
Screening and Enzymatic Evaluation of Saccharomyces cerevisiae Populations from Spontaneous Fermentation of Organic Verdejo Wines
 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and Yeast Isolation
2.2. Molecular Analysis of the Isolates
2.3. Enzymatic Activities
2.3.1. β-Lyase Activity
2.3.2. β-Glucosidase Activity
2.3.3. β-Glucanase Activity
2.3.4. Protease Activity
2.3.5. Sulfite Reductase Activity
2.3.6. Hydroxycinnamic Acid Decarboxylase (HCDC) Activity
2.3.7. Amino Acid Decarboxylation Activity
2.4. Statistical Analysis
3. Results and Discussion
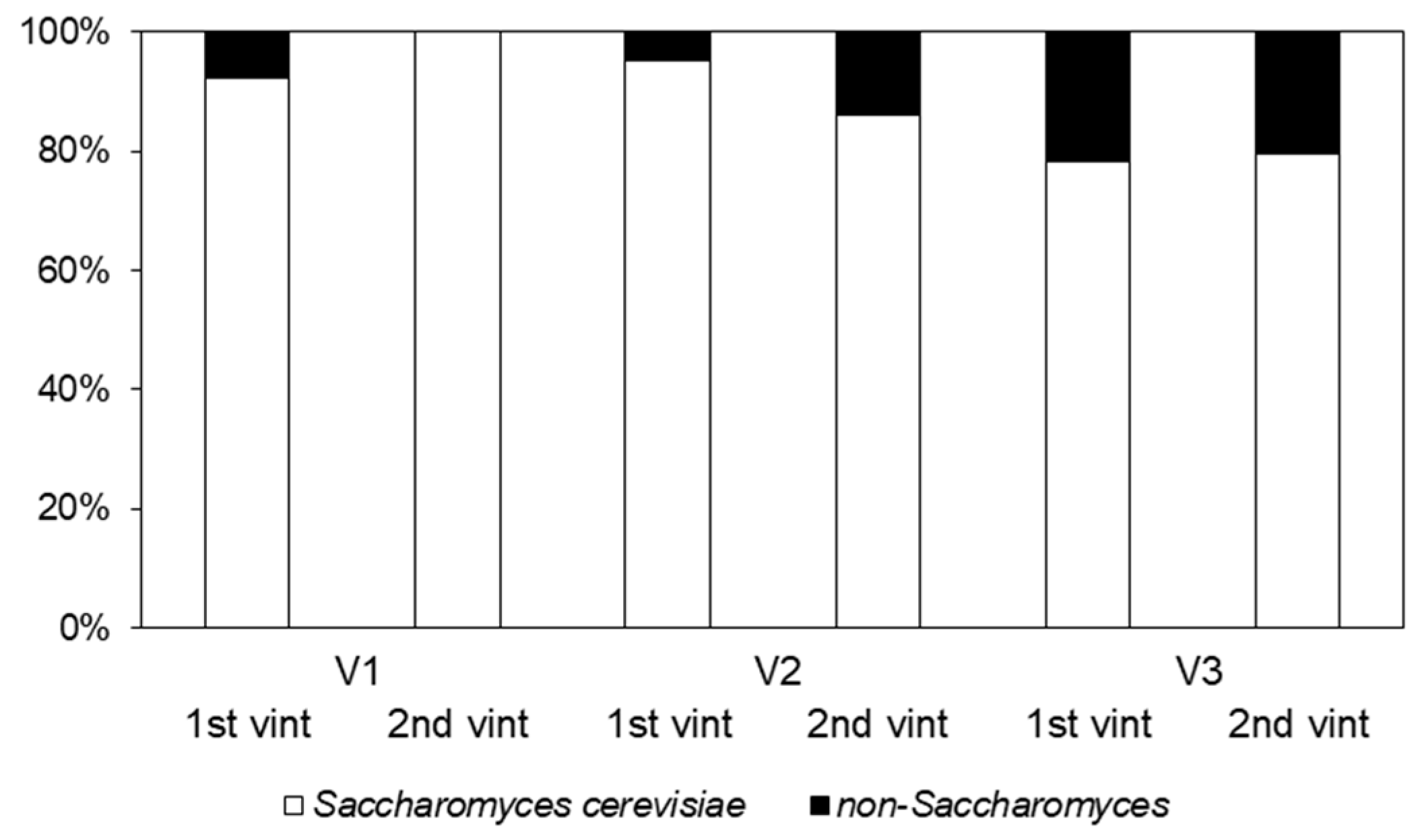
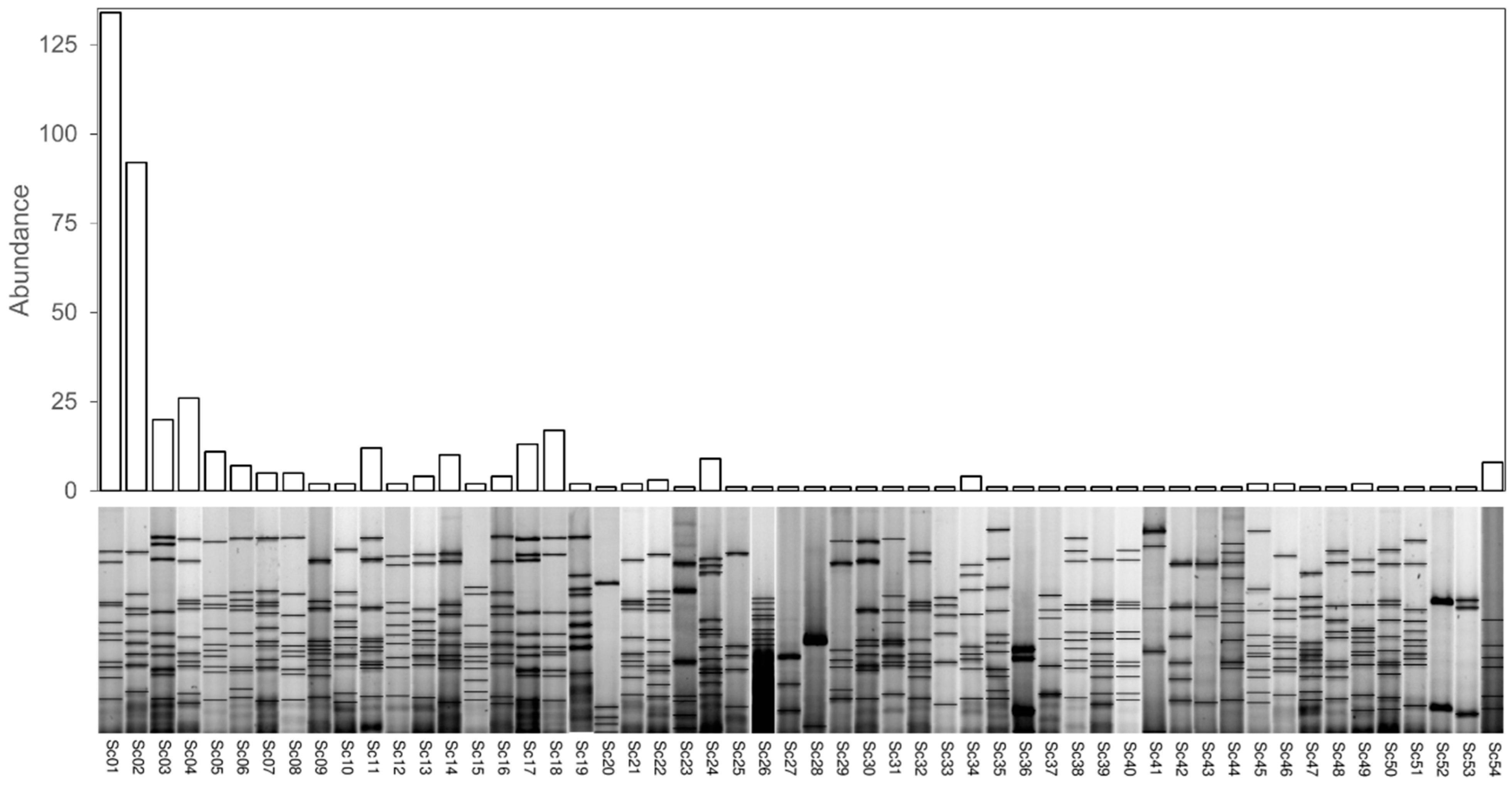
3.1. Identification and Genetic Classification of Yeasts
3.2. Analysis of Diversity of S. cerevisiae Population
3.3. Spatial and Temporal Concurrence of S. cerevisiae Strains
3.4. Evaluation of the Contribution of Yeast Populations to Wine Quality
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, D.; Legras, J.L.; Zhang, P.Z.; Chen, D.L.; Howell, K. Diversity and Dynamics of Fungi during Spontaneous Fermentations and Association with Unique Aroma Profiles in Wine. Int. J. Food Microbiol. 2021, 338, 11. [Google Scholar] [CrossRef] [PubMed]
- Tufariello, M.; Chiriatti, M.A.; Grieco, F.; Perrotta, C.; Capone, S.; Rampino, P.; Tristezza, M.; Mita, G.; Grieco, F. Influence of Autochthonous Saccharomyces cerevisiae Strains on Volatile of Negroamaro Wines. Lwt-Food Sci. Technol. 2014, 58, 35–48. [Google Scholar] [CrossRef]
- Varela, C.; Borneman, A.R. Yeasts Found in Vineyards and Wineries. Yeast 2017, 34, 111–128. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Zhang, P.Z.; Chen, D.L.; Howell, K. From the Vineyard to the Winery: How Microbial Ecology Drives Regional Distinctiveness of Wine. Front. Microbiol. 2019, 10, 13. [Google Scholar] [CrossRef] [PubMed]
- Castrillo, D.; Neira, N.; Blanco, P. Saccharomyces cerevisiae Strain Diversity Associated with Spontaneous Fermentations in Organic Wineries from Galicia (NW Spain). Fermentation 2020, 6, 89. [Google Scholar] [CrossRef]
- Berbegal, C.; Spano, G.; Tristezza, M.; Grieco, F.; Capozzi, V. Microbial Resources and Innovation in the Wine Production Sector. S. Afr. J. Enol. Vitic. 2017, 38, 156–166. [Google Scholar] [CrossRef]
