
Effect of Freeze Drying and Simulated Gastrointestinal Digestion on Phenolic Metabolites and Antioxidant Property of the Natal Plum (Carissa macrocarpa)

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents and Standards
2.2. Preparation of Natal Plum Fruit Powder (NPFP)
2.3. In Vitro Digestion of NPFP
2.4. Extraction of Phenolic Compounds
2.5. Total Phenolic Content
2.6. Antioxidant Capacity
2.7. In Vitro α-Glucosidase Inhibitory Activity
2.8. Quantification of Targeted and Untargeted Phenolic Metabolites
2.9. Statistical Analysis
3. Results and Discussion
3.1. Changes in Physicochemical Properties of Natal Plum during Freeze Drying
3.2. Characterisation, Identification of Phenolic Components from Natal Plum and NPFP
3.3. Changes in Total Phenolic Content, Phenolic Components, and Antioxidant Properties during Freeze Drying
3.4. Changes in Total Phenolic Content of NPFP during In Vitro Gastrointestinal Digestion
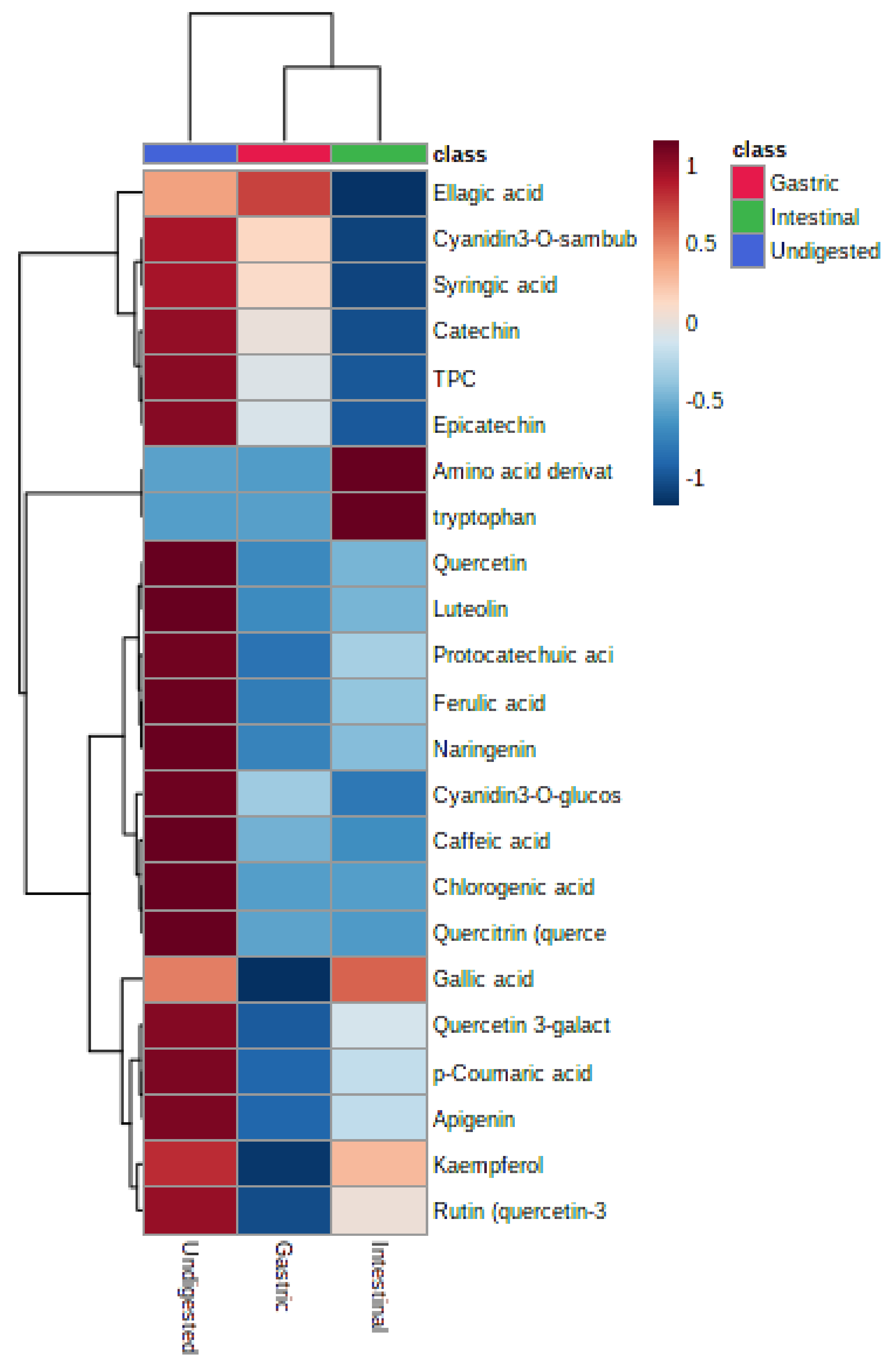
3.5. Release of Different Phenolic Components from NPFP during In Vitro Gastrointestinal Digestion
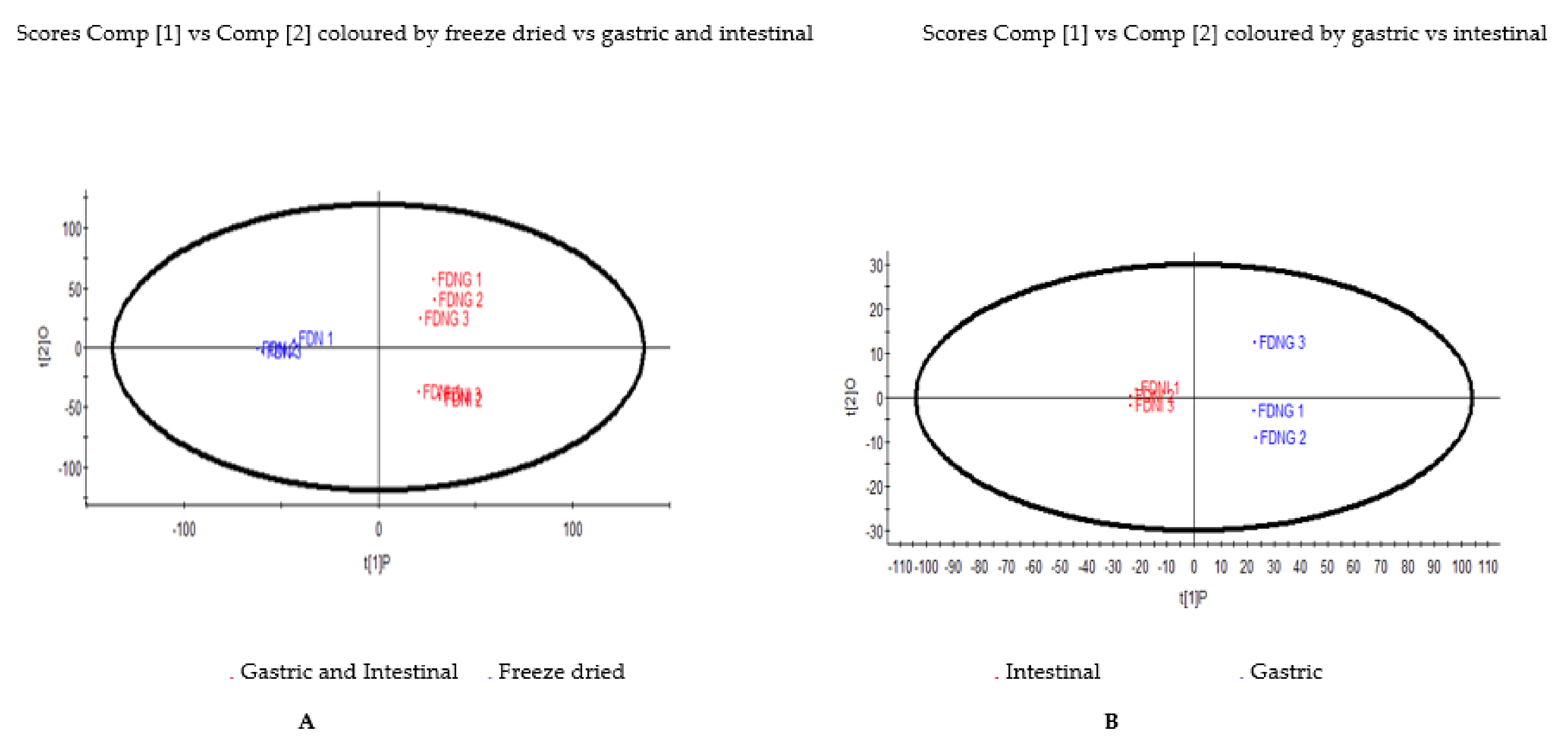
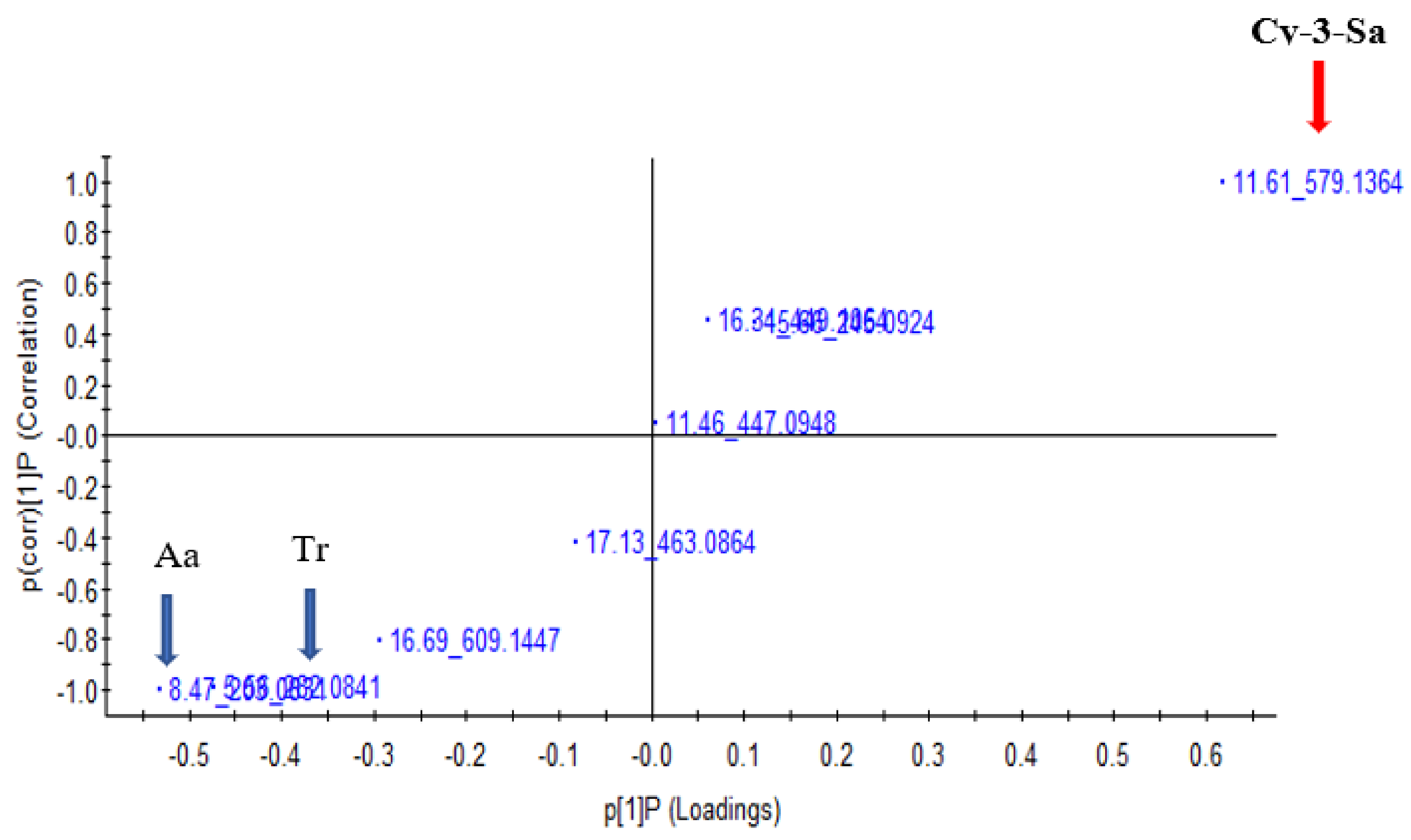
