Evidence for Anti-Inflammatory Activity of Isoliquiritigenin, 18β Glycyrrhetinic Acid, Ursolic Acid, and the Traditional Chinese Medicine Plants Glycyrrhiza glabra and Eriobotrya japonica, at the Molecular Level



Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Chemicals
2.2. Cell Culture and Cell Viability Assay with MTT
2.3. Griess Assay
2.4. NF-κB-Related Gene Expression Analysis by Real-Time PCR
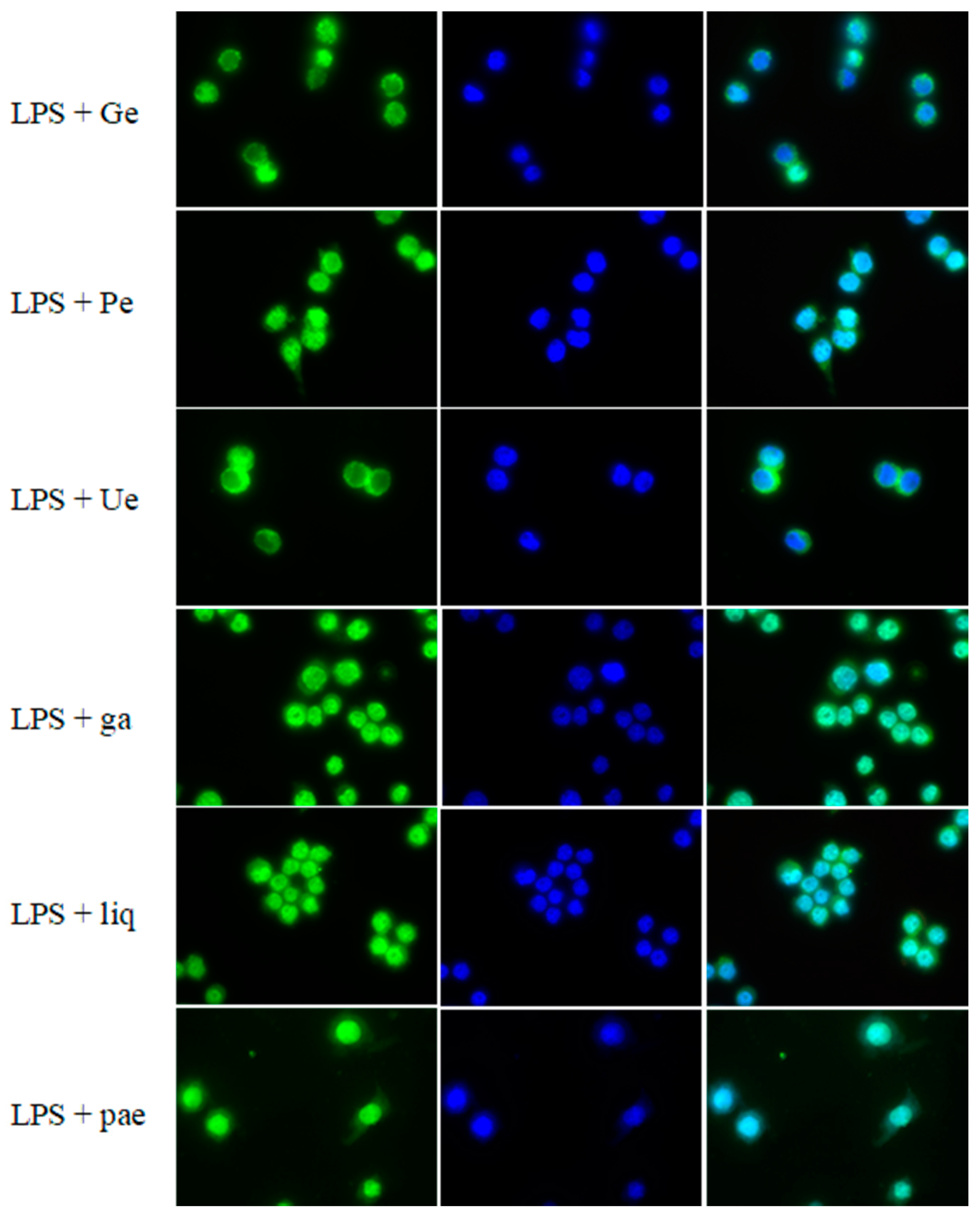
2.5. Imaging of NF-κB Nuclear Translocation
2.6. Statistical Analysis
3. Results
3.1. Cytotoxicity of Individual Substances in RAW 264.7 Cells
3.2. Nitrite Level after LPS Stimulation in RAW 264.7 Cells
3.3. Effect of Single Substances on Nitrite Production in RAW 264.7 Cells
3.4. Effect of Single Substances on the Gene Expressions of NF-κB Subunits and Pro-Inflammatory Factors in RAW 264.7 Cells
3.5. Effect of Single Substances on the Nuclear Translocation of NF-κB
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hayden, M.S.; Ghosh, S. Signaling to NF-κB. Genes Dev. 2004, 18, 2195–2224. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; May, M.J.; Kopp, E.B. NF-κB and rel proteins: Evolutionarily conserved mediators of immune responses. Ann. Rev. Immunol. 1998, 16, 225–260. [Google Scholar] [CrossRef] [PubMed]
- Pahl, H. Activators and target genes of Rel/NF-κB transcription factors. Oncogene 1999, 18, 6853–6866. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baeuerle, P.A.; Henkel, T. Function and activation of NF-κB in the immune system. Ann. Rev. Immunol. 1994, 12, 141–179. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Wu, J.; Wang, J. A database and functional annotation of NF-κB target genes. Int. J. Clin. Exp. Med. 2016, 9, 7986–7995. [Google Scholar]
- Kumar, A.; Takada, Y.; Boriek, A.M.; Aggarwal, B.B. Nuclear factor-kappaB: Its role in health and disease. J. Mol. Med. 2004, 82, 434–448. [Google Scholar] [CrossRef]
- Collart, M.A.; Baeuerle, P.; Vassalli, P. Regulation of tumor necrosis factor alpha transcription in macrophages: Involvement of four kappa B-like motifs and of constitutive and inducible forms of NF-kappa B. Mol. Cell. Biol. 1990, 10, 1498–1506. [Google Scholar] [CrossRef] [PubMed]
- Shakhov, A.N.; Collart, M.A.; Vassalli, P.; Nedospasov, S.A.; Jongeneel, C.V. Kappa B-type enhancers are involved in lipopolysaccharide-mediated transcriptional activation of the tumor necrosis factor alpha gene in primary macrophages. J. Exp. Med. 1990, 171, 35–47. [Google Scholar] [CrossRef] [Green Version]
- Popa, C.; Netea, M.G.; van Riel, P.L.; van der Meer, J.W.; Stalenhoef, A.F. The role of TNF-alpha in chronic inflammatory conditions, intermediary metabolism, and cardiovascular risk. J. Lipid Res. 2007, 48, 751–762. [Google Scholar] [CrossRef]
