
Anti-Obesity Effects of Morus alba L. and Aronia melanocarpa in a High-Fat Diet-Induced Obese C57BL/6J Mouse Model

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
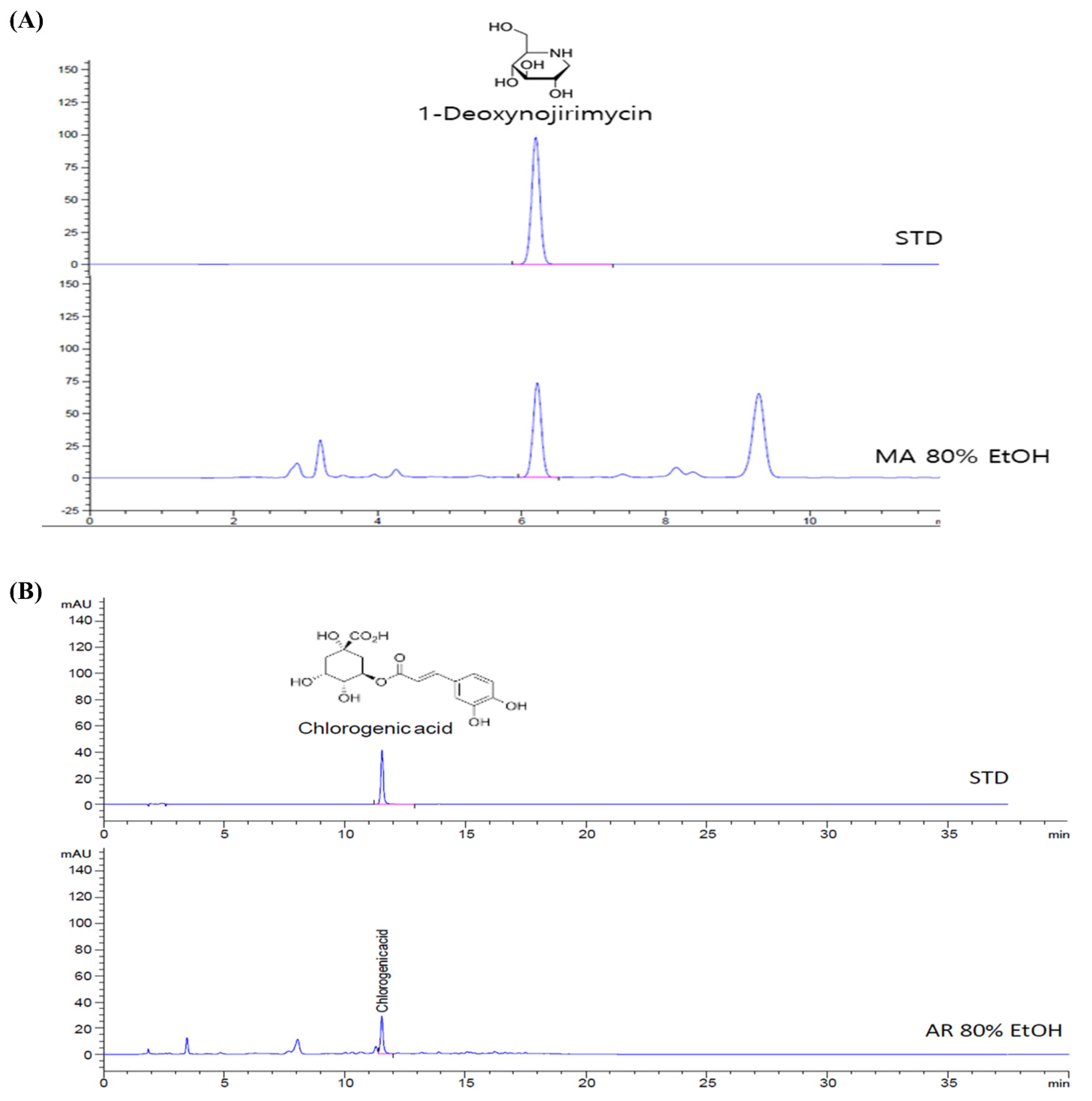
2.1. Preparation and HPLC Analysis of MA and AR
2.2. Animal Experimental Protocol
2.3. Analysis of Serum Biochemical Parameters
2.4. Micro-Computed Tomography (CT)
2.5. Histological Analysis
2.6. Gene Expression Analysis
2.7. Western Blot Analysis
2.8. Statistical Analysis
3. Results
3.1. Compounds Present in MA and AR
3.2. Effect of MA and AR Extracts on Body Weight, Tissue Weight, and Feed Intake
3.3. Effect of MA and AR Treatment on Serum Lipid Profiles and Adipokine Levels
3.4. Effect of MA and AR on Hepatic Lipids
3.5. Changes in Visceral Fat Volume and Histopathology after MA and AR Treatment
3.6. mRNA Expression of Genes in Liver and Adipose Tissue
3.7. Western Blot Analysis of Liver Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Obesity and Overweight. 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/obesityand-overweight (accessed on 17 August 2021).
