Multilocus Sequence Typing and Single Nucleotide Polymorphism Analysis in Tilletia indica Isolates Inciting Karnal Bunt of Wheat

Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection, Isolation and Maintenance of T. indica Isolates
2.2. DNA Isolation
2.3. Selection of MLST Loci and Amplification
2.4. Sequencing and Phylogenetic Analysis
2.5. Single Nucleotide Polymorphism Analysis
3. Results
3.1. PCR Amplification and BLAST Search
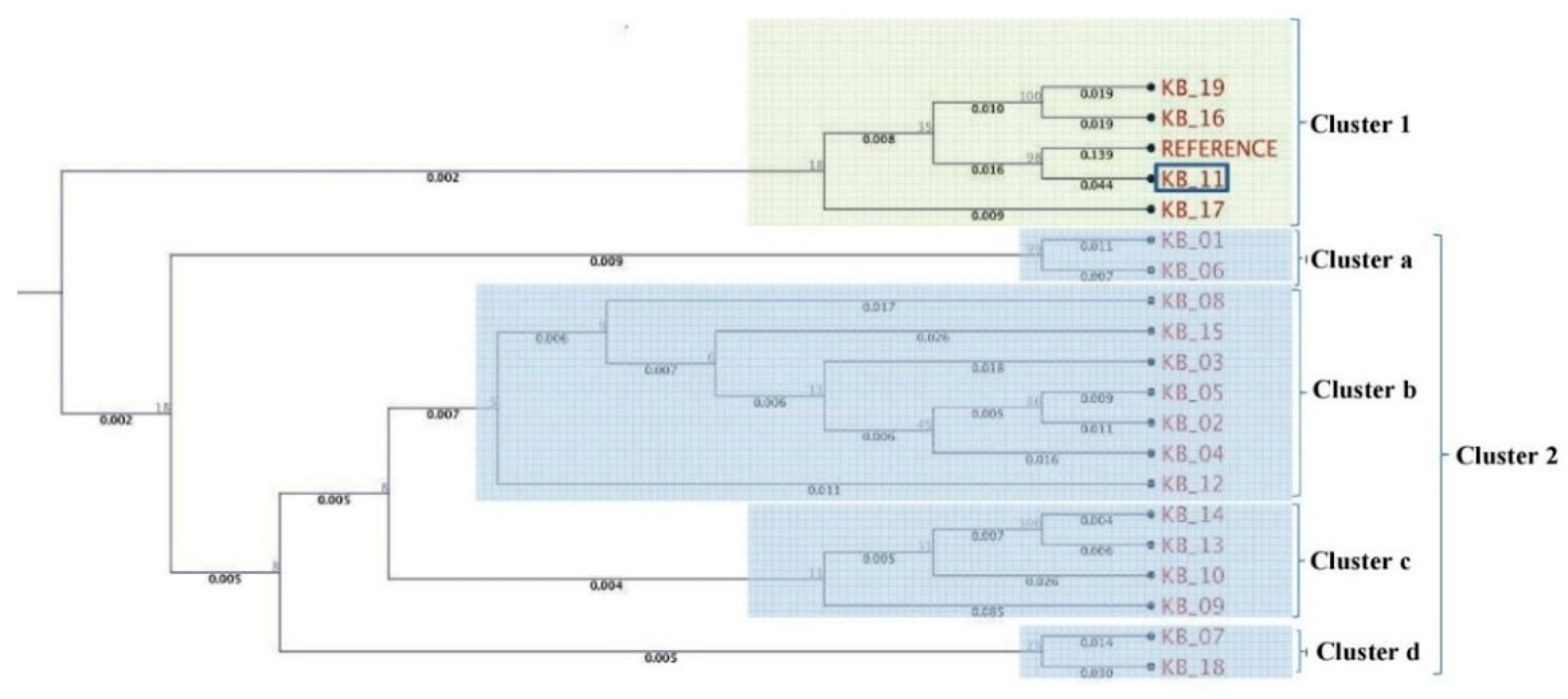
3.2. Multilocus Phylogenetic Analysis
3.3. Single Nucleotide Polymorphism (SNP) Analysis
3.4. Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ARP2 | Actin-related protein 2 |
| EIF3A | Eukaryotic translation initiation factor 3 subunit A |
| GAPDH | Glyceraldehyde-3-phosphate dehydrogenase |
| H2B | Histone 2B |
| KB | Karnal bunt |
| MLST | Multilocus sequence typing |
| PGK | Phosphoglycerate kinase |
| SNP | Single nucleotide polymorphism |
| STPK | serine/threonine-protein kinase |
| TUB | β-tubulin |
References
- Anonymous. IARI Annual Report (2015–2016); Indian Agricultural Research Institute: New Delhi, India, 2015; p. 83. Available online: www.iari.res.in (accessed on 3 January 2021).