- Parameswaran, N.; Patial, S. Tumor Necrosis Factor-α signaling in macrophages. Crit. Rev. Eukaryot. Gene Expr. 2010, 20, 87–103. [Google Scholar] [CrossRef]
- O’Neill, G.P.; Ford-Hutchinson, A.W. Expression of mRNA for cyclooxygenase-1 and cyclooxygenase-2 in human tissues. FEBS Lett. 1993, 330, 156–160. [Google Scholar] [CrossRef]
- Yamamoto, K.; Arakawa, T.; Ueda, N.; Yamamoto, S. Transcriptional roles of nuclear factor kappa B and nuclear factor-interleukin-6 in the tumor necrosis factor alpha-dependent induction of cyclooxygenase-2 in MC3T3-E1 cells. J. Biol. Chem. 1995, 270, 31315–31320. [Google Scholar] [CrossRef]
- Vane, J.R.; Bakhle, Y.S.; Botting, R.M. Cyclooxygenases 1 and 2. Ann. Rev. Pharmacol. Toxicol. 1998, 38, 97–120. [Google Scholar] [CrossRef]
- Kaltschmidt, B.; Linker, R.A.; Deng, J.; Kaltschmidt, C. Cyclooxygenase-2 is a neuronal target gene of NF-κB. BMC Mol. Biol. 2002, 3, 16. [Google Scholar] [CrossRef]
- Ackerman IV, W.E.; Summerfield, T.L.S.; Vandre, D.D.; Robinson, J.M.; Kniss, D.A. Nuclear Factor-kappa B regulates inducible prostaglandin E Synthase expression in human amnion mesenchymal cells. Biol. Reprod. 2008, 78, 68–76. [Google Scholar] [CrossRef]
- Harris, S.G.; Padilla, J.; Koumas, L.; Ray, D.; Phipps, R.P. Prostaglandins as modulators of immunity. Trends Immunol. 2002, 23, 144–150. [Google Scholar] [CrossRef]
- Chandrasekharan, N.V.; Simmons, D.L. The cyclooxygenases. Genome Biol. 2004, 5, 241. [Google Scholar] [CrossRef]
- Imhof, B.A.; Dunon, D. Leukocyte migration and adhesion. Adv. Immunol. 1995, 58, 345–416. [Google Scholar]
- Elgavish, A. Effects of Escherichia coli and E. coli lipopolysaccharides on the function of human ureteral epithelial cells cultured in serum-free medium. Infect Immun. 1993, 61, 3304–3312. [Google Scholar]
- Meager, A. Cytokine regulation of cellular adhesion molecule expression in infammation. Cytokine Growth Factor Rev. 1999, 10, 27–39. [Google Scholar] [CrossRef]
- Golias, C.; Tsoutsi, E.; Matziridis, A.; Makridis, P.; Batistatou, A.; Charalabopoulos, K. Leukocyte and endothelial cell adhesion molecules in inflammation focusing on inflammatory heart disease. In Vivo 2007, 21, 757–769. [Google Scholar]
- Frank, P.G.; Lisanti, M.P. ICAM-1: Role in inflammation and in the regulation of vascular permeability. Am. J. Physiol. Heart Circ. Physiol. 2008, 295, 926–927. [Google Scholar] [CrossRef]
- Hill, B.G.; Dranka, B.P.; Bailey, S.M.; Lancaster, J.R., Jr.; Darley-Usmar, V.M. What part of NO don’t you understand? Some answers to the cardinal questions in nitric oxide biology. J. Biol. Chem. 2010, 285, 19699–19704. [Google Scholar] [CrossRef]
- Nathan, C.; Xie, Q. Regulation of biosynthesis of nitric oxide. J. Biol. Chem. 1994, 269, 13725–13728. [Google Scholar]
