1. Introduction
Cronobacter sakazakii (
C. sakazakii) is an opportunistic foodborne pathogen isolated from a variety of food sources [
1]. Neonates are the most susceptible to fatal infections (e.g., meningitis and septicaemia), although other immune-compromised groups, such as the elderly, are also vulnerable. The organism is a food safety risk in powdered milk products, especially powdered infant formula (PIF) products, as
C. sakazakii is heat tolerant, resistant to desiccation and can form biofilms, and thus, it may survive in the processing environment for a long time [
2]. Environmental isolates of
C. sakazakii survive via stress response adaptations, making it difficult to be eradicated from a dairy processing environment.
Development of new sanitization technologies is contributing to the improved control of
C. sakazakii. Osaili et al. reported that 2 kGy of gamma radiation could eliminate 1.2 to 1.5 log CFU of desiccation-stressed
C. sakazakii isolates in dry infant milk formula [
3]. Inactivation of stressed
C. sakazakii after starvation, heat, cold, acid, alkaline, chlorine or ethanol treatments required 0.82 to 1.24 kGy of gamma radiation with D
10 value ranging from 1.35 to 1.95 [
4]. Ultraviolet (UV 200–260 nm) treatment at 2160 mJ/cm
2 reduced the bacterial level to the limit of detection (1.18 log CFU per coupon) on silicone, polycarbonate, and stainless steel [
5]. Another study reported that in vitro treatment of
C. sakazakii with conventional UV lamps (254 nm) achieved 1.3log CFU reduction at ambient temperature (25 °C) although higher temperatures (56–57 °C) resulted in higher reductions [
6]. However, the safety risks associated with UVC exposure its application as a germicidal technology.
Pulsed broad white light (Xenon flash lamp 190–1100 nm) has been found to achieve 4 log CFU reductions or above of
C. sakazakii [
7,
8]. However, it is still a UV-containing application. Visible light (especially blue light wavelengths from 405 to 465 nm) has been shown to have antimicrobial properties and inactivates microbes via photodynamic inactivation, where cellular molecules known as photosensitizers produce reactive oxygen species which react with cellular constituents such as lipids, proteins, and DNA to bring about a cytotoxic effect [
9]. Our previous study showed that blue-LED at 405 nm caused a significant reduction (to extinction) in the levels of
Escherichia coli (Shiga-toxin producing
E. coli) in vitro [
10]. With respect to
C. sakazakii, there is only limited evidence on its inactivation by blue-LED (light emitting diode) application. A recent study reported that 405-nm LED inactivated
C. sakazakii in PIF (up to 0.9 log reduction) at a dose of 546 J/cm
2 and decreased the tolerance of
C. sakazakii in PIF to desiccation, heat treatment at 50 and 55 °C, low PH (4.75) of simulated gastric fluid, and bile salt [
11]. Another study showed that about 375 J/cm
2 of 405 nm LED illumination significantly reduced the population of a
C. sakazakii (ATCC 29004) biofilm by 2.0–2.5 log CFU at different temperatures (25, 10, and 4 °C) [
12]. In this study, we investigated the antimicrobial effects of blue-LED at 405 nm against three strains of
C. sakazakii, one reference strain and two isolates from obtained from dairy processing environments. Further, phenotypic characteristics (such as metabolic rate, the outer membrane permeability, and ROS production) when exposed to blue-LED treatment were compared among the tested strains to better understand the antimicrobial mechanism of blue light.
2. Materials and Methods
2.1. Bacterial Strains
Three
C. sakazakii strains, ATCC BAA-894 (isolated from powdered formula associated with a neonatal intensive care unit outbreak), ES191 and AGRFS2961 (dairy environment isolates), were used in the study. ES191 has previously been identified as super biofilm former in different nutrient matrices at 37 °C [
13]. AGRFS2961 is an isolate from bacterial collection of Food System Integrity Team (AgResearch, Palmerston North, New Zealand) and was isolated from a New Zealand dairy processing plant. Single colonies of each strain were statically incubated under aerobic conditions in tryptic soy broth (TSB) or sterile 10% skim milk at 37 °C for 20 h to obtain overnight bacterial culture (~8 log CFU/mL).
2.2. Light Source and Application
Blue light (45 nm) was produced by a prototype LED array, a surface-mounted diode panel consisting of 400 LED bulbs arranged in strips (450 × 450 mm). The cell suspension or cell inoculated coupons were placed vertically at a distance of 25 cm from the light source, which corresponded to average light intensities of 50 mW/cm
2. Light intensity was measured as described in Wu et al. [
10]. The light treatments were all conducted in a temperature-controlled chamber (SKOPE PG600 VC, NZ). The average running temperature and humidity of the lighting surface were recorded per second using customized sensors and software (AgResearch Engineering Development Centre, Lincoln, New Zealand) during light exposure. Surface temperature under illumination was maintained at 28 ± 2 °C; surface humidity was between 20% and 30%.
2.3. Cytotoxicity/Antimicrobial Effect of LED against Stationary Phase Cells
The cytotoxicity of blue-LED light was measured using the XTT (sodium 3′-[1-[(phenylamino)-carbony]-3,4-tetrazolium]-bis(4-methoxy-6-nitro)benzene-sulfonic acid hydrate) assay against, tested cells over a 4 h time course [
14]. The antimicrobial effect of blue LED illumination was determined by the enumeration of viable bacterial cells on agar plates. Approximately 8 log CFU/mL overnight culture cells in TSB were resuspended in phosphate buffered saline (PBS) and 1.5 mL aliquots for each strain were introduced into 5 wells of a 24 well plate (Nunc™). Subsequently, samples in the 24-well plate were exposed to blue light. As a control, another 24-well plate was tested under the same condition but without light illumination. For both treated and control plates, the cell suspensions were sampled at 0, 1, 2, 3, and 4 h during the test.
