
Very High Gravity Bioethanol Revisited: Main Challenges and Advances
by
 , , and
, , and
Daniel Gomes
1,* ,
,
Mariana Cruz
2,
Miriam de Resende
2,
Eloízio Ribeiro
2,
José Teixeira
1 and
Lucília Domingues
1 1
CEB-Centre of Biological Engineering, Universidade do Minho, Campus de Gualtar, 4710-057 Braga, Portugal
2
Faculty of Chemical Engineering, Federal University of Uberlândia, Campus Santa Mônica, Uberlândia 38408-144, Brazil
*
Author to whom correspondence should be addressed.
Fermentation 2021, 7(1), 38; https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7010038
Submission received: 31 January 2021
/
Revised: 11 March 2021
/
Accepted: 11 March 2021
/
Published: 13 March 2021
(This article belongs to the Special Issue Biofuel Fermentation from Renewable Biomass)
Abstract
:Over the last decades, the constant growth of the world-wide industry has been leading to more and more concerns with its direct impact on greenhouse gas (GHG) emissions. Resulting from that, rising efforts have been dedicated to a global transition from an oil-based industry to cleaner biotechnological processes. A specific example refers to the production of bioethanol to substitute the traditional transportation fuels. Bioethanol has been produced for decades now, mainly from energy crops, but more recently, also from lignocellulosic materials. Aiming to improve process economics, the fermentation of very high gravity (VHG) mediums has for long received considerable attention. Nowadays, with the growth of multi-waste valorization frameworks, VHG fermentation could be crucial for bioeconomy development. However, numerous obstacles remain. This work initially presents the main aspects of a VHG process, giving then special emphasis to some of the most important factors that traditionally affect the fermentation organism, such as nutrients depletion, osmotic stress, and ethanol toxicity. Afterwards, some factors that could possibly enable critical improvements in the future on VHG technologies are discussed. Special attention was given to the potential of the development of new fermentation organisms, nutritionally complete culture media, but also on alternative process conditions and configurations.
1. Introduction
The application of process engineering strategies to achieve high-productivity fermentation systems is now considered a key issue in the bioethanol industry. A possible strategy to increase ethanol production and minimize the production cost is a fermentation with high final ethanol titers, achieved by using musts with high sugar concentrations, commonly known as very high gravity (VHG) fermentation. In general, sugar concentrations for ethanol production can be divided into normal gravity (NG) (<180 g/L of total sugars), high gravity (HG) (180–240 g/L of total sugars), and very high gravity (VHG) (≥250 g/L of total sugars) [1,2]. HG fermentation was proposed in the 1980s and was successfully commercialized thereafter, making final ethanol concentrations to increase drastically from a level of 7–8% (v/v) to 10–12% (v/v) [3]. As a result, VHG fermentations using sugar levels in excess of 250 g/L (enabling to achieve over 15% (v/v) ethanol) were then proposed in the 1990s; Thomas et al. [4] defined VHG technology as the fermentation of a media containing 270 g or more of dissolved solids per liter of mixture.
The application of VHG technology in bioethanol production has a number of benefits, namely: less process water and energy requirements; increased overall production plant productivity; higher final ethanol concentrations (with considerable savings on energy for distillation) [5]. An assessment of bioethanol production regarding energy requirements revealed that distillation is one of the most energy consuming steps, hence, a superior ethanol concentration in the final broth can considerably reduce the energy cost of this particular step [6]. Despite the potential benefits, VHG processes also expose cells to increased stressful conditions, which have been commonly associated to a loss of viability, reduced fermentation rates, and incomplete fermentations [7,8,9]. Therefore, the successful implementation of VHG requires the development of suitable organisms able to efficiently ferment high sugar concentrations, while being able to cope with the multiple stresses found in the process. In this context, Saccharomyces cerevisiae has been positioning itself as the most employed organism in VHG processes, and hence will be especially addressed throughout this review.
2. Main Challenges Arising from VHG Processes
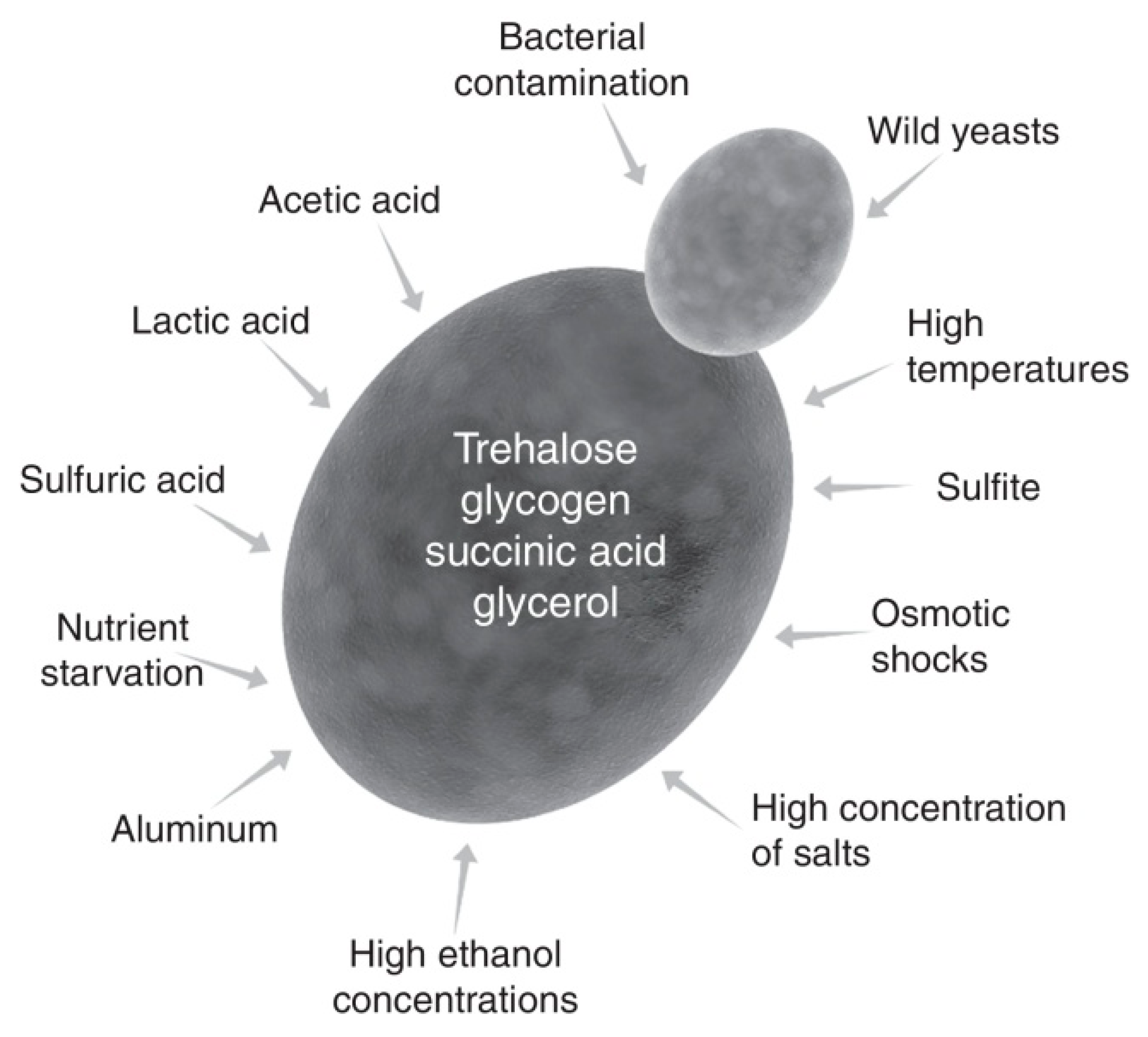
During ethanol production, cells are exposed to a variety of physical, chemical, and biological factors, typically referred to as stress factors. Yeast cells, however, have mechanisms of stress response towards industrial fermentation conditions [10], as presented by Figure 1.
In the context of VHG fermentations, yeast cells are exposed to a particular set of stresses: high osmotic pressure at the beginning of the process, caused by an high wort sugar concentration; nutrients starvation from a depletion of critical nutrients; lack of oxygen; accumulation of high ethanol concentrations, which in addition to the high levels of other toxic by-products, becomes lethal to cells [5,12,13,14].
2.1. Physiological Stress
High sugar and ethanol concentrations at the beginning and at the end of fermentation, respectively, expose yeast cells to physiological stress, supporting the need for more robust strains able to maintain cell viability and the fermentative capacity more efficiently over fermentation cycles. In this context, medium osmolality is possibly one of the main determinants of cells viability since it affects the control of water content, crucial for cells integrity. Cells exposed to hypertonic stress lose water, inducing an adaptive response to compensate for dehydration and to protect cellular structures [15]. The accumulation of solutes by the cell, in particular glycerol, acts as an essential response to acquire resistance to osmolality changes, as it decreases the intracellular potential of water. Changes on lipids metabolism may also be involved in osmolality regulation since modifications in the lipid composition of the plasma membrane may affect membrane fluidity and permeability to water and glycerol [16]. Another component whose cellular content increases under conditions of osmotic stress is trehalose. The importance of trehalose is proven by the correlation between its intracellular level and the ability to resist osmotic stress induced by NaCl or sorbitol. In fact, mutant strains of Saccharomyces cerevisiae unable to produce trehalose revealed a greater sensitivity to osmotic stress [17]. Besides being an energy reserve carbohydrate, trehalose is also a protecting agent for the membrane towards dehydration, high ethanol concentrations, and other industrial stress factors [18,19].
In what concerns ethanol, it is commonly regarded as one of the main stresses acting on yeasts due to its natural toxicity, which ultimately affects cells growth and viability [20]. Early studies by Casey and Ingledew [21] showed that the high ethanol concentrations obtained from the fermentation of sugar-rich substrates led to a reduction on the specific growth rate and fermentation rate of S. cerevisiae, but also on cells viability. Ethanol affects yeast cells on multiples levels and targeting different components of their structure [20]. One example is its effect over cells metabolism and the synthesis of specific molecules. According to Hu et al. [22], ethanol can not only induce the production of heat shock-like proteins, causing a reduction of mRNA and protein accumulation, but also a denaturation of intracellular proteins and glycolytic enzymes, which will directly affect critical metabolic capacities involved in cell growth and fermentation. Another key target structure on yeasts is their cell membrane where ethanol can cause an increase of fluidity, resulting in a loss of membrane integrity [23]. Additionally, according to Piper et al. [24], many of these changes induced from stressful levels of ethanol are identical to those caused by thermal stress.
2.2. Robustness of the Fermentation Organism
The capacity to withstand the stressful conditions mentioned before and to efficiently perform under a VHG environment is strongly dependent on cells robustness. In this context, S. cerevisiae is the yeast typically used to study the molecular mechanisms involved in stress response. Research on yeast physiology has revealed that many S. cerevisiae strains can potentially tolerate far higher ethanol concentrations than previously believed, generally without any conditioning or genetic modifications [21,25,26] (Table 1).
