
Comparative Effect of Cricket Protein Powder and Soy Protein Isolate on Gel Properties of Indian Mackerel Surimi



Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation of Surimi
2.3. Preparation of Surimi Gel Added with Protein Additives
2.4. Oscillatory Dynamic Rheology
2.5. Trichloroacetic acid (TCA)-Soluble Peptide
2.6. Breaking Force, Deformation, Gel Strength and Texture Profile Analysis (TPA)
2.7. Expressible Moisture Content (EMC)
2.8. Whiteness
2.9. Molecular Driving Forces
2.10. Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
2.11. Fourier Transform Infrared (FTIR) Spectra
2.12. Scanning Electron Microscopy (SEM)
2.13. Sensory Property
2.14. Thiobarbituric Acid Reactive Substances (TBARS)
2.15. Statistical Analysis
3. Results and Discussion
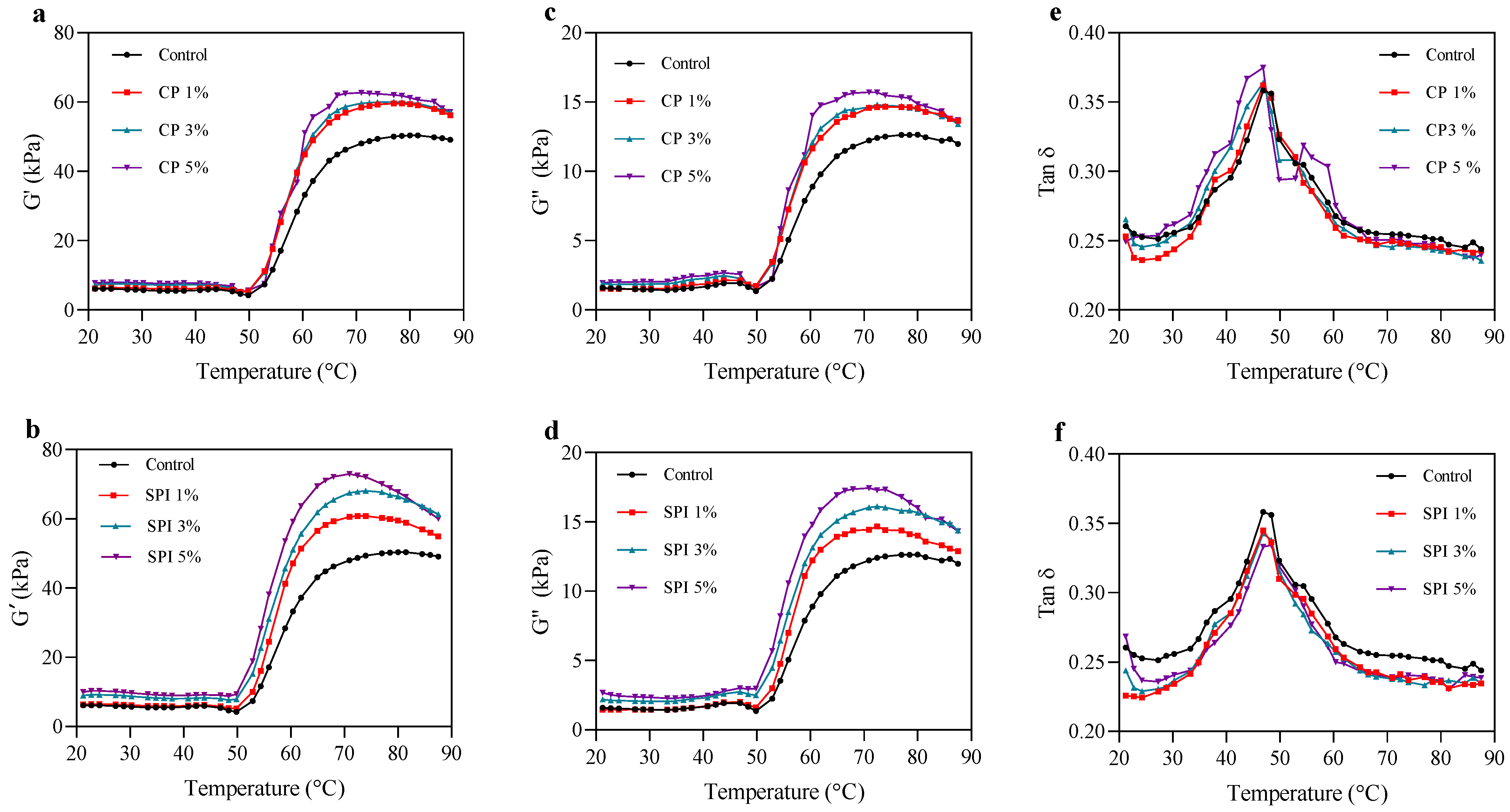
3.1. Dynamic Rheological Properties
3.2. Gel Properties
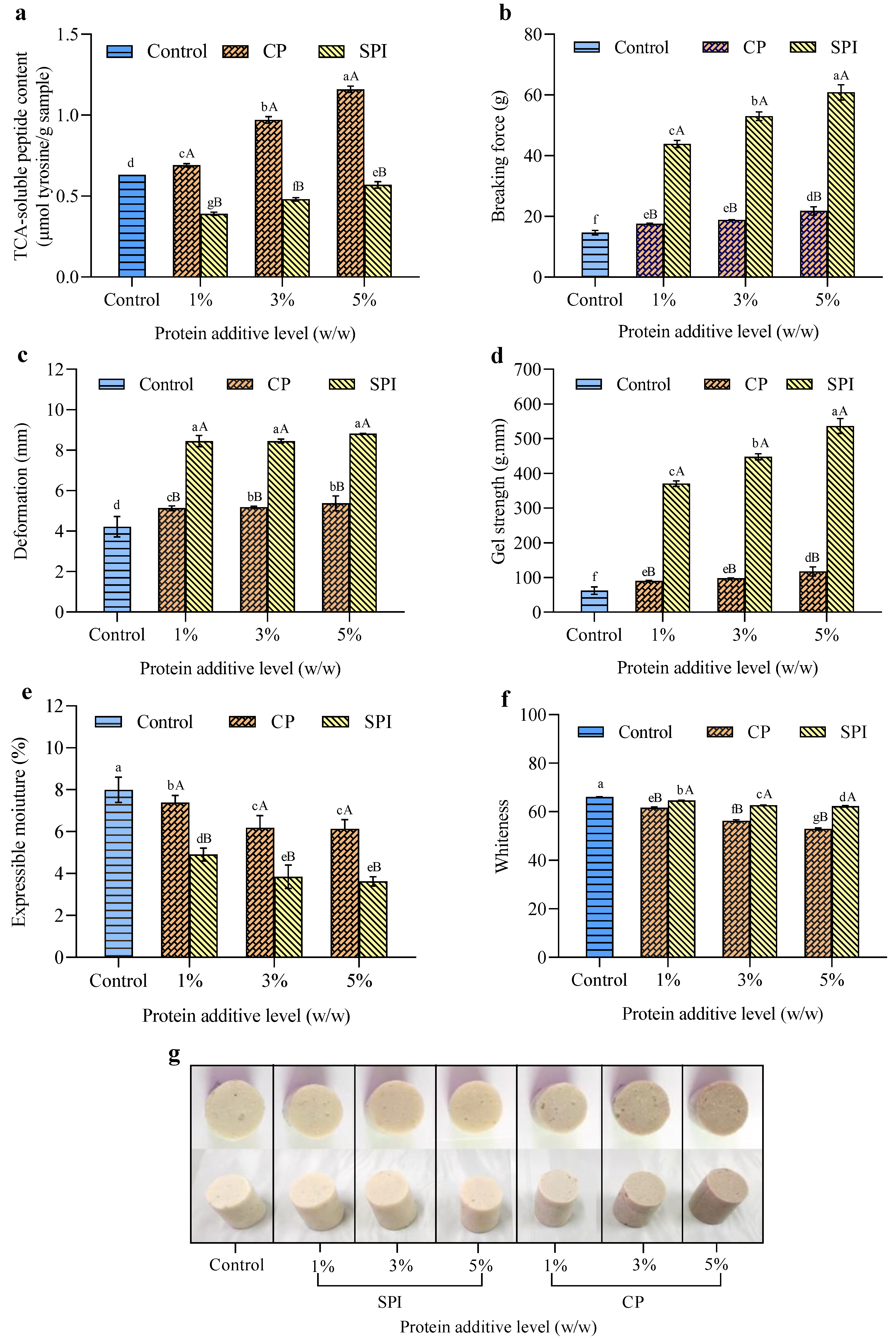
3.2.1. TCA-Soluble Peptide Content
3.2.2. Breaking Force, Deformation, and Gel Strength
3.2.3. EMC
3.2.4. Whiteness and Appearance
3.3. TPA
3.4. Molecular Driving Forces
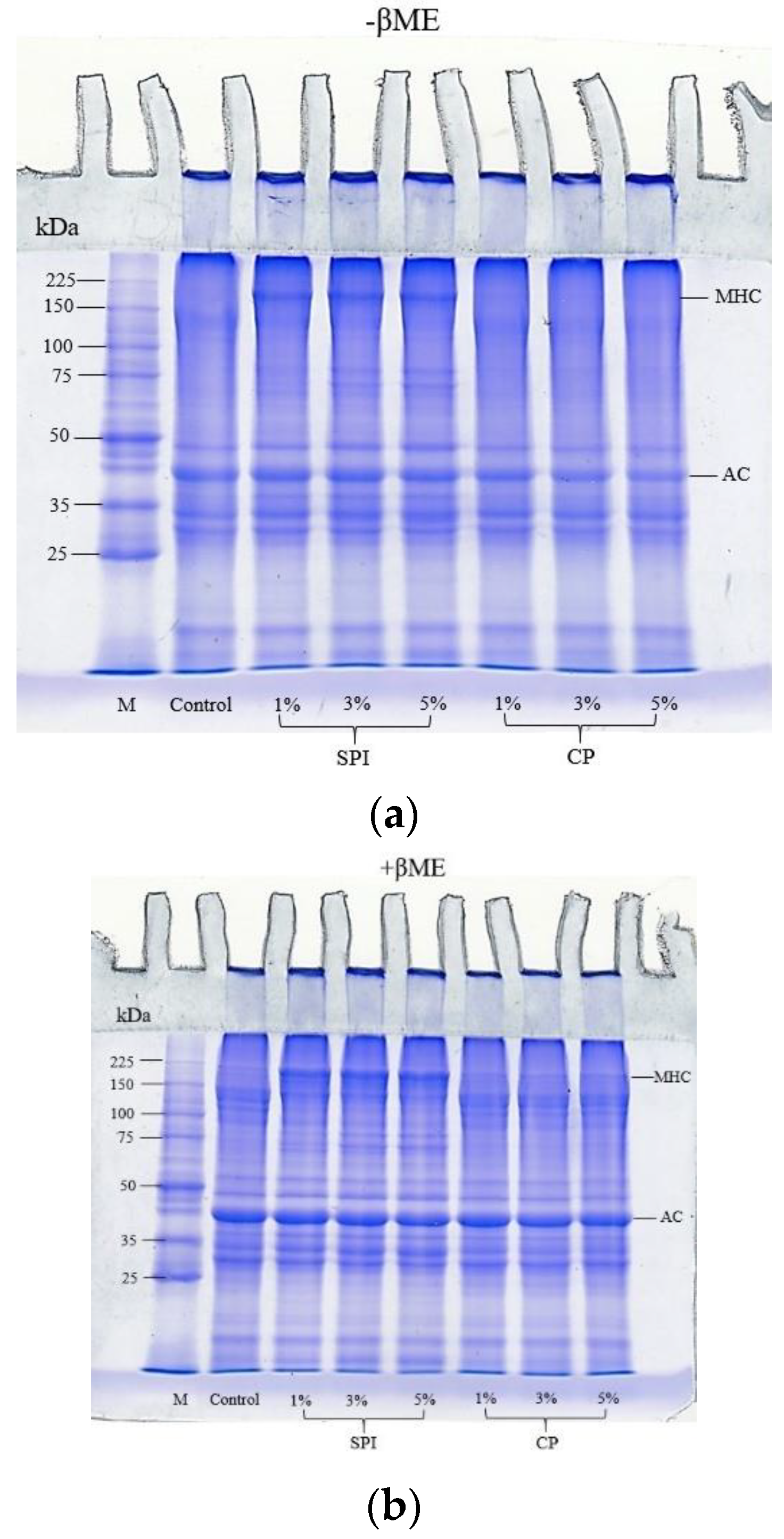
3.5. SDS-PAGE Protein Patterns
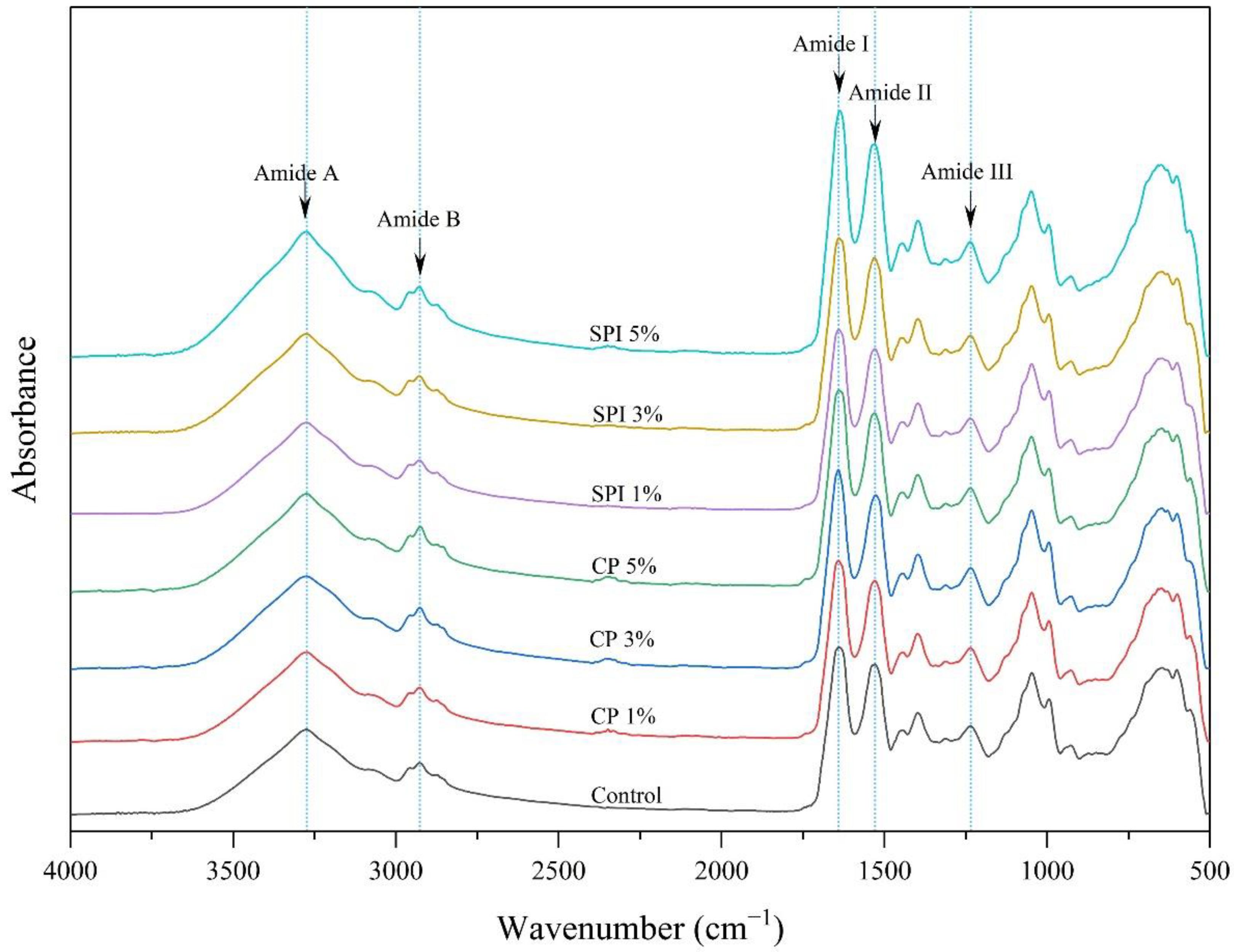
3.6. FTIR Spectroscopy of Gels
3.7. Microstructure
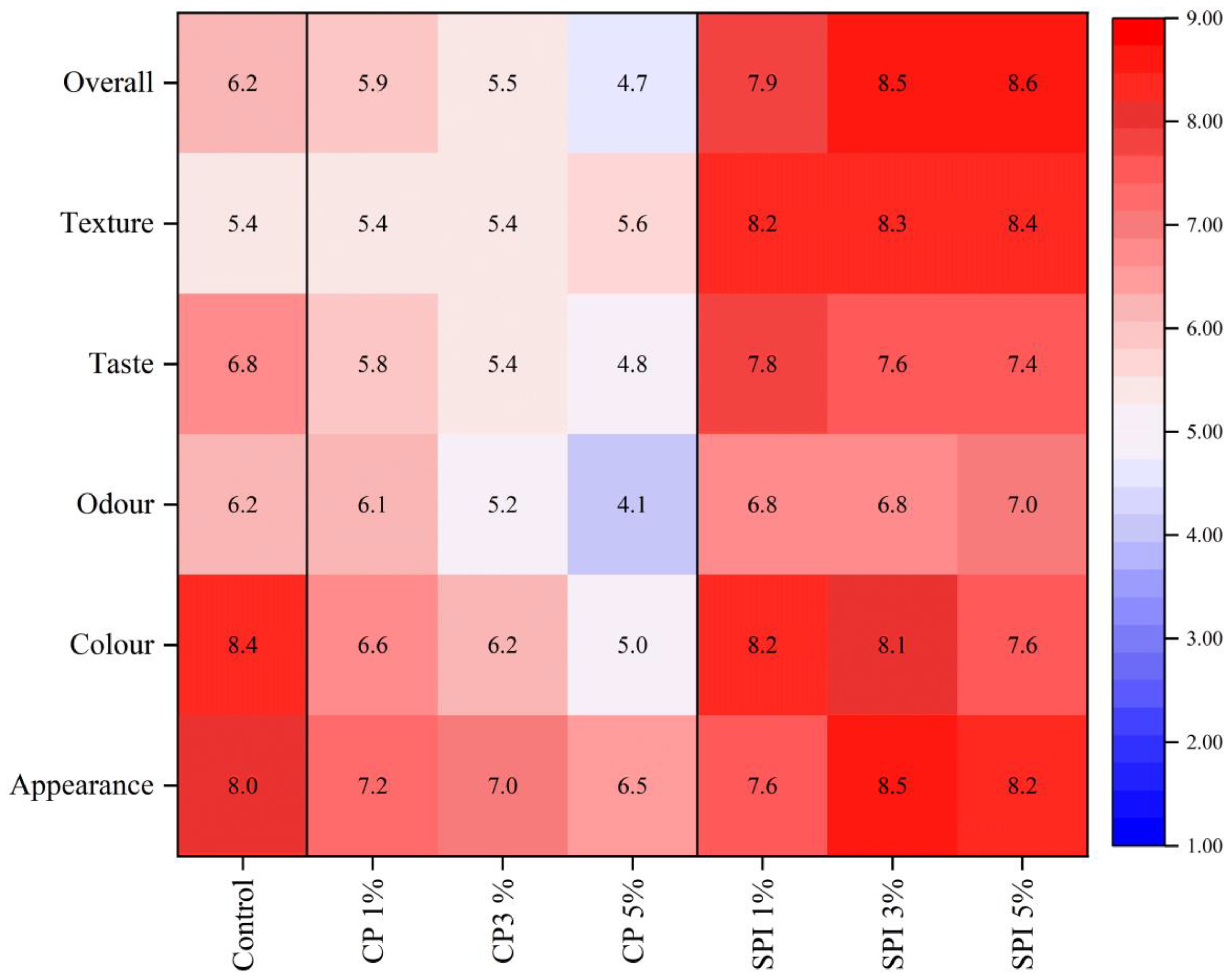
3.8. Sensory Properties
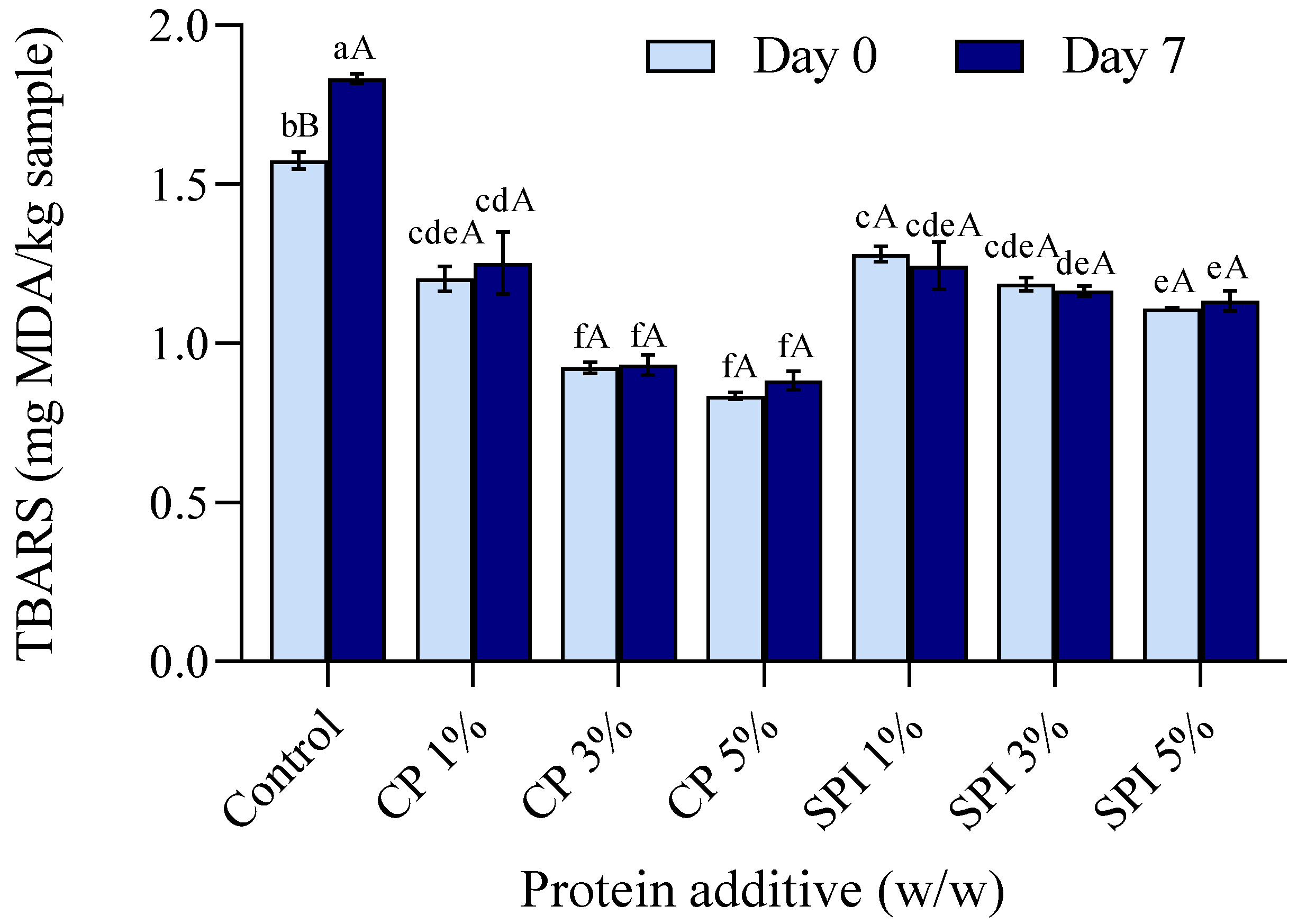
3.9. Lipid Oxidation
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tadpitchayangkoon, P.; Park, J.W.; Yongsawatdigul, J. Gelation characteristics of tropical surimi under water bath and ohmic heating. LWT 2012, 46, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Somjid, P.; Panpipat, W.; Cheong, L.Z.; Chaijan, M. Reduced washing cycle for sustainable mackerel (Rastrelliger kanagurta) surimi production: Evaluation of bio-physico-chemical, rheological, and gel-forming properties. Foods 2021, 10, 2717. [Google Scholar] [CrossRef]
- Chaijan, M.; Benjakul, S.; Visessanguan, W.; Faustman, C. Characteristics and gel properties of muscles from sardine (Sardinella gibbosa) and mackerel (Rastrelliger kanagurta) caught in Thailand. Food Res. Int. 2004, 37, 1021–1030. [Google Scholar] [CrossRef]
- Department of Fisheries. Fisheries Statistics of Thailand; Fishery Statistics Analysis and Research Group, Fisheries Development Policy and Strategy Division, Department of Fisheries: Bangkok, Thailand, 2022. [Google Scholar]
- Panpipat, W.; Chaijan, M.; Benjakul, S. Gel properties of croaker-mackerel surimi blend. Food Chem. 2010, 122, 1122–1128. [Google Scholar] [CrossRef]
- Lin, X.; Yang, W.; Xu, D.; Jie, Z.; Liu, W. Improving gel properties of hairtail surimi by electron irradiation. Radiat. Phys. Chem. 2015, 110, 1–5. [Google Scholar] [CrossRef]
- Liang, F.; Zhu, Y.; Ye, T.; Jiang, S.; Lin, L.; Lu, J. Effect of ultrasound assisted treatment and microwave combined with water bath heating on gel properties of surimi-crabmeat mixed gels. LWT 2020, 133, 110098. [Google Scholar] [CrossRef]
- Guo, Z.; Li, Z.; Wang, J.; Zheng, B. Gelation properties and thermal gelling mechanism of golden threadfin bream myosin containing CaCl2 induced by high pressure processing. Food Hydrocoll. 2019, 95, 43–52. [Google Scholar] [CrossRef]