The sampled cells were centrifuged to remove the supernatant and each cell pellet was resuspended in 1.5 mL PBS suspension. For the XTT assay, 500 µL of each sample was taken and mixed with 200 µL of 1 mg/mL XTT solution containing 25 nM PMS (phenazine methosulfate, MilliporeSigma, Burlington, MA, USA) for 3-h dark incubation at 37 °C. The change of colorimetric activity was quantitively measured by optical density at 450 nm with a spectrophotometer (MultiScan Go, Thermo Scientific, Waltham, MA, USA) as the maximum absorbance of the reduced XTT is around 450 nm. A reference reading on cell-free XTT solution with PMS was conducted for correction. Meanwhile, the number of viable cells was enumerated by plate counting (10-fold serial dilution for plating on TSB agar for 24 h incubation at 37 °C) using 1 mL of the cell suspension for each sample.
2.4. Change of the Outer Membrane Permeability When Exposed to Antimicrobial Light
Aliquots of 1 mL were taken from a PBS suspension containing stationary phase cells (7 log CFU/mL) and added into 24-well plates (NuncTM). For each strain, one plate received blue-LED treatment and the other was a control plate held under the same temperature and humidity condition but without any light exposure. Cells were sampled at time 0 min, 15 min, 30 min, 45 min, 1 h, 2 h, 3 h and 4 h. Sampled cells (1 mL) were centrifuged at 10,000× g for 3 min and resuspended in 500 µL of 5 mmol/L HEPE [N-(2-Hydroxyethyl) piperazine-N′-(2-ethanesulfonic acid] buffer. One hundred μL of bacterial suspension was mixed with 100 μL of 40 μmol/L NPN (1-N-phenylnapthylamine, Sigma, Burlington, MA, USA) in 96 well plates (Nunc™) for fluorescence measurement at an excitation wavelength of 350 nm and emission wavelength of 420 nm (Varioskan™ LUX, Thermo Scientific, Waltham, MA, USA) while the HEPE buffer mixing with NPN (1:1 ratio) was used as blank.
2.5. Endogenous ROS Levels
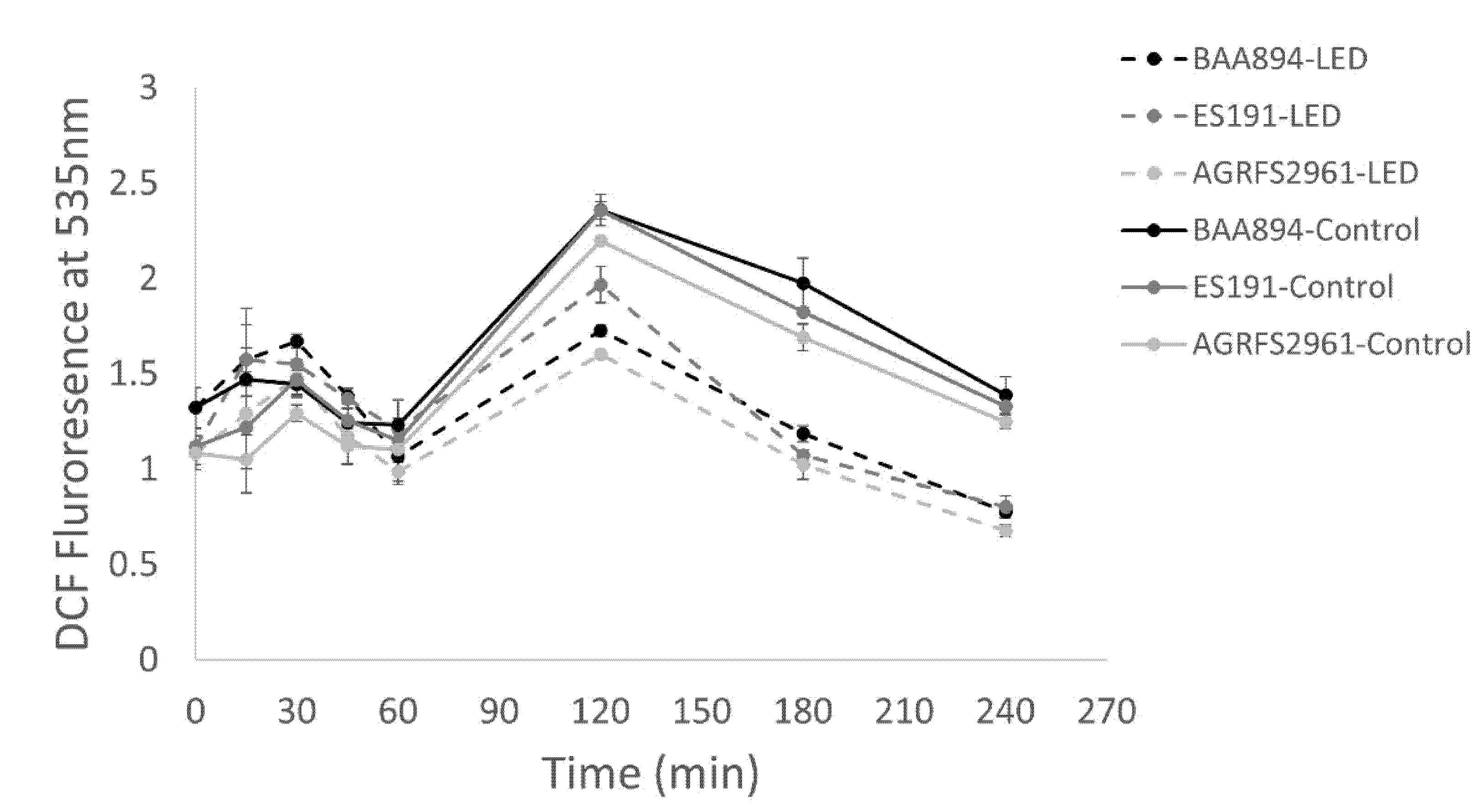