- Elagizi, A.; Kachur, S.; Lavie, C.J.; Carbone, S.; Pandey, A.; Ortega, F.B.; Milani, R.V. An Overview and Update on Obesity and the Obesity Paradox in Cardiovascular Diseases. Prog. Cardiovasc. Dis. 2018, 61, 142–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryder, J.R.; Fox, C.K.; Kelly, A.S. Treatment Options for Severe Obesity in the Pediatric Population: Current Limitations and Future Opportunities. Obesity 2018, 26, 951–960. [Google Scholar] [CrossRef] [Green Version]
- Kellogg, J.; Wang, J.; Flint, C.; Ribnicky, D.; Kuhn, P.; de Mejia, E.G.; Raskin, I.; Lila, M.A. Alaskan Wild Berry Resources and Human Health under the Cloud of Climate Change. J. Agric. Food Chem. 2009, 58, 3884–3900. [Google Scholar] [CrossRef] [Green Version]
- Kowalska, K.; Olejnik, A.; Szwajgier, D.; Olkowicz, M. Inhibitory activity of chokeberry, bilberry, raspberry and cranberry polyphenol-rich extract towards adipogenesis and oxidative stress in differentiated 3T3-L1 adipose cells. PLoS ONE 2017, 12, e0188583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ercisli, S.; Orhan, E. Chemical composition of white (Morus alba), red (Morus rubra) and black (Morus nigra) mulberry fruits. Food Chem. 2007, 103, 1380–1384. [Google Scholar] [CrossRef]
- Mahboubi, M. Morus alba (mulberry), a natural potent compound in management of obesity. Pharmacol. Res. 2019, 146, 104341. [Google Scholar] [CrossRef]
- Tian, S.; Wang, M.; Liu, C.; Zhao, H.; Zhao, B. Mulberry leaf reduces inflammation and insulin resistance in type 2 diabetic mice by TLRs and insulin Signalling pathway. BMC Complement. Altern. Med. 2019, 19, 326. [Google Scholar] [CrossRef] [Green Version]
- Li, H.X.; Jo, E.; Myung, C.-S.; Kim, Y.H.; Yang, S.Y. Lipolytic effect of compounds isolated from leaves of mulberry (Morus alba L.) in 3T3-L1 adipocytes. Nat. Prod. Res. 2017, 32, 1963–1966. [Google Scholar] [CrossRef]
- Chang, Y.-C.; Yang, M.-Y.; Chen, S.-C.; Wang, C.-J. Mulberry leaf polyphenol extract improves obesity by inducing adipocyte apoptosis and inhibiting preadipocyte differentiation and hepatic lipogenesis. J. Funct. Foods 2015, 21, 249–262. [Google Scholar] [CrossRef]
- Daskalova, E.; Delchev, S.; Topolov, M.; Dimitrova, S.; Uzunova, Y.; Valcheva-Kuzmanova, S.; Kratchanova, M.; Vladimirova-Kitova, L.; Denev, P. Aronia melanocarpa (Michx.) Elliot fruit juice reveals neuroprotective effect and improves cognitive and locomotor functions of aged rats. Food Chem. Toxicol. 2019, 132, 110674. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Gao, J.; Yu, W.; Hao, R.; Fan, J.; Wei, J. The effects and mechanism of Aronia melanocarpa Elliot anthocyanins on hepatic fibrosis. J. Funct. Foods 2020, 68, 103897. [Google Scholar] [CrossRef]
- Borissova, P.; Valcheva, S.; Belcheva, A. Antiinflammatory effect of flavonoids in the natural juice from Aronia melanocarpa, rutin and rutin-magnesium complex on an experimental model of inflammation induced by histamine and serotonin. Acta Physiol. Pharmacol. Bulg. 1994, 20, 25–30. [Google Scholar]
- Valcheva-Kuzmanova, S.; Kuzmanov, K.; Tancheva, S.; Belcheva, A. Hypoglycemic and hypolipidemic effects of Aronia melanocarpa fruit juice in streptozotocin-induced diabetic rats. Methods Find. Exp. Clin. Pharmacol. 2007, 29, 101–106. [Google Scholar] [CrossRef]