- Kleinert, H.; Pautz, A.; Linker, K.; Schwarz, P.M. Regulation of the expression of inducible nitric oxide synthase. Eur. J. Pharmacol. 2004, 500, 255–266. [Google Scholar] [CrossRef]
- Moncada, S.; Palmer, R.M.; Higgs, E.A. Nitric oxide: Physiology, pathophysiology, and pharmacology. Pharmacol. Rev. 1991, 43, 109–142. [Google Scholar]
- Moncada, S.; Higgs, E.A. The L-arginine-nitric oxide pathway. N. Engl. J. Med. 1993, 329, 2002–2012. [Google Scholar] [CrossRef]
- Coleman, J.W. Nitric oxide in immunity and inflammation. Int. Immunopharmacol. 2001, 1, 1397–1406. [Google Scholar] [CrossRef]
- Laskin, J.D.; Heck, D.E.; Laskin, D.L. Multifunctional role of nitric oxide in inflammation. Trends Endocrinol. Metab. 1994, 5, 377–382. [Google Scholar] [CrossRef]
- Wu, C. Nitric oxide and inflammation. Curr. Med. Chem. Anti-Inflamm. Anti-Allergy Agents 2004, 3, 217–222. [Google Scholar] [CrossRef]
- Kopp, E.; Ghosh, S. Inhibition of NF-κB by sodium salicylate and aspirin. Science 1994, 265, 956–959. [Google Scholar] [CrossRef]
- Gaddi, A.; Cicero, A.F.G.; Pedro, E.J. Clinical perspectives of anti-inflammatory therapy in the elderly: The lipoxigenase (LOX)/cycloxigenase (COX) inhibition concept. Arch. Gerontol. Geriatr. 2004, 38, 201–212. [Google Scholar] [CrossRef]
- Mendes, R.T.; Stanczyk, C.P.; Sordi, R.; Otuki, M.F.; dos Santos, F.A.; Fernandes, D. Selective inhibition of cyclooxygenase-2: Risks and benefits. Rev. Bras. Reumatol. 2012, 52, 767–782. [Google Scholar] [CrossRef]
- Pushpangadan, P.; Ijinu, T.P.; George, V. Plant based anti-inflammatory secondary metabolites. Ann. Phytomed. 2015, 4, 17–36. [Google Scholar]
- Van Wyk, B.E.; Wink, M. Medicinal Plants of the World, 2nd ed.; Briza: Pretoria, South Africa, 2017. [Google Scholar]
- Van Wyk, B.E.; Wink, M. Phytomedicines, Herbal Drugs and Poisons; University of Chicago Press: Chicago, IL, USA, 2015. [Google Scholar]
- Zhou, J.; Wink, M. Reversal of multidrug resistance in human colon cancer and human leukemia cells by three plant extracts and their major secondary metabolites. Medicines 2018, 5, 123. [Google Scholar] [CrossRef]
- Zhou, J.; Braun, M.S.; Wetterauer, P.; Wetterauer, B.; Wink, M. Antioxidant, cytotoxic, and antimicrobial activities of Glycyrrhiza glabra L., Paeonia lactiflora Pall., and Eriobotrya japonica (Thunb.) Lindl. extracts. Medicines 2019, 6, 43. [Google Scholar] [CrossRef]
- MacMicking, J.; Xie, Q.; Nathan, C. Nitric oxide and macrophage function. Ann. Rev. Immunol. 1997, 15, 323–350. [Google Scholar] [CrossRef]
- Sun, J.; Zhang, X.; Broderick, M.; Fein, H. Measurement of nitric oxide production in biological systems by using griess reaction assay. Sensors 2003, 3, 276–284. [Google Scholar] [CrossRef]
