
Occurrence of Polyamines in Foods and the Influence of Cooking Processes

 ,
,  , , and
, , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Polyamine Content in Foods
2.2. Effect of Different Cooking Processes on the Food Polyamine Content
2.3. Statistical Analysis
3. Results and Discussion
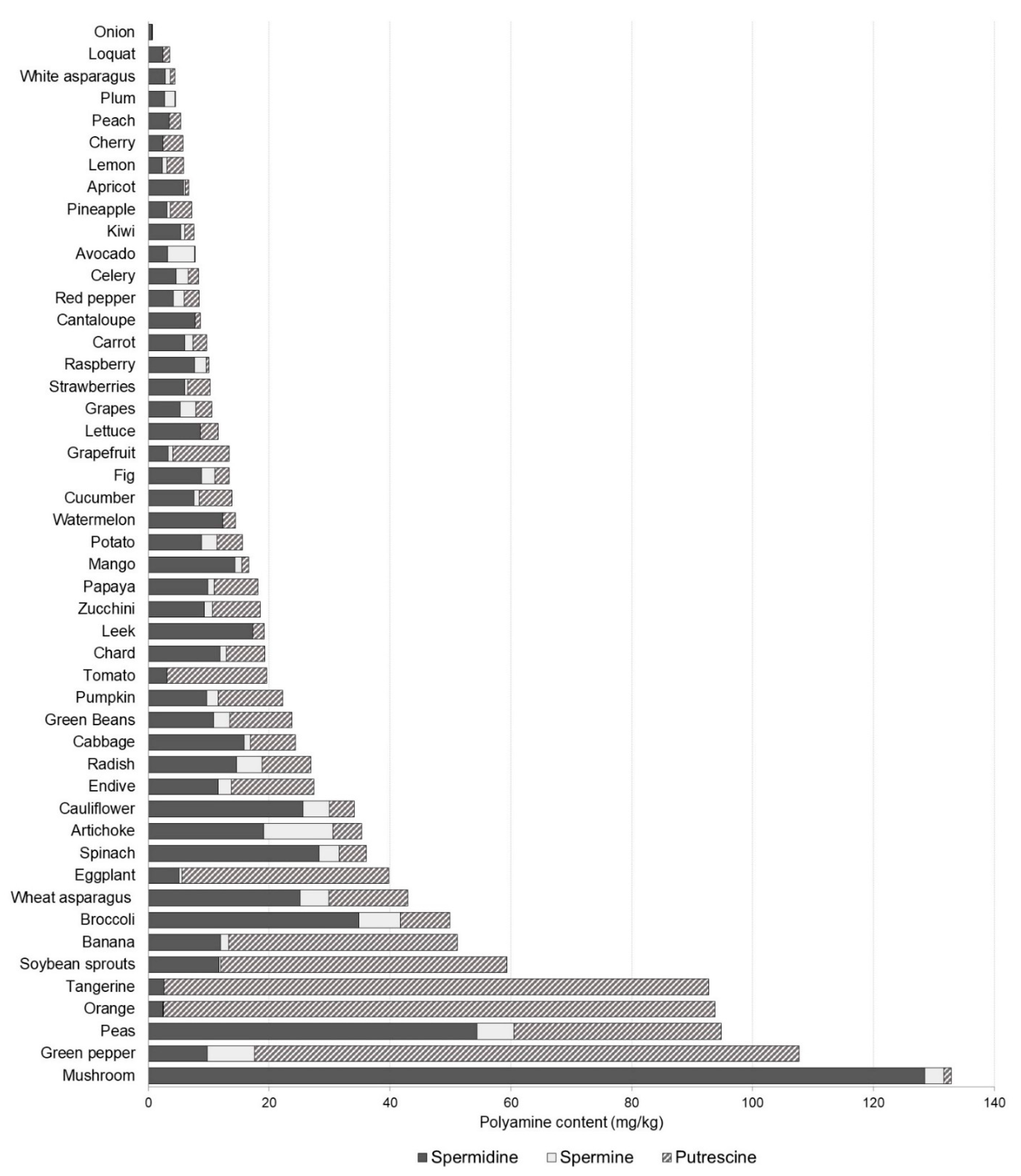
3.1. Polyamine Content in Foods
3.2. Effect of Different Cooking Processes on Food Polyamine Content
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kalač, P. Health effects and occurrence of dietary polyamines: A review for the period 2005–mid 2013. Food Chem. 2014, 161, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Toro-Funes, N.; Bosch-Fusté, J.; Veciana-Nogués, M.T.; Izquierdo-Pulido, M.; Vidal-Carou, M.C. In vitro antioxidant activity of dietary polyamines. Food Res. Int. 2013, 51, 141–147. [Google Scholar] [CrossRef]
- Gómez-Gallego, C.; Kumar, H.; García-Mantrana, I.; du Toit, E.; Suomela, J.-P.; Linderborg, K.M.; Zhang, Y.; Isolauri, E.; Yang, B.; Salminen, S.; et al. Breast Milk Polyamines and Microbiota Interactions: Impact of Mode of Delivery and Geographical Location. Ann. Nutr. Metab. 2017, 70, 184–190. [Google Scholar] [CrossRef] [PubMed]
- Handa, A.K.; Fatima, T.; Mattoo, A.K. Polyamines: Bio-Molecules with Diverse Functions in Plant and Human Health and Disease. Front. Chem. 2018, 6, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Madeo, F.; Eisenberg, T.; Pietrocola, F.; Kroemer, G. Spermidine in health and disease. Science 2018, 359, eaan2788. [Google Scholar] [CrossRef] [Green Version]
- Gugliucci, A.; Menini, T. The polyamines spermine and spermidine protect proteins from structural and functional damage by AGE precursors: A new role for old molecules? Life Sci. 2003, 72, 2603–2616. [Google Scholar] [CrossRef]
- Soda, K.; Dobashi, Y.; Kano, Y.; Tsujinaka, S.; Konishi, F. Polyamine-rich food decreases age-associated pathology and mortality in aged mice. Exp. Gerontol. 2009, 44, 727–732. [Google Scholar] [CrossRef]
- Soda, K. Spermine and gene methylation: A mechanism of lifespan extension induced by polyamine-rich diet. Amino Acids 2019, 52, 213–224. [Google Scholar] [CrossRef]
- Kiechl, S.; Pechlaner, R.; Willeit, P.; Notdurfter, M.; Paulweber, B.; Willeit, K.; Werner, P.; Ruckenstuhl, C.; Iglseder, B.; Weger, S.; et al. Higher spermidine intake is linked to lower mortality: A prospective population-based study. Am. J. Clin. Nutr. 2018, 108, 371–380. [Google Scholar] [CrossRef]