- Jiao, X.; Cao, H.; Fan, D.; Huang, J.; Zhao, J.; Yan, B.; Zhou, W.; Zhang, W.; Ye, W.; Zhang, H. Effects of fish oil incorporation on the gelling properties of silver carp surimi gel subjected to microwave heating combined with conduction heating treatment. Food Hydrocoll. 2019, 94, 164–173. [Google Scholar] [CrossRef]
- Rawdkuen, S.; Benjakul, S. Whey protein concentrate: Autolysis inhibition and effects on the gel properties of surimi prepared from tropical fish. Food Chem. 2008, 106, 1077–1084. [Google Scholar] [CrossRef]
- Kong, W.; Zhang, T.; Feng, D.; Xue, Y.; Wang, Y.; Li, Z.; Xue, C. Effects of modified starches on the gel properties of Alaska Pollock surimi subjected to different temperature treatments. Food Hydrocoll. 2016, 56, 20–28. [Google Scholar] [CrossRef]
- Zhou, X.; Chen, T.; Lin, H.; Chen, H.; Liu, J.; Lyu, F.; Ding, Y. Physicochemical properties and microstructure of surimi treated with egg white modified by tea polyphenols. Food Hydrocoll. 2019, 90, 82–89. [Google Scholar] [CrossRef]
- Balange, A.; Benjakul, S. Enhancement of gel strength of bigeye snapper (Priacanthus tayenus) surimi using oxidised phenolic compounds. Food Chem. 2009, 113, 61–70. [Google Scholar] [CrossRef]
- Arsyad, M.A.; Akazawa, T.; Ogawa, M. Effects of olive leaf powder on mechanical properties of heat-induced surimi gel. J. Aquat. Food Prod. Technol. 2019, 28, 2–13. [Google Scholar] [CrossRef]
- Li, Y.; Xiong, S.; Yin, T.; Hu, Y.; You, J. The gastric digestion kinetics of silver carp (Hypophthalmichthys molitrix) surimi gels induced by transglutaminase. Food Chem. 2019, 283, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Amiza, M.A.; Kang, W.C. Effect of chitosan on gelling properties, lipid oxidation, and microbial load of surimi gel made from African catfish (Clarias gariepinus). Int. Food Res. J. 2013, 20, 1585–1594. [Google Scholar]
- Hernández-Briones, A.; Velázquez, G.; Vázquez, M.; Ramírez, J.A. Effects of adding fish gelatin on Alaska pollock surimi gels. Food Hydrocoll. 2009, 23, 2446–2449. [Google Scholar] [CrossRef]
- Petcharat, T.; Chaijan, M.; Karnjanapratum, S. Effect of furcellaran incorporation on gel properties of sardine surimi. Int. J. Food Sci. Technol. 2021, 56, 5957–5967. [Google Scholar] [CrossRef]
- Zhang, H.; Xiong, Y.; Bakry, A.M.; Xiong, S.; Yin, T.; Zhang, B.; Huang, Q. Effect of yeast β-glucan on gel properties, spatial structure and sensory characteristics of silver carp surimi. Food Hydrocoll. 2019, 88, 256–264. [Google Scholar] [CrossRef]
- Shitole, S.S.; Balange, A.K.; Gangan, S.S. Use of seaweed (Sargassum tenerrimum) extract as gel enhancer for lesser sardine (Sardinella brachiosoma) surimi. Int. Aquat. Res. 2014, 6, 1–11. [Google Scholar] [CrossRef]
- Skotnicka, M.; Karwowska, K.; Kłobukowski, F.; Borkowska, A.; Pieszko, M. Possibilities of the development of edible insect-based foods in Europe. Foods 2021, 10, 766. [Google Scholar] [CrossRef]