- Mitra, M. A new bunt of wheat in India. Ann. Appl. Biol. 1931, 18, 178–179. [Google Scholar] [CrossRef]
- Bonde, M.R.; Nester, S.E.; Olsen, M.W.; Berner, D.K. Survival of teliospores of Tilletia indica in Arizona field soils. Plant Dis. 2004, 88, 804–810. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.V.; Gogoi, R. Karnal bunt of wheat (Triticum sp.): A global scenario. Indian J. Agric. Sci. 2011, 81, 3–14. [Google Scholar]
- Aggarwal, R.; Singh, D.V.; Srivastava, K.D. Host pathogen interaction in Karnal bunt of wheat. Indian Phytopathol. 1994, 47, 381–386. [Google Scholar]
- Aggarwal, R.; Tripathi, A.; Yadav, A. Pathogenic and genetic variability in Tilletia indica monosporidial culture lines using universal rice primer-PCR. Eur. J. Plant Pathol. 2010, 128, 333–342. [Google Scholar] [CrossRef]
- Carris, L.M.; Castlebury, L.A.; Goates, B.J. Nonsystemic Bunt Fungi-Tilletia indica and T. horrida: A Review of History, Systematics, and Biology. Annu. Rev. Phytopathol. 2006, 44, 113–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babadoost, M.; Mathre, D.E.; Johnston, R.H.; Bonde, M.R. Survival of teliospores of Tilletia indica in soil. Plant Dis. 2004, 88, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Goates, B.J. Survival of secondary sporidia of floret infecting Tilletia species: Implications for epidemiology. Phytopathology 2010, 100, 655–662. [Google Scholar] [CrossRef]
- Dhaliwal, H.S. Multiplication of secondary sporidia of Tilletia indica on soil and wheat leaves and spikes and incidence of Karnal bunt. Can. J. Bot. 1989, 67, 2387–2390. [Google Scholar] [CrossRef]
- Thirumalaisamy, P.P.; Singh, D.V. Variability of Indian isolates of Tilletia indica assessed by pathogenecity and molecular marker. J. Phytopathol. 2012, 160, 525–531. [Google Scholar] [CrossRef]
- Shabana, P.; Saharan, M.S.; Verma, A.; Sharma, I. Comparative analysis of RAPD and ISSR marker assays for detecting genetic polymorphism in Tilletia indica. Eur. J. Exp. Biol. 2013, 3, 380–387. [Google Scholar]
- Taylor, J.W.; Fisher, M.C. Fungal multilocus sequence typing—It’s not just for bacteria. Curr. Opin. Microbiol. 2003, 6, 351–356. [Google Scholar] [CrossRef]
- Bain, J.M.; Tavanti, A.; Davidson, A.D.; Jacobsen, M.D.; Shaw, D.; Gow, N.A.R.; Odds, F.C. Multilocus Sequence Typing of the Pathogenic Fungus Aspergillus fumigates. J. Clin. Microbiol. 2007, 5, 1469–1477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caicedo, J.D.; Karina, P.; Lalangui, A.N.; Pozo, P.A.; Cevallos, V.; Arahana, S.; Mendez, K.S. Multilocus molecular identification and phylogenetic analysis of Colletotrichum tamarilloi as the causal agent of Tamarillo (Solanum betaceum) anthracnose in the Ecuadorian highlands. Eur. J. Plant Pathol. 2017, 148, 983–996. [Google Scholar] [CrossRef]