- Soda, K.; Uemura, T.; Sanayama, H.; Igarashi, K.; Fukui, T. Polyamine-Rich Diet Elevates Blood Spermine Levels and Inhibits Pro-Inflammatory Status: An Interventional Study. Med. Sci. 2021, 9, 22. [Google Scholar] [CrossRef]
- Nishimura, K.; Shiina, R.; Kashiwagi, K.; Igarashi, K. Decrease in polyamines with aging and their ingestion from food and drink. J. Biochem. 2006, 139, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Larqué, E.; Sabater-Molina, M.; Zamora, S. Biological significance of dietary polyamines. Nutrition 2007, 23, 87–95. [Google Scholar] [CrossRef] [PubMed]
- Gómez Gallego, C.; Ros Berruezo, G.; Bernal Cava, M.J.; Pérez Conesa, D.; Periago Castón, M.J. Papel de las poliaminas en la alimentación. Importancia de las poliaminas en la alimentación infantil. Arch. Latinoam. Nutr. 2008, 58, 117–125. [Google Scholar] [PubMed]
- Miller-Fleming, L.; Olin-Sandoval, V.; Campbell, K.; Ralser, M. Remaining Mysteries of Molecular Biology: The Role of Polyamines in the Cell. J. Mol. Biol. 2015, 427, 3389–3406. [Google Scholar] [CrossRef]
- Muñoz-Esparza, N.C.; Latorre-Moratalla, M.L.; Comas-Basté, O.; Toro-Funes, N.; Veciana-Nogués, M.T.; Vidal-Carou, M.C. Polyamines in food. Front. Nutr. 2019, 6, 108. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Molina, B.; Queipo-Ortuño, M.I.; Lambertos, A.; Tinahones, F.J.; Peñafiel, R. Dietary and Gut Microbiota Polyamines in Obesity- and Age-Related Diseases. Front. Nutr. 2019, 6, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Tofalo, R.; Cocchi, S.; Suzzi, G. Polyamines and Gut Microbiota. Front. Nutr. 2019, 6, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Kozová, M.; Kalač, P.; Pelikánová, T. Contents of biologically active polyamines in chicken meat, liver, heart and skin after slaughter and their changes during meat storage and cooking. Food Chem. 2009, 116, 419–425. [Google Scholar] [CrossRef]
- Bover-Cid, S.; Latorre-Moratalla, M.L.; Veciana-Nogués, M.T.; Vidal-Carou, M.C. Processing Contaminants: Biogenic Amines. Encycl. Food Saf. 2014, 2, 381–391. [Google Scholar] [CrossRef]