One ml of a suspension of 7 log CFU/mL stationary phase cells in PBS of each strain was incubated with 2′, 7′-dichlorfluorescein-diacetate, Sigma (DCFH–DA), 2 μM, final concentration for 10 min at 25 °C and followed by a 4-h light application. Samples were collected at 0 min, 15 min, 30 min, 45 min, 1 h, 2 h, 3 h and 4 h and then resuspended in PBS to remove extracellular ROS. A total 200 µL of each sample was transferred into a 96 well plate (Nunc™) and measured for fluorescence intensity using a fluorescence microplate reader (Varioskan™ LUX, Thermo Scientific, Waltham, MA, USA) at an excitation wavelength of 485 nm and emission wavelength of 535 nm [
15]. PBS incubated with (2 μM) DCFH–DA was blank as a negative reference.
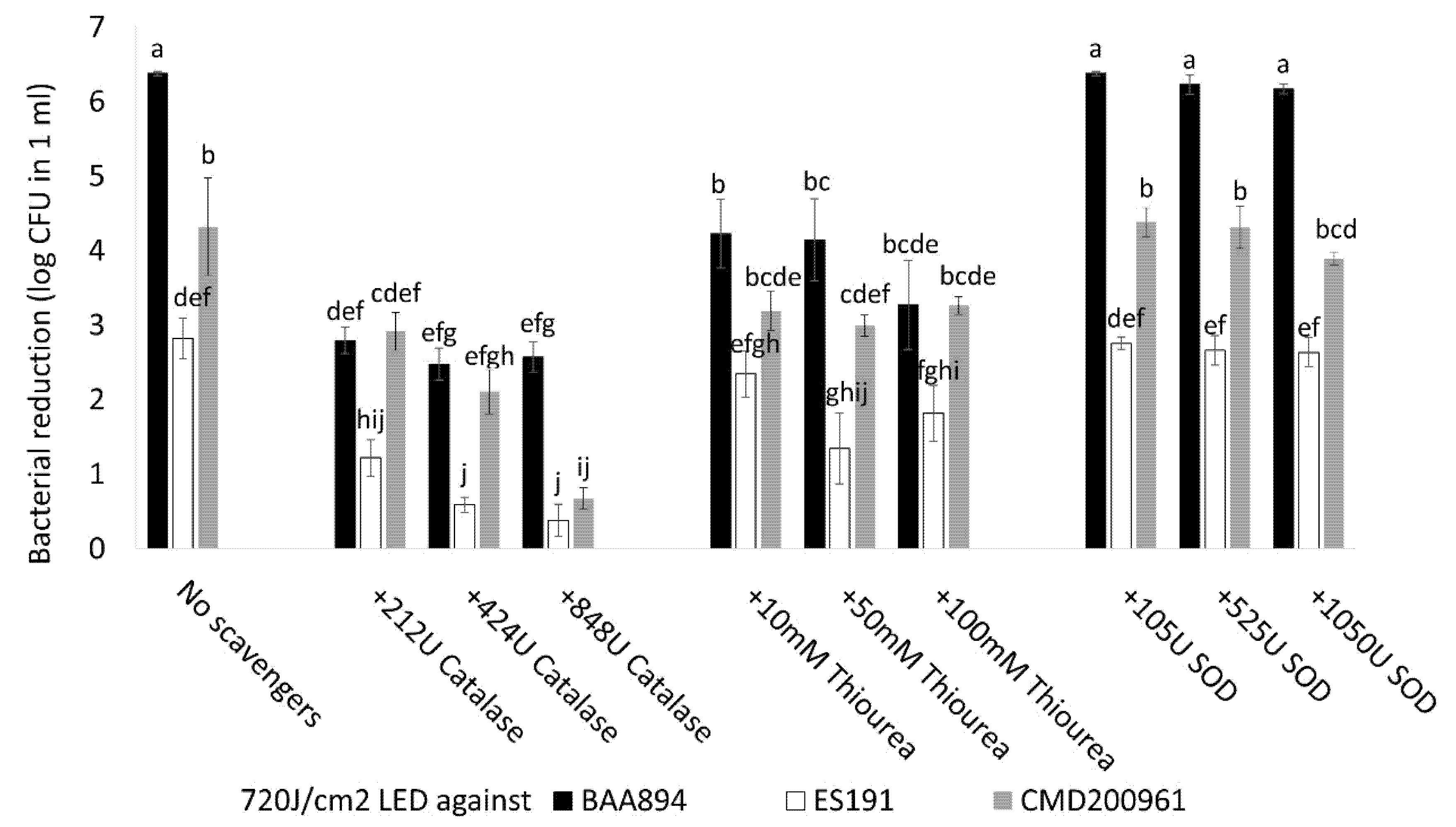
2.6. The Impact of External ROS Scavengers during Light Inactivation
Three different ROS scavengers were investigated, including two microbe-sourced ROS scavenger enzymes, catalase (
Aspergillus niger, Sigma, Burlington, MA, USA) and superoxide dismutase (
E. coli SOD, Sigma) and thiourea, (MilliporeSigma, Burlington, MA, USA) [
16]. For each strain, 6 log CFU/mL stationary phase cell suspensions containing different final concentrations of ROS scavenger were prepared. One ml aliquots of cell suspensions containing 212/424/848U catalase, 105/525/1050U SOD, and 10/50/100 mM thiourea were placed into each well of 24 well plates and treated with blue-LED illumination for 4 h, at a dose of 720 J/cm
2. The cytotoxic effect of the applied ROS scavenger agent was also estimated using a replicate plate as control (no light application under the same conditions). A cell suspension containing no external ROS scavenger was also included as a reference group for each strain. All groups were sampled, and serial 10-fold dilutions were prepared for viable cell enumeration. All plates were incubated at 37 °C for 48 h. The bacterial log CFU reduction per ml was determined.