3.6. Principal Component Analysis
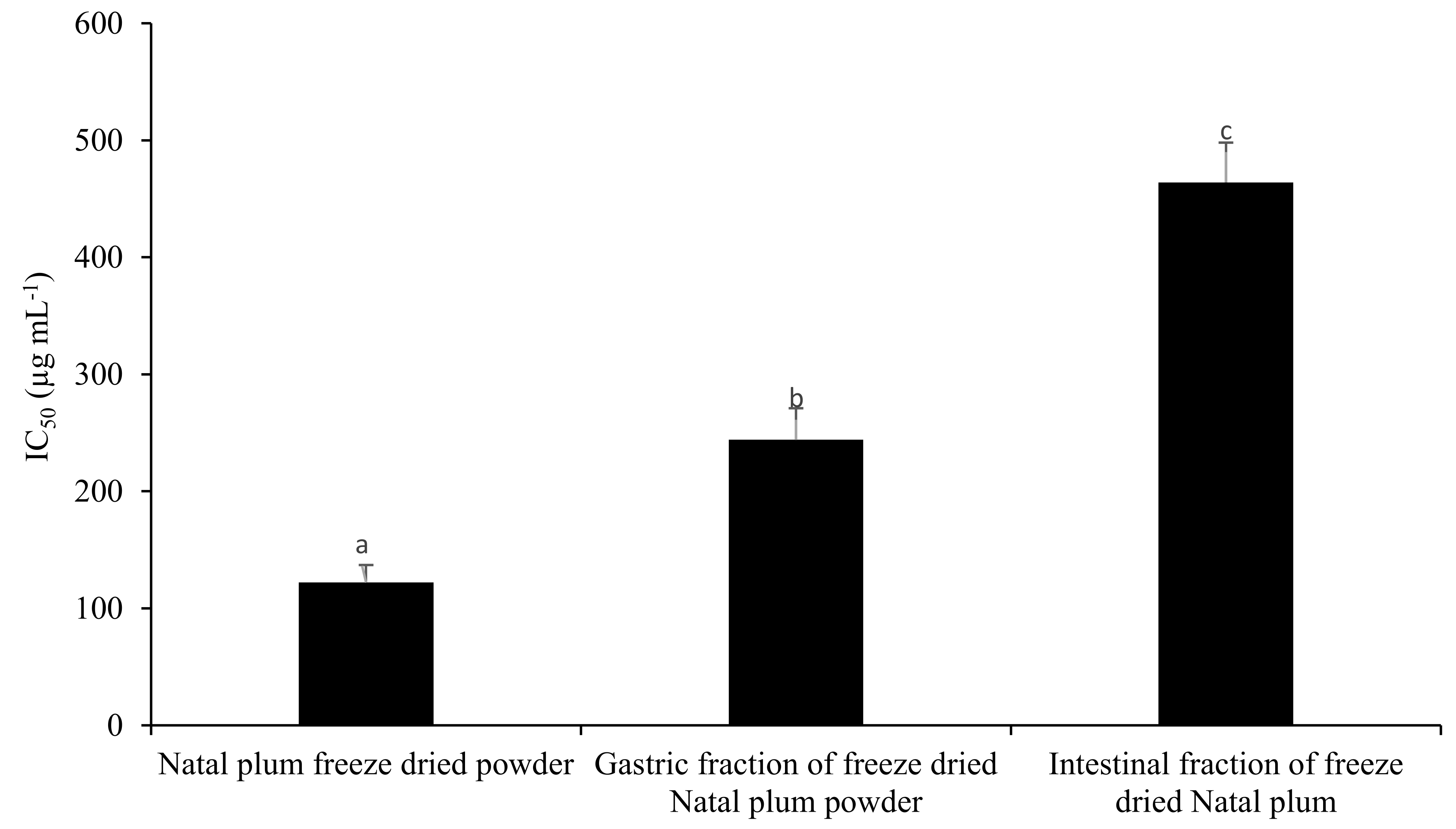
3.7. Antioxidant Capacity of Gastric and Intestinal Fraction of NPFP
3.8. α-Glucosidase Inhibitory Activity of Digested and Nondigested Natal Plum
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Report Link 2021. Functional Foods Market Size, Share & Trends Analysis Report by Ingredient, by Product, by Application and Segment Forecasts, 2019–2025. Available online: https://www.researchandmarkets.com/reports/4764576/functional-foods-market-size-share-and-trends (accessed on 2 February 2021).