- Nisiotou, A.; Mallouchos, A.; Tassou, C.; Banilas, G. Indigenous Yeast Interactions in Dual-Starter Fermentations May Improve the Varietal Expression of Moschofilero Wine. Front. Microbiol. 2019, 10, 1712. [Google Scholar] [CrossRef] [Green Version]
- Borren, E.; Tian, B. The Important Contribution of Non- Saccharomyces Yeasts to the Aroma Complexity of Wine: A Review. Foods 2021, 10, 13. [Google Scholar] [CrossRef]
- Drumonde-Neves, J.; Franco-Duarte, R.; Vieira, E.; Mendes, I.; Lima, T.; Schuller, D.; Pais, C. Differentiation of Saccharomyces Cerevisiae Populations from Vineyards of the Azores Archipelago: Geography vs Ecology. Food Microbiol. 2018, 74, 151–162. [Google Scholar] [CrossRef]
- Lappa, I.K.; Kachrimanidou, V.; Pateraki, C.; Koulougliotis, D.; Eriotou, E.; Kopsahelis, N. Indigenous Yeasts: Emerging Trends and Challenges in Winemaking. Curr. Opin. Food Sci. 2020, 32, 133–143. [Google Scholar] [CrossRef]
- Aponte, M.; Blaiotta, G. Selection of an Autochthonous Saccharomyces cerevisiae Strain for the Vinification of “Moscato Di Saracena”, a Southern Italy (Calabria Region) Passito Wine. Food Microbiol. 2016, 54, 30–39. [Google Scholar] [CrossRef]
- Belda, I.; Ruiz, J.; Navascues, E.; Marquina, D.; Santos, A. Improvement of Aromatic Thiol Release through the Selection of Yeasts with Increased Beta-Lyase Activity. Int. J. Food Microbiol. 2016, 225, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Escribano, R.; González-Arenzana, L.; Garijo, P.; Berlanas, C.; López Alfaro, I.; López, R.; Gutiérrez, A.R.; Santamaría, P. Screening of Enzymatic Activities within Different Enological Non-Saccharomyces Yeasts. J. Food Sci. Technol. 2017, 54, 1555–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikolaou, E.; Soufleros, E.H.; Bouloumpasi, E.; Tzanetakis, N. Selection of Indigenous Saccharomyces Cerevisiae Strains According to Their Oenological Characteristics and Vinification Results. Food Microbiol. 2006, 23, 205–211. [Google Scholar] [CrossRef]
- Tristezza, M.; Vetrano, C.; Bleve, G.; Grieco, F.; Tufariello, M.; Quarta, A.; Mita, G.; Spano, G. Autochthonous Fermentation Starters for the Industrial Production of Negroamaro Wines. J. Ind. Microbiol. Biotechnol. 2012, 39, 81–92. [Google Scholar] [CrossRef]