- Warham, E.J. Studies on Karnal Bunt of Wheat. Ph.D. Thesis, University of Wales, Aberystwyth, UK, 1987. [Google Scholar]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4325. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Rush, C.M.; Riemenschneider, R.; Stein, J.M.; Boratynski, T.; Bowden, R.L.; Matthew, H.R. Status of Karnal Bunt of Wheat in the Unites States 1996 to 2004. Plant Dis. 2005, 89, 212–223. [Google Scholar] [CrossRef] [Green Version]
- Gurjar, M.S.; Aggarwal, R.; Jogawat, A.; Kulshreshtha, D.; Sharma, S.; Solanke, A.U.; Dubey, H.; Jain, R.K. Genome Sequencing and Secretome analysis of Tilletia indica inciting Karnal bunt of wheat Provides Pathogenesis-related genes. 3 Biotech 2019, 9, 219. [Google Scholar] [CrossRef]
- Debourgogne, A.; Gueidan, C.; Henneguin, C.; Contet-Audonneau, N.; de Hoog, S.; Machouart, M. Development of a new MLST scheme for differentiation of Fusarium solani Species Complex (FSSC) isolates. J. Microbiol. Methods 2010, 82, 319–323. [Google Scholar] [CrossRef]
- Byrnes, E.J.; Bildfell, R.J.; Frank, S.A.; Mitchell, T.G.; Marr, K.A.; Heitman, J. Molecular evidence that the range of the Vancouver Island outbreak of Cryptococcus gattii infection has expanded into the Pacific Northwest in the United States. J. Infect. Dis. 2009, 199, 1081–1086. [Google Scholar] [CrossRef] [Green Version]
- Litvintseva, A.P.; Mitchell, T.G. Population Genetic Analyses Reveal the African origin and strain variation of Cryptococcus neoformans var. grubii. PLoS Pathog 2012, 8, e1002495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datta, R.; Rajebhosale, M.D.; Dhaliwal, H.S.; Singh, H.; Ranjekar, P.K.; Gupta, V.S. Intraspecific genetic variability analysis of Neovossia indica causing Karnal bunt of wheat using repetitive elements. Theor. Appl. Genet. 2000, 100, 569–575. [Google Scholar] [CrossRef]
- Tripathi, A.; Aggarwal, R.; Yadav, A. Determination of variability in monosporidial lines of Tilletia indica by RAPD analysis. Arch. Phytopathol. Plant Prot. 2011, 44, 1312–1321. [Google Scholar] [CrossRef]
- Urwin, R.; Maiden, M.C.J. Multi-locus sequence typing: A tool for global epidemiology. Trends Microbiol. 2003, 11, 479–487. [Google Scholar] [CrossRef]
- Meyer, W.; Aanensen, D.M.; Boekhout, T.; Cogliati, M.; Diaz, M.R.; Esposto, M.C.; Fisher, M.; Gilgado, F.; Hagen, F.; Kaocharoen, S.; et al. Consensus multi-locus sequence typing scheme for Cryptococcus neoformans and Cryptococcus gattii. Med. Mycol. 2009, 12, 1–14. [Google Scholar]
- Dhaliwal, H.S.; Singh, D.V. Up-to-date life cycle of Nevossia indica (Mitra) Mundkur. Curr. Sci. 1988, 57, 675–677. [Google Scholar]
- Duran, R.; Cromarty, R. Tilletia indica: A heterothallic wheat bunt 8 fungus with multiple alleles controlling incompatibility. Phytopathology 1977, 67, 812–815. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Frazzitta, A.E.; Litvintseva, A.P.; Fang, C.; Mitchell, T.G.; Springer, D.J.; Ding, Y.; Yuan, G.; Perfect, J.R. Next generation multilocus sequence typing (NGMLST) and the analytical software program MLSTEZ enable efficient, cost-effective, high-throughput, multilocus sequencing typing. Fungal Genet. Biol. 2015, 75, 64–71. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| S. No. | Genes | Sequences (5′-3′) |