An early study from Pereira et al. [26] showed that the industrial S. cerevisiae isolates PE-2 and CA1185 had an excellent performance in VHG fermentations, producing high ethanol titers (>19% v/v) with high productivities (>2.5 g/L·h), which suggests they are more physiologically robust under these conditions. Furthermore, these isolates showed to accumulate superior concentrations of trehalose, glycogen, and sterols when compared to the laboratory strains, which may explain the superior stress tolerance and fermentation performance. Trehalose and glycogen are energy reserve carbohydrates used by cells under sugar starvation conditions [27,28]. On a recent chemostat study, it was observed that the accumulations of trehalose and glycogen were strongly dependent of the cultivation temperature, with the highest glycogen accumulation at 12 °C and the highest trehalose accumulation at 39 °C [29]. Additionally, the thermotolerant industrial strains presented significantly higher trehalose accumulation compared to the reference laboratorial CEN.PK113-7D strain. At the proteome level, temperature responses also differ between S. cerevisiae strains [30]. Interestingly, thermotolerant S. cerevisiae Ethanol Red response to both supra and sub-optimal temperature involved the overexpression of Erg13—a protein involved in early ergosterol biosynthesis—and Gsy1—a glycogen synthetase [30].

{kind=link}
Table 1.
Comparative performance in the production of bioethanol by different strains of S. cerevisiae using different substrates, initial concentration, and temperature.
Table 1.
Comparative performance in the production of bioethanol by different strains of S. cerevisiae using different substrates, initial concentration, and temperature.
| Strain | Substrate | Initial Substrate (g/L) | Temperature (°C) | Final Ethanol (g/L) | Reference |
|---|---|---|---|---|---|
| AT-3 | Glucose | 180 | 40 | 68.5 | [31] |
| SEMF1 | Sweet sorghum juice | 185 | 33 | 86.2 | [32] |
| NRRL Y-2034 | Sweet sorghum juice | 200 | 30 | 70.6 | [33] |
| DMKU 3-S087 | Molasses | 200 | 40 | 72.4 | [34] |
| KKU-VN8 | Sweet sorghum juice | 238 | 40 | 89.3 | [35] |
| CCTCC M206111 | Sweet potato | 240 | 30 | 128.5 | [36] |
| Y-904 | Sugarcane juice and molasses | 300 | 27 | 135.0 | [37] |
| PE-2 | Glucose | 323 | 30 | 149.0 | [38] |
| YF10-5 | Glucose | 350 | 30 | 115.0 | [39] |
| KL17 | Galactose and glucose | 500 | 30 | 96.9 | [40] |
| C10 | Sugar beet syrup | 270 | 30 | 116.0 | [41] |
On a similar way, it was also possible to identify different critical ethanol concentrations for laboratory and industrial strains above which cells viability significantly decreased. For the industrial strains, cells viability remained above 85% for ethanol concentrations up to 140 g/L, sharply decreasing after that; on laboratory strains the decline on viability occurred for a much lower ethanol concentration (85–100 g/L), confirming its lower resistance to ethanol [26].
2.3. Nutrients Depletion
Previous research showed that yeast cells capacity to achieve high ethanol levels strongly depends on the nutritional conditions and the protective function that some nutrients can provide. A common limitation of the VHG technology refers to a lack of critical elements required by cells, which are usually present in insufficient amounts considering the levels of sugars available [1].
After carbon, assimilable nitrogen is the most important component in the fermentation media and has been reported to be a limiting nutrient in VHG fermentations. It represents one of the essential nutrients for ethanol production, especially under VHG conditions [1]; not only promotes yeast growth and ethanol production, but also tolerance to ethanol [42]. On lab-scale studies, yeast extract is widely used as a nitrogen source for yeast cells growth and as a nutrient supplement in ethanol production [43,44], however, it is not appropriate for industrial ethanol production due to its high cost. It is thus important to exploit low-cost nitrogen sources able to provide the nutritional requirements for yeast growth and fermentation. Several potential options have already been studied, such as millet flour [45,46], corn steep liquor [47,48], fresh yeast auto lysate [49], and spent yeast from the brewing sector [48,50].
In addition to carbon and nitrogen, trace elements such as zinc, magnesium, and manganese, all involved in several metabolic pathways, have been also widely reported to promote sugar conversion, allowing a superior ethanol tolerance and production levels, particularly under VHG conditions [5,47,51,52,53]. Zinc is an essential element for the normal growth, metabolism, and physiology of yeast cells, acting as an important co-factor of many proteins [32]. On an early work conducted by Zhao and Bai [53], medium supplementation with zinc was found to increase cells resistance to high temperatures and ethanol concentrations, leading to an increase in the ergosterol content of the cell membrane and in the levels of intracellular trehalose. The addition of 50 ppm zinc sulfate resulted in a 9.6% increase of ethanol production. On the other hand, glycerol production decreased with zinc supplementation, reaching a minimum of 3.21 g/L. A close association between cells viability and the levels of supplemented zinc was also observed, leading to an increase of the ethanol production yield. Similarly, calcium and magnesium, both present in sugarcane juice, were previously associated by Trofimova et al. [54] to a higher tolerance to the stress caused by hydration and dehydration, being important for the stabilization of yeast cell membranes, but also acting as protecting agents under high concentrations of ethanol, especially when subjected to large environmental changes such as on the osmolality levels [55].
An early study from Pereira et al. [47] compared the fermentation performance of S. cerevisiae strains under two distinct VHG mediums: a reference medium (RM) containing 100 g/L CSL as sole nutrient source; an optimized medium (OM) supplemented with low-cost nutrient sources (g/L: CSL 44.3, urea 2.3, MgSO4.7H2O 3.8 and CuSO4.5H2O 0.03). On fermentations conducted with approximately 300 g/L of glucose, CEN.PK 113-7D produced a maximum ethanol concentration of 130 g/L with the OM; opposing to that, only 120 g/L were achieved with the RM. For PE-2, the maximum ethanol concentration in fermentations with 300–330 g/L of glucose was 147 g/L using the OM, decreasing to 130 g/L with the RM. Similarly, the influence of nutrient supplements on cells viability was also assessed. Near the end of the fermentation, when ethanol titers were close to 15.0% (v/v), viability of CEN.PK 113-7D cells improved from 64%, with the RM, to 85% with the OM. In the same way, when 16.8% (v/v) of ethanol was obtained with PE-2, cells viability improved from 43% with the RM to 89% with the OM.
3. Critical Factors for Improved VHG Fermentations
Even though representing an interesting option from an economic point of view, VHG technology still faces important challenges towards its efficient implementation on an industrial scale. To achieve that, some major factors should be taken into account in the development of new/existing VHG systems, both by academia and industry (Table 2).
3.1. Temperature
Although high temperatures are commonly desirable in the bioethanol industry, mostly due to the potential savings on cooling requirements and reduced contaminations risk, in addition to leading to a higher ethanol production by yeast cells [29], their negative impacts on fermentation are more pronounced under VHG conditions. This is explained by the consequent increase of ethanol-related stresses, the intensification of non-growth-related energy demands [29] and by the fact that ethanol tolerance usually decreases as temperatures rises [56,57]. Indeed, in an early study from Gao and Fleet [58], the authors observed that S. cerevisiae cells incubated for 12 days in 15% ethanol at 10 or 15 °C did not lose any viability. Differently, for an incubation temperature of 30 °C, cells viability slightly decreased. In a previous work from Pereira et al. [59] on the development of cell recycling systems for VHG processes, the authors observed clear improvements over cells performance when the fermentation temperature was reduced from 30 to 27 °C. Specifically, for an initial glucose concentration of 400 g/L and a temperature of 30 °C, S. cerevisiae PE-2 cells were able to produce 18.2% (v/v) of ethanol, leaving a sugar residual around 80 g/L. Opposing that, when the temperature was reduced to 27 °C, cells were able to produce 20.1% (v/v) of ethanol and the glucose residual decreased to 60 g/L.
Another example points to the findings previously reported by Laluce et al. [60], who conducted an optimization study regarding critical variables on VHG fermentation, such as sugar concentration, temperature, and inoculum size. According to the authors, as sugar concentration increases, fermentation temperature must be reduced to attenuate ethanol-induced stresses, especially manifested by a decrease of cells viability. It is thus recommended to lower the fermentation temperature to minimize cell death and maintain high levels of ethanol production when the temperature is increasing in the industrial reactor.
Achieving efficient high-temperature fermentations on a VHG context will therefore rely on the organism capacity to withstand the accumulation of high concentrations of ethanol at elevated temperatures, without compromising cells viability. This will mostly depend on the utilization of organisms with an increased thermotolerance, such as the yeast Kluyveromyces marxianus. In an early study from Limtong et al. [61], the fermentation performance of K. marxianus strain DMKU 3-1042 (isolated by an enrichment technique in a sugar cane juice medium supplemented with 4% (w/v) ethanol at 35 °C) was assessed at different temperatures. In shaking flask cultivations and using a sugar cane juice media containing 22% of total sugars, 0.05% (NH4)2SO4, 0.05% KH2PO4, and 0.15% MgSO4.7H2O, for a temperature of 37 °C, the ethanol concentration reached 8.7% (w/v), the productivity 1.45 g/L·h, and the production yield 77.5%. At 40 °C, the maximum ethanol concentration was still 6.78% (w/v), while productivity decreased to 1.13 g/L·h and the production yield decreased to 60.4%. Additionally, in a recent work by Malairuang et al. [62], the authors observed that K. marxianus SS106 isolate was able to tolerate temperatures in the range of 35–45 °C, allowing to achieve high cell densities on the process. Similarly, a previous study from Techaparin et al. [35] reported the utilization of several thermotolerant yeast isolates to produce bioethanol under high fermentation temperatures. From the entire set of initial isolates, a selection of five isolates showed clear benefits towards an operation under high temperatures, specifically between 40–43 °C, inclusively outrunning the performance of an industrial strain commonly used for bioethanol production (S. cerevisiae TISTR 5606).
From the facts referred before, it seems clear that for an efficient VHG fermentation, a quantitative understanding of the effect of temperature on substrate consumption and ethanol production must be investigated to define the most suitable operating conditions. As an example, Rivera et al. [63] addressed the effect of temperature on ethanol production from sugarcane by S. cerevisiae using a mechanistic model formulated in the study. According to the authors, as the temperature increased, the maximum specific growth rate also increased; however, beyond approximately 37 °C, it started to decrease. Additionally, the maximum levels of ethanol and cell concentration were found to be inversely related to the fermentation temperature.
