
Evaluation of Autochthonous Non-Saccharomyces Yeasts by Sequential Fermentation for Wine Differentiation in Galicia (NW Spain)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Yeast Strains and Culture Media
2.2. Sequential Fermentations
2.3. Microbiological Control
2.4. Chemical Analysis
2.5. Statistical Analysis
3. Results and Discussion
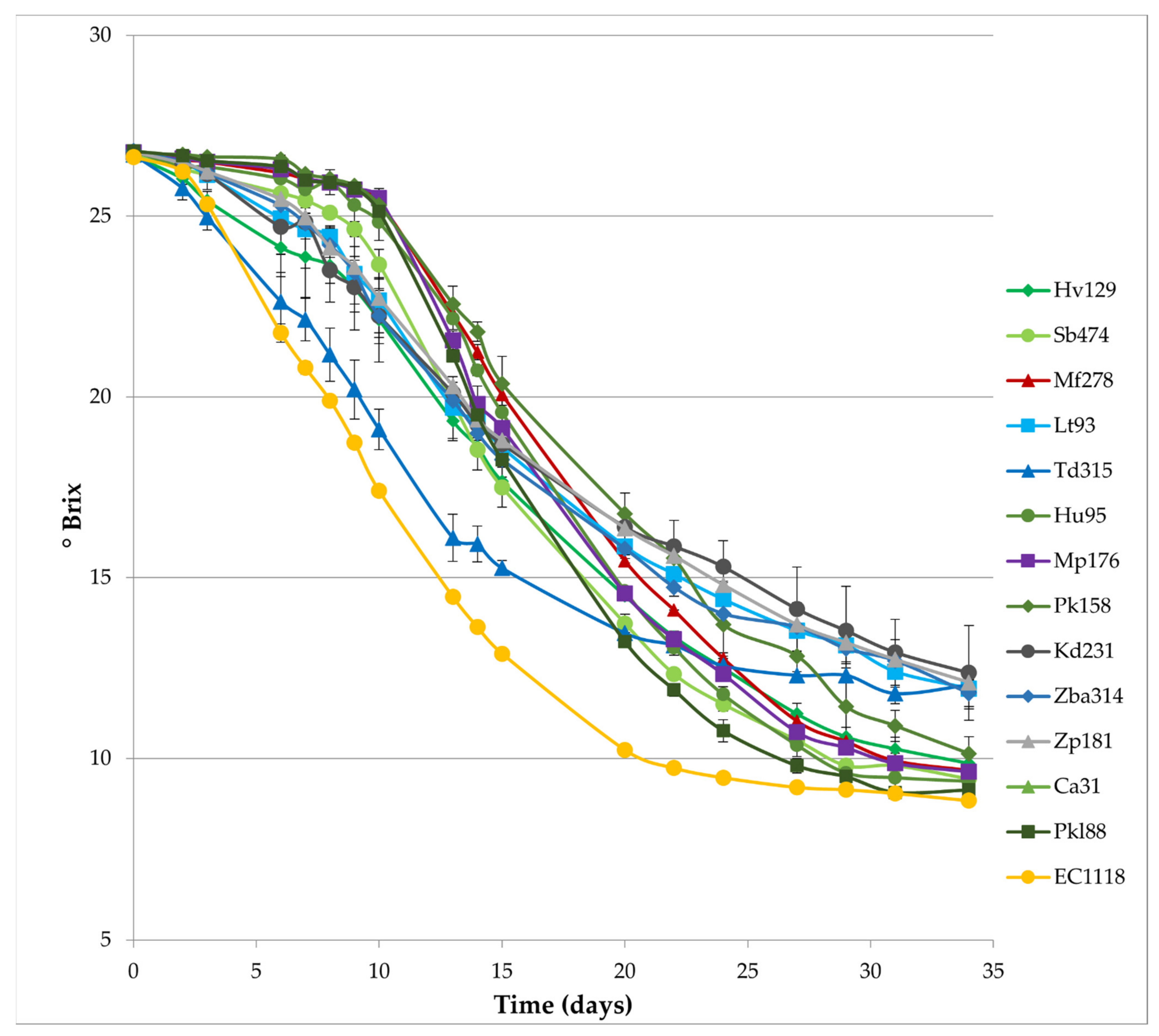
3.1. Kinetics of Sequential Fermentations and Yeast Population Dynamics
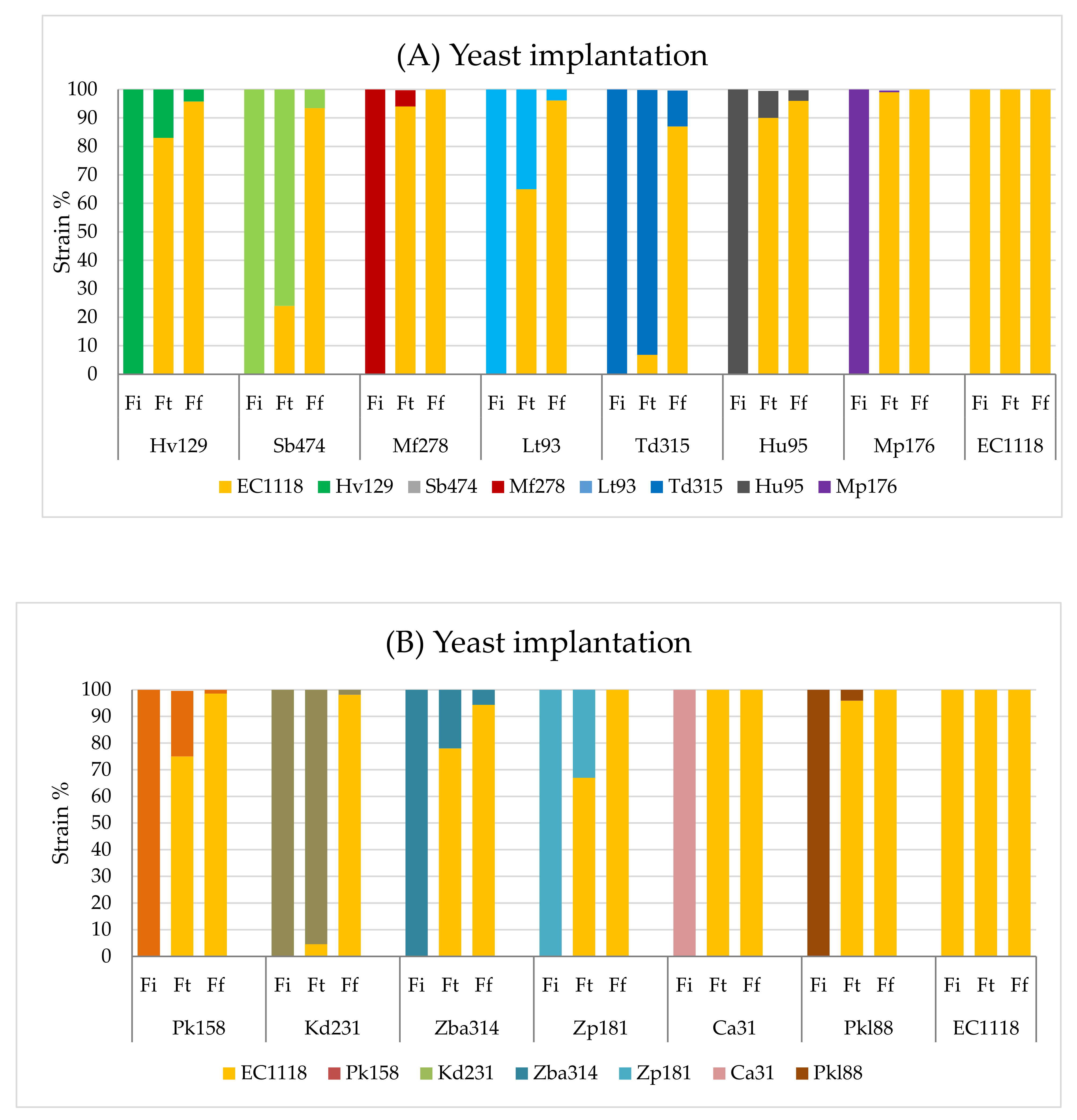
3.2. Yeast Implantation in Sequential Fermentations
3.3. Chemical Characteristics of Wine
3.4. Wine Aroma Composition
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Fleet, G.H.; Heard, G.M. Yeasts—Growth during fermentation. In Wine Microbiology and Biotechnology; Fleet, G.H., Ed.; Harwood Academic Publishers: Reading, UK, 1993; pp. 28–47. [Google Scholar]
- Fleet, G.H. Wine yeasts for the future. FEMS Yeast Res. 2008, 8, 979–995. [Google Scholar] [CrossRef] [Green Version]
- Jolly, N.P.; Augustyn, O.P.H.; Pretorius, I.S. The Role and Use of Non-Saccharomyces Yeasts in Wine Production. S. Afr. J. Enol. Vitic. 2006, 27, 15–39. [Google Scholar] [CrossRef]
- Jolly, N.P.; Varela, C.; Pretorius, I.S. Not your ordinary yeast: Non-Saccharomyces yeasts in wine production uncovered. FEMS Yeast Res. 2014, 14, 215–237. [Google Scholar] [CrossRef] [Green Version]
- Fleet, G.H. Yeast interactions and wine flavour. Int. J. Food Microbiol. 2003, 86, 11–22. [Google Scholar] [CrossRef]
- Ciani, M.; Comitini, F.; Mannazzu, I.; Domizio, P. Controlled mixed culture fermentation: A new perspective on the use of non-Saccharomyces yeasts in winemaking. FEMS Yeast Res. 2010, 10, 123–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padilla, B.; Gil, J.V.; Manzanares, P. Past and future of non-Saccharomyces yeasts: From spoilage microorganisms to biotechnological tools for improving wine aroma complexity. Front. Microbiol. 