2.7. The Combined Effect of Sublethal Hydrogen Peroxide (H2O2) and Light Inactivation against Stationary Phase Cells from Overnight Culture in 10% Sterile Skim Milk Media
Suspensions of 8 log CFU/mL cells of each strain were obtained after 20 h overnight incubation at 37 °C in sterile 10% skim milk media (Difco) and resuspended in PBS. One ml aliquots of diluted cell suspension (5 log CFU/mL) containing 0.02% H2O2 (final concentration) were added into a 24 well plate and subsequently exposed to light treatment for 1 h (180 J/cm2). A replicate plate received no light as a control to estimate the effect of H2O2 itself. Cell suspensions without the addition of H2O2 or light application were included as a reference. After the experiment, the cell suspension in each well was collected by centrifuging at 10,000× g for 5 min and then resuspended in PBS immediately for cell enumeration by plate counting on TSB agar for 48 h incubation at 37 °C. The bacterial log CFU reduction per ml was determined.
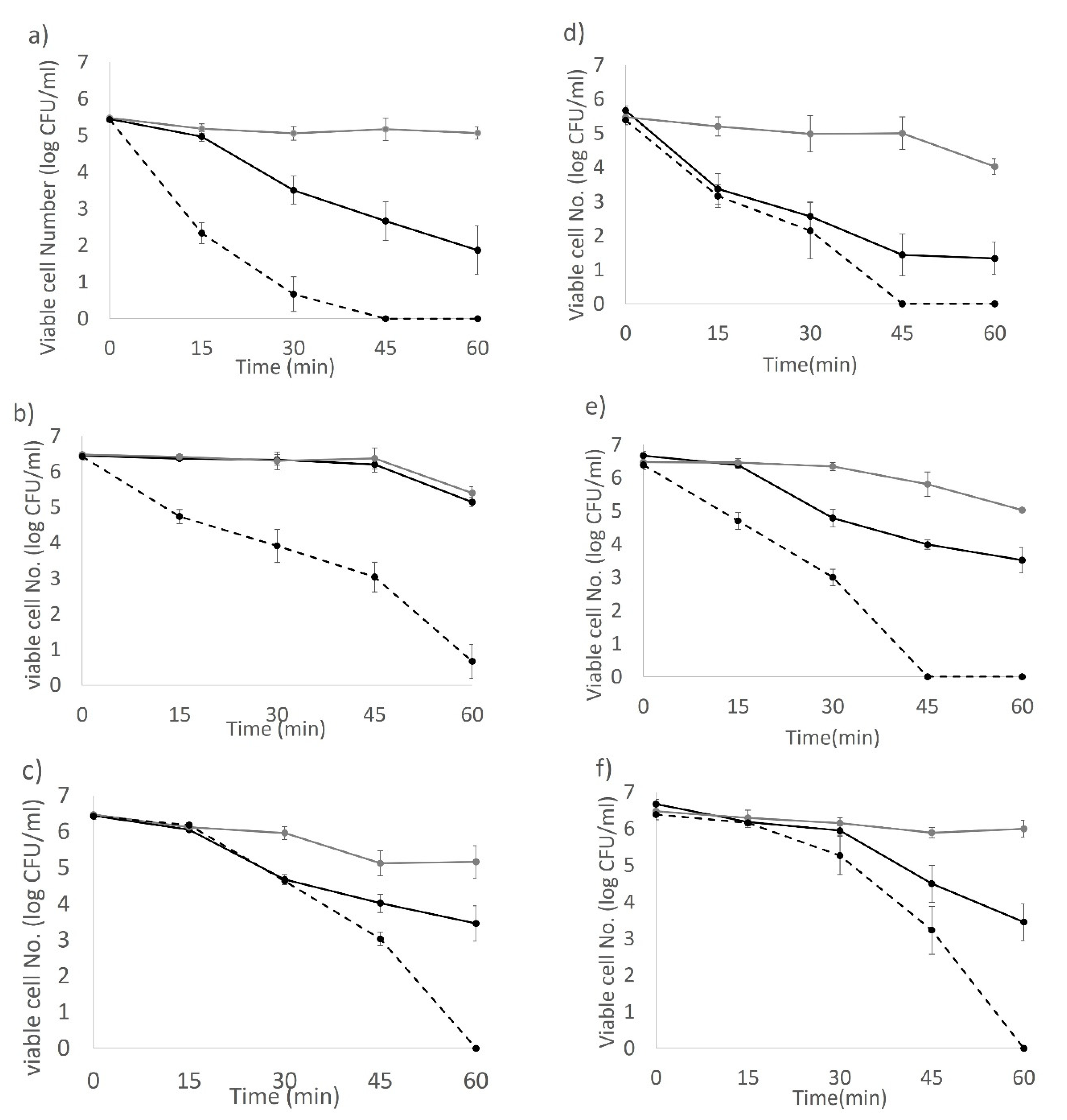
2.8. Against 3-Day-Old Biofilm Cells Grown from 10% Sterile Skim Milk
Sterile food grade stainless steel coupons (type 304, diameter of 2.5 cm, Agresearch Ltd., Palmerston North, New Zealand) and POM plastic (Sustarin C® polyoxymethylene copolymer commonly used for food conveyors, diameter of 2.5 cm, DOTMAR Ltd., Palmerston North, New Zealand) coupons were placed in 6-well cell culture plates (Nunclon DeltaTM). Coupon-containing wells were filled with 4 mL of sterile 10% skim milk media. Two hundred µL of 7 log CFU/mL stationary phase cells from an overnight culture in 10% skim milk was added into wells for each strain. Biofilms were grown on stainless steel and plastic coupons for 72 h at 37 °C statically. The 3-day-old biofilm on each coupon was washed in 30 mL of PBS three times to remove curdled milk and unattached bacterial cells. The blue-LED only treated coupons were immediately exposed to light illumination. For LED + H2O2 groups, external