- Sánchez-Pérez, S.; Comas-Basté, O.; Rabell-González, J.; Veciana-Nogués, M.; Latorre-Moratalla, M.; Vidal-Carou, M. Biogenic Amines in Plant-Origin Foods: Are They Frequently Underestimated in Low-Histamine Diets? Foods 2018, 7, 205. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.A.; Poortvliet, E.; Strömberg, R.; Yngve, A. Polyamines: Total daily intake in adolescents compared to the intake estimated from the Swedish Nutrition Recommendations Objectified (SNO). Food Nutr. Res. 2011, 55, 5455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eliassen, K.A.; Reistad, R.; Risøen, U.; Rønning, H.F. Dietary polyamines. Food Chem. 2002, 78, 273–280. [Google Scholar] [CrossRef]
- Krausová, P.; Kalač, P.; Křížek, M.; Pelikánová, T. Changes in the content of biologically active polyamines during storage and cooking of pig liver. Meat Sci. 2007, 77, 269–274. [Google Scholar] [CrossRef] [PubMed]
- Kozová, M.; Kalač, P.; Pelikánová, T. Changes in the content of biologically active polyamines during beef loin storage and cooking. Meat Sci. 2009, 81, 607–611. [Google Scholar] [CrossRef] [PubMed]
- Sagara, T.; Fiechter, G.; Pachner, M.; Mayer, H.K.; Vollmann, J. Soybean spermidine concentration: Genetic and environmental variation of a potential ‘anti-aging’ constituent. J. Food Compos. Anal. 2017, 56, 11–17. [Google Scholar] [CrossRef]
- Latorre-Moratalla, M.L.; Bosch-Fusté, J.; Lavizzari, T.; Bover-Cid, S.; Veciana-Nogués, M.T.; Vidal-Carou, M.C. Validation of an ultra high pressure liquid chromatographic method for the determination of biologically active amines in food. J. Chromatogr. A 2009, 1216, 7715–7720. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 17th ed.; Chemist, I., Ed.; AOAC: Washington, DC, USA, 2000. [Google Scholar]
- Nishibori, N.; Fujihara, S.; Akatuki, T. Amounts of polyamines in foods in Japan and intake by Japanese. Food Chem. 2007, 100, 491–497. [Google Scholar] [CrossRef]
- Lavizzari, T.; Veciana-Nogués, M.T.; Weingart, O.; Bover-Cid, S.; Mariné-Font, A.; Vidal-Carou, M.C. Occurrence of Biogenic Amines and Polyamines in Spinach and Changes during Storage under Refrigeration. J. Agric. Food Chem. 2007, 55, 9514–9519. [Google Scholar] [CrossRef]
- Kusano, T.; Yamaguchi, K.; Berberich, T.; Takahashi, Y. Advances in polyamine research in 2007. J. Plant Res. 2007, 120, 345–350. [Google Scholar] [CrossRef]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine Function in Plants: Metabolism, Regulation on Development, and Roles in Abiotic Stress Responses. Front. Plant Sci. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Glória, M.B.A.; Tavares-Neto, J.; Labanca, R.A.; Carvalho, M.S. Influence of cultivar and germination on bioactive amines in soybeans (Glycine max L. Merril). J. Agric. Food Chem. 2005, 53, 7480–7485. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.-J.; Dong, J.-F.; Zhu, M.-Y. Effects of germination conditions on ascorbic acid level and yield of soybean sprouts. J. Sci. Food Agric. 2005, 85, 943–947. [Google Scholar] [CrossRef]