2016, 7, 1–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drumonde-Neves, J.; Fernandes, T.; Lima, T.; Pais, C.; Franco-Duarte, R. Learning from 80 years of studies: A comprehensive catalogue of non-Saccharomyces yeasts associated with viticulture and winemaking. FEMS Yeast Res. 2021, 21, foab017. [Google Scholar] [CrossRef] [PubMed]
- Benito, S. The impacts of Lachancea thermotolerans yeast strains on winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 6775–6790. [Google Scholar] [CrossRef] [Green Version]
- Benito, S. The impact of Torulaspora delbrueckii yeast in winemaking. Appl. Microbiol. Biotechnol. 2018, 102, 3081–3094. [Google Scholar] [CrossRef] [PubMed]
- Vicente, J.; Ruiz, J.; Belda, I.; Benito-Vázquez, I.; Marquina, D.; Calderón, F.; Santos, A.; Benito, S. The Genus Metschnikowia in Enology. Microorganisms 2020, 8, 1038. [Google Scholar] [CrossRef]
- Roudil, L.; Russo, P.; Berbegal, C.; Albertin, W.; Spano, G.; Capozzi, V. Non-Saccharomyces Commercial Starter Cultures: Scientific Trends, Recent Patents and Innovation in the Wine Sector. Recent Pat. Food Nutr. Agric. 2020, 11, 27–39. [Google Scholar] [CrossRef]
- Ciani, M.; Morales, P.; Comitini, F.; Tronchoni, J.; Canonico, L.; Curiel, J.A.; Oro, L.; Rodrigues, A.J.; Gonzalez, R. Non-conventional Yeast Species for Lowering Ethanol Content of Wines. Front. Microbiol. 2016, 7, 642. [Google Scholar] [CrossRef] [Green Version]
- Rolle, L.; Englezos, V.; Torchio, F.; Cravero, F.; Río Segade, S.; Rantsiou, K.; Giacosa, S.; Gambuti, A.; Gerbi, V.; Cocolin, L. Alcohol reduction in red wines by technological and microbiological approaches: A comparative study. Aust. J. Grape Wine Res. 2018, 24, 62–74. [Google Scholar] [CrossRef]
- Varela, C.; Dry, P.R.; Kutyna, D.R.; Francis, I.L.; Henschke, P.A.; Curtin, C.D.; Chambers, P.J. Strategies for reducing alcohol concentration in wine. Aust. J. Grape Wine Res. 2015, 21, 670–679. [Google Scholar] [CrossRef]