- Ndiritu, A.K.; Kinyuru, J.N.; Gichuhi, P.N.; Kenji, G.M. Effects of NaCl and pH on the functional properties of edible crickets (Acheta domesticus) protein concentrate. J. Food Meas. Charact. 2019, 13, 1788–1796. [Google Scholar] [CrossRef]
- Hall, F.G.; Jones, O.G.; O'Haire, M.E.; Liceaga, A.M. Functional properties of tropical banded cricket (Gryllodes sigillatus) protein hydrolysates. Food Chem. 2017, 224, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.K.; Yong, H.I.; Jeong, C.H.; Han, S.G.; Kim, Y.B.; Paik, H.D.; Choi, Y.S. Technical functional properties of water-and salt-soluble proteins extracted from edible insects. Food Sci. Anim. Resour. 2019, 39, 643–654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scholliers, J.; Steen, L.; Fraeye, I. Gelation of a combination of insect and pork proteins as affected by heating temperature and insect: Meat ratio. Food Res. Int. 2020, 137, 109703. [Google Scholar] [CrossRef]
- Chaijan, M.; Chumthong, K.; Kongchoosi, N.; Chinarak, K.; Panya, A.; Phonsatta, N.; Cheong, L.Z.; Panpipat, W. Characterisation of pH-shift-produced protein isolates from sago palm weevil (Rhynchophorus ferrugineus) larvae. J. Insects Food Feed. 2022, 8, 313–324. [Google Scholar] [CrossRef]
- Osimani, A.; Milanović, V.; Cardinali, F.; Roncolini, A.; Garofalo, C.; Clementi, F.; Pasquini, M.; Mozzon, M.; Foligni, R.; Raffaelli, N.; et al. Bread enriched with cricket powder (Acheta domesticus): A technological, microbiological and nutritional evaluation. Innov. Food Sci. Emerg. Technol. 2018, 48, 150–163. [Google Scholar] [CrossRef]
- Kowalczewski, P.Ł.; Gumienna, M.; Rybicka, I.; Górna, B.; Sarbak, P.; Dziedzic, K.; Kmiecik, D. Nutritional value and biological activity of gluten-free bread enriched with cricket powder. Molecules 2021, 26, 1184. [Google Scholar] [CrossRef]
- Homann, A.M.; Ayieko, M.A.; Konyole, S.O.; Roos, N. Acceptability of biscuits containing 10% cricket (Acheta domesticus) compared to milk biscuits among 5-10-year-old Kenyan schoolchildren. J. Insects Food Feed. 2017, 3, 95–103. [Google Scholar] [CrossRef]
- Biró, B.; Sipos, M.A.; Kovács, A.; Badak-Kerti, K.; Pásztor-Huszár, K.; Gere, A. Cricket-enriched oat biscuit: Technological analysis and sensory evaluation. Foods 2020, 9, 1561. [Google Scholar] [CrossRef]
- Duda, A.; Adamczak, J.; Chełmińska, P.; Juszkiewicz, J.; Kowalczewski, P. Quality and nutritional/textural properties of durum wheat pasta enriched with cricket powder. Foods 2019, 8, 46. [Google Scholar] [CrossRef] [Green Version]
- Mi, H.; Zhao, Y.; Li, Y.; Chen, J.; Liu, H.; Yi, S.; Li, X.; Li, J. Combining effect of soybean protein isolate and transglutaminase on the gel properties of Zhikong scallop (Chlamys farreri) adductor muscle. LWT 2021, 138, 110727. [Google Scholar] [CrossRef]
- Wang, Z.; Liang, J.; Jiang, L.; Li, Y.; Wang, J.; Zhang, H.; Li, D.; Han, F.; Li, Q.; Wang, R.; et al. Effect of the interaction between myofibrillar protein and heat-induced soy protein isolates on gel properties. CyTA J. Food. 2015, 13, 527–534. [Google Scholar] [CrossRef]
- Luo, Y.; Shen, H.; Pan, D.; Bu, G. Gel properties of surimi from silver carp (Hypophthalmichthys molitrix) as affected by heat treatment and soy protein isolate. Food Hydrocoll. 2008, 22, 1513–1519. [Google Scholar] [CrossRef]