- Rees, D.D.; Palmer, R.M.; Schulz, R.; Hodson, H.F.; Moncada, S. Characterization of three inhibitors of endothelial nitric oxide synthase in vitro and in vivo. Br. J. Pharmacol. 1990, 101, 746–752. [Google Scholar] [CrossRef] [Green Version]
- Borowicz, K.K.; Starownik, R.; Kleinrok, Z.; Czuczwar, S.J. The influence of L-NG-nitroarginine methyl ester, an inhibitor of nitric oxide synthase, upon the anticonvulsive activity of conventional antiepileptic drugs against maximal electroshock in mice. J. Neural Transm. 1998, 105, 1–12. [Google Scholar] [CrossRef]
- Rawls, S.M.; Tallarida, R.J.; Gray, A.M.; Geller, E.B.; Adler, M.W. L-NAME (N-Nitro-L-Arginine Methyl Ester), a nitric-oxide synthase inhibitor, and WIN 55212-2 [4,5-dihydro-2-methyl-4(4-morpholinylmethyl)-1-(1-naphthalenyl-carbonyl)-6H-pyrrolo[3,2,1ij]quinolin-6-one], a cannabinoid agonist, interact to evoke synergistic hypothermia. J. Pharmacol. Exp. Ther. 2004, 308, 780–786. [Google Scholar] [CrossRef]
- Beutler, B.; Cerami, A. Cachectin and tumour necrosis factor as two sides of the same biological coin. Nature 1986, 320, 584–588. [Google Scholar] [CrossRef]
- Zhang, G.; Ghosh, S. Molecular mechanisms of NF-κB activation induced by bacterial lipopolysaccharide through Toll-like receptors. J. Endotoxin Res. 2000, 6, 453–457. [Google Scholar] [CrossRef]
- Thiyagarajan, P.; Chandrasekaran, C.V.; Deepak, H.B.; Agarwal, A. Modulation of lipopolysaccharide-induced pro-inflammatory mediators by an extract of Glycyrrhiza glabra and its phytoconstituents. Inflammopharmacology 2011, 19, 235–241. [Google Scholar] [CrossRef]
- Shishodia, S.; Majumdar, S.; Banerjee, S.; Aggarwal, B.B. Ursolic acid inhibits nuclear factor-kappa B activation induced by carcinogenic agents through suppression of Ikappa B alpha kinase and p65 phosphorylation: Correlation with down-regulation of cyclooxygenase 2, matrix metalloproteinase 9, and cyclin D1. Cancer Res. 2003, 63, 4375–4383. [Google Scholar]
- Kim, J.Y.; Park, S.J.; Yun, K.J.; Cho, Y.W.; Park, H.J.; Lee, K.T. Isoliquiritigenin isolated from the roots of Glycyrrhiza uralensis inhibits LPS-induced iNOS and COX-2 expression via the attenuation of NF-kappaB in RAW 264.7 macrophages. Eur. J. Pharmacol. 2008, 584, 175–184. [Google Scholar] [CrossRef]
- Lee, C.; Wu, S.; Chen, J.; Li, C.; Lo, H.; Cheng, W.; Lin, J.; Chang, Y.; Hsiang, C.; Ho, T. Eriobotrya japonica leaf and its triterpenes inhibited lipopolysaccharide-induced cytokines and inducible enzyme production via the Nuclear Factor-κB signaling pathway in lung epithelial cells. Am. J. Chin. Med. 2008, 36, 1185–1198. [Google Scholar] [CrossRef]
