Evaluation of Amyloid β42 Aggregation Inhibitory Activity of Commercial Dressings by A Microliter-Scale High-Throughput Screening System Using Quantum-Dot Nanoprobes

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
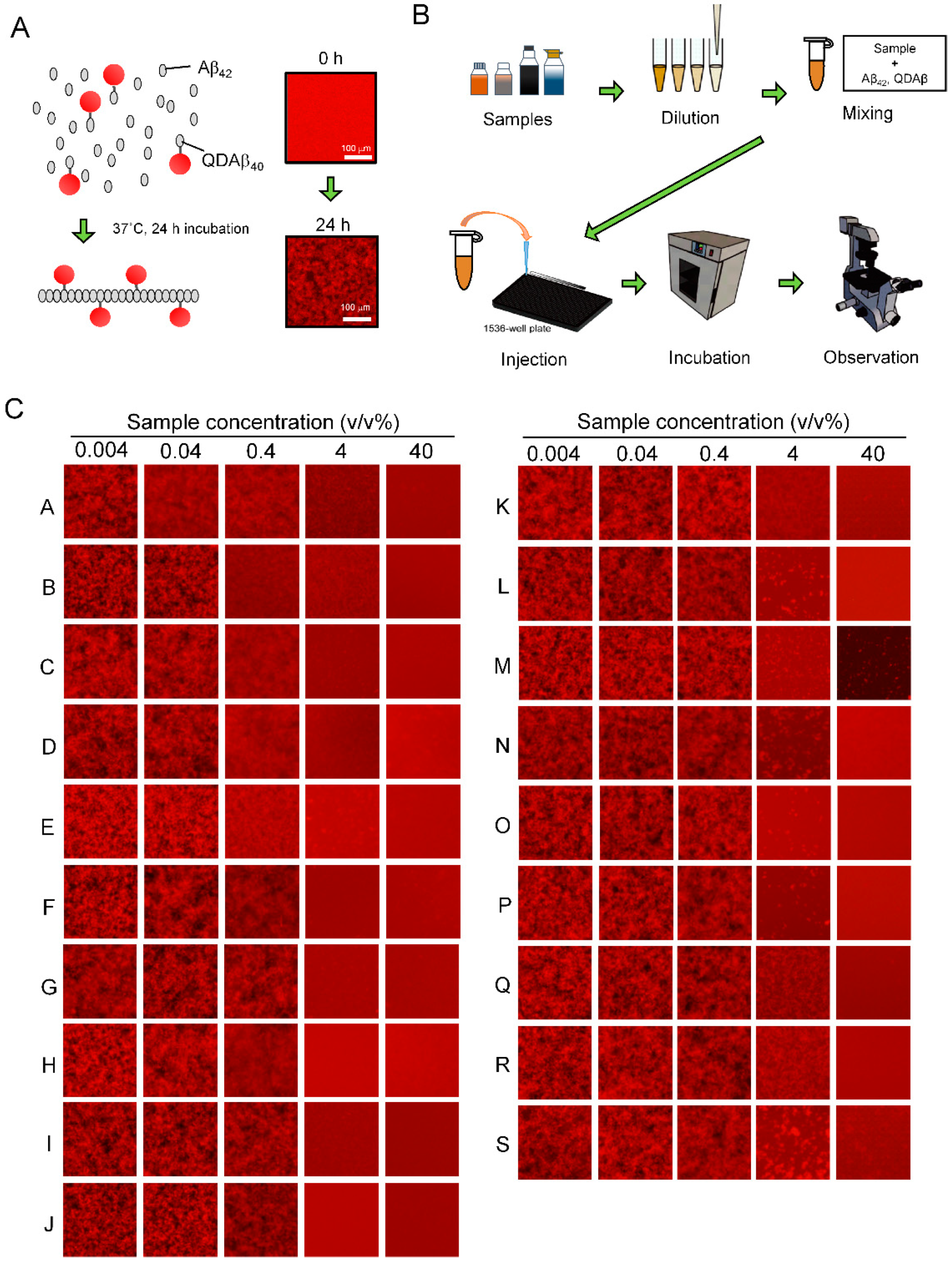
2.2. Preparation of QDAβ Nanoprobe
2.3. Estimation of EC50 by the MSHTS System
2.4. Fluorescence Microscopy
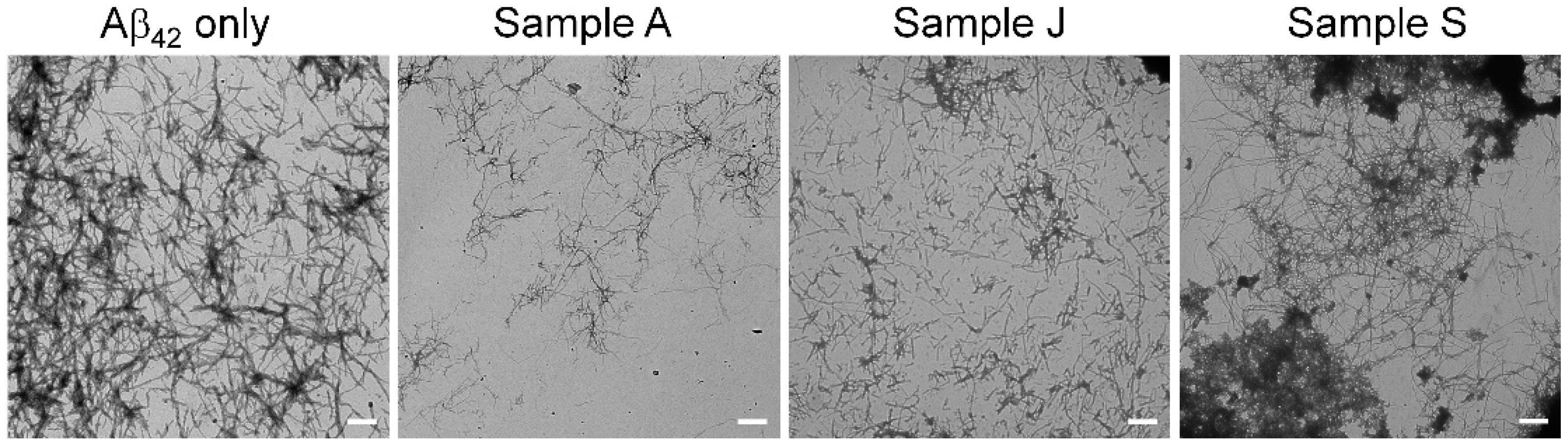
2.5. Transmission Electron Microscopy
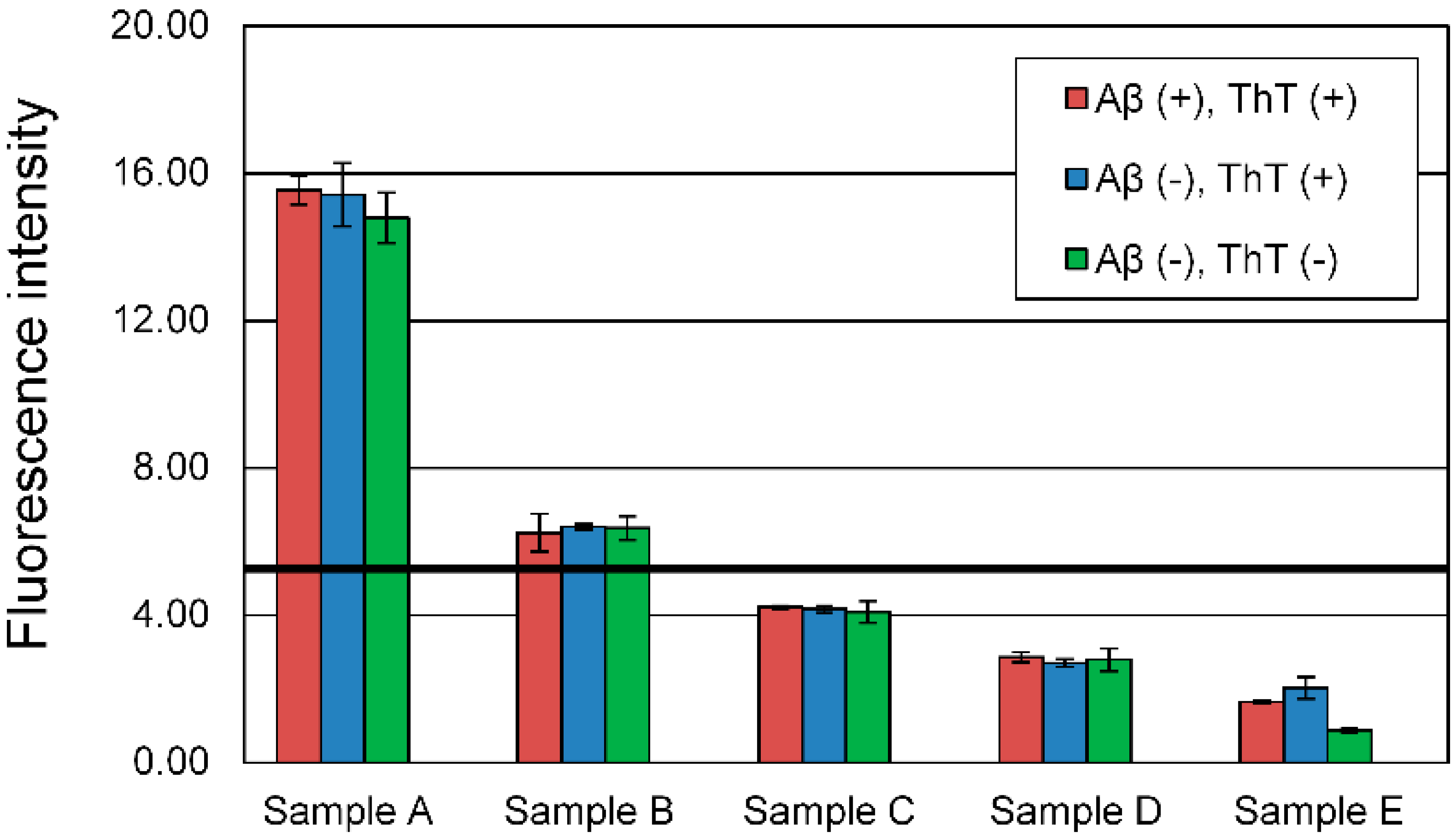
2.6. ThT Assay
2.7. SDS-Polyacrylamide Gel Electrophoresis
3. Results and Discussion
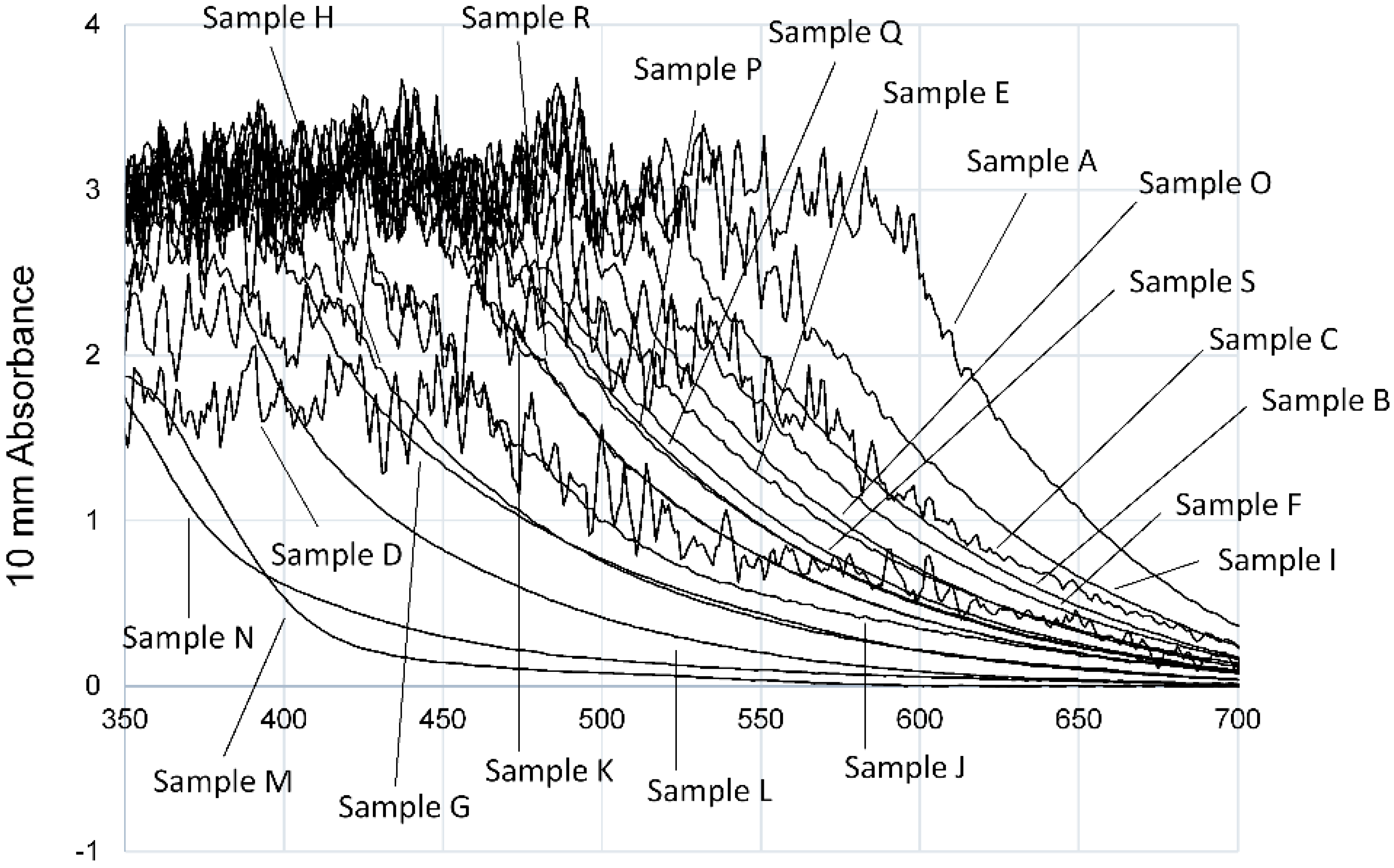
3.1. Evaluation of Aβ42 Aggregation Inhibitory Activity of Commercial Dressings by ThT Method
3.2. Evaluation of Aβ42 Aggregation Inhibitory Activity of Commercial Dressings Using MSHTS Sysytem
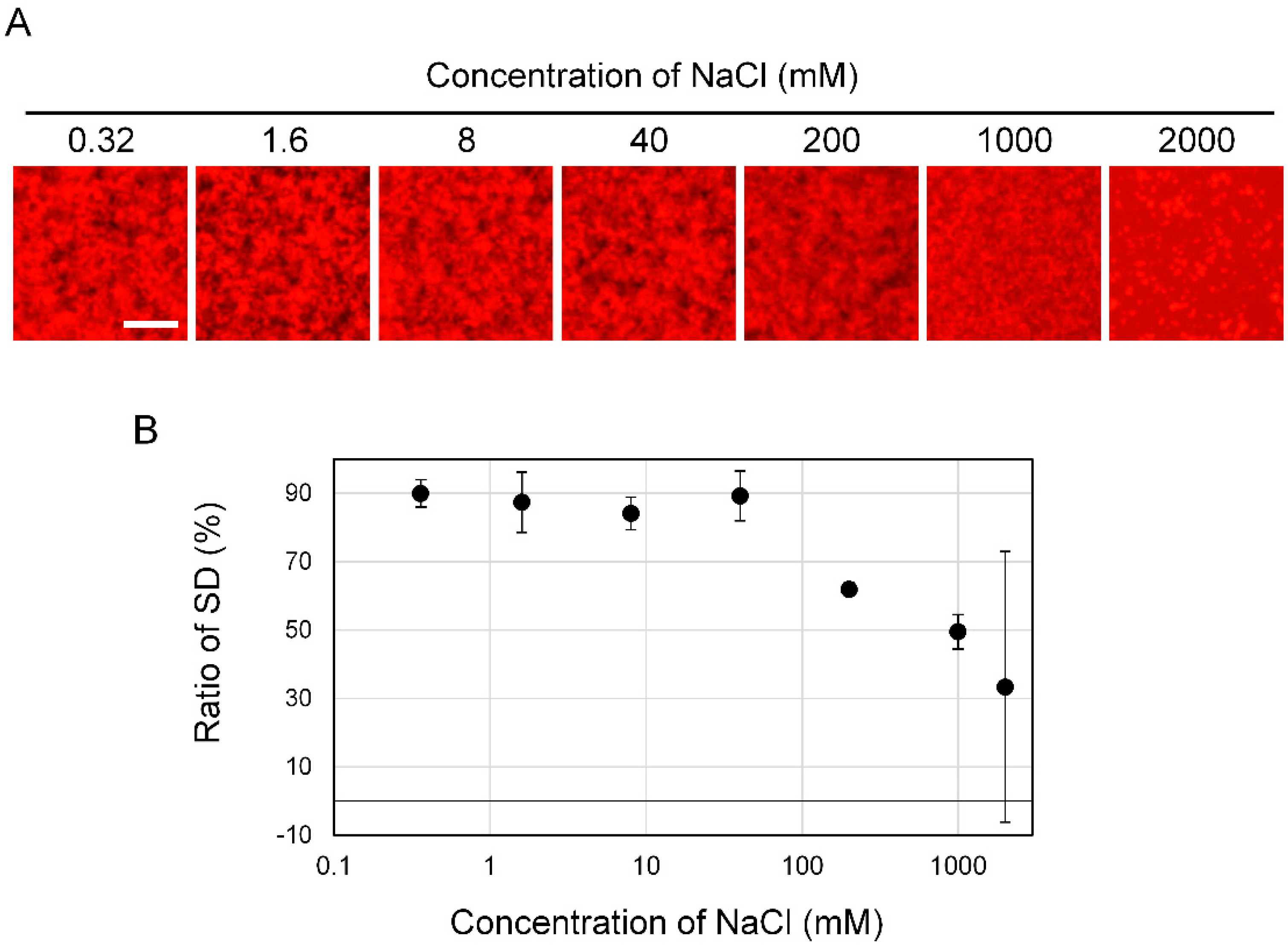
3.3. Effect of Salt Concentration on Aβ42 Aggregation
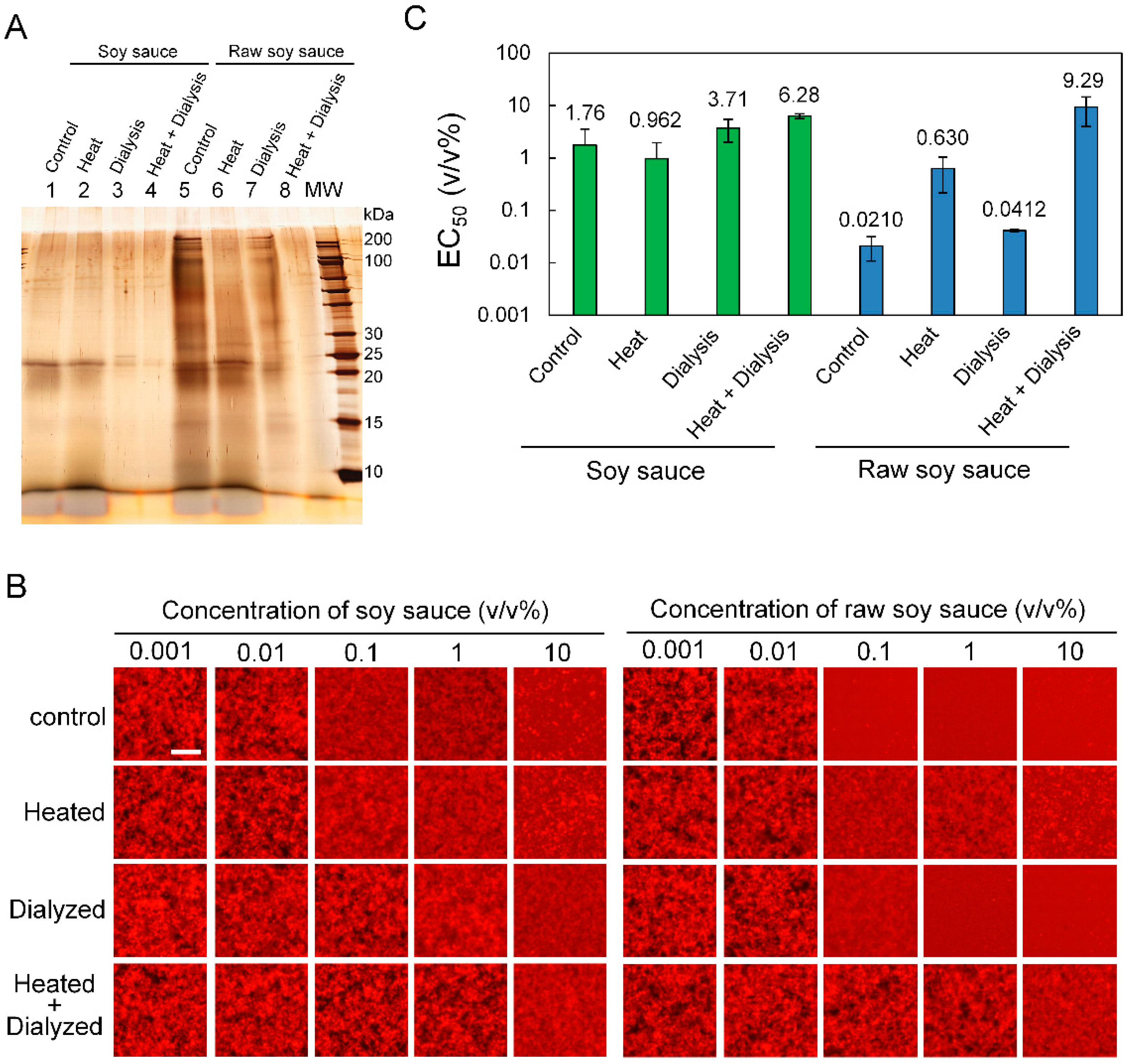
3.4. Influence of Heating and Dialysis Treatment on Aβ42 Aggregation Inhibitory Activity
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Aβ | amyloid β |
| MSHTS system | microliter-scale high-throughput screening system |
| PBS | phosphate buffered saline |
| QD | quantum dot |
| RA | rosmarinic acid |
| SD value | standard deviation value |