- Park, J.W. Functional protein additives in surimi gels. J. Food Sci. 1994, 59, 525–527. [Google Scholar] [CrossRef]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Morrissey, M.T.; Wu, J.W.; Lin, D.; An, H. Protease inhibitor effects on torsion measurements and autolysis of Pacific whiting surimi. J. Food Sci. 1993, 58, 1050–1054. [Google Scholar] [CrossRef]
- Chaijan, M.; Panpipat, W.; Benjakul, S. Physicochemical properties and gel-forming ability of surimi from three species of mackerel caught in Southern Thailand. Food Chem. 2010, 121, 85–92. [Google Scholar] [CrossRef]
- Panpipat, W.; Chaijan, M. Functional properties of pH-shifted protein isolates from bigeye snapper (Priacanthus tayenus) head by-product. Int. J. Food Prop. 2017, 20, 596–610. [Google Scholar] [CrossRef] [Green Version]
- Robinson, H.W.; Hogden, C.G. The biuret reaction in the determination of serum proteins I. A study of the conditions necessary for the production of a stable colour which bears a quantitative relationship to the protein concentration. J. Biol. Chem. 1940, 135, 707–725. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Somjid, P.; Panpipat, W.; Chaijan, M. Carbonated water as a novel washing medium for mackerel (Auxis thazard) surimi production. J. Food Sci. Technol. 2017, 54, 3979–3988. [Google Scholar] [CrossRef] [PubMed]
- Gani, A.; Benjakul, S. Impact of virgin coconut oil nanoemulsion on properties of croaker surimi gel. Food Hydrocoll. 2018, 82, 34–44. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Methods Enzymol. 1978, 52, 302–310. [Google Scholar] [PubMed]
- Huang, J.; Ye, B.; Wang, W.; Li, J.; Yi, S.; Li, X.; Xu, Y.; Mi, H. Incorporation effect of inulin and microbial transglutaminase on the gel properties of silver carp (Hypophthalmichthys molitrix) surimi. J. Food Meas. Charact. 2021, 15, 1–11. [Google Scholar] [CrossRef]
- Rao, M.A.; Cooley, H.J. Rheological behavior of tomato pastes in steady and dynamic shear. J. Texture Stud. 1992, 23, 415–425. [Google Scholar] [CrossRef]
- Zhang, F.; Fang, L.; Wang, C.; Shi, L.; Chang, T.; Yang, H.; Cui, M. Effects of starches on the textural, rheological, and color properties of surimi–beef gels with microbial tranglutaminase. Meat Sci. 2013, 93, 533–537. [Google Scholar] [CrossRef]
- Buamard, N.; Benjakul, S.; Konno, K. Improvement of gel quality of sardine surimi with low setting phenomenon by ethanolic coconut husk extract. J. Texture Stud. 2017, 48, 47–56. [Google Scholar] [CrossRef]
- Qiu, C.; Xia, W.; Jiang, Q. Effect of high hydrostatic pressure (HHP) on myofibril-bound serine proteinases and myofibrillar protein in silver carp (Hypophthalmichthys molitrix). Food Res. Int. 2013, 52, 199–205. [Google Scholar] [CrossRef]
- Singh, A.; Benjakul, S. Proteolysis and its control using protease inhibitors in fish and fish products: A review. Compr. Rev. Food Sci. Food Saf. 2018, 17, 496–509. [Google Scholar] [CrossRef] [Green Version]