- Business Wire Technovo Market Report 2021. Global Functional Powder Drinks Concentrate Market: Growth Analysis and Forecast by Technavio. Available online: https://www.businesswire.com/news/home/20170313006089/en/Global-Functional-Powder-Drinks-Concentrates-Market-Growth-Analysis-and-Forecast-by-Technavio (accessed on 2 February 2021).
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ndou, A.; Tinyani, P.P.; Slabbert, R.M.; Sultanbawa, Y.; Sivakumar, D. An integrated approach for harvesting Natal plum (Carissa macrocarpa) for quality and functional compounds related to maturity stages. Food Chem. 2019, 293, 499–510. [Google Scholar] [CrossRef] [PubMed]
- Kara, Ş.; Erçelebi, E.A. Thermal degradation kinetics of anthocyanins and visual colour of Urmu mulberry (Morus nigra L.). J. Food Eng. 2013, 116, 541–547. [Google Scholar] [CrossRef]
- Krokida, M.K.; Maroulis, Z.B.; Saravacos, G.D. The effect of the method of drying on the colour of dehydrated products. Int. J. Food Sci. Technol. 2001, 36, 53–59. [Google Scholar] [CrossRef]
- Melini, V.; Melini, F.; Acquistucci, R. Phenolic compounds and bioaccessibility thereof in functional pasta. Antioxidants 2020, 9, 343. [Google Scholar] [CrossRef] [Green Version]
- Tomas, M.; Beekwilder, J.; Hall, R.D.; Simon, C.D.; Sagdic, O.; Capanoglu, E. Effect of dietary fiber (inulin) addition on phenolics and in vitro bioaccessibility of tomato sauce. Food Res. Int. 2018, 106, 129–135. [Google Scholar] [CrossRef]
- da Silva, E.N.; de Farias, L.O.; Cadore, S. The total concentration and bioaccessible fraction of nutrients in purées, instant cereals and infant formulas by ICP OES: A study of Dietary Recommended Intakes and the importance of using a standardized in vitro digestion method. J. Food Compos. Anal. 2018, 68, 65–72. [Google Scholar] [CrossRef]
- Promchan, J.; Shiowatana, J.A. dynamic continuous-flow dialysis system with on-line electrothermal atomic-absorption spectrometric and pH measurements for in vitro determination of iron bioavailability by simulated gastrointestinal digestion. Anal. Bioanal. Chem. 2005, 382, 1360–1367. [Google Scholar] [CrossRef]
- Pérez-Vicente, A.; Gil-Izquierdo, A.; García-Viguera, C. In vitro gastrointestinal digestion study of pomegranate juice phenolic compounds, anthocyanins, and vitamin C. J. Agric. Food Chem. 2002, 50, 2308–2312. [Google Scholar] [CrossRef]
- Bouayed, J.; Hoffmann, L.; Bohn, T. Total phenolics, flavonoids, anthocyanins and antioxidant activity following simulated gastro-intestinal digestion and dialysis of apple varieties: Bioaccessibility and potential uptake. Food Chem. 2011, 128, 14–21. [Google Scholar] [CrossRef]
- Rodríguez, M.; Rojas-Graü, M.A.; Elez-Martínez, P.; Martín-Belloso, O. Changes in vitamin C, phenolic, and carotenoid profiles throughout in vitro gastrointestinal digestion of a blended fruit juice. J. Agric. Food Chem. 2013, 61, 1859–1867. [Google Scholar] [CrossRef] [PubMed]
- Gil-Izquierdo, A.; Gil, M.I.; Ferreres, F.; Tomás-Barberán, F.A. In vitro availability of flavonoids and other phenolics in orange juice. J. Agric. Food Chem. 2001, 49, 1035–1041. [Google Scholar] [CrossRef] [PubMed]
- Dalmau, M.E.; Bornhorst, G.M.; Eim, V.; Rosselló, C.; Simal, S. Effects of freezing, freeze drying and convective drying on in vitro gastric digestion of apples. Food Chem. 2017, 215, 7–16. [Google Scholar] [CrossRef] [Green Version]
- Kuljarachanan, T.; Chiewchan, N.; Devahastin, S. Mechanical Grinding Effects on Health-Related Functional Properties of Dietary Fiber Powder from White Cabbage By-products. J. Adv. Agric. Technol. 2019, 6, 154–160. [Google Scholar] [CrossRef]
- Jara-Palacios, M.J.; Gonçalves, S.; Hernanz, D.; Heredia, F.J.; Romano, A. Effects of in vitro gastrointestinal digestion on phenolic compounds and antioxidant activity of different white winemaking by products extracts. Food Res. Int. 2018, 109, 433–439. [Google Scholar] [CrossRef]