- Caruso, M.; Fiore, C.; Contursi, M.; Salzano, G.; Paparella, A.; Romano, P. Formation of Biogenic Amines as Criteria for the Selection of Wine Yeasts. World J. Microbiol. Biotechnol. 2002, 18, 159–163. [Google Scholar] [CrossRef]
- Guo, Y.Y.; Yang, Y.P.; Peng, Q.; Han, Y. Biogenic Amines in Wine: A Review. Int. J. Food Sci. Technol. 2015, 50, 1523–1532. [Google Scholar] [CrossRef]
- Restuccia, D.; Loizzo, M.; Spizzirri, U. Accumulation of Biogenic Amines in Wine: Role of Alcoholic and Malolactic Fermentation. Fermentation 2018, 4, 6. [Google Scholar] [CrossRef] [Green Version]
- Rodríguez-Nogales, J.; Fernández-Fernández, E.; Vila-Crespo, J. Characterisation and Classification of Spanish Verdejo Young White Wines by Volatile and Sensory Analysis with Chemometric Tools. J. Sci. Food Agric. 2009, 89, 1927–1935. [Google Scholar] [CrossRef]
- Querol, A.; Barrio, E.; Huerta, T.; Ramon, D. Molecular Monitoring of Wine Fermentations Conducted by Active Dry Yeast Strains. Appl. Environ. Microbiol. 1992, 58, 2948–2953. [Google Scholar] [CrossRef]
- Esteve-Zarzoso, B.; Belloch, C.; Uruburu, F.; Querol, A. Identification of Yeasts by RFLP Analysis of the 5.8S RRNA Gene and the Two Ribosomal Internal Transcribed Spacers. Int. J. Syst. Bacteriol. 1999, 49, 329–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Maio, S.; Polizzotto, G.; Di Gangi, E.; Foresta, G.; Genna, G.; Verzera, A.; Scacco, A.; Amore, G.; Oliva, D. Biodiversity of Indigenous Saccharomyces Populations from Old Wineries of South-Eastern Sicily (Italy): Preservation and Economic Potential. PLoS ONE 2012, 7, e30428. [Google Scholar] [CrossRef]
- Belda, I.; Ruiz, J.; Alastruey-Izquierdo, A.; Navascués, E.; Marquina, D.; Santos, A. Unraveling the Enzymatic Basis of Wine “Flavorome”: A Phylo-Functional Study of Wine Related Yeast Species. Front. Microbiol. 2016, 7, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suranská, H.; Vranova, D.; Omelkova, J. Isolation, Identification and Characterization of Regional Indigenous Saccharomyces cerevisiae Strains. Brazilian J. Microbiol. 2016, 47, 181–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Champaign, IL, USA, 1949. [Google Scholar]
- Paliy, O.; Shankar, V. Application of Multivariate Statistical Techniques in Microbial Ecology. Mol. Ecol. 2016, 25, 1032–1057. [Google Scholar] [CrossRef] [Green Version]
