M. The Simulator of the Human Intestinal Microbial Ecosystem (SHIME®). In The Impact of Food Bioactives on Health: In Vitro and Ex Vivo Models; Springer: Berlin/Heidelberg, Germany, 2015; pp. 305–317. [Google Scholar] [CrossRef] [Green Version]
- Possemiers, S.; Verthé, K.; Uyttendaele, S.; Verstraete, W. PCR-DGGE-Based Quantification of Stability of the Microbial Community in a Simulator of the Human Intestinal Microbial Ecosystem. FEMS Microbiol. Ecol. 2004, 49, 495–507. [Google Scholar] [CrossRef]
- Hildebrand, F.; Tadeo, R.; Voigt, A.Y.; Bork, P.; Raes, J. LotuS: An Efficient and User-Friendly OTU Processing Pipeline. Microbiome 2014, 2, 30. [Google Scholar] [CrossRef] [Green Version]
- Baenas, N.; Moreno, D.A.; García-Viguera, C. Selecting Sprouts of Brassicaceae for Optimum Phytochemical Composition. J. Agric. Food Chem. 2012, 60, 11409–11420. [Google Scholar] [CrossRef] [PubMed]
- Baenas, N.; Suárez-Martínez, C.; García-Viguera, C.; Moreno, D.A. Bioavailability and New Biomarkers of Cruciferous Sprouts Consumption. Food Res. Int. 2017, 100, 497–503. [Google Scholar] [CrossRef]
- Fritz, V.A.; Justen, V.L.; Bode, A.M.; Schuster, T.; Wang, M. Glucosinolate Enhancement in Cabbage Induced by Jasmonic Acid Application. HortScience 2010, 45, 1188–1191. [Google Scholar] [CrossRef] [Green Version]
- Hassini, I.; Baenas, N.; Moreno, D.A.; Carvajal, M.; Boughanmi, N.; Martinez Ballesta, M.D.C. Effects of Seed Priming, Salinity and Methyl Jasmonate Treatment on Bioactive Composition of Brassica Oleracea Var. Capitata (White and Red Varieties) Sprouts. J. Sci. Food Agric. 2017, 97, 2291–2299. [Google Scholar] [CrossRef]
- Loivamäki, M.; Holopainen, J.K.; Nerg, A.M. Chemical Changes Induced by Methyl Jasmonate in Oilseed Rape Grown in the Laboratory and in the Field. J. Agric. Food Chem. 2004, 52, 7607–7613. [Google Scholar] [CrossRef] [PubMed]
- Yi, G.E.; Robin, A.H.K.; Yang, K.; Park, J.I.; Hwang, B.H.; Nou, I.S. Exogenous Methyl Jasmonate and Salicylic Acid Induce Subspecies-Specific Patterns of Glucosinolate Accumulation and Gene Expression in Brassica oleracea L. Molecules 2016, 21, 1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thiruvengadam, M.; Baskar, V.; Kim, S.H.; Chung, I.M. Effects of Abscisic Acid, Jasmonic Acid and Salicylic Acid on the Content of Phytochemicals and Their Gene Expression Profiles and Biological Activity in Turnip (Brassica Rapa Ssp. Rapa). Plant Growth Regul. 2016, 80, 377–390. [Google Scholar] [CrossRef]
- Wu, Y.; Zou, L.; Mao, J.; Huang, J.; Liu, S. Stability and Encapsulation Efficiency of Sulforaphane Microencapsulated by Spray Drying. Carbohydr. Polym. 2014, 102, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Uppal, S.; Kaur, K.; Kumar, R.; Kaur, N.D.; Shukla, G.; Mehta, S.K. Chitosan Nanoparticles as a Biocompatible and Efficient Nanowagon for Benzyl Isothiocyanate. Int. J. Biol. Macromol. 2018, 115, 18–28. [Google Scholar] [CrossRef]
- Rios, J.J.; Yepes-Molina, L.; Martinez-Alonso, A.; Carvajal, M. Nanobiofertilization as a Novel Technology for Highly Efficient Foliar Application of Fe and B in Almond Trees: Nanobiofertilization in Almond Trees. R. Soc. Open Sci. 2020, 7, e200905. [Google Scholar] [CrossRef] [PubMed]
- Abukhabta, S.; Khalil Ghawi, S.; Karatzas, K.A.; Charalampopoulos, D.; McDougall, G.; Allwood, J.W.; Verrall, S.; Lavery, S.; Latimer, C.; Pourshahidi, L.K.; et al. Sulforaphane-Enriched Extracts from Glucoraphanin-Rich Broccoli Exert Antimicrobial Activity against Gut Pathogens in Vitro and Innovative Cooking Methods Increase in Vivo Intestinal Delivery of Sulforaphane. Eur. J. Nutr. 2020. [Google Scholar] [CrossRef] [PubMed]