3.2. Agitation and Aeration Conditions
On yeasts, ethanol is usually produced via the glycolytic route under anaerobic conditions. Nevertheless, numerous studies have already found that an appropriate aeration associated with an efficient agitation, especially in the beginning of yeast growth phase, can significantly improve the subsequent sugar consumption and ethanol production, even more under VHG conditions [44,57]. According to You et al. [64], S. cerevisiae requires a given amount of elemental oxygen to synthesize ergosterol and unsaturated fatty acids, both essential for plasma membrane integrity. The required levels of aeration are dependent of several factors, namely the specific yeast strain, nutrients availability, and the fermentation process [65,66,67]. A previous work from Deesuth et al. [2] addressed the potential role of aeration (and nutrients) on improving VHG processes. According to the authors, an aeration of 0.05 vvm for 12 h enabled an increase of final ethanol titers from 114.8 to 126.3 g/L, but also of ethanol productivity from 1.91 to 2.11 g/L·h. On the other hand, an early study from Lin et al. [68] demonstrated that the timing, duration, and intensity of the aeration step critically influence the amount of viable cells, and consequently, the overall levels of ethanol production. The authors observed that under the best aeration conditions, referring to a flow of 0.82 L/min in the log phase, the amount of viable cells had a 2-fold increase comparatively to a no-aeration scenario, and allowed to reduce the period of full glucose consumption from 48 to 36 h. Similarly, Khongsay et al. [44] observed that under optimum agitation conditions, the application of an aeration step (2.5 vvm; 4 h) allowed a general improvement of ethanol titers (118 to 133 g/L) and productivities (2.19 to 2.55 g/L·h) when compared to the absence of aeration.
From another perspective, and still in the scope of oxygen availability for cells, adding to its supply, it is equally critical its transference to cells. In this context, it is most relevant to discuss the close relation between oxygen diffusion in the media and efficient agitation methods, which could be especially relevant when substrates with a high gelatinization degree are employed, such as those based on starch. This aspect is commonly addressed in VHG research, namely with the optimization of mechanical agitation [44]. This may also refer to a new agitation apparatus or the development of new process schemes which may enable more favorable media rheology profiles, as are the cases of liquefaction-aiding enzymes. Specifically, growing interest has been given to the utilization of cell wall degrading enzymes to facilitate the liquefaction of the substrate solution. Poonsrisawat et al. [69] recently reported that the utilization of a combined mixture of different cell wall degrading enzymes (from different fungus) allowed clear improvements on medium rheology. When a cassava root mash was employed, the application of 0.05 mg protein/gsolid (with equal parts of three enzymes) for 2 h resulted on a significant decrease of complex viscosity from 832.4 to 1.01 Pa.s. According to the authors, this result could be largely attributed to an efficient synergism of complementary activities found on the different enzymes, explaining a much superior effect comparatively to the action of individual extracts. Another example refers to the study reported by Zhang et al. [70] on the application of a xylanase treatment to reduce the viscosity of a sweet potato mash for VHG processes. According to the authors, a xylanases treatment conducted during 90 min resulted into a major reduction of medium viscosity from 9.86 to 0.50 Pa.s, and posteriorly led to an ethanol titer above 17% (v/v) and an ethanol yield of 90.7%. In this regard, the advances in consolidated bioprocessing, which combines saccharolytic and fermentative abilities in a single microorganism, could significantly advance these VHG processes as industrial robust S. cerevisiae cells engineered with cell-surface display xylanases have been constructed [71].
3.3. Nutritional Supplementation
As mentioned above, the depletion on the fermentation medium of key components employed by cells to either grow or to ferment represent one of the main causes for incomplete and/or slow VHG fermentations. Indeed, often the preparation of high sugar content (VHG) mediums is not followed by an adequate balance of other important elements required by cells. Accordingly, the external supply of such elements through the supplementation with complementary sources can represent a viable solution to overcome nutrient starvation stresses.
On this context, several studies have been conducted on the utilization of a wide range of potential supplements, with a growing interest especially given to low-cost nutrient sources, which seems very appealing when framed to the economics of industrial scale processes. One example refers to the studies conducted by Kawa-Rygielska and Pietrzak [72] where VHG maize mashes were supplemented with spent brewer′s yeast. Resulting from that, the authors observed not only a clear improvement on the fermentation rate, where maximum ethanol yields were reached much earlier (at 48 h), but also on maximum ethanol titers, reaching 140 g/L comparing to 120 g/L in the control experiment. Similarly, Suwanapong et al. [73] reported the utilization of dried spent yeast and its hydrolysate as nitrogen supplements in VHG fermentations with sweet sorghum juice. Comparing to a no-supplementation scenario, where 89 g/L of ethanol were produced with a productivity of 1.58 g/L·h, the supplementation of 21 g/L of spent yeast resulted in an ethanol titer of 107 g/L and a productivity of 2.24 g/L·h. The utilization of yeast hydrolysate still resulted in further improvements of productivity, although ethanol titers and yields slightly decreased. Another example refers to an early work conducted by Sridee et al. [50] on the supplementation of a VHG sweet sorghum juice medium employing dried spent yeast as nitrogen source. Comparing to the control condition (no supplement), which produced 93.9 g/L of ethanol with a productivity of 1.96 g/L·h, the addition of 16 g/L of spent yeast led to 104.2 g/L of ethanol and a productivity of 2.17 g/L·h.
Another potential supplement worth discussing is urea, a commonly used low-cost nitrogen source [48]. According to Appiah-Nkansah et al. [74], when VHG fermentations conducted with mixtures of sweet sorghum juice and sorghum starch were supplemented with 16 mM urea, there were noticeable improvements over final ethanol titers and fermentation kinetics. Similarly, a previous study from Yue et al. [75] showed that the addition of 0.8 g/L in nitrogen-equivalent of urea resulted in an increase of ethanol concentration from 120 (control) to 135 g/L although the supplementation of similar levels of ammonium sulfate has not evidenced any benefits. In addition to the previous examples, numerous studies have also explored the application of more expensive components, such as yeast extract [76,77,78] and amino acids [79]. Nevertheless, on an industrial scale context, these will likely seem less attractive and thus have been receiving less and less attention these days.
Even though nitrogen supplementation represents a large part of the studies conducted on this topic, some efforts have also been focusing on other elements equally important for cells. One example refers to the important role played by different ions (e.g., Mg, Cu, Ca, Mn, etc.) as already addressed by the previous studies conducted by Pereira et al. [47], Hu et al. [80], Deesuth et al. [76], among others. Other interesting element especially relevant on a VHG environment is the application of osmoprotectants (e.g., glycine), as previously reported by Chan-u-tit et al. [77].
3.4. Operation Mode
Bioethanol production has been typically conducted by three distinct operation modes: batch, fed-batch, and continuous. According to Lopes et al. [11], fed-batch processes represents nearly 83% of the existing facilities, with the continuous operation only representing 17% of the cases.
3.4.1. Batch Operation
Batch fermentation is a closed culture system where both biomass and substrates are added into the fermenter on a single step, being the products only removed at the end of the process. This may significantly contribute to achieving high cell densities since nearly 99.5% of cells can be reused in a subsequent fermentation. High cell densities inside the fermentation vessel have been reported to contribute to reducing the fermentation time [59] and to increase production yields and productivities [81,82]. High cell densities may also result in reduced ethanol inhibition effects over cells, which will result in superior viabilities [83]. Nevertheless, this configuration has also some disadvantages, particularly when microorganisms are either slow growing or strongly affected by substrate inhibition [84,85]. Batch configuration presents limitations when processing substrates in a concentration of up to 200 g/L of total sugars, which corresponds to a maximum ethanol concentration of 102 g/L. Specifically, it can lead to reduced yields and productivities when the substrate is added at once at the beginning of the process, either by exerting inhibitory/repressing effects or diverting cellular metabolism to products of no interest [86,87].
3.4.2. Fed-Batch
Fed-batch operation usually begins with a batch stage, using a small amount of substrate in the fermenter. Afterwards, the fermentation medium is fed to the fermenter, stepwise or continuously, when most of the initial substrate has been consumed. This allows increasing the total levels of substrate uptake while maintaining a low substrate concentration within the fermentation vessel, thus reducing the negative effects of osmotic pressure or rheology-related constraints associated to highly viscous substrates. Among other advantages, this configuration can allow a reduction of substrate inhibition, higher productivity levels, reduced fermentation times, and a decrease of medium components toxicity, which are present in higher concentrations on this particular context [88].
Stepwise feeding in fed-batch fermentations has already been shown to effectively enhance ethanol production under HG conditions. In an early work from Laopaiboon et al. [89], the fermentation of sweet sorghum juice under HG conditions was assessed following different process configurations. From a 24° Brix medium, the traditional batch configuration resulted in 100 g/L of ethanol and a product yield of 0.42 g/g; using the optimum fed-batch strategy, which consisted in two-times substrate feeding, final ethanol concentration, and production yields increased to 120 g/L and 0.48 g/g, respectively. On the other hand, fed-batch fermentations with continuous feeding may also enable some improvements in fermentation efficiency although these seems highly dependent on numerous operating factors [90]. In a recent work from Phukoetphim et al. [78], the authors studied several fed-batch continuous schemes for the VHG fermentation of sweet-sorghum juice, which consisted of different feeding times and feeding rates. Overall, all feeding strategies did not result in visible improvements in final ethanol concentration. Nonetheless, the application of a specific regime referring to a feeding time of 9 h and a feeding rate of 40 g of sugar/h resulted in a clear improvement on process productivity, raising from 1.56 (batch) to 2.35 g/L·h. Other improvements in the process design and operation, which may allow to enhance the physiological environment of cells under stress, will be also economically interesting and a focus for continuous development.
3.4.3. Continuous Regime
Continuously stirred tank reactors (CSTRs) have been widely used for bioethanol production by both academia and industry for a long time. Although being characterized by their high mixing performance, superior product inhibition may also occur because of the high final product concentrations inside these reactors. On the other hand, although not fully suitable for batch VHG fermentations, multi-stage stirred tank reactors in series can lower product inhibition to some extent. Additionally, tubular bioreactors can also be used in a context of product inhibition since its concentration increases gradually along the axial direction, even though strong substrate inhibition can equally occur at the inlet sections, resulting from the very high substrate concentrations [8]. Independent of the exact reactor configuration—single reactor or a series of multiple vessels—operating under a continuous regime can potentially represent an interesting option under a VHG context, as long as some key issues are surpassed. While a continuous operation would allow to clearly reduce production times, a continuous exposure of yeast cells to high sugar and ethanol concentrations would also possibly affect cells growth, and ultimately lead to biomass washout [91]. Furthermore, for each case, it would be critical to find a proper dilution rate that simultaneously allows reasonable sugar levels on the residual stream and economically competitive ethanol productivities, which would essentially depend on increasing current sugar uptake capacities.