| SDS-PAGE | SDS-polyacrylamide gel electrophoresis |
| TEM | transmission electron microscope |
| ThT | thioflavin T |
References
- Ikejima, C.; Hisanaga, A.; Meguro, K.; Yamada, T.; Ouma, S.; Kawamuro, Y.; Hyouki, K.; Nakashima, K.; Wada, K.; Yamada, S.; et al. Multicentre population-based dementia prevalence survey in Japan: A preliminary report. Psychogeriatrics 2012, 12, 120–123. [Google Scholar] [CrossRef] [PubMed]
- Plassman, B.L.; Langa, K.M.; Fisher, G.G.; Heeringa, S.G.; Weir, D.R.; Ofstedal, M.B.; Burke, J.R.; Hurd, M.D.; Potter, G.G.; Rodgers, W.L.; et al. Prevalence of Dementia in the United States: The Aging, Demographics, and Memory Study. Neuroepidemiology 2007, 29, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Lane, C.A.; Hardy, J.; Schott, J.M. Alzheimer’s disease. Eur. J. Neurol. 2018, 25, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, A. A review on Alzheimer’s disease pathophysiology and its management: An update. Pharmacol. Rep. 2015, 67, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J. The Amyloid Hypothesis of Alzheimer’s Disease: Progress and Problems on the Road to Therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [Green Version]
- Koo, E.H.; Lansbury, P.T.; Kelly, J.W. Amyloid diseases: Abnormal protein aggregation in neurodegeneration. Proc. Natl. Acad. Sci. USA 1999, 96, 9989–9990. [Google Scholar] [CrossRef] [Green Version]