- Hwang, E.S.; Bornhorst, G.M.; Oteiza, P.I.; Mitchell, A.E. Assessing the Fate and Bioavailability of Glucosinolates in Kale (Brassica Oleracea) Using Simulated Human Digestion and Caco-2 Cell Uptake Models. J. Agric. Food Chem. 2019, 67, 9492–9500. [Google Scholar] [CrossRef] [PubMed]
- Moreda-Piñeiro, J.; Moreda-Piñeiro, A.; Romarís-Hortas, V.; Moscoso-Pérez, C.; López-Mahía, P.; Muniategui-Lorenzo, S.; Bermejo-Barrera, P.; Prada-Rodríguez, D. In-Vivo and in-Vitro Testing to Assess the Bioaccessibility and the Bioavailability of Arsenic, Selenium and Mercury Species in Food Samples. Trends Anal. Chem. 2011, 30, 324–345. [Google Scholar] [CrossRef]
- Lv, X.; Meng, G.; Li, W.; Fan, D.; Wang, X.; Espinoza-Pinochet, C.A.; Cespedes-Acuña, C.L. Sulforaphane and Its Antioxidative Effects in Broccoli Seeds and Sprouts of Different Cultivars. Food Chem. 2020, 316, 126216. [Google Scholar] [CrossRef]
- Del Carmen Martínez-Ballesta, M.; Pérez-Sánchez, H.; Moreno, D.A.; Carvajal, M. Plant Plasma Membrane Aquaporins in Natural Vesicles as Potential Stabilizers and Carriers of Glucosinolates. Colloids Surf. B Biointerfaces 2016, 143, 318–326. [Google Scholar] [CrossRef]
- Shertzer, H.G.; Senft, A.P. The Micronutrient Indole-3-Carbinol: Implications for Disease and Chemoprevention. Drug Metabol. Drug Interact. 2000, 17, 159–188. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, T.T.Y.; Teng, Z.; Chen, P.; Sun, J.; Wang, Q. Encapsulation of Indole-3-Carbinol and 3,3′-Diindolylmethane in Zein/Carboxymethyl Chitosan Nanoparticles with Controlled Release Property and Improved Stability. Food Chem. 2013, 139, 224–230. [Google Scholar] [CrossRef]
- Cirilli, R.; Gallo, F.R.; Multari, G.; Palazzino, G.; Mustazza, C.; Panusa, A. Study of Solvent Effect on the Stability of Isothiocyanate Iberin, a Breakdown Product of Glucoiberin. J. Food Compos. Anal. 2020, 92, 103515. [Google Scholar] [CrossRef]
- Luang-In, V.; Albaser, A.A.; Nueno-Palop, C.; Bennett, M.H.; Narbad, A.; Rossiter, J.T. Glucosinolate and Desulfo-Glucosinolate Metabolism by a Selection of Human Gut Bacteria. Curr. Microbiol. 2016, 73, 442–451. [Google Scholar] [CrossRef] [PubMed]
- Cuomo, V.; Luciano, F.B.; Meca, G.; Ritieni, A.; Mañes, J. Bioaccessibility of Glucoraphanin from Broccoli Using an in Vitro Gastrointestinal Digestion Model. CYTA J. Food 2015, 13, 361–365. [Google Scholar] [CrossRef]
- Pongmalai, P.; Fu, N.; Soponronnarit, S.; Chiewchan, N.; Devahastin, S.; Chen, X.D. Microwave Pretreatment Enhances the Formation of Cabbage Sulforaphane and Its Bioaccessibility as Shown by a Novel Dynamic Soft Rat Stomach Model. J. Funct. Foods 2018, 43, 186–195. [Google Scholar] [CrossRef]
- Chang, C.J.; Lin, C.S.; Lu, C.C.; Martel, J.; Ko, Y.F.; Ojcius, D.M.; Tseng, S.F.; Wu, T.R.; Chen, Y.Y.M.; Young, J.D.; et al. Ganoderma Lucidum Reduces Obesity in Mice by Modulating the Composition of the Gut Microbiota. Nat. Commun. 2015, 6, 7489. [Google Scholar] [CrossRef] [Green Version]
- Llopis, M.; Cassard, A.M.; Wrzosek, L.; Boschat, L.; Bruneau, A.; Ferrere, G.; Puchois, V.; Martin, J.C.; Lepage, P.; Le Roy, T.; et al. Intestinal Microbiota Contributes to Individual Susceptibility to Alcoholic Liver Disease. Gut 2016, 65, 830–839. [Google Scholar] [CrossRef]
- Cani, P.D. Human Gut Microbiome: Hopes, Threats and Promises. Gut 2018, 67, 1716–1725. [Google Scholar] [CrossRef]
- Zhang, Z.; Mocanu, V.; Cai, C.; Dang, J.; Slater, L.; Deehan, E.C.; Walter, J.; Madsen, K.L. Impact of Fecal Microbiota Transplantation on Obesity and Metabolic Syndrome—A Systematic Review. Nutrients 2019, 11, 2291. [Google Scholar] [CrossRef] [Green Version]