Referring to some examples, an early work by Bayrock and Ingledew [92] reported the production of bioethanol employing a multistage system constituted by a train of 5 sequential vessels. Different mash concentrations were assessed, and accordingly, different dilution rates were established so the glucose residuals were always inferior to 0.3% (w/v). Using a 32% (w/v) glucose mash, this system allowed an ethanol production around 16.73% (v/v); considering the correspondent dilution rate of 0.05 h-1, an ethanol productivity of 6.6 g/L·h can be estimated, which can be considered fairly competitive with other works found in literature. From another report, Bai et al. [9] described the continuous production of bioethanol by S. cerevisiae under VHG conditions, specifically with a medium containing 280 g/L of glucose fed at a dilution rate of 0.012 h-1 (to guarantee over 90% of glucose conversion). Under these conditions, the authors achieved an average ethanol concentration of 15.8% (v/v) over 2 months of operation, which corresponds instead to a very modest productivity around 1.5 g/L·h.
3.5. Cells Immobilization
Bioethanol production is usually conducted employing free cells, where they proliferate in the media and carry out their metabolic functions. However, for this case, the specific growth rate of cells can be affected by many factors related to either product or substrate [93]. To overcome these, as well as enhance ethanol tolerance and promote a reduction of production costs, alternative strategies for bioethanol production have been studied, among which is the application of cells immobilization. This has been explored in bioethanol research during the last several decades, and yet, it still holds much interest in the field [93].
Cells immobilization offers numerous benefits over free cells systems: prolonged cellular stability; increased tolerance to osmotic stress; increased ethanol yield and productivities; reduced end-product inhibition; lower risk of contamination due to high cell densities; inferior energy demand and process costs due to an easier product recovery; reutilization of cells for extended periods, namely by cell recycling in repeated fermentations; cells protection against toxic compounds [94]. Cells of S. cerevisiae, which is the most common microorganism used in fermentation, can be immobilized via two main approaches: immobilization on a physical support; and immobilization by self-flocculation [95].
Commonly known for their high auto-immobilization capacity, self-flocculating strains seems to be superior to those immobilized on a physical support; they are naturally retained inside the reactor (when flocs of an appropriate size are formed) with no visible problems on cell growth, being recovered by a simple sedimentation, rather than using centrifuges [1,38,96]. Different reactor configurations have been tested with these strains including air-lift reactors [97,98,99,100], single-tower reactors [101,102], and two-tower reactors connected in series [103]. The particular use of flocculent yeasts on continuous processes goes back to the 1980s. In these processes, high yields can be reached since it is possible to operate with high cell concentrations (e.g., 45% (v/v)) [101], and consequently, high ethanol titers are achieved in the fermented juice, a major goal of industry [104]. This self-flocculation capacity does not only provide a more convenient way to maintain high cell concentrations on the system but will also prevent contamination [105], besides enabling multiple batch cycles. In a previous study from Li et al. [106], the authors reported the utilization of a self-flocculation yeast of S. cerevisiae to conduct a process of VHG fermentation through several consecutive batches. According to the authors, cells were repeatedly recovered and recycled only by flocculation through a total of 9 batches, which enabled a continuous use of high cell-densities and a consequent reduction of fermentation times. Thus, with a VHG medium containing approximately 255 g/L of glucose, the system allowed an average production of 15% (v/v) ethanol within 8–14 h. As pointed out by the authors, after including cells sedimentation time, an average process productivity of 8.2 g/L·h was obtained, which represent a very attractive value in a VHG context. In another report, Gomes et al. [38] described the development and utilization of a self-flocculation yeast on a repeated batch system for VHG fermentations. Using an air-lift reactor, the engineered flocculent cells were easily recovered and recycled by flocculation-sedimentation throughout a total of 10 batches, enabling an average production of 142 g/L of ethanol per operational batch.
In regards to cells immobilization through a physical support, four main methods are traditionally used: adsorption, crosslinking, encapsulation, and entrapment [107]. Adsorption is one of the most attractive options because of its simplicity and low-cost; through electrostatic forces (e.g., ionic bonds, Van der Waals forces, etc.), cells are adsorbed to a support material, usually not requiring any toxic chemicals [108]. Another interesting technique is cell entrapment on a polymeric matrix, such as calcium alginate. This option has important advantages, among which inferior mass transfer limitations and cells leakage, and the possibility to operate under high dilution rates [109]. On the other hand, with cells encapsulation, they are contained inside a semi-permeable membrane, allowing the diffusion of nutrients and products. Even though presenting a high chemical and mechanical stability and being suitable for high cell loads, this technique can also constrain cell growth due to limitation of capsule dimensions and diffusion rates of important compounds [110].
Among possible materials that can be used as immobilization support one of the most common options is calcium alginate, which present good biocompatibility, low cost, high availability, and an easy preparation. However, calcium alginate beads are also reported for gel degradation, severe mass transfer limitations, low mechanical strength (enabling cells release from the support), and large pore size [111,112]. Other potential support options may refer to sugarcane bagasse, spent grains, corn cobs, zeolites, among others [113]. The utilization of lignocellulosic residues may be found especially attractive as they correspond in many cases to materials with no value, hence very cheap. This selection should consider not only their cost and toxicity but also their performance as efficient mass transfer structures, enabling a proper diffusion of nutrients and products. Traditionally, natural polymers, like sugarcane bagasse and corn cobs, are less expensive and do not present impurities from a chemical synthesis. However, synthetic polymers present a higher stability and resistance to abrasion, but also a superior surface area and permeability [114]. Even though cells immobilization has still not received much attention in this context, ethanol production employing immobilized cells still harbors considerable potential; it can potentiate a lower product and/or substrate inhibition, higher productivities (enabled by increased cell concentrations), the possibility to reuse the cells, and a relatively easy product separation process [112].
3.6. Development of the Fermentation Organism
The development of robust strains more tolerant to the stresses found in VHG fermentations represents one of the key aspects on the improvement of this type of processes. Considerable research has been conducted in this regard over the last decades, which has addressed distinct aspects of VHG-related stresses.
Previous studies conducted by Pereira et al. [115] identified different sets of genes necessary for yeast resistance to multiple fermentation-related stresses, namely high concentrations of glucose and ethanol, both relevant in VHG fermentation. By comparing the fermentative performance of different single-gene deleted strains under VHG conditions, it was possible to infer five genes whose expression is required for maximum performance on VHG fermentations: BUD31 and HPR1 were found to have a crucial effect in both ethanol yield and fermentation rate; PHO85, VRP1, and YGL024w were required for maximal ethanol production. According to the authors, these could represent interesting candidates for further genetic engineering strategies to achieve more robust yeasts. Another example refers to recent work conducted by Hong et al. [116] where the authors truncated the promoter of CYR1 of S. cerevisiae. The obtained mutants, carrying different types of promoter truncation, presented a 2–3-fold decrease in adenylate cyclase activity and showed a superior heat and ethanol tolerance. In VHG fermentation (nearly 276 g/L of glucose) at 40 °C, these mutants enabled a 14–15% increase in the ethanol yield. On a different approach, genome shuffling has been also frequently used as a simple tool for yeast development. For example, Tao et al. [117] combined an initial deletion of GPD2 with three rounds of genome shuffling over S. cerevisiae Z5 strain, resulting in a mutant strain able to achieve a 8% higher ethanol yield. Liu et al. [118] combined an initial step of chemical mutagenesis using EMS (ethyl methanesulfonate) with a meiotic recombination-mediated genome shuffling. According to the authors, under VHG conditions, the resulting S. cerevisiae mutants showed a noticeable increase in osmolarity and ethanol tolerance, enabling a 16% increase on ethanol yields.
Under a slightly distinct strategy, significant improvements can be also achieved by addressing other aspects besides directly targeting yeast cells tolerance. In a recent study by Wang et al. [119], the authors reported that the overexpression of the metB/yfdZ operon in Zymomonas mobilis enabled the mutant strain to grow on a chemically defined medium without amino acids and vitamins, which ultimately lowers the medium cost. Another example consists in the work developed by Guo et al. [120] on the expression of an aspartic protease from Neurospora crassa over an ethanol producing yeast (cell wall-anchored) to obtain process-relevant traits. The mutant strain not only showed a superior cell viability and growth rate, but also a 7.2% superior ethanol yield. According to the authors, this could be attributed to an improvement of starch saccharification led by the lysing effect associated to the proteases, but also to the positive nutritional effect of the amino acids released during proteases action.
3.7. VHG Fermentation for Cellulosic Ethanol Production
The application of VHG fermentation to produce bioethanol is typically associated with energy crops, or sucrose/starch materials in general, having a high carbohydrates content. Nonetheless, this technology may also be considered for another important class of feedstocks known as lignocellulosic materials (LCMs). These are complex materials found abundantly in nature, which are mainly composed by cellulose, hemicellulose, lignin, and other minor components, like ash, protein, and fat [121]. Because their high availability and diversity of possible sources, they are usually cheaper and do not compete with food crops. However, when comparing with traditional feedstocks of 1G-bioethanol, VHG operation with LCMs is much more challenging due to several factors. Firstly, LCMs typically present large fractions of insoluble components that are not sugars (e.g., ash, lignin) [122], resulting on a dilution of its sugar fraction. Because of that, and since the total carbohydrates fraction is in many cases inferior to 50%, reaching an initial sugar concentration above 250 g/L can frequently require using an initial hydrolysis slurry with at least 50% (w/v) of solids. On the other hand, this level of solids consistency cannot be achieved with LCMs because they traditionally have low densities and high water-retentions capacities [123], originating high-viscosity solutions and mass transfer limitations when solids content raises above 18–20% [124]. Adding to rheology-related issues, a VHG operation will also bring a superior concentration of sugar-derived inhibitors formed during solid pre-treatment such as HMF, furfural, and acetic acid, which have been widely reported to affect both cells and enzymes [125]. Similarly, lignin and derivatives will be also present in superior amounts, with several consequences in the process. Phenolic compounds originated from lignin, such as ferulic and p-coumaric acid, have been reported to inhibit cell growth and fermentation [126,127]. Furthermore, lignin has been associated to a strong inhibition of cellulases action towards cellulose, either by forming a physical barrier to cellulose [122] or due to a non-productive binding of the enzymes to lignin [128], affecting their adsorption to cellulose.
Some efforts have been conducted towards a partial/total elimination of the effects from these compounds, which may involve the application of detoxification steps, the utilization of more robust organisms, the utilization of more adequate pre-treatments, among others [124]. A potentially effective strategy that may not only decrease inhibitors toxicity but also attenuate medium rheology constraints refers to the application of alternative operation modes such as the fed-batch process [129], which has already been discussed above in the context of a reduction of osmolarity-related stress (c.f. Section 3.4.2). Gradually feeding the LCM substrate will allow, in some cases, that the fermentation organism could gradually convert some of the inhibitors present on the medium (e.g., furfural and HMF) [130] and/or an adaptation to them [131]. On the other hand, a fed-batch operation would also guarantee a constant availability of appropriate levels of free water as the LCM suspension is continuously liquefied [132,133], a critical element for an efficient mobility of cellulases [124]. This opposes a single addition of the LCM, where reduced levels of free water can drastically affect enzymes action.