H2O2 (0.02%) was introduced before the light application by individually dipping each coupon into 0.02% H2O2 solution (100 mL) for 3 s then transferring into new 6 well plates for light illumination. A reference group was also set up, which consisted of coupons treated with 0.02% H2O2 in the absence of light illumination. All coupons were sampled by swabbing at 0, 15, 30, 45, and 60 min following light illumination (180 J/cm2), and the swabs were re-suspended in 10 mL 0.1% peptone H2O plus sterile glass beads. Undiluted sample suspension and 10-fold serial dilutions were plated on tryptic soy agar (TSA) plates and incubated at 37 °C for 48 h for counting detectable colonies.
2.9. Statistical Analysis
All experiments were performed in triplicate with biological duplicates. Experimental data were subjected to one-way ANOVA with Tukey’s HSD on Minitab 19 Statistical Software. Statistical significance was determined when p < 0.05.
4. Discussion
Huang et al. recently reported that the treatment of stainless steel using 405 nm LED illumination (375 J/cm
2 of; 26 mW/cm
2 for 4 h) reduced the population of
C. sakazakii in biofilm by 2.0 log CFU at 25 °C, 2.5 log CFU at 10 °C, and 2.0 log CFU at 4 °C [
12]. We used a surface mounted diode LED prototype with a stronger light intensity (50 mW/cm
2). At 2 h of light application (360 J/cm
2), 3 log CFU reductions were observed for all tested strains on both stainless steel and plastic surfaces (
Supplementary Figures S1 and S2). A biofilm developed by BAA894 cells was inactivated further to undetectable levels within 3-h light exposure while biofilms of either ES191 or AGRFS2961 persisted over 6 h. These findings indicate that the efficacy of biofilm removal may be related to strain-specific abilities of cell attachment or affinity to different materials.
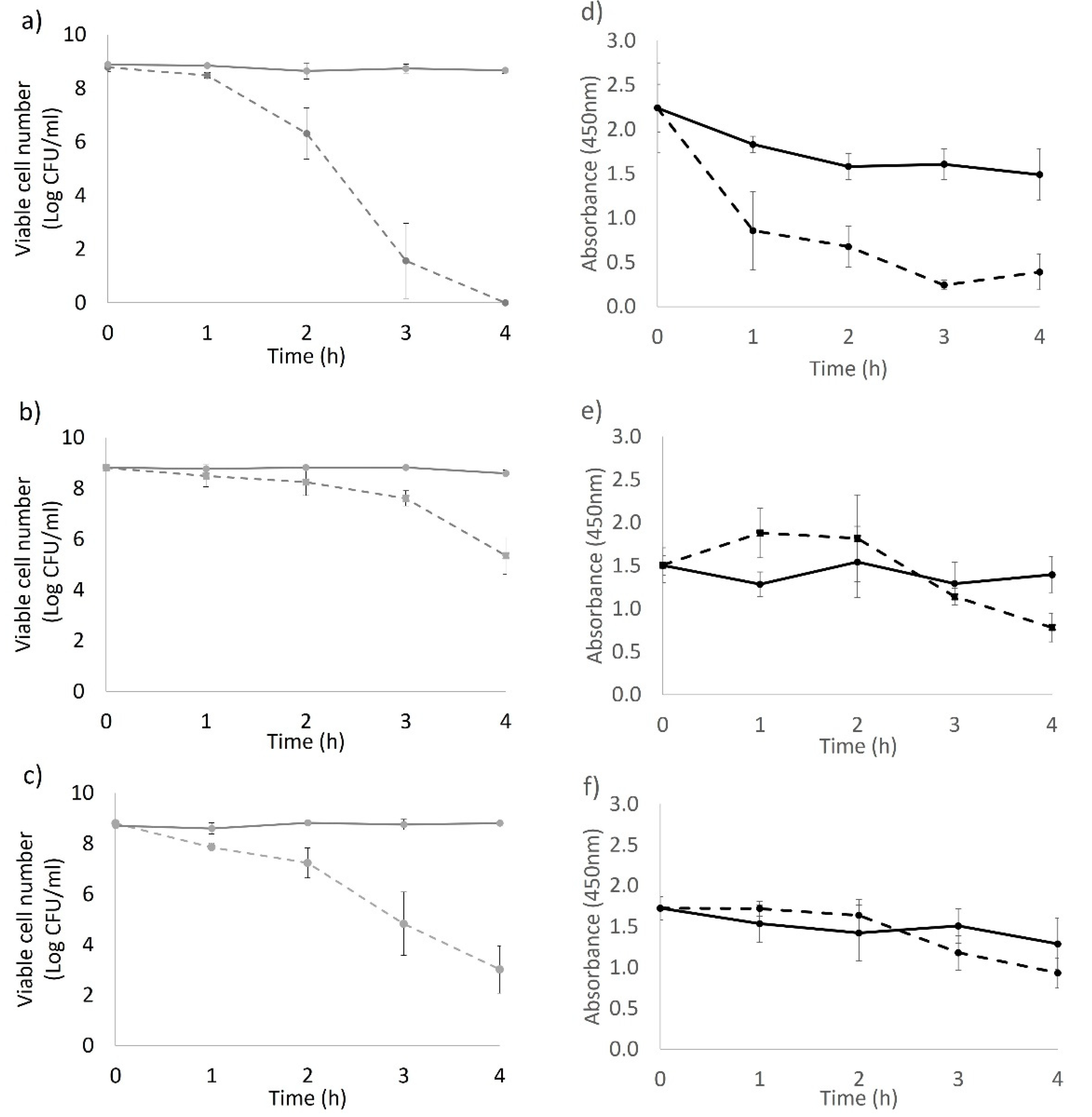
The bacterial response to the applied 405 nm light stress varied among the tested strains. For the BAA894 strain, although at a low metabolic rate after 1 h light treatment (