- Mpai, S.; Du Preez, R.; Sultanbawa, Y.; Sivakumar, D. Phytochemicals and nutritional composition in accessions of Kei-apple (Dovyalis caffra): Southern African indigenous fruit. Food Chem. 2018, 253, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Apostolidis, E.; Lee, C.M. In vitro potential of Ascophyllum nodosum phenolic antioxidant-mediated α-glucosidase and α-amylase inhibition. J. Food Sci. 2010, 75, 97–102. [Google Scholar] [CrossRef]
- Rezaei, F.; Vandergheynst, J.S. Critical moisture content for microbial growth in dried food-processing residues. J. Sci. Food Agric. 2010, 90, 2000–2005. [Google Scholar] [CrossRef]
- Hammami, C.; René, F. Determination of freeze-drying process variables for strawberries. J. Food Eng. 1997, 32, 133–154. [Google Scholar] [CrossRef]
- Sinela, A.; Rawat, N.; Mertz, C.; Achir, N.; Fulcrand, H.; Dornier, M. Anthocyanins degradation during storage of Hibiscus sabdariffa extract and evolution of its degradation products. Food Chem. 2017, 214, 234–241. [Google Scholar] [CrossRef]
- Sun, J.; Lin, L.; Chen, P. Study of the mass spectrometric behaviors of anthocyanins in negative ionization mode and its applications for characterization of anthocyanins and non-anthocyanin polyphenols. Rapid Commun. Mass. Spectrom. 2012, 26, 1123–1133. [Google Scholar] [CrossRef]
- Ezzat, S.M.; Salama, M.M.; Seif el-Din, S.H.; Saleh, S.; El-Lakkany, N.M.; Hammam, O.A.; Botros, S.S. Metabolic profile and hepatoprotective activity of the anthocyanin-rich extract of Hibiscus sabdariffa calyces. Pharm Biol. 2016, 54, 3172–3181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDougall, G.J.; Dobson, P.; Smith, P.; Blake, A.; Stewart, D. Assessing potential bioavailability of raspberry anthocyanins using an in vitro digestion system. J. Agric. Food Chem. 2005, 53, 5896–5904. [Google Scholar] [CrossRef]
- Mashitoa, F.M.; Shoko, T.; Shai, J.L.; Slabbert, R.M.; Sivakumar, D. Changes in phenolic metabolites and biological activities of pumpkin leaves (Cucurbita moschata Duchesne ex Poir.) during blanching. Front. Nutr. 2021, 8, 86. [Google Scholar] [CrossRef]
- Shofian, N.M.; Hamid, A.A.; Osman, A.; Saari, N.; Anwar, F.; Pak Dek, M.S.; Hairuddin, M.R. Effect of freeze-drying on the antioxidant compounds and antioxidant activity of selected tropical fruits. Int. J. Mol. Sci. 2011, 12, 4678–4692. [Google Scholar] [CrossRef] [Green Version]
- Lucas, B.F.; Zambiazi, R.C.; Costa, J.A.V. Biocompounds and physical properties of açaí pulp dried by different methods. LWT Food Sci. Technol. 2018, 98, 335–340. [Google Scholar] [CrossRef]
- Michalska-Ciechanowska, A.; Wojdyło, A.; Lech, K.; Łysiak, G.P.; Figiel, A. Effect of different drying techniques on physical properties, total polyphenols and antioxidant capacity of blackcurrant pomace powder. LWT Food Sci. Technol. 2017, 78, 114–121. [Google Scholar] [CrossRef]
- Leusink, G.J.; Kitts, D.D.; Yaghmaee, P.; Durance, T. Retention of Antioxidant Capacity of Vacuum Microwave Dried Cranberry. J. Food Sci. 2010, 75, 311–316. [Google Scholar] [CrossRef]
- De Torres, C.; Díaz-Maroto, M.C.; Hermosín-Gutiérrez, I.; Pérez-Coello, M.S. Effect of freeze-drying and oven-drying on volatiles and phenolics composition of grape skin’. Anal. Chim. Acta. 2010, 660, 177–182. [Google Scholar] [CrossRef]
- Bae, H.S.; Kim, H.J.; Kang, J.H.; Kudo, R.; Hosoya, T.; Kumazawa, S.; Ahn, M. Anthocyanin profile and antioxidant activity of various berries cultivated in Korea. Nat. Prod. Commun. 2015, 10. [Google Scholar] [CrossRef] [Green Version]
- Ketnawa, S.; Suwannachot, J.; Ogawa, Y. In vitro gastrointestinal digestion of Crisphead lettuce: Changes in bioactive compounds and antioxidant potential. Food Chem. 2020, 311, 125885. [Google Scholar] [CrossRef] [PubMed]