- Zhang, T.; Xue, Y.; Li, Z.; Wang, Y.; Yang, W.; Xue, C. Effects of ozone-induced oxidation on the physicochemical properties of myofibrillar proteins recovered from bighead carp (Hypophthalmichthys nobilis). Food Bioprocess Technol. 2015, 8, 181–190. [Google Scholar] [CrossRef]
- Singh, A.; Prabowo, F.F.; Benjakul, S.; Pranoto, Y.; Chantakun, K. Combined effect of microbial transglutaminase and ethanolic coconut husk extract on the gel properties and in-vitro digestibility of spotted golden goatfish (Parupeneus heptacanthus) surimi gel. Food Hydrocoll. 2020, 109, 106107. [Google Scholar] [CrossRef]
- Mleko, S.; Foegeding, E.A. pH induced aggregation and weak gel formation of whey protein polymers. J. Food Sci. 2000, 65, 139–143. [Google Scholar] [CrossRef]
- Taherian, A.R.; Mondor, M.; Labranche, J.; Drolet, H.; Ippersiel, D.; Lamarche, F. Comparative study of functional properties of commercial and membrane processed yellow pea protein isolates. Food Res. Int. 2011, 44, 2505–2514. [Google Scholar] [CrossRef]
- Wang, S.F.; Smith, D.M. Dynamic rheological properties and secondary structure of chicken breast myosin as influenced by isothermal heating. J. Agric. Food Chem. 1994, 42, 1434–1439. [Google Scholar] [CrossRef]
- Klomklao, S.; Benjakul, S. Effect of trypsin inhibitor in adzuki bean (Vigna angularis) on proteolysis and gel properties of threadfin bream (Nemipterus bleekeri). LWT 2015, 63, 906–911. [Google Scholar] [CrossRef]
- Kudre, T.; Benjakul, S.; Kishimura, H. Effects of protein isolates from black bean and mungbean on proteolysis and gel properties of surimi from sardine (Sardinella albella). LWT 2013, 50, 511–518. [Google Scholar] [CrossRef]
- Nakashima, T.; Tokuyasu, K.; Funatsu, M. Studies on proteolytic enzyme of the cricket, Gryllulus taiwanemma Ohmachi. Agric. Biol. Chem. 1965, 29, 307–314. [Google Scholar] [CrossRef]
- Jiang, J.; Xiong, Y.L. Extreme pH treatments enhance the structure-reinforcement role of soy protein isolate and its emulsions in pork myofibrillar protein gels in the presence of microbial transglutaminase. Meat Sci. 2013, 93, 469–476. [Google Scholar] [CrossRef]
- Kosečková, P.; Zvěřina, O.; Pěchová, M.; Krulíková, M.; Duborská, E.; Borkovcová, M. Mineral profile of cricket powders, some edible insect species and their implication for gastronomy. J. Food Compos. Anal. 2022, 107, 104340. [Google Scholar] [CrossRef]
- Shah, A.A.; Wanapat, M. Gryllus testaceus walker (crickets) farming management, chemical composition, nutritive profile, and their effect on animal digestibility. Entomol. Res. 2021, 51, 639–649. [Google Scholar] [CrossRef]
- Benjakul, S.; Visessanguan, W.; Tanaka, M.; Ishizaki, S.; Suthidham, R.; Sungpech, O. Effect of chitin and chitosan on gelling properties of surimi from barred garfish (Hemiramphus far). J. Sci. Food Agric. 2001, 81, 102–108. [Google Scholar] [CrossRef]
- Chaijan, M.; Benjakul, S.; Visessanguan, W.; Faustman, C. Physicochemical properties, gel-forming ability and myoglobin content of sardine (Sardinella gibbosa) and mackerel (Rastrelliger kanagurta) surimi produced by conventional method and alkaline solubilisation process. Eur. Food Res. Technol. 2006, 222, 58–63. [Google Scholar] [CrossRef]