- Canonico, L.; Comitini, F.; Oro, L.; Ciani, M. Sequential fermentation with selected immobilized non-Saccharomyces yeast for reduction of ethanol content in wine. Front. Microbiol. 2016, 7, 278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Englezos, V.; Giacosa, S.; Rantsiou, K.; Rolle, L.; Cocolin, L. Starmerella bacillaris in winemaking: Opportunities and risks. Curr. Opin. Food Sci. 2017, 17, 30–35. [Google Scholar] [CrossRef]
- Varela, C.; Sengler, F.; Solomon, M.; Curtin, C. Volatile flavour profile of reduced alcohol wines fermented with the non-conventional yeast species Metschnikowia pulcherrima and Saccharomyces uvarum. Food Chem. 2016, 209, 57–64. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, J.; Belda, I.; Beisert, B.; Navascués, E.; Marquina, D.; Calderón, F.; Rauhut, D.; Santos, A.; Benito, S. Analytical impact of Metschnikowia pulcherrima in the volatile profile of Verdejo white wines. Appl. Microbiol. Biotechnol. 2018, 102, 8501–8509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pallmann, C.L.; Brown, J.A.; Olineka, T.L.; Cocolin, L.; Mills, D.A.; Bisson, L.F. Use of WL Medium to Profile Native Flora Fermentations. Am. J. Enol. Vitic. 2001, 52, 198–203. [Google Scholar]
- Esteve-Zarzoso, B.; Belloch, C.; Uruburu, F.; Querol, A. Identification of yeasts by RFLP analysis of the 5.8S rRNA gene and the two ribosomal internal transcribed spacers. Int. J. Syst. Bacteriol. 1999, 49, 329–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Querol, A.; Barrio, E.; Huerta, T.; Ramon, D. Molecular monitoring of wine fermentations conducted by active dry yeast strains. Appl. Environ. Microbiol. 1992, 58, 2948–2953. [Google Scholar] [CrossRef] [Green Version]
- OIV. Office International de la Vigne et du Vin Compendium of International Methods of Wine and Must Analysis. Vol. 1 and 2. Paris, France, 2018. Available online: http://www.oiv.int (accessed on 25 August 2021).