- Cebova, M.; Klimentova, J.; Janega, P.; Pechanova, O. Effect of bioactive compound of Aronia melanocarpa on cardiovascular system in experimental hypertension. Oxid. Med. Cell. Longev. 2017, 2017, 8156594. [Google Scholar] [CrossRef] [Green Version]
- Jurikova, T.; Mlcek, J.; Skrovankova, S.; Sumczynski, D.; Sochor, J.; Hlavacova, I.; Snopek, L.; Orsavová, J. Fruits of Black Chokeberry Aronia melanocarpa in the Prevention of Chronic Diseases. Molecules 2017, 22, 944. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhang, J.-Y.; Wei, Y.-L.; Hao, J.-Y.; Lei, Y.-Q.; Zhao, W.-B.; Xiao, Y.-H.; Sun, A.-D. The polyphenol-rich extract from chokeberry (Aronia melanocarpa L.) modulates gut microbiota and improves lipid metabolism in diet-induced obese rats. Nutr. Metab. 2020, 17, 54. [Google Scholar] [CrossRef]
- Lim, S.-M.; Lee, H.S.; Jung, J.I.; Kim, S.M.; Kim, N.Y.; Seo, T.S.; Bae, J.-S.; Kim, E.J. Cyanidin-3-O-Galactoside-Enriched Aronia melanocarpa Extract Attenuates Weight Gain and Adipogenic Pathways in High-Fat Diet-Induced Obese C57BL/6 Mice. Nutrients 2019, 11, 1190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.I.; Lee, D.H.; Park, S.H.; Jang, Y.J.; Ahn, J.; Ha, T.Y.; Jung, C.H. Antiobesity effects of the combination of Patrinia scabiosaefolia root and Hippophae rhamnoides leaf extracts. J. Food Biochem. 2020, 44, e13214. [Google Scholar] [CrossRef] [PubMed]
- Leng, E.; Xiao, Y.; Mo, Z.; Li, Y.; Zhang, Y.; Deng, X.; Zhou, M.; Zhou, C.; He, Z.; He, J.; et al. Synergistic effect of phytochemicals on cholesterol metabolism and lipid accumulation in HepG2 cells. BMC Complement. Altern. Med. 2018, 18, 122. [Google Scholar] [CrossRef]
- Thomas, S.S.; Kim, M.; Lee, S.J.; Cha, Y.S. Antiobesity Effects of Purple Perilla (Perilla frutescens var. acuta) on Adipocyte Differentiation and Mice Fed a High-fat Diet. J. Food Sci. 2018, 83, 2384–2393. [Google Scholar] [CrossRef] [PubMed]
- Chooi, Y.C.; Ding, C.; Magkos, F. The epidemiology of obesity. Metabolism 2018, 92, 6–10. [Google Scholar] [CrossRef] [Green Version]
- Kopelman, P.G. Obesity as a medical problem. Nature 2000, 404, 635–643. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, M.O.; Horvath, T.L. Limitations in anti-obesity drug development: The critical role of hunger-promoting neurons. Nat. Rev. Drug Discov. 2012, 11, 675–691. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sun, M.; Yao, H.; Liu, Y.; Gao, R. Herbal Medicine for the Treatment of Obesity: An Overview of Scientific Evidence from 2007 to 2017. Evid. Based Complement. Altern. Med. 2017, 2017, 8943059. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Smith, D.L., Jr.; Keating, K.D.; Allison, D.B.; Nagy, T.R. Variations in body weight, food intake and body composition after long-term high-fat diet feeding in C57BL/6J mice. Obesity 2014, 22, 2147–2155. [Google Scholar] [CrossRef] [PubMed]
- Kakimoto, P.A.; Kowaltowski, A.J. Effects of high fat diets on rodent liver bioenergetics and oxidative imbalance. Redox Biol. 2016, 8, 216–225. [Google Scholar] [CrossRef] [Green Version]