Figure 1d), the light stressed cells remained recoverable until they were inactivated by an intracellular ROS level (triggered with extended light dose) that was high enough to cause irreversible cell death [
17]. The XTT activity of strain ES191 and AGRFS2961 showed that the whole population could sustain comparable metabolic rates as the control cells during the light treatment (
p > 0.05) (
Figure 1e,f), suggesting that the light treated populations were still metabolically active (
p < 0.05). The DCFH–DA assay demonstrated that ROS production increased after 1 h of light treatment (where the cell loss became significant) (
Figure 1 and
Figure 3), supporting the finding that light induced ROS against many different bacteria was time-dependent [
9,
10]. After 2 h, blue-LED treated samples had a reduced number of viable cells, which led to the decrease of overall ROS levels in the whole population. The ROS level of the control groups was also increased at 1 h. The higher ROS in control population could be corelated with the higher number of viable cells actively responding to electron transportation and respiration. In other words, ROS level per cell could be varied between the irradiated groups and the control groups. However, the assumption needs to confirm by other methods (e.g., flow cytometry) in the future. A review of literature has not revealed which metabolic responses could be associated with the increase of blue-light resistance.
Notably, we observed that ES191 and AGRFS2961 produced distinct yellow pigment on TSB plates compared with the colony colour of BAA894. The yellow pigment of
C. sakazakii strain was previously identified with having carotenogenic nature on molecular/chemical level and BAA894 strain has been reported to carry gene clusters responsible for carotenoid biosynthesis [
18,
19]. Carotenoid could function as antioxidants to remove intracellular ROS and influence cellular membrane structure/fluidity in the harmful environment [