- Ponce De León, C.; Torija, E.; Matallana, M.C. Utilidad en la alimentación de algunas semillas germinadas: Brotes de soja y trigo. Bol. R. Soc. Esp. Hist. Nat. Sec. Biol. 2013, 107, 47–55. [Google Scholar]
- Menéndez, A.B.; Calzadilla, P.I.; Sansberro, P.A.; Espasandin, F.D.; Gazquez, A.; Bordenave, C.D.; Maiale, S.J.; Rodríguez, A.A.; Maguire, V.G.; Campestre, M.P.; et al. Polyamines and Legumes: Joint Stories of Stress, Nitrogen Fixation and Environment. Front. Plant Sci. 2019, 10, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zoumas-Morse, C.; Rock, C.L.; Quintana, E.L.; Neuhouser, M.L.; Gerner, E.W.; Meyskens, F.L. Development of a Polyamine Database for Assessing Dietary Intake. J. Am. Diet. Assoc. 2007, 107, 1024–1027. [Google Scholar] [CrossRef] [Green Version]
- Bandeira, C.M.; Evangelista, W.P.; Gloria, M.B.A. Bioactive amines in fresh, canned and dried sweet corn, embryo and endosperm and germinated corn. Food Chem. 2012, 131, 1355–1359. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Tan, Y.; Kan, J. Regulation of quality and biogenic amine production during sufu fermentation by pure Mucor strains. LWT 2020, 117, 108637. [Google Scholar] [CrossRef]
- Park, Y.K.; Lee, J.H.; Mah, J.-H. Occurrence and reduction of biogenic amines in traditional Asian fermented soybean foods: A review. Food Chem. 2019, 278, 1–9. [Google Scholar] [CrossRef]
- Lan, G.; Li, C.; He, L.; Zeng, X.; Zhu, Q. Effects of different strains and fermentation method on nattokinase activity, biogenic amines, and sensory characteristics of natto. J. Food Sci. Technol. 2020, 57, 4414–4423. [Google Scholar] [CrossRef]
- Kobayashi, K.; Horii, Y.; Watanabe, S.; Kubo, Y.; Koguchi, K.; Hoshi, Y.; Matsumoto, K.I.; Soda, K. Comparison of soybean cultivars for enhancement of the polyamine contents in the fermented soybean natto using Bacillus subtilis (natto). Biosci. Biotechnol. Biochem. 2017, 81, 587–594. [Google Scholar] [CrossRef] [Green Version]
- Dawood, M.F.A.; Abeed, A.H.A. Spermine-priming restrained water relations and biochemical deteriorations prompted by water deficit on two soybean cultivars. Heliyon 2020, 6, e04038. [Google Scholar] [CrossRef]