- Green, C.J.; Hodson, L. The Influence of Dietary Fat on Liver Fat Accumulation. Nutrients 2014, 6, 5018–5033. [Google Scholar] [CrossRef]
- Carr, M.C.; Brunzell, J.D. Abdominal Obesity and Dyslipidemia in the Metabolic Syndrome: Importance of Type 2 Diabetes and Familial Combined Hyperlipidemia in Coronary Artery Disease Risk. J. Clin. Endocrinol. Metab. 2004, 89, 2601–2607. [Google Scholar] [CrossRef] [Green Version]
- Ann, J.-Y.; Eo, H.; Lim, Y. Mulberry leaves (Morus alba L.) ameliorate obesity-induced hepatic lipogenesis, fibrosis, and oxidative stress in high-fat diet-fed mice. Genes Nutr. 2015, 10, 46. [Google Scholar] [CrossRef] [Green Version]
- Arner, P.; Rydén, M. Fatty acids, obesity and insulin resistance. Obes. Facts 2015, 8, 147–155. [Google Scholar] [CrossRef]
- Bergman, R.N.; Ader, M. Free Fatty Acids and Pathogenesis of Type 2 Diabetes Mellitus. Trends Endocrinol. Metab. 2000, 11, 351–356. [Google Scholar] [CrossRef]
- Izquierdo, A.G.; Crujeiras, A.B.; Casanueva, F.F.; Carreira, M.C. Leptin, obesity, and leptin resistance: Where are we 25 years later? Nutrients 2019, 11, 2704. [Google Scholar] [CrossRef] [Green Version]
- Ricci, R.; Bevilacqua, F. The potential role of leptin and adiponectin in obesity: A comparative review. Vet. J. 2012, 191, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.-H.; Lin, H.-T.; Chung, D.-J.; Huang, C.-N.; Wang, C.-J. Mulberry Leaf Extracts prevent obesity-induced NAFLD with regulating adipocytokines, inflammation and oxidative stress. J. Food Drug Anal. 2018, 26, 778–787. [Google Scholar] [CrossRef]
- Kong, W.-H.; Oh, S.-H.; Ahn, Y.-R.; Kim, K.-W.; Kim, J.-H.; Seo, S.-W. Antiobesity Effects and Improvement of Insulin Sensitivity by 1-Deoxynojirimycin in Animal Models. J. Agric. Food Chem. 2008, 56, 2613–2619. [Google Scholar] [CrossRef] [PubMed]
- Utzschneider, K.M.; Carr, D.B.; Barsness, S.M.; Kahn, S.E.; Schwartz, R.S. Diet-Induced Weight Loss Is Associated with an Improvement in β-Cell Function in Older Men. J. Clin. Endocrinol. Metab. 2004, 89, 2704–2710. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuduki, T.; Kikuchi, I.; Kimura, T.; Nakagawa, K.; Miyazawa, T. Intake of mulberry 1-deoxynojirimycin prevents diet-induced obesity through increases in adiponectin in mice. Food Chem. 2013, 139, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Rosen, E.D.; Sarraf, P.; Troy, A.E.; Bradwin, G.; Moore, K.; Milstone, D.S.; Spiegelman, B.M.; Mortensen, R.M. PPARγ Is Required for the Differentiation of Adipose Tissue In Vivo and In Vitro. Mol. Cell 1999, 4, 611–617. [Google Scholar] [CrossRef]
- Foretz, M.; Guichard, C.; Ferré, P.; Foufelle, F. Sterol regulatory element binding protein-1c is a major mediator of insulin action on the hepatic expression of glucokinase and lipogenesis-related genes. Proc. Natl. Acad. Sci. USA 1999, 96, 12737–12742. [Google Scholar] [CrossRef] [Green Version]
- Fajas, L.; Schoonjans, K.; Gelman, L.; Kim, J.B.; Najib, J.; Martin, G.; Fruchart, J.-C.; Briggs, M.; Spiegelman, B.M.; Auwerx, J. Regulation of Peroxisome Proliferator-Activated Receptor γ Expression by Adipocyte Differentiation and Determination Factor 1/Sterol Regulatory Element Binding Protein 1: Implications for Adipocyte Differentiation and Metabolism. Mol. Cell Biol. 1999, 19, 5495–5503. [Google Scholar] [CrossRef] [Green Version]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. SREBPs: Activators of the complete program of cholesterol and fatty acid synthesis in the liver. J. Clin. Investig. 2002, 109, 1125–1131. [Google Scholar] [CrossRef]