- Sze-Tao, K.W.C.; Sathe, S.K. Functional properties and in vitro digestibility of almond (Prunus dulcis L.) protein isolate. Food Chem. 2000, 69, 153–160. [Google Scholar] [CrossRef]
- Li, J.; Wu, M.; Wang, Y.; Li, K.; Du, J.; Bai, Y. Effect of pH-shifting treatment on structural and heat induced gel properties of peanut protein isolate. Food Chem. 2020, 325, 126921. [Google Scholar] [CrossRef] [PubMed]
- Bawa, M.; Songsermpong, S.; Kaewtapee, C.; Chanput, W. Nutritional, sensory, and texture quality of bread and cookie enriched with house cricket (Acheta domesticus) powder. J. Food Process. Preserv. 2020, 44, e14601. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, W.; Liu, X.; Qiao, M.; Yi, S.; Li, X.; Li, J. Effects of chickpea and peanut protein isolates on the gelling properties of hairtail (Trichiurus haumela) myosin. LWT 2022, 15, 113562. [Google Scholar] [CrossRef]
- Duangmal, K.; Taluengphol, A. Effect of protein additives, sodium ascorbate, and microbial transglutaminase on the texture and colour of red tilapia surimi gel. Int. J. Food Sci. Technol. 2010, 45, 48–55. [Google Scholar] [CrossRef]
- Chen, J.; Deng, T.; Wang, C.; Mi, H.; Yi, S.; Li, X.; Li, J. Effect of hydrocolloids on gel properties and protein secondary structure of silver carp surimi. J. Sci. Food Agric. 2020, 100, 2252–2260. [Google Scholar] [CrossRef]
- Pietrasik, Z.; Jarmoluk, A.; Shand, P.J. Effect of non-meat proteins on hydration and textural properties of pork meat gels enhanced with microbial transglutaminase. LWT 2007, 40, 915–920. [Google Scholar] [CrossRef]
- Lin, D.; Zhang, L.; Li, R.; Zheng, B.; Rea, M.C.; Miao, S. Effect of plant protein mixtures on the microstructure and rheological properties of myofibrillar protein gel derived from red sea bream (Pagrosomus major). Food Hydrocoll. 2019, 96, 537–545. [Google Scholar] [CrossRef]
- Liu, R.; Zhao, S.M.; Xie, B.J.; Xiong, S.B. Contribution of protein conformation and intermolecular bonds to fish and pork gelation properties. Food Hydrocoll. 2011, 25, 898–906. [Google Scholar] [CrossRef]
- Peng, I.C.; Nielsen, S.S. Protein-protein interactions between soybean beta-conglycinin (B1-B6) and myosin. J. Food Sci. 1986, 51, 588–590. [Google Scholar] [CrossRef]
- O'Kane, F.E.; Happe, R.P.; Vereijken, J.M.; Gruppen, H.; van Boekel, M.A. Heat-induced gelation of pea legumin: Comparison with soybean glycinin. J. Agric. Food Chem. 2004, 52, 5071–5078. [Google Scholar] [CrossRef] [PubMed]
- Alishahi, A.; Aïder, M. Applications of chitosan in the seafood industry and aquaculture: A review. Food Bioprocess Technol. 2012, 5, 817–830. [Google Scholar] [CrossRef]
- Huang, X.; Li, C.; Yang, F.; Xie, L.; Xu, X.; Zhou, Y.; Pan, S. Interactions and gel strength of mixed myofibrillar with soy protein, 7S globulin and enzyme-hydrolyzed soy proteins. Eur. Food Res. Technol. 2010, 231, 751–762. [Google Scholar] [CrossRef]