- Gull, A.; Ahmad Lone, A.; Ul Islam Wani, N. Biotic and Abiotic Stresses in Plants. In Abiotic and Biotic Stress in Plants; IntechOpen: London, UK, 2019; pp. 3–6. [Google Scholar]
- Khajuria, A.; Sharma, N.; Bhardwaj, R.; Ohri, P. Emerging Role of Polyamines in Plant Stress Tolerance. Curr. Protein Pept. Sci. 2018, 19, 1114–1123. [Google Scholar] [CrossRef] [PubMed]
- Latorre-Moratalla, M.L.; Comas-Basté, O.; Bover-Cid, S.; Vidal-Carou, M.C. Tyramine and histamine risk assessment related to consumption of dry fermented sausages by the Spanish population. Food Chem. Toxicol. 2017, 99, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Paulsen, P.; Hagen, U.; Bauer, F. Changes in biogenic amine contents, non-protein nitrogen and crude protein during curing and thermal processing of M. longissimus, pars lumborum of pork. Eur. Food Res. Technol. 2006, 223, 603–608. [Google Scholar] [CrossRef]
- Krausová, P.; Kalač, P.; Křížek, M.; Pelikánová, T. Changes in the content of biologically active polyamines during pork loin storage and culinary treatments. Eur. Food Res. Technol. 2008, 226, 1007–1012. [Google Scholar] [CrossRef]
- Veciana-Nogués, M.T.; Latorre-Moratalla, M.L.; Toro-Funes, N.; Bosh-Fusté, J.; Vidal-Carou, M.C. Efecto de la cocción con y sin sal en el contenido de poliaminas de las verduras. Nutr. Hosp. 2014, 30, 53. [Google Scholar]
- Ziegler, W.; Hahn, M.; Wallnöfer, P.R. Influence of food processing on the decrease of biogenic amines in various vegetables. Dtsch. Leb. 1994, 90, 108–112. [Google Scholar]
- Bardocz, S.; Grant, G.; MacDonald, A.; Walker, T.J.; Pustzai, A.; Coutts, J. Biogenically active amines in food. In Biogenically Active Amines in Food; Morgan, D.M.L., Milovic, V., Krizek, M., White, A., Eds.; European Comission: Brussels, Belgium, 2001; pp. 119–122. ISBN 92-828-7008-1. [Google Scholar]
- Zamora, R.; Hidalgo, F.J. The Maillard reaction and lipid oxidation. Lipid Technol. 2011, 23, 59–62. [Google Scholar] [CrossRef]
- Jiang, W.; He, X.; Yang, H.; Xiang, X.; Hu, S.; Li, S.; Liu, Y. Histamine reduction by Maillard reaction with glucose. Food Control. 2017, 82, 136–144. [Google Scholar] [CrossRef]
- Latorre-Moratalla, M.; Comas-Basté, L.; Veciana-Nogués, O.; Teresa, M.; Vidal-Carou, M.C. La cocción reduce el contenido de histamina de las espinacas. In 11ª Reunón anual de la Sociedad Española de Seguridad Alimentaria; Sociedad Española de Seguridad Alimentaria: Pamplona, Spain, 2015. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cooking Method | Conditions | Cooking Time |