- Pepys, M.B. Amyloidosis. Annu. Rev. Med. 2006, 57, 223–241. [Google Scholar] [CrossRef]
- Selkoe, D.J.; Hardy, J. The amyloid hypothesis of Alzheimer’s disease at 25 years. EMBO Mol. Med. 2016, 8, 595–608. [Google Scholar] [CrossRef]
- Sevigny, J.; Chiao, P.; Bussière, T.; Weinreb, P.H.; Williams, L.; Maier, M.; Dunstan, R.; Salloway, S.; Chen, T.; Ling, Y.; et al. The antibody aducanumab reduces Aβ plaques in Alzheimer’s disease. Nature 2016, 537, 50–56. [Google Scholar] [CrossRef]
- Selkoe, D.J. Alzheimer disease and aducanumab: Adjusting our approach. Nat. Rev. Neurol. 2019, 15, 365–366. [Google Scholar] [CrossRef] [PubMed]
- Bateman, R.J.; Xiong, C.; Benzinger, T.L.S.; Fagan, A.M.; Goate, A.; Fox, N.C.; Marcus, D.S.; Cairns, N.J.; Xie, X.; Blazey, T.M.; et al. Clinical and Biomarker Changes in Dominantly Inherited Alzheimer’s Disease. N. Engl. J. Med. 2012, 367, 795–804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, K.; Hasegawa, K.; Naiki, H.; Yamada, M. Curcumin Has Potent Anti-Amyloidogenic Effects for Alzheimer’s β-Amyloid Fibrils In Vitro. J. Neurosci. Res. 2004, 75, 742–750. [Google Scholar] [CrossRef]