- Kim, J.B.; Sarraf, P.; Wright, M.; Yao, K.M.; Mueller, E.; Solanes, G.; Lowell, B.B.; Spiegelman, B.M. Nutritional and insulin regulation of fatty acid synthetase and leptin gene expression through ADD1/SREBP1. J. Clin. Investig. 1998, 101, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, L.; Haryono, R.; Keast, R. Functionality of fatty acid chemoreception: A potential factor in the development of obesity? Nutrients 2013, 5, 1287–1300. [Google Scholar] [CrossRef]
- Jeong, S.; Yoon, M. Fenofibrate inhibits adipocyte hypertrophy and insulin resistance by activating adipose PPARα in high fat diet-induced obese mice. Exp. Mol. Med. 2009, 41, 397–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, E.; Lim, S.-M.; Kim, M.-S.; Yoo, S.-H.; Kim, Y. Phyllodulcin, a Natural Sweetener, Regulates Obesity-Related Metabolic Changes and Fat Browning-Related Genes of Subcutaneous White Adipose Tissue in High-Fat Diet-Induced Obese Mice. Nutrients 2017, 9, 1049. [Google Scholar] [CrossRef] [PubMed]
- Duval, C.; Müller, M.; Kersten, S. PPARα and dyslipidemia. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2007, 1771, 961–971. [Google Scholar] [CrossRef]
- Fan, C.-Y.; Pan, J.; Chu, R.; Lee, D.; Kluckman, K.D.; Usuda, N.; Singh, I.; Yeldandi, A.V.; Rao, M.S.; Maeda, N.; et al. Hepatocellular and Hepatic Peroxisomal Alterations in Mice with a Disrupted Peroxisomal Fatty Acyl-coenzyme A Oxidase Gene. J. Biol. Chem. 1996, 271, 24698–24710. [Google Scholar] [CrossRef] [Green Version]
- Song, S.; Attia, R.R.; Connaughton, S.; Niesen, M.I.; Ness, G.C.; Elam, M.B.; Hori, R.T.; Cook, G.A.; Park, E.A. Peroxisome proliferator activated receptor α (PPARα) and PPAR gamma coactivator (PGC-1α) induce carnitine palmitoyltransferase IA (CPT-1A) via independent gene elements. Mol. Cell. Endocrinol. 2010, 325, 54–63. [Google Scholar] [CrossRef] [Green Version]
- Lindholm, C.R.; Ertel, R.L.; Bauwens, J.D.; Schmuck, E.G.; Mulligan, J.D.; Saupe, K.W. A high-fat diet decreases AMPK activity in multiple tissues in the absence of hyperglycemia or systemic inflammation in rats. J. Physiol. Biochem. 2012, 69, 165–175. [Google Scholar] [CrossRef]
- Bort, A.; Sánchez, B.G.; Mateos-Gómez, P.A.; Díaz-Laviada, I.; Rodríguez-Henche, N. Capsaicin Targets Lipogenesis in HepG2 Cells Through AMPK Activation, AKT Inhibition and PPARs Regulation. Int. J. Mol. Sci. 2019, 20, 1660. [Google Scholar] [CrossRef] [Green Version]
- Hameed, A.; Galli, M.; Adamska-Patruno, E.; Krętowski, A.; Ciborowski, M. Select Polyphenol-Rich Berry Consumption to Defer or Deter Diabetes and Diabetes-Related Complications. Nutrients 2020, 12, 2538. [Google Scholar] [CrossRef] [PubMed]
- Chan, E.W.-C.; Phui-Yan, L.; Wong, S.-K. Phytochemistry, pharmacology, and clinical trials of Morus alba. Chin. J. Nat. Med. 2016, 14, 17–30. [Google Scholar] [PubMed]