|---|---|---|
| Boiling | Sample was placed in 500 mL of boiling water. | 15 min |
| Grilling | Sample was placed in the pan once it was hot (140 °C–160 °C). | 7 min |
| Microwave | Sample was placed in a special microwave device made of silicone and cooked at 600 watts. | 5 min |
| Sous-vide | Sample was vacuumed and sealed in a special plastic bag of polyamide and polyethylene with a thickness of 90 microns. It was subsequently immersed in water at 70 °C. | 25 min |
| Food | Raw | Boiling | Grilling | Microwave | Sous-Vide | |
|---|---|---|---|---|---|---|
| Mushroom | SD | 1658.9 ± 7.0 | * 1274.3 ± 19.1 | 1495.5 ± 18.4 | 1646.2 ± 30.4 | 1658.0 ± 31.9 |
| SM | 39.6 ± 0.2 | 39.6 ± 2.2 | 35.9 ± 1.7 | 38.9 ± 2.3 | 39.1 ± 0.6 | |
| PU | 7.0 ± 0.1 | 6.9 ± 0.1 | 6.5 ± 0.2 | 6.7 ± 0.1 | 6.3 ± 0.4 | |
| Asparagus | SD | 645.8 ± 40.8 | 645.7 ± 13.0 | * 484.9 ± 20.2 | 641.8 ± 44.1 | 638.2 ± 4.9 |
| SM | 125.8 ± 0.3 | 125.4 ± 0.1 | * 91.0 ± 2.5 | 117.2 ± 8.1 | 122.8 ± 7.6 | |
| PU | 160.1 ± 3.5 | * 121.0 ± 10.1 | 145.0 ± 7.4 | 143.9 ± 2.5 | 159.3 ± 1.2 | |
| Green beans | SD | 185.6 ± 0.3 | * 157.6 ± 0.1 | * 156.1 ± 2.7 | 179.0 ± 0.4 | 174.1 ± 5.6 |
| SM | 88.9 ± 2.0 | * 73.3 ± 1.4 | * 67.7 ± 1.1 | 80.0 ± 1.9 | 89.5 ± 2.2 | |
| PU | 12.9 ± 0.4 | 12.6 ± 0.4 | 12.5 ± 0.3 | 12.5 ± 0.2 | 12.4 ± 0.1 | |
| Cauliflower | SD | 327.1 ± 9.0 | 325.8 ± 5.3 | 324.4 ± 8.3 | 320.3 ± 23.8 | 324.0 ± 35.9 |
| SM | 66.6 ± 1.7 | 62.7 ± 2.1 | 65.5 ± 3.1 | 63.4 ± 1.5 | 64.4 ± 3.0 | |
| PU | 49.7 ± 0.6 | 48.4 ± 0.3 | 48.2 ± 2.1 | 47.2 ± 4.8 | 45.7 ± 4.3 | |
| Cabbage | SD | 105.1 ± 1.8 | 105.3 ± 0.2 | * 69.6 ± 0.2 | 96.1 ± 3.4 | 105.3 ± 0.3 |
| SM | 36.4 ± 0.3 | 33.7 ± 7.2 | * 15.9 ± 0.2 | 33.0 ± 0.3 | 35.9 ± 0.2 | |
| PU | 2.9 ± 0.1 | 2.37 ± 0.2 | * 1.7 ± 0.1 | 2.8 ± 0.1 | 2.7 ± 0.1 | |
| Broccoli | SD | 389.1 ± 23.9 | 382.6 ± 29.5 | 396.0 ± 17.4 | 389.9 ± 21.6 | 394.9 ± 4.0 |
| SM | 77.2 ± 6.6 | 69.8 ± 4.0 | 73.8 ± 0.1 | 71.2 ± 0.7 | 72.1 ± 1.7 | |
| PU | 91.7 ± 5.6 | 91.7 ± 6.5 | 89.5 ± 4.0 | 85.9 ± 5.4 | 88.5 ± 0.9 | |
| Zucchini | SD | 344.2 ± 3.4 | * 298.0 ± 2.0 | * 282.0 ± 1.7 | * 301.7 ± 1.8 | 341.7 ± 12.0 |
| SM | 62.8 ± 3.9 | * 27.4 ± 4.9 | * 41.6 ± 2.7 | 59.3 ± 1.4 | 51.0 ± 1.9 | |
| PU | 160.7 ± 5.6 | * 133.1 ± 6.61 | * 116.7 ± 1.1 | * 123.2 ± 2.6 | 163.5 ± 12.9 | |