- Carding, S.; Verbeke, K.; Vipond, D.T.; Corfe, B.M.; Owen, L.J. Dysbiosis of the Gut Microbiota in Disease. Microb. Ecol. Heal. Dis. 2015, 26, 2619. [Google Scholar] [CrossRef] [PubMed]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What Is the Healthy Gut Microbiota Composition? A Changing Ecosystem across Age, Environment, Diet, and Diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Luz Calle, M. Statistical Analysis of Metagenomics Data. Genom. Inform. 2019, 17, e6. [Google Scholar] [CrossRef] [PubMed]
- Harmsen, H.; de Goffau, M. The Human Gut Microbiota. In Microbiota of the Human Body; Springer: Berlin/Heidelberg, Germany, 2016; Volume 902, pp. 5–32. [Google Scholar] [CrossRef]
- Liu, Y.; Song, X.; Zhou, H.; Zhou, X.; Xia, Y.; Dong, X.; Zhong, W.; Tang, S.; Wang, L.; Wen, S.; et al. Gut Microbiome Associates with Lipid-Lowering Effect of Rosuvastatin in Vivo. Front. Microbiol. 2018, 9, 530. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.K.; Chang, H.W.; Yan, D.; Lee, K.M.; Ucmak, D.; Wong, K.; Abrouk, M.; Farahnik, B.; Nakamura, M.; Zhu, T.H.; et al. Influence of Diet on the Gut Microbiome and Implications for Human Health. J. Transl. Med. 2017, 15, 73. [Google Scholar] [CrossRef] [Green Version]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial Signature of Dysbiosis in Gut Microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Fei, N.; Zhao, L. An Opportunistic Pathogen Isolated from the Gut of an Obese Human Causes Obesity in Germfree Mice. ISME J. 2013, 7, 880–884. [Google Scholar] [CrossRef]
- Wu, Y.; Shen, Y.; Zhu, Y.; Mupunga, J.; Zou, L.; Liu, C.; Liu, S.; Mao, J. Broccoli Ingestion Increases the Glucosinolate Hydrolysis Activity of Microbiota in the Mouse Gut. Int. J. Food Sci. Nutr. 2019, 70, 585–594. [Google Scholar] [CrossRef]
- Ridaura, V.K.; Faith, J.J.; Rey, F.E.; Cheng, J.; Duncan, A.E.; Kau, A.L.; Griffin, N.W.; Lombard, V.; Henrissat, B.; Bain, J.R.; et al. Gut Microbiota from Twins Discordant for Obesity Modulate Metabolism in Mice. Science 2013, 341, 1241214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial Ecology: Human Gut Microbes Associated with Obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Zou, Y.; Ju, X.; Chen, W.; Yuan, J.; Wang, Z.; Aluko, R.E.; He, R. Rice Bran Attenuated Obesity via Alleviating Dyslipidemia, Browning of White Adipocytes and Modulating Gut Microbiota in High-Fat Diet-Induced Obese Mice. Food Funct. 2020, 11, 2406–2417. [Google Scholar] [CrossRef]
- Larsen, N.; Vogensen, F.K.; Gøbel, R.J.; Michaelsen, K.F.; Forssten, S.D.; Lahtinen, S.J.; Jakobsen, M. Effect of Lactobacillus Salivarius Ls-33 on Fecal Microbiota in Obese Adolescents. Clin. Nutr. 2013, 32, 935–940. [Google Scholar] [CrossRef] [PubMed]
- Finucane, M.M.; Sharpton, T.J.; Laurent, T.J.; Pollard, K.S. A Taxonomic Signature of Obesity in the Microbiome? Getting to the Guts of the Matter. PLoS ONE 2014, 9, e84689. [Google Scholar] [CrossRef] [Green Version]
- Cortés-martín, A.; Iglesias-aguirre, C.E.; Meoro, A.; Selma, M.V.; Espín, J.C. There Is No Distinctive Gut Microbiota Signature in the Metabolic Syndrome: Contribution of Cardiovascular Disease Risk Factors and Associated Medication. Microorganisms 2020, 8, 416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoun, A.; Darwish, F.; Hamod, N. The Influence of the Gut Microbiome on Obesity in Adults and the Role of Probiotifcs Prebiotics and Synbiotics for Weight Loss. Prev. Nutr. Food Sci. 2020, 25, 113–123. [Google Scholar] [CrossRef]