- Hase, T.; Shishido, S.; Yamamoto, S.; Yamashita, R.; Nukima, H.; Taira, S.; Toyoda, T.; Abe, K.; Hamaguchi, T.; Ono, K.; et al. Rosmarinic acid suppresses Alzheimer’s disease development by reducing amyloid β aggregation by increasing monoamine secretion. Sci. Rep. 2019, 9, 8711. [Google Scholar] [CrossRef]
- Hamaguchi, T.; Ono, K.; Murase, A.; Yamada, M. Phenolic Compounds Prevent Alzheimer’s Pathology through Different Effects on the Amyloid-β Aggregation Pathway. Am. J. Pathol. 2009, 175, 2557–2565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noguchi-Shinohara, M.; Ono, K.; Hamaguchi, T.; Iwasa, K.; Nagai, T.; Kobayashi, S.; Nakamura, H.; Yamada, M. Pharmacokinetics, Safety and Tolerability of Melissa officinalis Extract which Contained Rosmarinic Acid in Healthy Individuals: A Randomized Controlled Trial. PLoS ONE 2015, 10, e0126422. [Google Scholar] [CrossRef] [PubMed]
- Momoyo, K.; Aiko, S.; Ryuichiro, S. Neuroprotective Action of Miso. New Food Ind. 2018, 60, 79–83. [Google Scholar] [CrossRef]
- Hsu, R.-L.; Lee, K.-T.; Wang, J.-H.; Lee, L.Y.L.; Chen, R.P.Y. Amyloid-Degrading Ability of Nattokinase from Bacillus subtilis Natto. J. Agric. Food Chem. 2009, 57, 503–508. [Google Scholar] [CrossRef]
- LeVine III, H. Thioflavin T interaction with synthetic Alzhei- mers’sdisease β-amyloid peptides: Detection of amyloid aggre- gation in solution. Prot. Protein Sci. 1993, 404–410. [Google Scholar]