|---|---|---|
| 1. | actin-related protein 2F actin-related protein 2R | TGCAGCAGCAGGCCAGATCAAG GGAAGGTAGGCCGGGGTACAT |
| 2. | β-tubulin F β-tubulin R | GTCCTTATCGACTTGGAGCCC AGTCCTGGATAGCAGTGCTG |
| 3. | eukaryotic translation initiation factor 3 subunit A F Eukaryotic translation initiation factor 3 subunit A R | TGCACGGTGACCAAGAAACTC CGGAGTTCTTCTTGTCGAGCA |
| 4. | glyceraldeyde-3-phosphate dehydrogenase F glyceraldeyde-3-phosphate dehydrogenase R | TGAAGGGTGGTGCCAAGAAG GAAGACGGAGCTGGCGGAGT |
| 5. | histone 2B F histone 2B R | TCTACAGGCGGCAAGGCTCC GCGGGCAGAGATGGTCGACTT |
| 6. | phosphoglycerate kinase F phosphoglycerate kinase R | GTCCCGATGGTCAGAAGGTCGA AGCATACTCTTCGAGCCCGC |
| 7. | Serine/threonine-protein kinase F Serine/threonine-protein kinase R | CAGATTGTCGCCGCCTGTCAA TTTCTTGCAACAGCCGACGCT |
| Isolates | Locations | NCBI GeneBank Accession Numbers * | ||||||
|---|---|---|---|---|---|---|---|---|
| ARP2 | βTUB | EIF3A | GAPDH | H2B | PGK | STPK | ||
| KB-1 | Aligarh, Uttar Pradesh | MG655312 | MG655332 | MG386622 | MG386602 | MG870326 | MG701241 | MG655332 |
| KB-2 | New Delhi, Delhi | MG655313 | MG655333 | MG386623 | MG386603 | MG870327 | MG701242 | MG655333 |
| KB-3 | Karnal, Haryana | MG655314 | MG655334 | MG386624 | MG386604 | MG870328 | MG701243 | MG655334 |
| KB-4 | Kurukshetra, Haryana | MG655315 | MG655335 | MG386625 | MG386605 | MG870329 | MG701244 | MG655335 |
| KB-5 | Nilokheri, Haryana | MG655316 | MG655336 | MG386626 | MG386606 | MG870330 | MG701245 | MG655336 |
| KB-6 | Sonipat, Haryana | MG655317 | MG655337 | MG386627 | MG386607 | MG870331 | MG701246 | MG655337 |
| KB-7 | Jind, Haryana | MG655318 | MG655338 | MG386628 | MG386608 | MG870332 | MG701247 | MG655338 |
| KB-8 | Panipat, Haryana | MG655319 | MG655339 | MG386629 | MG386609 | MG870333 | MG701248 | MG655339 |
| KB-9 | Bareilly, Uttar Pradesh | MG655320 | MG655340 | MG386630 | MG386610 | MG870334 | MG701249 | MG655340 |
| KB-10 | Bhiwani, Haryana | MG655321 | MG655341 | MG386631 | MG386611 | MG870335 | MG701250 | MG655341 |
| KB-11 | Kaithal, Haryana | MG655322 | MG655342 | MG386632 | MG386612 | MG870336 | MG701251 | MG655342 |
| KB-12 | Ambala, Haryana | MG655323 | MG655343 | MG386633 | MG386613 | MG870337 | MG701252 | MG655343 |
| KB-13 | Pantnagar, Uttarakhand | MG655324 | MG655344 | MG386634 | MG386614 | MG870338 | MG701253 | MG655344 |
| KB-14 | Sultanpur, Uttar Pradesh | MG655325 | MG655345 | MG386635 | MG386615 | MG870339 | MG701254 | MG655345 |
| KB-15 | Allahabad, UP | MG655326 | MG655346 | MG386636 | MG386616 | MG870340 | MG701255 | MG655346 |