- Chang, Y.L.; Chen, C.L.; Kuo, C.L.; Chen, B.C.; You, J.S. Glycyrrhetinic acid inhibits ICAM-1 expression via blocking JNK and NF-kappaB pathways in TNF-alpha-activated endothelial cells. Acta Pharmacol. Sin. 2010, 31, 546–553. [Google Scholar] [CrossRef]
- Uto, T.; Suangkaew, N.; Morinaga, O.; Kariyazono, H.; Oiso, S.; Shoyama, Y. Eriobotryae Folium extract suppresses LPS-induced iNOS and COX-2 expression by inhibition of NF-κB and MAPK activation in murine macrophages. Am. J. Chin. Med. 2010, 38, 985–994. [Google Scholar] [CrossRef]
- Jin, L.; Zhang, L.; Xie, K.; Ye, Y.; Feng, L. Paeoniflorin suppresses the expression of intercellular adhesion molecule-1 (ICAM-1) in endotoxintreated human monocytic cells. Br. J. Pharmacol. 2011, 164, 694–703. [Google Scholar] [CrossRef]
- Wang, C.; Kao, T.; Lo, W.; Yen, G. Glycyrrhizic Acid and 18β glycyrrhetinic acid acid modulate lipopolysaccharide-induced inflammatory response by suppression of NF-κB through PI3K p110δ and p110γ Inhibitions. J. Agric. Food Chem. 2011, 59, 7726–7733. [Google Scholar] [CrossRef]
- Kwon, H.M.; Choi, Y.J.; Choi, J.S.; Kang, S.W.; Bae, J.Y.; Kang, I.J.; Jun, J.G.; Lee, S.S.; Lim, S.S.; Kang, Y.H. Blockade of cytokine-induced endothelial cell adhesion molecule expression by licorice isoliquiritigenin through NF-kappaB signal disruption. Exp. Biol. Med. 2007, 232, 235–245. [Google Scholar]
- Ma, X.; Zhang, Y.; Wang, Z.; Shen, Y.; Zhang, M.; Nie, Q.; Hou, Y.; Bai, G. Ursolic acid, a natural nutraceutical agent, targets caspase3 and alleviates inflammation-associated downstream signal transduction. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Kao, T.; Shyu, M.; Yen, G. Glycyrrhizic acid and 18β-glycyrrhetinic acid inhibit inflammation via PI3K/Akt/GSK3β signaling and glucocorticoid receptor activation. J. Agric. Food Chem. 2010, 58, 8623–8629. [Google Scholar] [CrossRef]
- Chen, H.J.; Kang, S.P.; Lee, I.J.; Lin, Y.L. Glycyrrhetinic acid suppressed NF-κB activation in TNF-α-induced hepatocytes. J. Agric. Food Chem. 2014, 62, 618–625. [Google Scholar] [CrossRef]
- Chen, B.; Chen, Y.; Lin, W. Involvement of p38 mitogen-activated protein kinase in lipopolysaccharide-induced iNOS and COX-2 expression in J774 macrophages. Immunology 1999, 97, 124–129. [Google Scholar] [CrossRef]
- Sasaki, K.; Yonebayashi, S.; Yoshida, M.; Shimizu, K.; Aotsuka, T.; Takayama, K. Improvement in the bioavailability of poorly absorbed glycyrrhizin via various non-vascular administration routes in rats. Int. J. Pharm. 2003, 265, 95–102. [Google Scholar] [CrossRef]
- Jin, S.; Fu, S.; Han, J.; Jin, S.; Lv, Q.; Lu, Y.; Qi, J.; Wu, W.; Yuan, H. Improvement of oral bioavailability of glycyrrhizin by sodium deoxycholate/phospholipid-mixed nanomicelles. J. Drug Target. 2012, 20, 615–622. [Google Scholar] [CrossRef]