- Fawole, O.A.; Opara, U.L. Stability of total phenolic concentration and antioxidant capacity of extracts from pomegranate co-products subjected to in vitro digestion. BMC Compl. Altern. Med. 2016, 16, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ariza, M.T.; Forbes-Hernandez, T.Y.; Giampieri, F.; Gasparrini, M.; Soria, C.; Martínez-Ferri, E.; Battino, M. Effects of in vitro gastrointestinal digestion on strawberry polyphenols stability. Acta. Hortic. 2017, 1156, 396. [Google Scholar] [CrossRef]
- Marhuenda, J.; Alemán, M.D.; Gironés-Vilaplana, A.; Pérez, A.; Caravaca, G.; Figueroa, F.; Zafrilla, P. Phenolic composition, antioxidant activity, and in vitro availability of four different berries. J. Chem. 2016, 2016, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Las Heras, R.; Pinazo, A.; Heredia, A.; Andrés, A. Evaluation studies of persimmon plant (Diospyros kaki) for physiological benefits and bioaccessibility of antioxidants by in vitro simulated gastrointestinal digestion. Food Chem. 2017, 214, 478–541. [Google Scholar] [CrossRef]
- Alminger, M.; Aura, A.M.; Bohn, T.; Dufour, C.; El, S.N.; Gomes, A.; Santos, C.N. In vitro models for studying secondary plant metabolite digestion and bioaccessibility. Compr. Rev. Food Sci. Food Saf. 2014, 13, 413–436. [Google Scholar] [CrossRef] [Green Version]
- Gil-Izquierdo, A.; Zafrilla, P.; Tomás-Barberán, F.A. An in vitro method to simulate phenolic compound release from the food matrix in the gastrointestinal tract. Eur. Food Res. Technol. 2002, 214, 155–159. [Google Scholar] [CrossRef]
- da Silva, F.L.; Escribano-Bailón, M.T.; Alonso, J.J.P.; Rivas-Gonzalo, J.C.; Santos-Buelga, C. Anthocyanin pigments in strawberry. LWT Food Sci. Technol. 2007, 40, 374–382. [Google Scholar] [CrossRef]
- Schreckinger, M.E.; Lotton, J.; Lila, M.A.; de Mejia, E.G. Berries from South America: A Comprehensive Review on Chemistry, Health Potential, and Commercialization. J. Med. Food 2010, 13, 233–246. [Google Scholar] [CrossRef]
- Hao, Y.; Yang, J.; Cui, J.; Fan, Y.; Li, N.; Wang, C.; Dong, Y. Stability and mechanism of phenolic compounds from raspberry extract under in vitro gastrointestinal digestion. LWT Food Sci. Technol. 2021, 139, 110552. [Google Scholar] [CrossRef]
- Talavera, S.; Felgines, C.; Texier, O.; Besson, C.; Lamaison, J.L.; Rémésy, C. Anthocyanins are efficiently absorbed from the stomach in anesthetized rats. J. Nutr. 2003, 133, 4178–4182. [Google Scholar] [CrossRef]
- Crozier, A.; Jaganath, I.B.; Clifford, M.N. Dietary phenolics: Chemistry, bioavailability and effects on health. Nat. Prod. Rep. 2009, 26, 1001–1043. [Google Scholar] [CrossRef]
- Kamonpatana, K.; Failla, M.L.; Kumar, P.S.; Giusti, M.M. Anthocyanin structure determines susceptibility to microbial degradation and bioavailability to the buccal mucosa. J. Agric. Food Chem. 2014, 62, 6903–6910. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, H.; Zhang, W.; Ding, Y.; Zhao, T.; Zhang, M.; Yang, L. Bioaccessibility and biotransformation of anthocyanin monomers following in vitro simulated gastric-intestinal digestion and in vivo metabolism in rats. Food Funct. 2019, 10, 6052–6061. [Google Scholar] [CrossRef]
- Oliveira, G.; Tylewicz, U.; Dalla Rosa, M.; Andlid, T.; Alminger, M. Effects of pulsed electric field-assisted osmotic dehydration and edible coating on the recovery of anthocyanins from in vitro digested berries. Foods 2019, 8, 505. [Google Scholar] [CrossRef] [Green Version]
- Oidtmann, J.; Schantz, M.; Mäder, K.; Baum, M.; Berg, S.; Betz, M.; Richling, E. Preparation and comparative release characteristics of three anthocyanin encapsulation systems. J. Agric. Food Chem. 2012, 60, 844–851. [Google Scholar] [CrossRef]