- Tokuraku, K.; Marquardt, M.; Ikezu, T. Real-Time Imaging and Quantification of Amyloid-β Peptide Aggregates by Novel Quantum-Dot Nanoprobes. PLoS ONE 2009, 4, e8492. [Google Scholar] [CrossRef]
- Ishigaki, Y.; Tanaka, H.; Akama, H.; Ogara, T.; Uwai, K.; Tokuraku, K. A Microliter-Scale High-throughput Screening System with Quantum-Dot Nanoprobes for Amyloid-β Aggregation Inhibitors. PLoS ONE 2013, 8, e72992. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, R.; Tainaka, R.; Ando, Y.; Hashi, Y.; Deepak, H.V.; Suga, Y.; Murai, Y.; Anetai, M.; Monde, K.; Ohta, K.; et al. An automated microliter-scale high-throughput screening system (MSHTS) for real-time monitoring of protein aggregation using quantum-dot nanoprobes. Sci. Rep. 2019, 9, 2587. [Google Scholar] [CrossRef]
- Ogara, T.; Takahashi, T.; Yasui, H.; Uwai, K.; Tokuraku, K. Evaluation of the effects of amyloid β aggregation from seaweed extracts by a microliter-scale high-throughput screening system with a quantum dot nanoprobe. J. Biosci. Bioeng. 2015, 120, 45–50. [Google Scholar] [CrossRef]
- Kanda, Y. Investigation of the freely available easy-to-use software ‘EZR’ for medical statistics. Bone Marrow Transplant. 2013, 48, 452–458. [Google Scholar] [CrossRef] [Green Version]