- Ramette, A. Multivariate Analyses in Microbial Ecology. Fems Microbiol. Ecol. 2007, 62, 142–160. [Google Scholar] [CrossRef] [Green Version]
- Hammer, O.; Harper, D.; Ryan, P. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Querol, A.; Barrio, E.; Ramón, D. A Comparative Study of Different Methods of Yeast Strain Characterization. Syst. Appl. Microbiol. 1992, 15, 439–446. [Google Scholar] [CrossRef]
- López, V.; Querol, A.; Ramón, D.; Fernández-Espinar, M.T.T. A Simplified Procedure to Analyse Mitochondrial DNA from Industrial Yeasts. Int. J. Food Microbiol. 2001, 68, 75–81. [Google Scholar] [CrossRef]
- Fernandez-Espinar, M.T.; Llopis, S.; Querol, A.; Barrio, E. Molecular Identification and Characterization of Wine Yeasts. In Molecular Wine Microbiology; Carrascosa, A.V., Munoz, R., Gonzalez, R., Eds.; Elsevier Academic Press Inc.: San Diego, CA, USA, 2011; pp. 111–141. ISBN1 978-0-08-096258-0. ISBN2 978-0-12-375021-1. [Google Scholar]
- Alonso, A.; de Celis, M.; Ruiz, J.; Vicente, J.; Navascues, E.; Acedo, A.; Ortiz-Alvarez, R.; Belda, I.; Santos, A.; Gomez-Flechoso, M.A.; et al. Looking at the Origin: Some Insights into the General and Fermentative Microbiota of Vineyard Soils. Fermentation 2019, 5, 78. [Google Scholar] [CrossRef] [Green Version]
- Kioroglou, D.; Kraeva-Deloire, E.; Schmidtke, L.M.; Mas, A.; Portillo, M.C. Geographical Origin Has a Greater Impact on Grape Berry Fungal Community than Grape Variety and Maturation State. Microorganisms 2019, 7, 669. [Google Scholar] [CrossRef] [PubMed]
- Knight, S.J.; Karon, O.; Goddard, M.R. Small Scale Fungal Community Differentiation in a Vineyard System. Food Microbiol. 2020, 87, 8. [Google Scholar] [CrossRef] [PubMed]
- Ribéreau-Gayon, P.; Darriet, P.; Towey, J. Handbook of Enology. Volume 1, The Microbiology of Wine and Vinifications, 3rd ed.; John Wiley: Chichester, UK, 2021; ISBN 9781119584681. [Google Scholar]
- Granchi, L.; Ganucci, D.; Buscioni, G.; Mangani, S.; Guerrini, S. The Biodiversity of Saccharomyces cerevisiae in Spontaneous Wine Fermentation: The Occurrence and Persistence of Winery-Strains. Fermentation 2019, 5, 86. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Martín, F.; Seseña, S.; Fernández-González, M.; Arévalo, M.; Palop, M.L. Microbial Communities in Air and Wine of a Winery at Two Consecutive Vintages. Int. J. Food Microbiol. 