Overall, the application of VHG technologies to efficiently process lignocellulosic materials faces multiples challenges and will still possibly require considerable research efforts in the upcoming years. This may explain why studies of VHG in the context of LCMs are currently very scarce and/or still not present attractive indicators.
4. Concluding Remarks
Although already been studied over several decades, very-high gravity technology is still the subject of considerable research nowadays. Overall, and despite the substantial amount of studies conducted so far, many technical hurdles remain. Among these, special attention has been given to the physiological stresses undertaken by cells, either from high initial sugar levels or from high final ethanol concentrations. Additionally, nutrients starvation has been also commonly found in VHG processes referring to, in many cases, to the cause of incomplete and inefficient fermentations. In the face of that, many current research efforts have been focused on optimizing the supplementation of fermentation media, which has mostly relied on the search of new low-cost nutrients able to meet the core needs of the fermentation organism. In respect to the fermentation organism, special attention has been dedicated to the development and screening of more robust organisms towards the multiple stresses found in a characteristically challenging environment. Additionally, alternative process designs such as fed-batch or continuous operation, or using immobilized cells, has been also considered. Finally, to meet a growing utilization of new and more economic substrates, many of them raising new operational challenges to the process, recent research has been also focused on new agitation apparatus and/or substrate feeding schemes. As long as some of these issues are efficiently addressed, VHG technology will likely contribute to significant economic improvements in biofuels production and for their definitive affirmation as a viable alternative to fossil fuels and sustainable solution for simultaneous multi-waste valorization processes.
Author Contributions
Conceptualization, D.G. and L.D.; Writing—Original Draft Preparation, D.G. and M.C.; Writing—review & editing, D.G., M.C., M.d.R., E.R., J.T. and L.D. All authors have read and agreed to the published version of the manuscript.
Funding
This work has been carried out at the Biomass and Bioenergy Research Infrastructure (BBRI)-LISBOA-01-0145-FEDER-022059, supported by Operational Programme for Competitiveness and Internationalization (PORTUGAL2020), by Lisbon Portugal Regional Operational Programme (Lisboa 2020) and by North Portugal Regional Operational Program (Norte 2020) under the Portugal 2020 Partnership Agreement, through the European Regional Development Fund (ERDF) and has been supported by the Portuguese Foundation for Science and Technology (FCT) under the scope of the strategic funding of UIDB/04469/2020 and through Project EcoTech (POCI-01-0145-FEDER-032206/FAPESP 2018/07522-6) funded by the European Regional Development Fund under the scope of Norte2020—Programa Operacional Regional do Norte. The authors also acknowledge the financial support of Research Support Foundation of the state of Minas Gerais-FAPEMIG, National Counsel of Technological and Scientific Development-CNPq and Coordination for the Improvement of Higher Education Personnel-CAPES (Finance Code 001).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analyzed in this study. Data sharing is not applicable to this article.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bai, F.W.; Anderson, W.A.; Moo-Young, M. Ethanol fermentation technologies from sugar and starch feedstocks. Biotechnol. Adv. 2008, 26, 89–105. [Google Scholar] [CrossRef] [PubMed]
- Deesuth, A.; Laopaiboon, P.; Klanrit, P.; Laopaiboon, L. Improvement of ethanol production from sweet sorghum juice under high gravity and very high gravity conditions: Effect of nutrient supplementation and aeration. Ind. Crops Prod. 2015, 74, 95–102. [Google Scholar] [CrossRef]
- Arshad, M.; Hussain, T.; Iqbal, M.; Abbas, M. Enhanced ethanol production at commercial scale from molasses using high gravity technology by mutant S. cerevisiae. Braz. J. Microbiol. 2017, 48, 403–409. [Google Scholar] [CrossRef]
- Thomas, K.C.; Hynes, S.H.; Jones, A.M.; Ingledew, W.M. Production of fuel alcohol from wheat by VHG technology: Effect of sugar concentration and fermentation temperature. Appl. Biochem. Biotechnol. 1993, 43, 211–226. [Google Scholar] [CrossRef]
- Wang, F.Q.; Gao, C.J.; Yang, C.Y.; Xu, P. Optimization of an ethanol production medium in very high gravity fermentation. Biotechnol. Lett. 2007, 29, 233–236. [Google Scholar] [CrossRef]
- Maiorella, B.; Blanch, H.; Wilke, C.; Wyman, C.E. Economic evaluation of alternative ethanol fermentation processes. Biotechnol. Bioeng. 2009, 104, 419–443. [Google Scholar] [CrossRef]
- Wang, S.; Thomas, K.C.; Sosulski, K.; Ingledew, W.M. Grain pearling and very high gravity (VHG) fermentation technologies for fuel alcohol production from rye and triticale. Process Biochem. 1999, 34, 421–428. [Google Scholar] [CrossRef]
- Bai, F.W.; Chen, L.J.; Anderson, W.A.; Moo-Young, M. Parameter oscillations in very high gravity medium continuous ethanol fermentation and their attenuation on multi-stage packed column bioreactor system. Biotechnol. Bioeng. 2004, 88, 558–566. [Google Scholar] [CrossRef] [PubMed]
- Bai, F.W.; Chen, L.J.; Zhang, Z.; Anderson, W.A.; Moo-Young, M. Continuous ethanol production and evaluation of yeast cell lysis and viability loss under very high gravity medium conditions. J. Biotechnol. 2004, 110, 287–293. [Google Scholar] [CrossRef]
- Reis, V.R.; Bassi, A.P.G.; da Silva, J.C.G.; Ceccato-Antonini, S.R. Characteristics of Saccharomyces cerevisiae yeasts exhibiting rough colonies and pseudohyphal morphology with respect to alcoholic fermentation. Braz. J. Microbiol. 2013, 44, 1121–1131. [Google Scholar] [CrossRef] [Green Version]
- Lopes, M.L.; Paulillo, S.C.L.; Godoy, A.; Cherubin, R.A.; Lorenzi, M.S.; Giometti, F.H.C.; Bernardino, C.D.; Neto, H.B.A.; Amorim, H.V. Ethanol production in Brazil: A bridge between science and industry. Braz. J. Microbiol. 2016, 47, 64–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gibson, B.R.; Lawrence, F.M.; Leclaire, J.P.R.; Powell, C.D.; Smart, K.A. Yeast responses to stresses associated with industrial brewery handling. FEMS Microbiol. Rev. 2007, 31, 535–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mira, N.P.; Palma, M.; Guerreiro, J.F.; Sá-Correia, I. Genome-wide identification of Saccharomyces cerevisiae genes required for tolerance to acetic acid. Microb. Cell Fact. 2010, 9, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, M.C.; Mira, N.P.; Sá-Correia, I. A genome-wide perspective on the response and tolerance to food-relevant stresses in Saccharomyces cerevisiae. Curr. Opin. Biotechnol. 2011, 22, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Estruch, F. Stress-controlled transcription factor, stress-induced genes and stress tolerance in budding yeast. FEMS Microbiol. Rev. 2000, 24, 469–486. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, S. Osmotic stress signaling and osmoadaptation in yeasts. Microbiol. Mol. Biol. Rev. 2002, 66, 300–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hounsa, C.G.; Brandt, E.V.; Trevelein, J.; Hohmann, S.; Prior, B.A. Role of trehalose in survival of Saccharomyces cerevisiae under osmotic stress. Microbiology 1998, 144, 671–680. [Google Scholar] [CrossRef]
- Panek, A.C.; Vânia, J.J.M.; Paschoalin, M.F.; Panek, D. Regulation of trehalose metabolism in Saccharomyces cerevisiae mutants during temperature shifts. Biochimie 1990, 72, 77–79. [Google Scholar] [CrossRef]
- D’Amore, T.; Crumplen, R.; Stewart, G.G. The involvement of trehalose in yeast stress tolerance. J. Ind. Microbiol. 1991, 7, 191–195. [Google Scholar] [CrossRef]
- Stanley, D.; Bandara, A.; Fraser, S.; Chambers, P.J.; Stanley, G.A. The ethanol stress response and ethanol tolerance of Saccharomyces cerevisiae. J. Appl. Microbiol. 2010, 109, 13–24. [Google Scholar] [CrossRef]
- Casey, G.P.; Ingledew, W.M. Ethanol tolerance in yeasts. Crit. Rev. Microbiol. 1986, 13, 219–280. [Google Scholar] [CrossRef]
- Hu, X.H.; Wang, M.H.; Tan, T.; Li, J.R.; Yang, H.; Leach, L.; Zhang, R.M.; Luo, Z.W. Genetic dissection of ethanol tolerance in the budding yeast Saccharomyces cerevisiae. Genetics 2007, 175, 1479–1487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, P.; Prasad, R. Relationship between ethanol tolerance and fatty acyl composition of Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 1989, 30, 294–298. [Google Scholar] [CrossRef]
- Piper, P.W.; Talreja, K.; Panaretou, B.; Moradas-Ferreira, P.; Byrne, K.; Praekelt, U.M.; Meacock, P.; Récnacq, M.; Boucherie, H. Induction of major heat-shock proteins of Saccharomyces cerevisiae, including plasma membrane Hsp30, by ethanol levels above a critical threshold. Microbiology 1994, 140 Pt 11, 3031–3038. [Google Scholar] [CrossRef] [Green Version]
- Thomas, K.C.; Ingledew, W.M. Production of 21% (v/v) ethanol by fermentation of very high gravity (VHG) wheat mashes. J. Ind. Microbiol. 1992, 10, 61–68. [Google Scholar] [CrossRef]
- Pereira, F.B.; Guimarães, P.M.R.; Teixeira, J.A.; Domingues, L. Robust industrial Saccharomyces cerevisiae strains for very high gravity bio-ethanol fermentations. J. Biosci. Bioeng. 2011, 112, 130–136. [Google Scholar] [CrossRef] [Green Version]