- Mosele, J.I.; Macià, A.; Romero, M.P.; Motilva, M.J. Stability and metabolism of Arbutus unedo bioactive compounds (phenolics and antioxidants) under in vitro digestion and colonic fermentation. Food Chem. 2016, 201, 120–130. [Google Scholar] [CrossRef]
- Tagliazucchi, D.; Verzelloni, E.; Bertolini, D.; Conte, A. In vitro bio-accessibility and antioxidant activity of grape polyphenols. Food Chem. 2010, 120, 599–606. [Google Scholar] [CrossRef]
- Rha, C.S.; Seong, H.; Jung, Y.S.; Jang, D.; Kwak, J.G.; Kim, D.O.; Han, N.S. Stability and fermentability of green tea flavonols in in vitro-simulated gastrointestinal digestion and human faecal fermentation. Int. J. Mol. Sci. 2019, 20, 5890. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez, M.; TironI, V.A. Polyphenols in amaranth (A. manteggazianus) flour and protein isolate: Interaction with other components and effect of the gastrointestinal digestion. Food Res. Int. 2020, 137, 109524. [Google Scholar] [CrossRef]
- Dutra, R.L.T.; Dantas, A.M.; de Araújo Marques, D.; Batista, J.D.F.; de Albuquerque Meireles, B.R.L.; de Magalhães Cordeiro, Â.M.T.; Borges, G.D.S.C. Bioaccessibility and antioxidant activity of phenolic compounds in frozen pulps of Brazilian exotic fruits exposed to simulated gastrointestinal conditions. Food Res. Int. 2017, 100, 650–657. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.Q.; Zhang, A.; Tsang, D.; Huang, Y.; Chen, Z.Y. Stability of Green tea catechins. J. Agric. Food Chem. 1997, 45, 4624–4628. [Google Scholar] [CrossRef]
- Xiao, J. Dietary flavonoid aglycones and their glycosides: Which show better biological significance? Crit. Rev. Food Sci. Nutr. 2017, 57, 1874–1905. [Google Scholar] [CrossRef] [PubMed]
- Hollman, P.C.; Van Trijp, J.M.; Buysman, M.N.; vd Gaag, M.S.; Mengelers, M.J.; De Vries, J.H.; Katan, M.B. Relative bioavailability of the antioxidant flavonoid quercetin from various foods in man. FEBS Lett. 1997, 418, 152–156. [Google Scholar] [CrossRef] [Green Version]
- Celep, E.; Charehsaz, M.; Akyüz, S.; Acar, E.T.; Yesilada, E. Effect of in vitro gastrointestinal digestion on the bioavailability of phenolic components and the antioxidant potentials of some Turkish fruit wines. Food Res. Int. 2015, 78, 209–215. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Ambigaipalan, P. Phenolics and polyphenolics in foods, beverages and spices: Antioxidant activity and health effects–A review. J. Funct. Foods 2015, 18, 820–897. [Google Scholar] [CrossRef]
- Kim, D.O.; Jeong, S.W.; Lee, C.Y. Antioxidant capacity of phenolic phytochemicals from various cultivars of plums. Food Chem. 2003, 81, 321–326. [Google Scholar] [CrossRef]
- Gowd, V.; Jia, Z.; Chen, W. Anthocyanins as promising molecules and dietary bioactive components against diabetes–A review of recent advances. Trends Food Sci. Technol. 2017, 68, 1–13. [Google Scholar] [CrossRef]
- Poovitha, S.; Parani, M. In vitro and in vivo α-amylase and α-glucosidase inhibiting activities of the protein extracts from two varieties of bitter gourd (Momordica charantia L.). BMC Complement. Altern. Med. 2016, 16, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Tadera, K.; Minami, Y.; Takamatsu, K.; Matsuoka, T. Inhibition of α-glucosidase and α-amylase by flavonoids. J. Nutr. Sci. Vitaminol. 2006, 52, 149–153. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Physicochemical Property | Fresh Natal Plum | NPFP |
|---|---|---|
| Moisture content (g /100 g) | 77.2 ± 1.3 a | 6.4 ± 0.8 b |
| pH | 3.19 ± 0.1 a | 3.21 ± 0.1 a |
| Colour coordinates | ||
| L* | 36.6 ± 1.2 b | 41.4 ± 2.2 a |
| a* | 20.2 ± 2.1 a | 17.4 ± 0.7 b |
| b* | 6.3 ± 0.5 a | 7.7 ± 0.9 a |
| Fresh Natal Plum (DW) | Freeze-Dried Natal Plum (Non-Digested) | % Loss | Gastric Digested Natal Plum (DW) | Recovery % | Intestinal Digested Natal Plum (DW) | Bioaccessibility (%) | |
|---|---|---|---|---|---|---|---|