20]. The ability of carotenoid biosynthesis or similar pigmentation pathways [
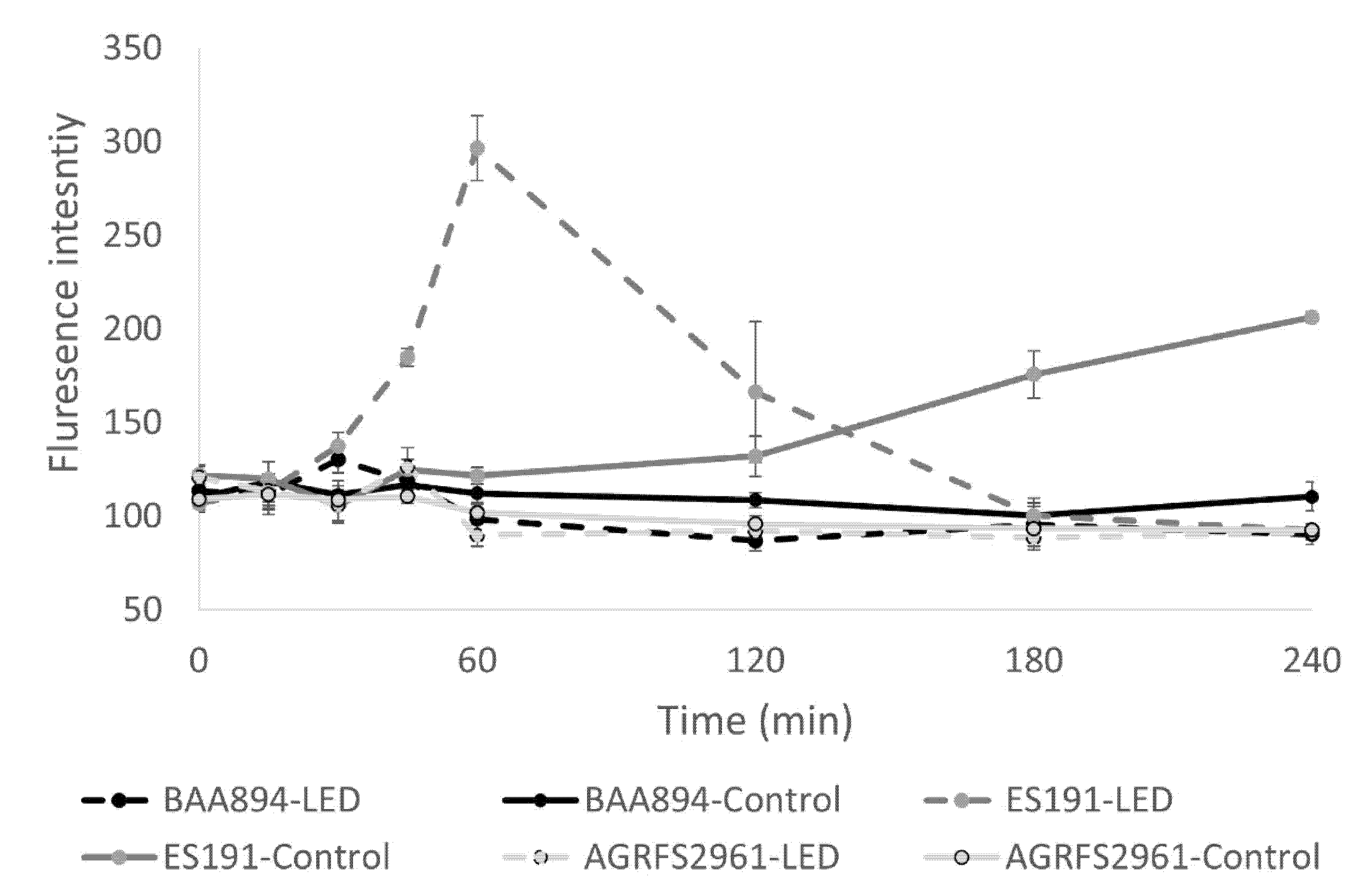
6] should be further investigated for ES191/AGRFS2961 to clarify the possible mechanism of antimicrobial-blue-light resistance. We hypothesize that the contribution of pigment may play varied roles in different strains since our NPN test showed that the permeability of the outer cell membrane was distinct between ES191 and AGRFS2961 (
Figure 2). Edward et al. [
21] demonstrated that the efficiency of NPN binding to phospholipids was influenced by the redox state of cytochrome o incorporated into phospholipid vesicles in
E. coli. The addition of H
2O
2 resulted in the immediate reoxidation of cytochrome o and loss of the NPN fluorescence increase observed [
21]. We hypothesize that blue-LED induced ROS, particular H
2O
2 (the focus in our study), possibly inhibited NPN uptake in a strain-dependent manner, as evidenced by constant cell permeability in AGRFS2961/BAA894 strain whereas ES191 had a significant change on the cell permeability, resulting in a differentiated light-susceptible phenotype. Another report found that
E. coli carrying the cloned cytochrome d terminal oxidase complex (membrane-bound cytochrome) are sensitive to near-UV inactivation [
22], suggesting that reoxidation of cytochrome could also be linked to the observed change in permeability of
C. sakazakii outer membrane when exposed to lethal ROS induced by 405nm blue-LED. However, we admitted that the cell permeability measured by NPN was a phenotypic character for the whole tested cell group. The tested population was assumed to be comprised of viable/damaged/inactivated cells and the ratio of cells in different fitness could be in a dynamic change during the test. Therefore, it is important to determine the cell permeability for viable cells or inactivated cells individually. Again, an investigation by flow cytometry would facilitate to understand at this point.