- Takeda, S.; Ishthara, K.; Wakui, Y.; Amagaya, S.; Maruno, M.; Akao, T.; Kobashi, K. Bioavailability study of glycyrrhetic acid after oral administration of glycyrrhizin in rats; relevance to the intestinal bacterial hydrolysis. J. Pharm. Pharmacol. 1996, 48, 902–905. [Google Scholar] [CrossRef]
- Kang, H.E.; Cho, Y.K.; Jung, H.Y.; Choi, K.Y.; Sohn, S.I.; Baek, S.R.; Lee, M.G. Pharmacokinetics and first-pass effects of liquiritigenin in rats: Low bioavailability is primarily due to extensive gastrointestinal first-pass effect. Xenobiotica 2009, 39, 465–475. [Google Scholar] [CrossRef]
- Dai, J.Y.; Yang, J.L.; Li, C. Transport and metabolism of flavonoids from Chinese herbal remedy Xiaochaihu-tang across human intestinal Caco-2 cell monolayers. Acta Pharmacol. Sin. 2008, 29, 1086–1093. [Google Scholar] [CrossRef]
- Takeda, S.; Isono, T.; Wakui, Y.; Matsuzaki, Y.; Sasaki, H.; Amagaya, S.; Maruno, M. Absorption and excretion of paeoniflorin in rats. J. Pharm. Pharmacol. 1995, 7, 1036–1040. [Google Scholar] [CrossRef]
- Hirsh, S.; Huber, L.; Zhang, P.; Stein, R.; Joyal, S. A single ascending dose, initial clinical pharmacokinetic and safety study of ursolic acid in healthy adult volunteers. Nutrition 2014, 28, 1044.6. [Google Scholar]
- Yang, L.; Sun, Z.; Zu, Y.; Zhao, C.; Sun, X.; Zhang, Z.; Zhang, L. Physicochemical properties and oral bioavailability of ursolic acid nanoparticles using supercritical anti-solvent (SAS) process. Food Chem. 2012, 132, 319–325. [Google Scholar] [CrossRef]
- Ge, Z.; Du, X.; Huang, X.; Qiao, B. Enhanced oral bioavailability of ursolic acid nanoparticles via antisolvent precipitation with TPGS1000 as a stabilizer. J. Drug Deliv. Sci. Technol. 2015, 29, 210–217. [Google Scholar] [CrossRef]
- Biswas, S.; Mukherjee, P.K.; Harwansh, R.K.; Bannerjee, S.; Bhattacharjee, P. Enhanced bioavailability and hepatoprotectivity of optimized ursolic acid-phospholipid complex. Drug Dev. Ind. Pharm. 2019, 45. [Google Scholar] [CrossRef]
- Garg, M.; Garg, P. Encapsulation of bio active compound ursolic acid as proniosomes and its evaluation. Asian J. Pharm. 2013, 7, 158–162. [Google Scholar] [CrossRef]
- Tian, Q.; Wang, W.; He, X.; Zhu, X.; Huang, W.; Zhang, C.; Yuan, Z.; Chen, X. Glycyrrhetinic acid-modified nanoparticles for drug delivery: Preparation and characterization. Chin. Sci. Bull. 2009, 54, 3121–3126. [Google Scholar] [CrossRef]
- Wu, F.; Li, X.; Jiang, B.; Yan, J.; Zhang, Z.; Qin, J.; Yu, W.; Gao, Z. Glycyrrhetinic acid functionalized nanoparticles for drug delivery to liver cancer. J. Biomed. Nanotechnol. 2018, 14, 1837–1852. [Google Scholar] [CrossRef]
- Boyapelly, K.; Bonin, M.; Traboulsi, H.; Cloutier, A.; Phaneuf, S.C.; Fortin, D.; Cantin, A.M.; Richter, M.V.; Marsault, E. synthesis and characterization of a phosphate prodrug of isoliquiritigenin. J. Nat. Prod. 2017, 80, 879–886. [Google Scholar] [CrossRef]