- Hawkins, J.; Hires, C.; Baker, C.; Keenan, L.; Bush, M. Daily supplementation with Aronia melanocarpa (chokeberry) reduces blood pressure and cholesterol: A meta analysis of controlled clinical trials. J. Diet. Suppl. 2020, 1–14. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name 1 | Primers | Sequence (5′→ 3′) |
|---|---|---|
| SREBP-1c | Forward | ACGGAGCCATGGATTGCACA |
| Reverse | AAGGGTGCAGGTGTCACCTT | |
| PPAR-γ | Forward | GCCCACCAACTTCGGAATC |
| Reverse | TGCGAGTGGTCTTCCATCAC | |
| C/EBP-α | Forward | GTGTGCACGTCTATGCTAAACCA |
| Reverse | GCCGTTAGTGAAGAGTCTCAGTTTG | |
| FAS | Forward | GAAGTGTCTGGACTGTGTCATTTTTAC |
| Reverse | TTAATTGTGGGATCAGGAGAGCAT | |
| CD36 | Forward | GCTTGCAACTGTCAGCACAT |
| Reverse | GCCTTGCTGTAGCCAAGAAC | |
| PPAR-α | Forward | AGGCTGTAAGGGCTTCTTTCG |
| Reverse | GGCATTTGTTCCGGTTCTTC | |
| ACOX1 | Forward | GTATAAACTCTTCCCGCTCCTG |
| Reverse | CCAGGTAGTAAAAGCCTTCAGC | |
| CPT-1a | Forward | TGGCATCATCACTGGTGTGTT |
| Reverse | GTCTAGGGTCCGATTGATCTTTG |
| Parameters | ND | HD | MA | AR | MA + AR |
|---|---|---|---|---|---|
| TC (mg/dL) | 111.2 ± 21.2 c | 192.4 ± 11.4 a | 184.7 ± 12.5 a | 178.4 ± 17.9 ab | 163.3 ± 22.2 b |
| TG (mg/dL) | 88.0 ± 13.6 b | 120.3 ± 20.7 a | 102.1 ± 13.5 b | 99.5 ± 10.6 b | 96.1 ± 11.6 b |
| HDL-C (mg/dL) | 47.5 ± 4.7 b | 56.8 ± 4.4 a | 60.5 ± 3.6 a | 59.3 ± 2.2 a | 60.4 ± 6.4 a |
| LDL-C (mg/dL) | 48.8 ± 20.7 c | 112.4 ± 14.6 a | 106.5 ± 10.3 ab | 99.4 ± 15.7 ab | 91.7 ± 13.6 b |
| VLDL (mg/dL) | 17.6 ± 2.7 b | 24.1 ± 4.1 a | 20.4 ± 2.7 b | 20.7 ± 3.1 b | 19.2 ± 2.3 b |
| Free fatty acid (nmol/μL) | 0.90 ± 0.01 ab | 0.99 ± 0.11 a | 0.71 ± 0.14 b | 0.75 ± 0.22 b | 0.74 ± 0.14 b |
| Glycerol (nmol/μL) | 0.22 ± 0.03 b | 0.28 ±0.03 a | 0.24 ± 20.02 b | 0.24 ± 0.03 b | 0.22 ± 0.03 b |
| Adiponectin (ng/mL) | 10.32 ± 0.30 a | 9.54 ±0.38 b | 11.05 ± 0.39 a | 10.51 ± 0.97 a | 10.50 ± 0.88 a |
| Leptin (ng/mL) | 4.15 ± 1.81 c | 75.24 ± 13.84 a | 61.19 ± 15.78 ab | 59.93 ± 9.51 ab | 56.08 ± 13.7 b |
| Insulin (ng/mL) | 0.06 ± 0.03 c | 1.44 ± 0.56 a | 0.66 ± 0.38 b | 0.71 ± 0.46 b | 0.98 ± 0.63 b |
| Parameters | ND | HD | MA | AR | MA + AR |
|---|---|---|---|---|---|
| TC (mg/g tissue) | 17.3 ± 1.6 b | 23.7 ± 4.0 a | 18.8 ± 3.0 b | 21.0 ± 3.6 b | 21.9 ± 3.9 ab |
| TG (mg/g tissue) | 50.3 ± 7.7 b | 66.1 ± 8.0 a | 55.0± 8.7 b | 49.9 ± 10.2 b | 55.9 ± 8.6 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, N.-Y.; Thomas, S.S.; Hwang, D.-I.; Lee, J.-H.; Kim, K.-A.; Cha, Y.-S. Anti-Obesity Effects of Morus alba L. and Aronia melanocarpa in a High-Fat Diet-Induced Obese C57BL/6J Mouse Model. Foods 2021, 10, 1914. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10081914
Kim N-Y, Thomas SS, Hwang D-I, Lee J-H, Kim K-A, Cha Y-S. Anti-Obesity Effects of Morus alba L. and Aronia melanocarpa in a High-Fat Diet-Induced Obese C57BL/6J Mouse Model. Foods. 2021; 10(8):1914. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10081914
Chicago/Turabian StyleKim, Na-Yeon, Shalom Sara Thomas, Dae-Il Hwang, Ji-Hye Lee, Kyung-Ah Kim, and Youn-Soo Cha. 2021. "Anti-Obesity Effects of Morus alba L. and Aronia melanocarpa in a High-Fat Diet-Induced Obese C57BL/6J Mouse Model" Foods 10, no. 8: 1914. https://0-doi-org.brum.beds.ac.uk/10.3390/foods10081914