Photosensitization is an intrinsic process that enhances ROS production in bacteria during photoinactivation and can be influenced by using additional photosensitizers or external ROS scavengers [
23]. Our study suggests that the antimicrobial activity of 405 nm blue light was associated with the type of ROS as three different ROS scavengers (catalase, thiourea, and SOD) produced different responses (
Figure 4). Catalase showed the most significant effect for population survival under a lethal light dose (720 J/cm
2) compared with thiourea and SOD (
p < 0.05). Catalases reduce H
2O
2 into H
2O and O
2. Effective removal/reduction of H
2O
2 by catalase is assumed to be critical for the survival of tested
C. sakazakii strains when exposed to the blue-LED induced stress. Thiourea is known to reduce OH·, which is a type of ROS derived from H
2O
2 and its effect in mitigating antimicrobial light efficacy was found to be significant for BAA894 strain than the other two strains (
Figure 4), indicating that BAA894 strain could be more sensitive to a particular ROS stress, e.g., H
2O
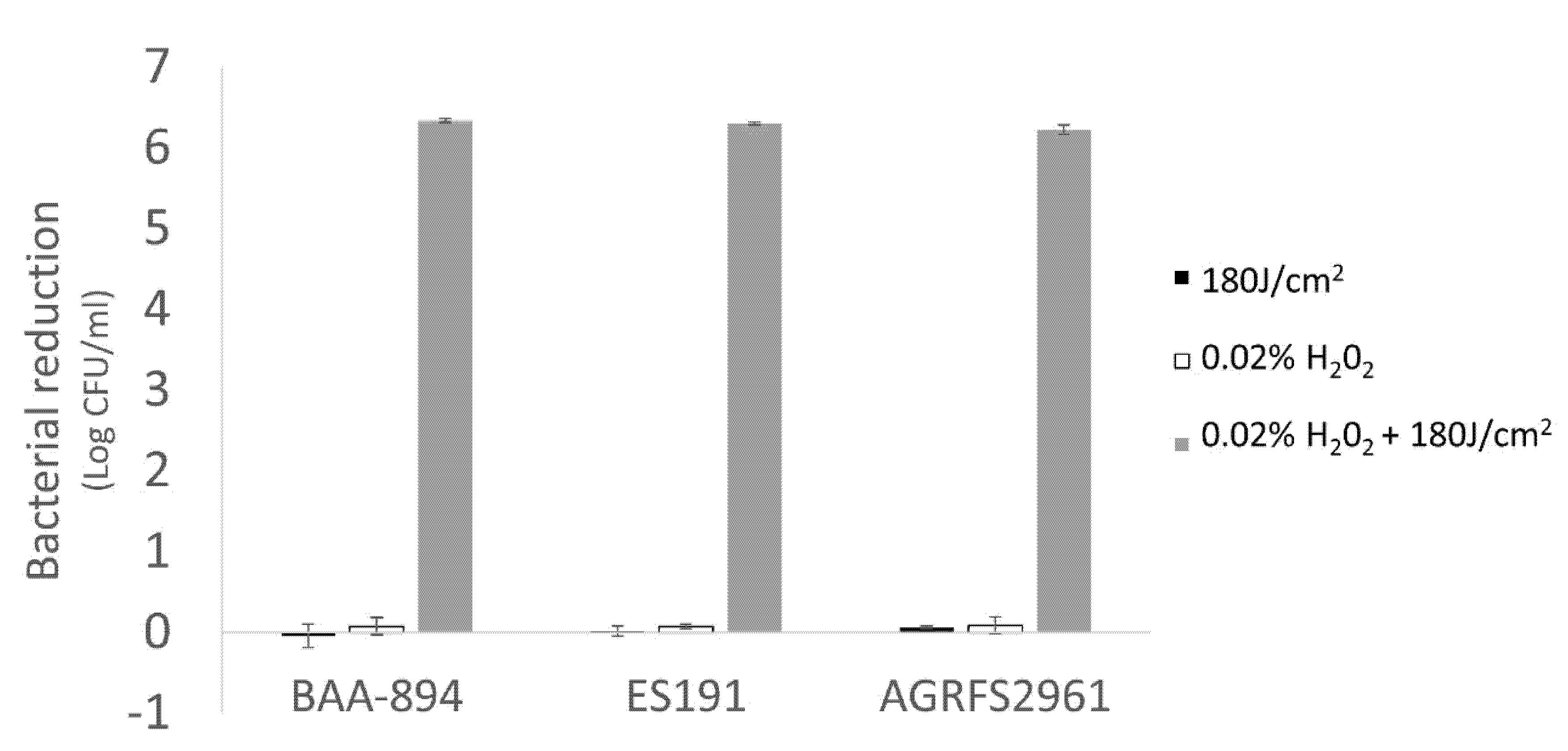
2 associated species. A supplementary experiment determined that our tested strains had different sensitivities to H
2O
2 after a long-time incubation (up to 4 h) at 25 °C and strain BAA894 was significantly more sensitive compared with ES191 and AGRFS2961 (
Supplementary Figure S3). Exogenous H
2O
2 could participate in cell damaging and could act as an inhibitor of catalase [
24]. Our investigation showed that H
2O
2 (either endogenous or exogenous) had a significant role during the 405 nm light inactivation (
Figure 4,
Figure 5 and
Figure 6). Biofilm cells of strains ES191 and AGRFS2961 were resistant to 6 h of the light exposure on both stainless steel and POM coupons (
Supplementary Figures S1 and S2). ES191 strain was the most light-resistant strain among three tested strains as it is a known super biofilm former [
13] and produced higher amounts of EPS than BAA894/AGRFS2961 (
Supplementary Figure S4). However, the combined effect of 0.02% H
2O
2 and 405 nm blue-LED (sublethal light dose at 180 J/cm
2; sublethal concentration of H
2O
2 at 0.02%) could increase the inactivation efficiency against persistent biofilm cells by achieving shortened treatment time (reduced from 6 h to 1 h) and additional 2–4 log reduction (
Figure 6). Feuerstein et al. reported that the combination of blue light (450–490 nm) and 0.3 mM H
2O
2 yielded 96% growth inhibition of
Streptococcus mutans, whereas single treatments of blue light orH
2O
2 only resulted in a bacterial growth reduction of 3% or 30%, respectively [
25]. Therefore, the combined of antimicrobial blue light and H
2O
2 could enhance the cytotoxic effect and induce irreversible cell damage rapidly. The current evidence could serve as a scientific guidance for future development/adaption of antimicrobial visible LED technology into dairy processing scenarios.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





 , control groups (with 0.02% H2O2);
, control groups (with 0.02% H2O2);  , blue-LED illuminated group;
, blue-LED illuminated group;  ; blue-LED with 0.02% H2O2.
; blue-LED with 0.02% H2O2.