- Paulillo, S.C.L.; Yokoya, F.; Basso, L.C. Mobilization of endogenous glycogen and trehalose of industrial yeasts. Braz. J. Microbiol. 2003, 34, 249–254. [Google Scholar] [CrossRef] [Green Version]
- François, J.M.; Walther, T.; Parrou, J.L. Genetics and Regulation of Glycogen and Trehalose Metabolism in Saccharomyces cerevisiae. In Microbial Stress Tolerance for Biofuels—Microbiology Monographs; Liu, Z., Ed.; Springer: Berlin/Heidelberg, Germany, 2012; Volume 22, pp. 29–55. [Google Scholar]
- Lip, K.; García-Ríos, E.; Costa, C.E.; Guillamón, J.M.; Domingues, L.; Teixeira, J.; van Gulik, W.M. Selection and subsequent physiological characterization of industrial Saccharomyces cerevisiae strains during continuous growth at sub- and- supra optimal temperatures. Biotechnol. Rep. 2020, 26, e00462. [Google Scholar] [CrossRef]
- Pinheiro, T.; Lip, K.; García-Ríos, E.; Querol, A.; Teixeira, J.; van Gulik, W.; Guillamón, J.M.; Domingues, L. Differential proteomic analysis by SWATH-MS unravels the most dominant mechanisms underlying yeast adaptation to non-optimal temperatures under anaerobic conditions. Sci. Rep. 2020, 10, 22329. [Google Scholar] [CrossRef] [PubMed]
- Tofighi, A.; Assadi, M.M.; Asadirad, M.H.A.; Karizi, S.Z. Bio-ethanol production by a novel autochthonous thermo-tolerant yeast isolated from wastewater. J. Environ. Health Sci. Eng. 2014, 12, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Wang, B.; Liang, X. Enhanced ethanol production from stalk juice of sweet sorghum by response surface methodology. Afr. J. Biotechnol. 2012, 11, 6117–6122. [Google Scholar]
- Phutela, U.G.; Kaur, J. Process optimization for ethanol production from sweet sorghum juice using Saccharomyces cerevisiae strain NRRL Y-2034 by response surface methodology. Sugar Tech. 2014, 16, 411–421. [Google Scholar] [CrossRef]
- Pattanakittivorakul, S.; Lertwattanasakul, N.; Yamada, M.; Limtong, S. Selection of thermotolerant Saccharomyces cerevisiae for high temperature ethanol production from molasses and increasing ethanol production by strain improvement. Antonie Leeuwenhoek 2019, 112, 975–990. [Google Scholar] [CrossRef]
- Techaparin, A.; Thanonkeo, P.; Klanrit, P. High-temperature ethanol production using thermotolerant yeast newly isolated from Greater Mekong Subregion. Braz. J. Microbiol. 2017, 48, 461–475. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhao, H.; Gan, M.; Jin, Y.; Gao, X.; Chen, Q.; Guan, J.; Wang, Z. Application of simultaneous saccharification and fermentation (SSF) from viscosity reducing of raw sweet potato for bioethanol production at laboratory, pilot and industrial scales. Bioresour. Technol. 2011, 102, 4573–4579. [Google Scholar] [CrossRef] [PubMed]
- Cruz, M.L.; de Resende, M.M.; Ribeiro, E.J. Improvement of ethanol production in fed-batch fermentation using a mixture of sugarcane juice and molasse under very high-gravity conditions. Bioprocess Biosyst. Eng. 2021, 44, 617–625. [Google Scholar] [CrossRef]
- Gomes, D.G.; Guimarães, P.M.R.; Pereira, F.B.; Teixeira, J.A.; Domingues, L. Plasmid-mediate transfer of FLO1 into industrial Saccharomyces cerevisiae PE-2 strain creates a strain useful for repeat-batch fermentations involving flocculation–sedimentation. Bioresour. Technol. 2012, 108, 162–168. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Jin, Y.-L.; Fang, Y.; Zhao, H. Adaptive evolution and selection of stress-resistant Saccharomyces cerevisiae for very high gravity bioethanol fermentation. Electron. J. Biotechnol. 2019, 41, 88–94. [Google Scholar] [CrossRef]
- Kim, J.H.; Ryu, J.; Huh, I.Y. Ethanol production from galactose by a newly isolated Saccharomyces cerevisiae KL17. Bioprocess Biosyst. Eng. 2014, 37, 1871–1878. [Google Scholar] [CrossRef]
- Joannis-Cassan, C.; Riess, J.; Jolibert, F.; Taillandier, P. Optimization of very high gravity fermentation process for ethanol production from industrial sugar beet syrup. Biomass Bioenergy 2014, 70, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Bafrncová, P.; šmogrovičová, D.; Sláviková, I.; Pátková, J.; Dömény, Z. Improvement of very high gravity ethanol fermentation by media supplementation using Sacchromyces serevisiae. Biotechnol. Lett. 1999, 21, 337–341. [Google Scholar] [CrossRef]
- Laopaiboon, L.; Nuanpeng, S.; Srinophakun, P.; Klanrit, P.; Laopaiboon, P. Ethanol production from sweet sorghum juice using very high gravity technology: Effects of carbon and nitrogen supplementations. Bioresour. Technol. 2009, 100, 4176–4182. [Google Scholar] [CrossRef]
- Khongsay, N.; Laopaiboon, L.; Jaisil, P.; Laopaiboon, P. Optimization of agitation and aeration for very high gravity ethanol fermentation from sweet sorghum juice by Saccharomyces cerevisiae using an orthogonal array design. Energies 2012, 5, 561–576. [Google Scholar] [CrossRef] [Green Version]
- Reddy, L.V.A.; Reddy, O.V.S. Improvement of ethanol production in very high gravity fermentation by horse gram (Dolichos biflorus) flour supplementation. Lett. Appl. Microbiol. 2005, 41, 440–444. [Google Scholar] [CrossRef]
- Reddy, L.V.A.; Reddy, O.V.S. Rapid and enhanced production of ethanol in very high gravity (VHG) sugar fermentations by Saccharomyces cerevisiae: Role of finger millet (Eleusine coracana L.) flour. Process Biochem. 2006, 41, 726–729. [Google Scholar] [CrossRef]
- Pereira, F.B.; Guimarães, P.M.R.; Teixeira, J.A.; Domingues, L. Optimization of low-cost medium for very high gravity ethanol fermentations by Saccharomyces cerevisiae using statistical experimental designs. Bioresour. Technol. 2010, 101, 7856–7863. [Google Scholar] [CrossRef] [Green Version]
- Kelbert, M.; Romaní, A.; Coelho, E.; Pereira, F.B.; Teixeira, J.A.; Domingues, L. Lignocellulosic bioethanol production with revalorization of low-cost agroindustrial by-products as nutritional supplements. Ind. Crops Prod. 2015, 64, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Jones, A.M.; Ingledew, W.M. Fuel alcohol production: Optimization of temperature for efficient very-high-gravity fermentation. Appl. Microbiol. Biotechnol. 1994, 60, 1048–1051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sridee, W.; Laopaiboon, L.; Jaisil, P.; Laopaiboon, P. The use of dried spent yeast as a low-cost nitrogen supplement in ethanol fermentation from sweet sorghum juice under very high gravity conditions. Electron. J. Biotechnol. 2011, 14, 1–15. [Google Scholar]
- Palukurty, M.A.; Telgana, N.K.; Bora, H.S.R.; Mulampaka, S.N. Screening and optimization of metal ions to enhance ethanol production using statistical experimental designs. Afr. J. Microbiol. Res. 2008, 2, 87–94. [Google Scholar]
- Xue, C.; Zhao, X.Q.; Yuan, W.J.; Bai, F.W. Improving ethanol tolerance of a self-flocculating yeast by optimization of medium composition. World J. Microbiol. Biotechnol. 2008, 24, 2257–2261. [Google Scholar] [CrossRef]
- Zhao, X.Q.; Bai, F.W. Yeast flocculation: New story in fuel ethanol production. Biotechnol. Adv. 2009, 27, 849–856. [Google Scholar] [CrossRef]
- Trofimova, Y.; Walker, G.; Rapoport, A. Anhydrobiosis in yeast: Influence of calcium and magnesium ions on yeast resistance to dehydration–rehydration. FEMS Microbiol. Lett. 2010, 308, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Courchesne, W.E.; Vlasek, C.; Klukovich, R.; Coffee, S. Ethanol induces calcium influx via the Cch1-Mid1 transporter in Saccharomyces cerevisiae. Arch. Microbiol. 2011, 193, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.W.; Oliver, S.G. The effect of temperature on the ethanol tolerance of the yeast Saccharomyces uvarum. Biotechnol. Lett. 1982, 4, 269–273. [Google Scholar] [CrossRef]
- Patrascu, E.; Rapeanu, G.; Hopulele, T. Current approaches to efficient biotechnological production of ethanol. Innovative Rom. Food Biotechnol. 2009, 4, 1–11. [Google Scholar]
- Gao, C.; Fleet, G. The effects of temperature and pH on the ethanol tolerance of the wine yeasts, Saccharomyces cerevisiae, Candida stellata and Kloeckera apiculata. J. Appl. Bacteriol. 1988, 65, 405–409. [Google Scholar] [CrossRef]
- Pereira, F.B.; Gomes, D.G.; Guimarães, P.M.; Teixeira, J.A.; Domingues, L. Cell recycling during repeated very high gravity bio-ethanol fermentations using the industrial Saccharomyces cerevisiae strain PE-2. Biotechnol. Lett. 2012, 34, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Laluce, C.; Tognolli, J.O.; de Oliveira, K.F.; Souza, C.S.; Morais, M.R. Optimization of temperature, sugar concentration, and inoculum size to maximize ethanol production without significant decrease in yeast cell viability. Appl. Microbiol. Biotechnol. 2009, 83, 627–637. [Google Scholar] [CrossRef]
- Limtong, S.; Sringiew, C.; Yongmanitchai, W. Production of fuel ethanol at high temperature from sugar cane juice by a newly isolated Kluyveromyces marxianus. Bioresour. Technol. 2007, 98, 3367–3374. [Google Scholar] [CrossRef]
- Malairuang, K.; Krajang, M.; Rotsattarat, R.; Chamsart, S. Intensive Multiple Sequential Batch Simultaneous Saccharification and Cultivation of Kluyveromyces marxianus SS106 Thermotolerant Yeast Strain for Single-Step Ethanol Fermentation from Raw Cassava Starch. Processes 2020, 8, 898. [Google Scholar] [CrossRef]
- Rivera, E.C.; Costa, A.C.; Atala, D.I.P.; Maugeri, F.; Maciel, M.R.W.; Filho, R.M. Evaluation of optimization techniques for parameter estimation: Application to ethanol fermentation considering the effect of temperature. Process Biochem. 2006, 41, 1682–1687. [Google Scholar] [CrossRef]
- You, K.M.; Rosenfield, C.L.; Knipple, D.C. Ethanol tolerance in the yeast Saccharomyces cerevisiae is dependent on cellular oleic acid content. Appl. Environ. Microbiol. 2003, 69, 1499–1503. [Google Scholar] [CrossRef] [Green Version]