| KB-16 | Bulandshahr, Uttar Pradesh | MG655327 | MG655347 | MG386637 | MG386617 | MG870341 | MG701256 | MG655347 |
| KB-17 | Saharanpur, UP | MG655328 | MG655348 | MG386638 | MG386618 | MG870342 | MG701257 | MG655348 |
| KB-18 | Mujaffar Nagar, UP | MG655329 | MG655349 | MG386639 | MG386619 | MG870343 | MG701258 | MG655349 |
| KB-19 | Pataudi, Haryana | MG655330 | MG655350 | MG386640 | MG386620 | MG870344 | MG701259 | MG655350 |
| KB-20 | Shamli, Haryana | MG655331 | MG655351 | MG386641 | MG386621 | MG870345 | MG701260 | MG655351 |
| Genes # | ARP2 | βTUB | EIF3A | GAPDH | H2B | PGK | STPK | Total Identified SNPs/Isolates |
|---|---|---|---|---|---|---|---|---|
| Sequence Length/Reference | 1100/1200 bp | 960/1350 bp | 1400/3300 bp | 700/1000 bp | 300/430 bp | 900/1250 bp | 900/1250 bp | |
| KB-01 | 42 | 5 | 26 | 8 | 2 | 36 | 9 | 128 |
| KB-02 | 28 | 13 | 26 | 9 | 1 | 34 | 7 | 118 |
| KB-03 | 7 | 3 | 13 | 8 | 2 | 32 | 3 | 68 |
| KB-04 | 22 | 11 | 26 | 9 | 0 | 35 | 10 | 113 |
| KB-05 | 29 | 12 | 17 | 10 | 1 | 35 | 14 | 118 |
| KB-06 | 40 | 6 | 26 | 10 | 2 | 37 | 14 | 135 |
| KB-07 | 41 | 5 | 26 | 22 | 0 | 34 | 6 | 134 |
| KB-08 | 23 | 0 | 19 | 5 | 0 | 34 | 16 | 97 |
| KB-09 | 37 | 7 | 26 | 14 | 2 | 0 | 23 | 109 |
| KB-10 | 6 | 5 | 16 | 6 | 0 | 41 | 17 | 91 |
| KB-11 | 7 | 4 | 13 | 10 | 0 | 30 | 3 | 67 |
| KB-12 | 38 | 5 | 15 | 10 | 2 | 32 | 20 | 122 |
| KB-13 | 58 | 3 | 26 | 10 | 1 | 42 | 13 | 153 |
| KB-14 | 58 | 4 | 26 | 8 | 1 | 42 | 25 | 164 |
| KB-15 | 7 | 10 | 17 | 22 | 0 | 35 | 14 | 105 |
| KB-16 | 41 | 12 | 26 | 18 | 0 | 35 | 17 | 149 |
| KB-17 | 40 | 3 | 27 | 9 | 0 | 33 | 8 | 120 |
| KB-18 | 38 | 5 | 26 | 43 | 2 | 40 | 11 | 165 |
| KB-19 | 40 | 4 | 26 | 17 | 2 | 37 | 17 | 143 |
| KB-20 | 38 | 34 | 27 | 7 | 0 | 31 | 8 | 145 |
| Total | 640 | 151 | 450 | 255 | 18 | 675 | 255 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gurjar, M.S.; Aggarwal, R.; Jain, S.; Sharma, S.; Singh, J.; Gupta, S.; Agarwal, S.; Saharan, M.S. Multilocus Sequence Typing and Single Nucleotide Polymorphism Analysis in Tilletia indica Isolates Inciting Karnal Bunt of Wheat. J. Fungi 2021, 7, 103. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7020103
Gurjar MS, Aggarwal R, Jain S, Sharma S, Singh J, Gupta S, Agarwal S, Saharan MS. Multilocus Sequence Typing and Single Nucleotide Polymorphism Analysis in Tilletia indica Isolates Inciting Karnal Bunt of Wheat. Journal of Fungi. 2021; 7(2):103. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7020103
Chicago/Turabian StyleGurjar, Malkhan Singh, Rashmi Aggarwal, Shekhar Jain, Sapna Sharma, Jagmohan Singh, Sangeeta Gupta, Shweta Agarwal, and Mahender Singh Saharan. 2021. "Multilocus Sequence Typing and Single Nucleotide Polymorphism Analysis in Tilletia indica Isolates Inciting Karnal Bunt of Wheat" Journal of Fungi 7, no. 2: 103. https://0-doi-org.brum.beds.ac.uk/10.3390/jof7020103