- Kitagawa, M.; Shiraishi, T.; Yamamoto, S.; Kutomi, R.; Ohkoshi, Y.; Sato, T.; Wakui, H.; Itoh, H.; Miyamoto, A.; Yokota, S.-I. Novel antimicrobial activities of a peptide derived from a Japanese soybean fermented food, Natto, against Streptococcus pneumoniae and Bacillus subtilis group strains. AMB Express 2017, 7, 127. [Google Scholar] [CrossRef] [Green Version]
- Lee, Y.-B.; Lee, H.J.; Sohn, H.S. Soy isoflavones and cognitive function. J. Nutr. Biochem. 2005, 16, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Katayama, S.; Imai, R.; Sugiyama, H.; Nakamura, S. Oral Administration of Soy Peptides Suppresses Cognitive Decline by Induction of Neurotrophic Factors in SAMP8 Mice. J. Agric. Food Chem. 2014, 62, 3563–3569. [Google Scholar] [CrossRef] [PubMed]
- Sugano, M.; Yamada, Y.; Yoshida, K.; Hashimoto, Y.; Matsuo, T.; Kimoto, M. The hypocholesterolemic action of the undigested fraction of soybean protein in rats. Atherosclerosis 1988, 72, 115–122. [Google Scholar] [CrossRef]
- Park, Y.-J.; Ko, J.; Jeon, S.; Kwon, Y. Protective Effect of Genistein against Neuronal Degeneration in ApoE−/− Mice Fed a High-Fat Diet. Nutrients 2016, 8, 692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weng, Y.; Yao, J.; Sparks, S.; Wang, K. Nattokinase: An Oral Antithrombotic Agent for the Prevention of Cardiovascular Disease. Int. J. Mol. Sci. 2017, 18, 