- Visser, W.; Scheffers, W.A.; Batenburg-van der Vegte, W.H.; van Dijken, J.P. Oxygen requirements of yeasts. Appl. Environ. Microbiol. 1990, 56, 3785–3792. [Google Scholar] [CrossRef] [Green Version]
- Rosenfeld, E.; Beauvoit, B.; Blondin, B.; Salmon, J.M. Oxygen consumption by anaerobic Saccharomyces cerevisiae under enological conditions: Effect on fermentation kinetics. Appl. Environ. Microbiol. 2003, 69, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Jouhten, P.; Rintala, E.; Huuskonen, A.; Tamminen, A.; Toivari, M.; Wiebe, M.; Ruohonen, L.; Penttilä, M.; Maaheimo, H. Oxygen dependence of metabolic fluxes and energy generation of Saccharomyces cerevisiae CEN.PK113-1A. BMC Syst. Biol. 2008, 2, 60. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-H.; Chien, W.-S.; Duan, K.-J.; Chang, P.R. Effect of aeration timing and interval during very-high-gravity ethanol fermentation. Process Biochem. 2011, 46, 1025–1028. [Google Scholar] [CrossRef]
- Poonsrisawat, A.; Wanlapatit, S.; Wansuksri, R.; Piyachomkwan, K.; Paemanee, A.; Gamonpilas, C.; Eurwilaichitr, L.; Champreda, V. Synergistic effects of cell wall degrading enzymes on rheology of cassava root mash. Process Biochem. 2016, 51, 2104–2111. [Google Scholar] [CrossRef]
- Zhang, L.; Chen, Q.; Jin, Y.; Xue, H.; Guan, J.; Wang, Z.; Zhao, H. Energy-saving direct ethanol production from viscosity reduction mash of sweet potato at very high gravity (VHG). Fuel Process. Technol. 2010, 91, 1845–1850. [Google Scholar] [CrossRef]
- Cunha, J.T.; Romaní, A.; Inokuma, K.; Johansson, B.; Hasunuma, T.; Kondo, A.; Domingues, L. Consolidated bioprocessing of corn cob-derived hemicellulose: Engineered industrial Saccharomyces cerevisiae as efficient whole cell biocatalysts. Biotechnol. Biofuels 2020, 13, 138. [Google Scholar] [CrossRef] [PubMed]
- Kawa-Rygielska, J.; Pietrzak, W. Ethanol fermentation of very high gravity (VHG) maize mashes by Saccharomyces cerevisiae with spent brewer’s yeast supplementation. Biomass Bioenergy 2014, 60, 50–57. [Google Scholar] [CrossRef]
- Suwanapong, S.; Khongsay, N.; Laopaiboon, L.; Jaisil, P.; Laopaiboon, P. Dried Spent Yeast and Its Hydrolysate as Nitrogen Supplements for Single Batch and Repeated-Batch Ethanol Fermentation from Sweet Sorghum Juice. Energies 2013, 6, 1618–1631. [Google Scholar] [CrossRef]
- Appiah-Nkansah, N.B.; Zhang, K.; Rooney, W.; Wang, D. Ethanol production from mixtures of sweet sorghum juice and sorghum starch using very high gravity fermentation with urea supplementation. Ind. Crops Prod. 2018, 111, 247–253. [Google Scholar] [CrossRef]
- Yue, G.; Yu, J.; Zhang, X.; Tan, T. The influence of nitrogen sources on ethanol production by yeast from concentrated sweet sorghum juice. Biomass Bioenergy 2012, 39, 48–52. [Google Scholar] [CrossRef]
- Deesuth, O.; Laopaiboon, P.; Jaisil, P.; Laopaiboon, L. Optimization of Nitrogen and Metal Ions Supplementation for Very High Gravity Bioethanol Fermentation from Sweet Sorghum Juice Using an Orthogonal Array Design. Energies 2012, 5, 3178–3197. [Google Scholar] [CrossRef]
- Chan-u-tit, P.; Laopaiboon, L.; Jaisil, P.; Laopaiboon, P. High Level Ethanol Production by Nitrogen and Osmoprotectant Supplementation under Very High Gravity Fermentation Conditions. Energies 2013, 6, 884–899. [Google Scholar] [CrossRef] [Green Version]
- Phukoetphim, N.; Salakkam, A.; Laopaiboon, P.; Laopaiboon, L. Improvement of ethanol production from sweet sorghum juice under batch and fed-batch fermentations: Effects of sugar levels, nitrogen supplementation, and feeding regimes. Electron. J. Biotechnol. 2017, 26, 84–92. [Google Scholar] [CrossRef]
- Yang, H.; Zong, X.; Cui, C.; Mu, L.; Zhao, H. Peptide (Lys-Leu) and amino acids (Lys and Leu) supplementations improve physiological activity and fermentation performance of brewer’s yeast during very high-gravity (VHG) wort fermentation. Biotechnol. Appl. Biochem. 2018, 65, 630–638. [Google Scholar] [CrossRef]
- Hu, C.; Qin, Q.; Gao, P. Medium optimization for improved ethanol production in very high gravity fermentation. Chin. J. Chem. Eng. 2011, 19, 1017–1022. [Google Scholar] [CrossRef]
- Liu, C.G.; Wang, N.; Lin, Y.H.; Bai, F.W. Very high gravity ethanol fermentation by flocculating yeast under redox potential-controlled conditions. Biotechnol. Biofuels 2012, 5, 61. [Google Scholar] [CrossRef] [Green Version]
- Westman, J.O.; Franzén, C.J. Current progress in high cell density yeast bioprocesses for bioethanol production. Biotechnol. J. 2015, 10, 1185–1195. [Google Scholar] [CrossRef] [PubMed]
- Vega, J.L.; Navarro, A.R.; Clausen, E.C.; Gaddy, J.L. Effects of inoculum size on ethanol inhibition modeling and other fermentation parameters. Biotechnol. Bioeng. 1987, 29, 633–638. [Google Scholar] [CrossRef]
- Srivastava, A.K.; Gupta, S. 2.38—Fed-Batch Fermentation—Design Strategies. In Comprehensive Biotechnology, 2nd ed.; Moo-Young, M., Ed.; Academic Press: Cambridge, MA, USA, 2011; pp. 515–526. [Google Scholar]
- Stanbury, P.F.; Whitaker, A.; Hall, S.J. Chapter 2—Microbial growth kinetics. In Principles of Fermentation Technology, 3rd ed.; Stanbury, P.F., Whitaker, A., Hall, S.J., Eds.; Butterworth-Heinemann: Oxford, UK, 2017; pp. 21–74. [Google Scholar]
- Rolland, F.; Winderickx, J.; Thevelein, J.M. Glucose-sensing and -signalling mechanisms in yeast. FEMS Yeast Res. 2002, 2, 183–201. [Google Scholar] [CrossRef]
- Gomar-Alba, M.; Morcillo-Parra, M.Á.; Olmo, M.L. Response of yeast cells to high glucose involves molecular and physiological differences when compared to other osmostress conditions. FEMS Yeast Res. 2015, 15, fov039. [Google Scholar] [CrossRef] [PubMed]
- Stanbury, P.F.; Whitaker, A.; Hall, S.J. Principles of Fermentation Technology; Butterworth-Heinemann: Oxford, UK, 1995. [Google Scholar]
- Laopaiboon, L.; Thanonkeo, P.; Jaisil, P.; Laopaiboon, P. Ethanol production from sweet sorghum juice in batch and fed-batch fermentations by Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 2007, 23, 1497–1501. [Google Scholar] [CrossRef]
- Cruz, L.M.; Resende, M.M.; Ribeiro, E.J. Evaluation of process conditions in the performance of yeast on alcoholic fermentation. Chem. Eng. Commun. 2018, 205, 846–855. [Google Scholar] [CrossRef]
- Puligundla, P.; Smogrovicova, D.; Mok, C.; Obulam, V.S.R. A review of recent advances in high gravity ethanol fermentation. Renew. Energy 2019, 133, 1366–1379. [Google Scholar] [CrossRef]
- Bayrock, D.P.; Ingledew, W.M. Application of multistage continuous fermentation for production of fuel alcohol by very-high-gravity fermentation technology. J. Ind. Microbiol. Biotechnol. 2001, 27, 87–93. [Google Scholar] [CrossRef]
- Mishra, A.; Sharma, A.K.; Sharma, S.; Bagai, R.; Mathur, A.S.; Gupta, R.P.; Tuli, D.K. Lignocellulosic ethanol production employing immobilized Saccharomyces cerevisiae in packed bed reactor. Renew. Energy 2016, 98, 57–63. [Google Scholar] [CrossRef]
- Soares, E.V. Flocculation in Saccharomyces cerevisiae: A review. J. Appl. Microbiol. 2010, 110, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kourkoutas, Y.; Bekatorou, A.; Banat, I.M.; Marchant, R.; Koutinas, A. Immobilization technologies and support materials suitable in alcohol beverages production: A review. Food Microbiol. 2004, 21, 377–397. [Google Scholar] [CrossRef]
- Brandao, A.C.T.; de Resende, M.M.; Ribeiro, E.J. Alcoholic fermentation with high sugar and cell concentrations at moderate temperatures using flocculant yeasts. Korean J. Chem. Eng. 2020, 37, 1717–1725. [Google Scholar] [CrossRef]
- Domingues, L.; Dantas, M.M.; Lima, N.; Teixeira, J.A. Continuous ethanol fermentation of lactose by a recombinant flocculating Saccharomyces cerevisiae strain. Biotechnol. Bioeng. 1999, 64, 692–697. [Google Scholar] [CrossRef]
- Domingues, L.; Lima, N.; Teixeira, J.A. Alcohol production from cheese whey permeate using genetically modified flocculent yeast cells. Biotechnol. Bioeng. 2001, 72, 507–514. [Google Scholar] [CrossRef]
- Domingues, L.; Lima, N.; Teixeira, J.A. Aspergillus niger β-galactosidase production by yeast in a continuous high cell density reactor. Process Biochem. 2005, 40, 1151–1154. [Google Scholar] [CrossRef] [Green Version]
- Klein, J.; Maia, J.; Vicente, A.A.; Domingues, L.; Teixeira, J.A.; Jurascík, M. Relationships between hydrodynamics and rheology of flocculating yeast suspensions in a high-cell-density airlift bioreactor. Biotechnol. Bioeng. 2005, 89, 393–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrietta, S.R.; Seckelberg, C.; Andrietta, M.G.S. Study of flocculent yeast performance in tower reactors for bioethanol production in a continuous fermentation process with no cell recycling. Bioresour. Technol. 2008, 99, 3002–3008. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, T.F.; Júnior, W.G.M.; Guidini, C.Z.; Marquez, L.D.S.; Cardoso, V.L.; Resende, M.M.; Ribeiro, E.J. Alcoholic fermentation with self-flocculating yeast in a tower upflow reactor. Chem. Eng. Technol. 2015, 38, 345–354. [Google Scholar] [CrossRef]
- Viegas, M.C.; Andrietta, M.G.S.; Andrietta, S.R. Use of tower reactors for continuous ethanol production. Braz. J. Chem. Eng. 2002, 2, 167–173. [Google Scholar] [CrossRef] [Green Version]
- Landaeta, R.; Aroca, G.; Acevedo, F.; Teixeira, J.A.; Mussatto, S.I. Adaptation of a flocculent Saccharomyces cerevisiae strain to lignocellulosic inhibitors by cell recycle batch fermentation. Appl. Energy 2013, 102, 124–130. [Google Scholar] [CrossRef] [Green Version]