- Xu, J.; Huang, Y.; Ruan, S.; Chi, Z.; Qin, K.; Cai, B.; Cai, T. Cocrystals of isoliquiritigenin with enhanced pharmacokinetic performance. CrystEngComm 2016, 18, 8776–8786. [Google Scholar] [CrossRef]
- Xie, Y.; Wang, Q.; Adu-Frimpong, M.; Liu, J.; Zhang, K.; Xu, X.; Yu, J. Preparation and evaluation of Isoliquiritigenin-loaded F127/P123 polymeric micelles. Drug Dev. Ind. Pharm. 2019, 45. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence | Amplicon |
|---|---|---|
| RPS201 * | Forward: CCCGAAGTGGCGATTCACC | 77 nt |
| Reverse: TCCGCACAAACCTTCTCCAG | ||
| NF-κB1 | Forward: AAGGCAAAGCGAATCCAAA | 72 nt |
| Reverse: GAAGGCCTTGAATGAAATCG | ||
| NF-κB2 | Forward: GTAATCACTGGGCAGACAAGG | 95 nt |
| Reverse: AAGTGGAGGGCGGAGTCT | ||
| RelA | Forward: TCACCAAGGATCCACCTCA | 87 nt |
| Reverse: GGCAGAGGTCAGCCTCATAG | ||
| RelB | Forward: TGTTCAAAACGCCACCCTAC | 70 nt |
| Reverse: CGCTGCAAGAACACATTGAC | ||
| c-Rel # | Forward: AGACTGCGACCTCAATGTGG | 117 nt |
| Reverse: GCACGGTTGTCATAAATTGGGTT | ||
| ICAM-1 | Forward: TGGAAGCTGTTTGAGCTGAG | 67 nt |
| Reverse: TGCCACAGTTCTCAAAGCAC | ||
| TNF-α | Forward: TTGTCTTAATAACGCTGATTTGGT | 61 nt |
| Reverse: GGGAGCAGAGGTTCAGTGAT | ||
| iNOS | Forward: CAGACTGGATTTGGCTGGTC | 68 nt |
| Reverse: CAACATCTCCTGGTGGAACA | ||
| COX-2 + | Forward: TGAGCAACTATTCCAAACCAGC | 74 nt |
| Reverse: GCACGTAGTCTTCGATCACTATC |
| Substances | RAW 264.7 | Substances | RAW 264.7 |
|---|---|---|---|
| Dox # | 0.26 ± 0.05 | 18ga | 90.19 ± 2.05 |
| Ge # | 94.11 ± 4.88 | liq | 262.84 ± 26.88 |
| Pe # | 67.08 ± 5.31 | iso | 42.56 ± 5.70 |
| Ue # | 60.53 ± 4.03 | pae | 1034.91 ± 883.85 |
| ga | 163.95 ± 74.98 | urs | 23.40 ± 1.42 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.-X.; Wink, M. Evidence for Anti-Inflammatory Activity of Isoliquiritigenin, 18β Glycyrrhetinic Acid, Ursolic Acid, and the Traditional Chinese Medicine Plants Glycyrrhiza glabra and Eriobotrya japonica, at the Molecular Level. Medicines 2019, 6, 55. https://0-doi-org.brum.beds.ac.uk/10.3390/medicines6020055
Zhou J-X, Wink M. Evidence for Anti-Inflammatory Activity of Isoliquiritigenin, 18β Glycyrrhetinic Acid, Ursolic Acid, and the Traditional Chinese Medicine Plants Glycyrrhiza glabra and Eriobotrya japonica, at the Molecular Level. Medicines. 2019; 6(2):55. https://0-doi-org.brum.beds.ac.uk/10.3390/medicines6020055
Chicago/Turabian StyleZhou, Jun-Xian, and Michael Wink. 2019. "Evidence for Anti-Inflammatory Activity of Isoliquiritigenin, 18β Glycyrrhetinic Acid, Ursolic Acid, and the Traditional Chinese Medicine Plants Glycyrrhiza glabra and Eriobotrya japonica, at the Molecular Level" Medicines 6, no. 2: 55. https://0-doi-org.brum.beds.ac.uk/10.3390/medicines6020055