- Louis, P.; Flint, H.J. Formation of Propionate and Butyrate by the Human Colonic Microbiota. Environ. Microbiol. 2017, 19, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Yi, C.X.; Katiraei, S.; Kooijman, S.; Zhou, E.; Chung, C.K.; Gao, Y.; Van Den Heuvel, J.K.; Meijer, O.C.; Berbée, J.F.P.; et al. Butyrate Reduces Appetite and Activates Brown Adipose Tissue via the Gut-Brain Neural Circuit. Gut 2018, 67, 1269. [Google Scholar] [CrossRef] [Green Version]
- Coppola, S.; Avagliano, C.; Calignano, A.; Berni Canani, R. The Protective Role of Butyrate against Obesity and Obesity-Related Diseases. Molecules 2021, 26, 682. [Google Scholar] [CrossRef]
- Cleophas, M.C.P.; Ratter, J.M.; Bekkering, S.; Quintin, J.; Schraa, K.; Stroes, E.S.; Netea, M.G.; Joosten, L.A.B. Effects of Oral Butyrate Supplementation on Inflammatory Potential of Circulating Peripheral Blood Mononuclear Cells in Healthy and Obese Males. Sci. Rep. 2019, 9, 775. [Google Scholar] [CrossRef] [PubMed]
- Goswami, C.; Iwasaki, Y.; Yada, T. Short-Chain Fatty Acids Suppress Food Intake by Activating Vagal Afferent Neurons. J. Nutr. Biochem. 2018, 57, 130–135. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Glucosinolate (mg g D.W.−1) | Control | 200 μM SA | 100 μM MeJA | SA + MeJA |
|---|---|---|---|---|
| GIB | * | * | * | * |
| GRA | * | * | * | * |
| SIN | 4.12 ± 0.13 a | 3.73 ± 0.08 b | 4.95 ± 0.04 a | 4.12 ± 0.06 ab |
| GNA | * | * | * | * |
| HGB | 0.77 ± 0.02 b | 0.41 ± 0.03 c | 1.79 ± 0.02 a | 0.54 ± 0.02 c |
| GB | 2.93 ± 0.05 c | 2.24 ± 0.08 c | 8.1 ± 0.06 a | 5.01 ± 0.1 b |
| Total GSLs | 8.13 ± 0.2 b | 6.39 ± 0.03 c | 14.82 ± 0.01 a | 9.71 ± 0.08 b |
| Red Cabbage Aqueous Extract | ||
|---|---|---|
| ITCs Composition (μg/mL) | Free Extract | Nanoencapsulated |
| Sulforaphane (SFN) | 6.72 ± 0.68 a | 5.64 ± 0.28 a |
| Indole-3-carbinol (I3C) | 1.81 ± 0.15 a | 1.05 ± 0.1 a |
| Iberin | 1.82 ± 0.04 a | 1.97 ± 0.06 a |
| Bacteroidetes/Firmicutes Ratio | Free Red Cabbage Aqueous Extract | Red Cabbage Nanoencapsulated Aqueous Extract | ||
|---|---|---|---|---|
| Stabilization | 14 Days Treatment | Stabilization | 14 Days Treatment | |
| Inoculation | 1.00 ± 0.02 a | - | 0.98 ± 0.07 a | - |
| Ascending colon | 0.15 ± 0.01 c | 0.19 ± 0.08 c | 0.5 ± 0.03 a | 0.39 ± 0.02 b |
| Transversal colon | 1.08 ± 0.1 ab | 1.14 ± 0.08 a | 1.21 ± 0.16 a | 0.95 ± 0.05 b |
| Descending colon | 1.08 ± 0.03 ab | 1.09 ± 0.04 ab | 1.16 ± 0.08 a | 0.90 ± 0.01 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Garcia-Ibañez, P.; Roses, C.; Agudelo, A.; Milagro, F.I.; Barceló, A.M.; Viadel, B.; Nieto, J.A.; Moreno, D.A.; Carvajal, M. The Influence of Red Cabbage Extract Nanoencapsulated with Brassica Plasma Membrane Vesicles on the Gut Microbiome of Obese Volunteers. Foods 2021, 10, 1038. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10051038
Garcia-Ibañez P, Roses C, Agudelo A, Milagro FI, Barceló AM, Viadel B, Nieto JA, Moreno DA, Carvajal M. The Influence of Red Cabbage Extract Nanoencapsulated with Brassica Plasma Membrane Vesicles on the Gut Microbiome of Obese Volunteers. Foods. 2021; 10(5):1038. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10051038
Chicago/Turabian StyleGarcia-Ibañez, Paula, Carles Roses, Agatha Agudelo, Fermin I. Milagro, Ana M. Barceló, Blanca Viadel, Juan Antonio Nieto, Diego A. Moreno, and Micaela Carvajal. 2021. "The Influence of Red Cabbage Extract Nanoencapsulated with Brassica Plasma Membrane Vesicles on the Gut Microbiome of Obese Volunteers" Foods 10, no. 5: 1038. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10051038