| Spinach | SD | 330.2 ± 0.3 | * 238.5 ± 20.89 | * 236.1 ± 17.0 | * 278.6 ± 1.1 | 317 ± 4.9 |
| SM | 42.2 ± 2.5 | * 31.5 ± 1.9 | * 25.9 ± 1.0 | 38.8 ± 1.6 | 39.0 ± 0.5 | |
| PU | 40.8 ± 0.6 | * 22.3 ± 4.7 | * 31.4 ± 2.3 | 40.2 ± 1.7 | 38.1 ± 0.3 | |
| Pumpkin | SD | 80.3 ± 0.6 | * 66.6 ± 4.2 | * 60.4 ± 1.1 | 78.1 ± 0.5 | 76.7 ± 0.6 |
| SM | 840.3 ± 8.4 | 834.0 ± 10.4 | * 664.8 ± 5.4 | 839.5 ± 0.8 | 833.2 ± 2.8 | |
| PU | 19.3 ± 0.5 | * 15.1 ± 1.0 | 17.4 ± 2.5 | 19.8 ± 0.6 | 19.5 ± 0.1 | |
| Chard | SD | 239.7 ± 11.4 | * 178.0 ± 0.3 | 234.7 ± 2.7 | * 196.6 ± 1.0 | 215.3 ± 14.5 |
| SM | 28.4 ± 1.8 | 27.1 ± 1.0 | 26.0 ± 0.2 | 25.9 ± 0.6 | 28.1 ± 0.5 | |
| PU | 38.6 ± 1.7 | * 13.7 ± 0.2 | * 26.1 ± 0.6 | 35.1 ± 0.3 | 32.8 ± 0.7 | |
| Chicken meat | SD | 1.5 ± 0.1 | 1.0 ± 0.1 | 1.4 ± 0.2 | 1.11 ± 0.0 | 1.2 ± 0.1 |
| SM | 10.0 ± 0.9 | 8.2 ± 0.4 | 8.5 ± 0.0 | 8.4 ± 0.3 | 9.9 ± 0.4 | |
| PU | 0.4 ± 0.0 | 0.3 ± 0.0 | 0.3 ± 0.0 | 0.3 ± 0.1 | 0.4 ± 0.1 | |
| Beef meat | SD | 9.3 ± 0.4 | 8.1 ± 0.1 | * 7.5 ± 0.1 | * 7.6 ± 0.5 | 8.6 ± 0.1 |
| SM | 130.0 ± 9.5 | * 94.4 ± 3.4 | 115.7 ± 0.4 | 120.8 ± 0.9 | 127.2 ± 9.5 | |
| PU | 10.4 ± 0.6 | * 5.9 ± 0.1 | * 7.1 ± 0.1 | * 7.7 ± 0.3 | 9.4 ± 0.6 | |
| Pork meat | SD | 9.4 ± 0.5 | * 5.9 ± 0.1 | * 7.7 ± 0.2 | 8.8 ± 0.3 | 9.0 ± 0.2 |
| SM | 138.1 ± 1.0 | * 89.0 ± 0.3 | * 118.6 ± 3.4 | 137.6 ± 0.1 | 136.9 ± 0.8 | |
| PU | 0.4 ± 0.0 | 0.3 ± 0.1 | 0.4 ± 0.0 | ± 0.0 | 0.4 ± 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muñoz-Esparza, N.C.; Costa-Catala, J.; Comas-Basté, O.; Toro-Funes, N.; Latorre-Moratalla, M.L.; Veciana-Nogués, M.T.; Vidal-Carou, M.C. Occurrence of Polyamines in Foods and the Influence of Cooking Processes. Foods 2021, 10, 1752. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10081752
Muñoz-Esparza NC, Costa-Catala J, Comas-Basté O, Toro-Funes N, Latorre-Moratalla ML, Veciana-Nogués MT, Vidal-Carou MC. Occurrence of Polyamines in Foods and the Influence of Cooking Processes. Foods. 2021; 10(8):1752. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10081752
Chicago/Turabian StyleMuñoz-Esparza, Nelly C., Judit Costa-Catala, Oriol Comas-Basté, Natalia Toro-Funes, M. Luz Latorre-Moratalla, M. Teresa Veciana-Nogués, and M. Carmen Vidal-Carou. 2021. "Occurrence of Polyamines in Foods and the Influence of Cooking Processes" Foods 10, no. 8: 1752. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10081752