| TPC (mg GAE kg−1) | 2576.05 a ± 4.85 | 2265.77 a ± 1.23 * | 12.04 | 1786.13 b ± 0.08 | 78.8 | 1399.18 c ± 0.16 | 61.8 |
| Phenolic components (mg kg−1) | |||||||
| Anthocyanins | |||||||
| Cyanidin-3-O-sambubioside | 1688.1 a ± 4.3 | 1631.4 a ± 1.2 | 3.36 | 1193.2 b ± 1.5 | 73.1 | 525.3 c ± 3.1 | 32.2 |
| Cyanidin-3-O-glucoside | 1049.5 a± 2.2 | 955.2 a ± 1.9 | 8.99 | 347.6 b ± 1.5 | 36.4 | 155.7 c ± 1.8 | 16.3 |
| Hydroxybenzoic acids | |||||||
| Ellagic acid | 26.0 c ± 0.4 | 42.4 b ± 0.1 | +63.07 | 49.6 a ± 0.5 | 116.9 | 11.3 d ± 0.2 | 26.7 |
| Gallic acid | 3.8 c ± 0.4 | 5.5 b ± 0.1 | +44.74 | 2.1 c ± 0.1 | 38.2 | 8.8 a ± 0.1 | 160 |
| Protocatechuic acid | 158.8 b ± 5.0 | 167.6 a ± 0.9 | +5.54 | 59.6 d ± 0.2 | 35.6 | 88.6 c ± 0.5 | 52.9 |
| Hydroxycinnamic acids | |||||||
| p-Coumaric acid | 226.4 a ± 5.2 | 140.3 b ± 1.6 | 38.03 | 55.4 d ± 0.5 | 39.5 | 85.7 c ± 0.2 | 61 |
| Ferulic acid | 126.4 b ± 4.6 | 165.1 a ± 0.3 | +30.61 | 49.6 d ± 0.3 | 30 | 73.5 c ± 0.3 | 44.5 |
| Caffeic acid | 120.1 b ± 1.26 | 168.4 a ± 0.1 | +40.22 | 53.8 c ± 0.5 | 31.9 | 40.8 d ± 0.4 | 24.2 |
| Syringic acid | 100.4 a ± 4.2 | 62.2 b ± 0.2 | 38.05 | 47.5 c ± 0.6 | 76.4 | 26.5 d ± 0.2 | 42.6 |
| Flavonoids | |||||||
| Catechin | 497.7 a ± 9.7 | 491.8 a ± 2.3 | 0.0 | 354.1 b ± 2.8 | 72 | 210.4 c ± 1.4 | 42.8 |
| Epicatechin | 21.3 a ± 0.6 | 19.3 a ± 0.2 | 9.39 | 12.2 b ± 0.2 | 63 | 6.7 c ± 0.2 | 34.7 |
| Kaempferol | 493.5 a ± 2.3 | 333.5 b ± 1.2 | 32.42 | 70.9 d ± 3.5 | 21.3 | 259.9 c ± 3.1 | 21.3 |
| Quercetin | 210.2 b ± 0.8 | 279.7 a ± 1.4 | +33.06 | 88.6 d ± 5.9 | 31.7 | 112.1 c ± 0.9 | 40 |
| Naringenin | 37.4 b ± 1.3 | 48.3 a ± 0.2 | +29.14 | 17.2 c ± 0.1 | 35.6 | 22.3 c ± 0.2 | 46 |
| Apigenin | 4.69 | 9.7 a ± 0.6 | +106.82 | 3.8 c ± 0.1 | 39 | 5.9 b ± 0.1 | 60.8 |
| Luteolin | 3.4 b ± 0.5 | 4.6 a ± 0.1 | +26.09 | 1.3 c ± 0.1 | 28.3 | 1.7 c ± 0.1 | 36.9 |
| Flavonoid glycoside | |||||||
| Quercitrin (quercetin-O-glycoside) | 30.5 a ± 1.2 | 5.3 b ± 1.9 | 82.62 | 1.0 c ± 0.1 | 18.9 | 0.9 c ± 0.1 | 16.9 |
| Quercetin-3-galactoside 7-rhamnoside | 73.6 a ± 1.6 | 7.8 b ± 0.6 | 89.40 | 0.9 d ± 0.1 | 11.5 | 3.8 c ± 0.1 | 48.7 |
| Quercetin 3-O-rutinoside (Rutin) | 50.4 a ± 0.98 | 34.6 b ± 1.8 | 31.35 | 0.9 d ± 0.1 | 25 | 18.5 c ± 1.4 | 53.5 |
| Amino Acid components (mg kg−1) | |||||||
| Amino acid derivatives | ND | 0.2 b ± 0.1 | 0.08 c ± 0.4 | 40 | 7.39 a ± 0.05 | 3695 | |
| Tryptophan | ND | 0.01 b ± 0.0 | 0.03 b ± 0.0 | 9.12 a ± 0.2 | 912 | ||
| Antioxidant capacity | |||||||
| DPPH (IC50 for mg mL−1) | 0.77 c ± 0.09 | 0.81 c ± 0.03 | 0.94 a ± 0.18 | 1.15 a ± 0.08 | |||
| ABTS (IC50 mg mL−1) | 0.65 b ± 0.03 | 0.69 b ± 0.12 | 0.717 b ± 0.05 | 0.835 a ± 0.16 | |||
| FRAP (mM VCEAC 100 g−1) | 603.5 a ± 4.3 | 493.54 b 603.5 a ± 1.40 | 482.54 b ± 1.11 | 399.42 c ± 0.88 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seke, F.; Manhivi, V.E.; Shoko, T.; Slabbert, R.M.; Sultanbawa, Y.; Sivakumar, D. Effect of Freeze Drying and Simulated Gastrointestinal Digestion on Phenolic Metabolites and Antioxidant Property of the Natal Plum (Carissa macrocarpa). Foods 2021, 10, 1420. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10061420
Seke F, Manhivi VE, Shoko T, Slabbert RM, Sultanbawa Y, Sivakumar D. Effect of Freeze Drying and Simulated Gastrointestinal Digestion on Phenolic Metabolites and Antioxidant Property of the Natal Plum (Carissa macrocarpa). Foods. 2021; 10(6):1420. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10061420
Chicago/Turabian StyleSeke, Faith, Vimbainashe E. Manhivi, Tinotenda Shoko, Retha M. Slabbert, Yasmina Sultanbawa, and Dharini Sivakumar. 2021. "Effect of Freeze Drying and Simulated Gastrointestinal Digestion on Phenolic Metabolites and Antioxidant Property of the Natal Plum (Carissa macrocarpa)" Foods 10, no. 6: 1420. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10061420