2014, 190, 44–53. [Google Scholar] [CrossRef]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not Your Ordinary Yeast: Non-Saccharomyces Yeasts in Wine Production Uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Ciani, M.; Capece, A.; Comitini, F.; Canonico, L.; Siesto, G.; Romano, P. Yeast Interactions in Inoculated Wine Fermentation. Front. Microbiol. 2016, 7, 555. [Google Scholar] [CrossRef] [Green Version]
- Fleet, G.H. Yeast Interactions and Wine Flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Harlé, O.; Legrand, J.; Tesnière, C.; Pradal, M.; Mouret, J.R.; Nidelet, T. Investigations of the Mechanisms of Interactions between Four Non-Conventional Species with Saccharomyces cerevisiae in Oenological Conditions. PLoS ONE 2020, 15, e0233285. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermentation Stages | Vineyard 1 | Vineyard 2 | Vineyard 3 | Total | |||
|---|---|---|---|---|---|---|---|
| 1st vint | 2nd vint | 1st vint | 2nd vint | 1st vint | 2nd vint | ||
| CM | 13 | 14 | 16 | 15 | 16 | 12 | 86 |
| RM | 15 | 17 | 16 | 17 | 16 | 14 | 95 |
| SF | 16 | 17 | 17 | 16 | 17 | 15 | 98 |
| TF | 17 | 17 | 16 | 17 | 17 | 15 | 99 |
| EF | 17 | 17 | 17 | 21 | 17 | 17 | 106 |
| Total | 78 | 82 | 82 | 86 | 83 | 73 | 484 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Enríquez, L.; Vila-Crespo, J.; Rodríguez-Nogales, J.M.; Fernández-Fernández, E.; Ruipérez, V. Screening and Enzymatic Evaluation of Saccharomyces cerevisiae Populations from Spontaneous Fermentation of Organic Verdejo Wines. Foods 2022, 11, 3448. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11213448
López-Enríquez L, Vila-Crespo J, Rodríguez-Nogales JM, Fernández-Fernández E, Ruipérez V. Screening and Enzymatic Evaluation of Saccharomyces cerevisiae Populations from Spontaneous Fermentation of Organic Verdejo Wines. Foods. 2022; 11(21):3448. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11213448
Chicago/Turabian StyleLópez-Enríquez, Lorena, Josefina Vila-Crespo, José Manuel Rodríguez-Nogales, Encarnación Fernández-Fernández, and Violeta Ruipérez. 2022. "Screening and Enzymatic Evaluation of Saccharomyces cerevisiae Populations from Spontaneous Fermentation of Organic Verdejo Wines" Foods 11, no. 21: 3448. https://0-doi-org.brum.beds.ac.uk/10.3390/foods11213448