523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferland, G. Vitamin K and the Nervous System: An Overview of its Actions. Adv. Nutr. 2012, 3, 204–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsukamoto, Y.; Kasai, M.; Kakuda, H. Construction of a Bacillus subtilis (natto) with high productivity of vitamin K2 (menaquinone-7) by analog resistance. Biosci. Biotechnol. Biochem. 2001, 65, 2007–2015. [Google Scholar] [CrossRef] [PubMed]
- Schwalfenberg, G.K. Vitamins K1 and K2: The Emerging Group of Vitamins Required for Human Health. J. Nutr. Metab. 2017, 2017, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Galaqin, N.; Tainaka, R.; Shimamori, K.; Kuragano, M.; Noguchi, T.Q.P.; Tokuraku, K. Real-Time 3D Imaging and Inhibition Analysis of Various Amyloid Aggregations Using Quantum Dots. Int. J. Mol. Sci. 2020, 21, 1978. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | EC50 (v/v%) | Oil Type |
|---|---|---|
| RA (positive control) | 0.122 ± 0.034 (w/v%) | - |
| A | 0.065 ± 0.020 | Non-oil |
| B | 0.094 ± 0.017 | Non-oil |
| C | 0.133 ± 0.021 | Non-oil |
| D | 0.227 ± 0.026 | Oil |
| E | 0.230 ± 0.026 | Non-oil |
| F | 0.334 ± 0.075 | Non-oil |
| G | 0.395 ± 0.130 | Non-oil |
| H | 0.413 ± 0.084 | Non-oil |
| I | 0.480 ± 0.101 | Non-oil |
| J | 0.508 ± 0.025 | Non-oil |
| K | 0.763 ± 0.607 | Oil |