- Domingues, L.; Lima, N.; Teixeira, J.A. Contamination of a high-cell-density continuous bioreactor. Biotechnol. Bioeng. 2000, 68, 584–587. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Zhao, X.Q.; Ge, X.M.; Bai, F.W. An innovative consecutive batch fermentation process for very high gravity ethanol fermentation with self-flocculating yeast. Appl. Microbiol. Biotechnol. 2009, 84, 1079–1086. [Google Scholar] [CrossRef]
- Azhar, S.H.M.; Abdulla, R.; Jambo, S.A.; Marbawi, H.; Gansau, J.A.; Faik, A.A.M.; Rodrigues, K.F. Yeasts in sustainable bioethanol production: A review. Biochem. Biophys. Rep. 2017, 10, 52–61. [Google Scholar]
- Margaritis, A.; Merchant, F.J.A. The technology of anaerobic yeast growth. In Yeast Biotechnology; Berry, D.R., Russell, B.I., Stewart, G.G., Eds.; Springer: Dordrecht, The Netherlands, 1987; pp. 37–53. [Google Scholar]
- Chibata, I.; Wingard, L.B. Immobilized Microbial Cells: Applied Biochemistry and Bioengineering, 4th ed.; Academic Press: Oxford, UK, 1983; pp. 203–215. [Google Scholar]
- Ylitervo, P.; Franzen, C.J.; Taherzadeh, M.J. Ethanol production at elevated temperatures using encapsulation of yeast. J. Biotechnol. 2011, 156, 22–29. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Li, G.; Li, Y. Immobilization of Saccharomyces cerevisiae alcohol dehydrogenase on hybrid alginate-chitosan beads. Int. J. Biol. Macromol. 2010, 47, 21–26. [Google Scholar] [CrossRef]
- Bangrak, P.; Limtong, S.; Phisalaphong, M. Continuous ethanol production using immobilized yeast cells entrapped in loofa-reinforced alginate carriers. Braz. J. Microbiol. 2011, 42, 676–684. [Google Scholar] [CrossRef]
- Yu, J.; Zhang, X.; Tan, T. A novel immobilization method of Saccharomyces cerevisiae to sorghum bagasse for ethanol production. J. Biotechnol. 2007, 129, 415–420. [Google Scholar] [CrossRef] [PubMed]
- Stolarzewicz, I.; Bialecka-Florjanczyk, E.; Majewska, E.; Krzyczkowska, J. Immobilization of yeast on polymeric supports. Chem. Biochem. Eng. Q. 2011, 25, 135–144. [Google Scholar]
- Pereira, F.B.; Guimarães, P.M.R.; Gomes, D.G.; Mira, N.P.; Teixeira, M.C.; Sá-Correia, I.; Domingues, L. Identification of candidate genes for yeast engineering to improve bioethanol production in very high gravity and lignocellulosic biomass industrial fermentations. Biotechnol. Biofuels 2011, 4, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, K.-Q.; Hou, X.-Y.; Hao, A.-L.; Wang, P.-F.; Fu, X.-M.; Lv, A.; Dong, J. Truncation of CYR1 promoter in industrial ethanol yeasts for improved ethanol yield in high temperature condition. Process Biochem. 2018, 65, 37–45. [Google Scholar] [CrossRef]
- Tao, X.; Zheng, D.; Liu, T.; Wang, P.; Zhao, W.; Zhu, M.; Jiang, X.; Zhao, Y.; Wu, X. A novel strategy to construct yeast Saccharomyces cerevisiae strains for very high gravity fermentation. PLoS ONE 2012, 7, e31235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.J.; Ding, W.T.; Zhang, G.C.; Wang, J.Y. Improving ethanol fermentation performance of Saccharomyces cerevisiae in very high-gravity fermentation through chemical mutagenesis and meiotic recombination. Appl. Microbiol. Biotechnol. 2011, 91, 1239–1246. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cao, S.; Wang, W.T.; Wang, K.T.; Jia, X. Very high gravity ethanol and fatty acid production of Zymomonas mobilis without amino acid and vitamin. J. Ind. Microbiol. Biotechnol. 2016, 43, 861–871. [Google Scholar] [CrossRef]
- Guo, Z.P.; Qiu, C.Y.; Zhang, L.; Ding, Z.Y.; Wang, Z.X.; Shi, G.Y. Expression of aspartic protease from Neurospora crassa in industrial ethanol-producing yeast and its application in ethanol production. Enzyme Microb. Technol. 2011, 48, 148–154. [Google Scholar] [CrossRef]
- Sun, Y.; Cheng, J. Hydrolysis of lignocellulosic materials for ethanol production: A review. Bioresour. Technol. 2002, 83, 1–11. [Google Scholar] [CrossRef]
- Gomes, D.G.; Rodrigues, A.C.; Domingues, L.; Gama, F.M. Cellulase recycling in biorefineries—Is it possible? Appl. Microbiol. Biotechnol. 2015, 99, 4131–4143. [Google Scholar] [CrossRef] [Green Version]
- Gomes, D.G.; Domingues, L.; Gama, F.M. Valorizing recycled paper sludge by a bioethanol production process with cellulase recycling. Bioresour. Technol. 2016, 216, 637–644. [Google Scholar] [CrossRef] [Green Version]
- Koppram, R.; Tomás-Pejó, E.; Xiros, C.; Olsson, L. Lignocellulosic ethanol production at high-gravity: Challenges and perspectives. Trends Biotechnol. 2014, 32, 46–53. [Google Scholar] [CrossRef]
- Almeida, J.R.M.; Bertilsson, M.; Gorwa-Grauslund, M.F.; Gorsich, S.; Lidén, G. Metabolic effects of furaldehydes and impacts on biotechnological process. Appl. Microbiol. Biotechnol. 2009, 82, 625–638. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.-Q.; Cai, L.-Y.; Ma, Y.-L. Study on inhibitors from acid pretreatment of corn stalk on ethanol fermentation by alcohol yeast. RSC Adv. 2020, 10, 38409–38415. [Google Scholar] [CrossRef]
- Gu, H.; Zhu, Y.; Peng, Y.; Liang, X.; Liu, X.; Shao, L.; Xu, Y.; Xu, Z.; Liu, R.; Li, J. Physiological mechanism of improved tolerance of Saccharomyces cerevisiae to lignin-derived phenolic acids in lignocellulosic ethanol fermentation by short-term adaptation. Biotechnol. Biofuels 2019, 12, 268. [Google Scholar] [CrossRef] [Green Version]
- Gomes, D.G.; Gama, F.M.; Domingues, L. Determinants on an efficient cellulase recycling process for the production of bioethanol from recycled paper sludge under high solid loadings. Biotechnol. Biofuels 2018, 11, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alkasrawi, M.; Rudolf, A.; Lidén, G.; Zacchi, G. Influence of strain and cultivation procedure on the performance of simultaneous saccharification and fermentation of steam pretreated spruce. Enzyme Microb. Technol. 2006, 38, 279–286. [Google Scholar] [CrossRef]
- Hawkins, G.M.; Doran-Peterson, J. A strain of Saccharomyces cerevisiae evolved for fermentation of lignocellulosic biomass displays improved growth and fermentative ability in high solids concentrations and in the presence of inhibitory compounds. Biotechnol. Biofuels 2011, 4, 49. [Google Scholar] [CrossRef] [Green Version]
- Rudolf, A.; Alkasrawi, M.; Zacchi, G.; Lidén, G. A comparison between batch and fed-batch simultaneous saccharification and fermentation of steam pretreated spruce. Enzyme Microb. Technol. 2005, 37, 195–204. [Google Scholar] [CrossRef]
- Rosgaard, L.; Andric, P.; Dam-Johansen, K.; Pedersen, S.; Meyer, A.S. Effects of substrate loading on enzymatic hydrolysis and viscosity of pretreated barley straw. Appl. Biochem. Biotechnol. 2007, 143, 27–40. [Google Scholar] [CrossRef]
- Sotaniemi, V.-H.; Taskila, S.; Ojamo, H.; Tanskanen, J. Controlled feeding of lignocellulosic substrate enhances the performance of fed-batch enzymatic hydrolysis in a stirred tank reactor. Biomass Bioenergy 2016, 91, 271–277. [Google Scholar] [CrossRef]
Figure 1.
Main stress factors affecting yeast cells in industrial fermentations for ethanol production and response mechanisms commonly involved (reproduced from Lopes et al. [11]).
Figure 1.
Main stress factors affecting yeast cells in industrial fermentations for ethanol production and response mechanisms commonly involved (reproduced from Lopes et al. [11]).

Table 2.
Main challenges to efficient VHG ethanol production and current research efforts addressing them.
Table 2.
Main challenges to efficient VHG ethanol production and current research efforts addressing them.
| Process Element | Challenge | Research Efforts |
|---|---|---|
| Fermentation organism | Stress induced by high ethanol concentrations | New high ethanol-tolerance organisms |
| Optimization of process conditions (e.g., temperature) | ||
| Alternative process configurations (e.g., cells immobilization) | ||
| Optimization of cells pitching strategy | ||
| Stress induced by high sugar concentrations | ||
| New high osmolites-tolerance organisms | ||
Alternative process configurations
| ||
| Nutrients starvation (especially nitrogen) | New low-cost nutrient sources | |
| Optimization of media composition | ||
| Cost of cells | Cells immobilization | |
| Process productivity | ||
| Sugar feedstock | Competition with food crops | New feedstocks not competing with food crops and/or with an inferior cost |
| Feedstock price | ||
| Sugar Yields | Plant development for high sugar contents | |
| Highly viscous materials | Process adjuvants (e.g., cell wall degrading enzymes) | |
| Optimization of mixing/improved mixing alternatives | ||
| Process | Mass transferlimitations | |
| Oxygen limitation | Optimization of oxygen supply/new strategies for oxygen supply | |
| Contamination risk | More robust organisms towards microbial contamination | |
| More thermotolerant organisms |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gomes, D.; Cruz, M.; de Resende, M.; Ribeiro, E.; Teixeira, J.; Domingues, L. Very High Gravity Bioethanol Revisited: Main Challenges and Advances. Fermentation 2021, 7, 38. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7010038
AMA Style
Gomes D, Cruz M, de Resende M, Ribeiro E, Teixeira J, Domingues L. Very High Gravity Bioethanol Revisited: Main Challenges and Advances. Fermentation. 2021; 7(1):38. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7010038
Chicago/Turabian StyleGomes, Daniel, Mariana Cruz, Miriam de Resende, Eloízio Ribeiro, José Teixeira, and Lucília Domingues. 2021. "Very High Gravity Bioethanol Revisited: Main Challenges and Advances" Fermentation 7, no. 1: 38. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7010038
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.