- Liu, H.; Gao, L.; Ren, Y.; Zhao, Q. Chemical interactions and protein conformation changes during silver carp (Hypophthalmichthys molitrix) surimi gel formation. Int. J. Food Prop. 2014, 17, 1702–1713. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.Q.; Luo, S.Z.; Zhong, X.Y.; Cai, J.; Jiang, S.T.; Zheng, Z. Changes in chemical interactions and protein conformation during heat-induced wheat gluten gel formation. Food Chem. 2017, 214, 393–399. [Google Scholar] [CrossRef]
- Feng, J.; Xiong, Y.L. Interaction of myofibrillar and preheated soy proteins. J. Food Sci. 2002, 67, 2851–2856. [Google Scholar] [CrossRef]
- Zheng, M.; Liu, X.; Chuai, P.; Jiang, Z.; Zhu, Y.; Zhang, B.; Ni, H.; Li, Q. Effects of crude fucoidan on physicochemical properties, antioxidation and bacteriostasis of surimi products. Food Control 2021, 122, 107806. [Google Scholar] [CrossRef]
- Moreno, H.M.; Bargiela, V.; Tovar, C.A.; Cando, D.; Borderias, A.J.; Herranz, B. High pressure applied to frozen flying fish (Parexocoetus brachyterus) surimi: Effect on physicochemical and rheological properties of gels. Food Hydrocoll. 2015, 48, 127–134. [Google Scholar] [CrossRef]
- Ahmad, S.; Rizawi, J.A.; Srivastava, P.K. Effect of soy protein isolate incorporation on quality characteristics and shelf-life of buffalo meat emulsion sausage. J. Food Sci. Technol. 2010, 47, 290–294. [Google Scholar] [CrossRef]
- Mozhaev, V.V.; Heremans, K.; Frank, J.; Masson, P.; Balny, C. High pressure effects on protein structure and function. Proteins Struct. Funct. Bioinform. 1996, 24, 81–91. [Google Scholar] [CrossRef]
- Zhuang, X.; Jiang, X.; Zhou, H.; Chen, Y.; Zhao, Y.; Yang, H.; Zhou, G. Insight into the mechanism of physicochemical influence by three polysaccharides on myofibrillar protein gelation. Carbohydr. Polym. 2020, 229, 115449. [Google Scholar] [CrossRef] [PubMed]
- Chaijan, M.; Panpipat, W. Mechanism of oxidation in foods of animal origin. In Natural Antioxidants, 1st ed.; Banerjee, R., Verma, A.K., Siddiqui, M.W., Eds.; Apple Academic Press: Palm Bay, FL, USA, 2017; pp. 1–37. [Google Scholar]
- Phetsang, H.; Panpipat, W.; Undeland, I.; Panya, A.; Phonsatta, N.; Chaijan, M. Comparative quality and volatilomic characterisation of unwashed mince, surimi, and pH-shift-processed protein isolates from farm-raised hybrid catfish (Clarias macrocephalus × Clarias gariepinus). Food Chem. 2021, 364, 130365. [Google Scholar] [CrossRef] [PubMed]
- St. Angelo, A.J.; Vercellotti, J.; Jacks, T.; Legendre, M. Lipid oxidation in foods. Crit. Rev. Food Sci. Nutr. 1996, 36, 175–224. [Google Scholar]
- Hayes, R.E.; Bookwalter, G.N.; Bagley, E.B. Antioxidant activity of soybean flour and derivatives–A review. J. Food Sci. 1977, 42, 1527–1532. [Google Scholar] [CrossRef]
- Peña-Ramos, E.A.; Xiong, Y.L. Whey and soy protein hydrolysates inhibit lipid oxidation in cooked pork patties. Meat Sci. 2003, 64, 259–263. [Google Scholar] [CrossRef]
- Zielińska, E.; Baraniak, B.; Karaś, M. Antioxidant and anti-inflammatory activities of hydrolysates and peptide fractions obtained by enzymatic hydrolysis of selected heat-treated edible insects. Nutrients 2017, 9, 970. [Google Scholar] [CrossRef] [Green Version]