| L | 1.350 ± 0.247 | Non-oil |
| M | 1.360 ± 0.590 | Non-oil |
| N | 2.067 ± 0.728 | Oil |
| O | 2.132 ± 1.473 | Non-oil |
| P | 2.150 ± 0.887 | Non-oil |
| Q | 2.313 ± 0.490 | Non-oil |
| R | 2.580 ± 0.173 | Non-oil |
| S | 6.737 ± 5.054 | Non-oil |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuragano, M.; Yoshinari, W.; Lin, X.; Shimamori, K.; Uwai, K.; Tokuraku, K. Evaluation of Amyloid β42 Aggregation Inhibitory Activity of Commercial Dressings by A Microliter-Scale High-Throughput Screening System Using Quantum-Dot Nanoprobes. Foods 2020, 9, 825. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9060825
Kuragano M, Yoshinari W, Lin X, Shimamori K, Uwai K, Tokuraku K. Evaluation of Amyloid β42 Aggregation Inhibitory Activity of Commercial Dressings by A Microliter-Scale High-Throughput Screening System Using Quantum-Dot Nanoprobes. Foods. 2020; 9(6):825. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9060825
Chicago/Turabian StyleKuragano, Masahiro, Wataru Yoshinari, Xuguang Lin, Keiya Shimamori, Koji Uwai, and Kiyotaka Tokuraku. 2020. "Evaluation of Amyloid β42 Aggregation Inhibitory Activity of Commercial Dressings by A Microliter-Scale High-Throughput Screening System Using Quantum-Dot Nanoprobes" Foods 9, no. 6: 825. https://0-doi-org.brum.beds.ac.uk/10.3390/foods9060825