- Mohan, C.O.; Ravishankar, C.N.; Lalitha, K.V.; Gopal, T.S. Effect of chitosan edible coating on the quality of double filleted Indian oil sardine (Sardinella longiceps) during chilled storage. Food Hydrocoll. 2012, 26, 167–174. [Google Scholar] [CrossRef]
- Habinshuti, I.; Chen, X.; Yu, J.; Mukeshimana, O.; Duhoranimana, E.; Karangwa, E.; Muhoza, B.; Zhang, M.; Xia, S.; Zhang, X. Antimicrobial, antioxidant and sensory properties of Maillard reaction products (MRPs) derived from sunflower, soybean and corn meal hydrolysates. LWT 2019, 101, 694–702. [Google Scholar] [CrossRef]
- Mshayisa, V.V.; Van Wyk, J. Hermetia illucens protein conjugated with glucose via Maillard reaction: Antioxidant and techno-functional properties. Int. J. Food Sci. 2021, 2021, 5572554. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Levels (%) | Texture Profile Analysis Parameters | ||||
|---|---|---|---|---|---|---|
| Hardness (N) | Springiness (mm) | Cohesiveness | Chewiness (N) | Gumminess (N) | ||
| Control | - | 10.53 ± 0.73 e | 6.68 ± 0.27 d | 0.13 ± 0.01 d | 9.08 ± 0.92 e | 1.42 ± 0.13 e |
| CP | 1 | 10.67 ± 0.72 eB | 6.43 ± 0.08 dB | 0.13 ± 0.00 dB | 9.11 ± 0.88 eB | 1.44 ± 0.11 eB |
| 3 | 12.28 ± 0.94 eB | 6.58 ± 0.18 dB | 0.13 ± 0.02 dB | 10.70 ± 1.48 eB | 1.62 ± 0.18 deB | |
| 5 | 14.63 ± 1.47 dB | 8.48 ± 0.12 cB | 0.14 ± 0.01 cB | 17.70 ± 3.08 dB | 1.87 ± 0.04 dB | |
| SPI | 1 | 44.22 ± 4.91 cA | 10.09 ± 0.02 bA | 0.39 ± 0.01 bA | 172.05 ± 17.09 cA | 17.05 ± 1.80 cA |
| 3 | 52.52 ± 5.00 bA | 10.36 ± 0.07 aA | 0.41 ± 0.01 aA | 223.22 ± 17.17 bA | 21.48 ± 1.75 bA | |
| 5 | 60.86 ± 3.03 aA | 10.32 ± 0.07 aA | 0.41 ± 0.01 aA | 255.32 ± 13.90 aA | 24.80 ± 1.38 aA | |
| Samples | Level (%) | Soluble Protein Contents (g/L) | |||
|---|---|---|---|---|---|
| Ionic Bonds | Hydrogen Bonds | Hydrophobic Interactions | Disulfide Bonds | ||
| Control | - | 0.64 ± 0.01 c | 0.34 ± 0.01 c | 2.44 ± 0.06 e | 1.51 ± 0.02 b |
| CP | 1 | 0.62 ± 0.01 cdB | 0.37 ± 0.03 cA | 2.41 ± 0.01 eB | 1.26 ± 0.07 cA |
| 3 | 0.64 ± 0.04 cA | 0.35 ± 0.04 cB | 2.89 ± 0.06 dB | 1.18 ± 0.05 cA | |
| 5 | 0.80 ± 0.01 bA | 0.47 ± 0.01 bB | 2.84 ± 0.11 dB | 1.77 ± 0.10 aA | |
| SPI | 1 | 0.85 ± 0.01 aA | 0.35 ± 0.01 cA | 4.06 ± 0.06 cA | 0.87 ± 0.07 dB |
| 3 | 0.60 ± 0.03 dA | 0.49 ± 0.02 bA | 5.55 ± 0.15 bA | 0.73 ± 0.10 dB | |
| 5 | 0.50 ± 0.01 eB | 0.56 ± 0.02 aA | 6.88 ± 0.08 aA | 0.36 ± 0.08 eB | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Somjid, P.; Panpipat, W.; Cheong, L.-Z.; Chaijan, M. Comparative Effect of Cricket Protein Powder and Soy Protein Isolate on Gel Properties of Indian Mackerel Surimi. Foods 2022, 11, 3445. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11213445
Somjid P, Panpipat W, Cheong L-Z, Chaijan M. Comparative Effect of Cricket Protein Powder and Soy Protein Isolate on Gel Properties of Indian Mackerel Surimi. Foods. 2022; 11(21):3445. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11213445
Chicago/Turabian StyleSomjid, Panumas, Worawan Panpipat, Ling-Zhi Cheong, and Manat Chaijan. 2022. "Comparative Effect of Cricket Protein Powder and Soy Protein Isolate on Gel Properties of Indian Mackerel Surimi" Foods 11, no. 21: 3445. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11213445