- Ortega, C.; López, R.; Cacho, J.; Ferreira, V. Fast analysis of important wine volatile compounds: Development and validation of a new method based on gas chromatographic–flame ionisation detection analysis of dichloromethane microextracts. J. Chromatogr. A 2001, 923, 205–214. [Google Scholar] [CrossRef]
- Contreras, A.; Hidalgo, C.; Henschke, P.; Chambers, P.; Curtin, C.; Varela, C. Evaluation of Non-Saccharomyces Yeasts for the Reduction of Alcohol Content in Wine. Appl. Environ. Microbiol. 2014, 80, 1670–1678. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García, M.; Esteve-Zarzoso, B.; Cabellos, J.M.; Arroyo, T. Sequential Non-Saccharomyces and Saccharomyces cerevisiae Fermentations to Reduce the Alcohol Content in Wine. Fermentation 2020, 6, 60. [Google Scholar] [CrossRef]
- Viana, F.; Gil, J.V.; Genovés, S.; Vallés, S.; Manzanares, P. Rational selection of non-Saccharomyces wine yeasts for mixed starters based on ester formation and enological traits. Food Microbiol. 2008, 25, 778–785. [Google Scholar] [CrossRef]
- Mestre Furlani, M.V.; Maturano, Y.P.; Combina, M.; Mercado, L.A.; Toro, M.E.; Vazquez, F. Selection of non-Saccharomyces yeasts to be used in grape musts with high alcoholic potential: A strategy to obtain wines with reduced ethanol content. FEMS Yeast Res. 2017, 17, fox010. [Google Scholar] [CrossRef] [Green Version]
- Gobbi, M.; De Vero, L.; Solieri, L.; Comitini, F.; Oro, L.; Giudici, P.; Ciani, M. Fermentative aptitude of non-Saccharomyces wine yeast for reduction in the ethanol content in wine. Eur. Food Res. Technol. 2014, 239, 41–48. [Google Scholar] [CrossRef]
- Contreras, A.; Hidalgo, C.; Schmidt, S.; Henschke, P.A.; Curtin, C.; Varela, C. The application of non-Saccharomyces yeast in fermentations with limited aeration as a strategy for the production of wine with reduced alcohol content. Int. J. Food Microbiol. 2015, 205, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Binati, R.L.; Lemos Junior, W.J.F.; Luzzini, G.; Slaghenaufi, D.; Ugliano, M.; Torriani, S. Contribution of non-Saccharomyces yeasts to wine volatile and sensory diversity: A study on Lachancea thermotolerans, Metschnikowia spp. and Starmerella bacillaris strains isolated in Italy. Int. J. Food Microbiol. 2020, 318, 108470. [Google Scholar] [CrossRef] [PubMed]
- Comitini, F.; Gobbi, M.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 2011, 28, 873–882. [Google Scholar] [CrossRef] [PubMed]
- Shekhawat, K.; Bauer, F.F.; Setati, M.E. Impact of oxygenation on the performance of three non-Saccharomyces yeasts in co-fermentation with Saccharomyces cerevisiae. Appl. Microbiol. Biotechnol. 2017, 101, 2479–2491. [Google Scholar] [CrossRef]
- Englezos, V.; Cravero, F.; Torchio, F.; Rantsiou, K.; Ortiz-Julien, A.; Lambri, M.; Gerbi, V.; Rolle, L.; Cocolin, L. Oxygen availability and strain combination modulate yeast growth dynamics in mixed culture fermentations of grape must with Starmerella bacillaris and Saccharomyces cerevisiae. Food Microbiol. 2018, 69, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Albergaria, H.; Arneborg, N. Dominance of Saccharomyces cerevisiae in alcoholic fermentation processes: Role of physiological fitness and microbial interactions. Appl. Microbiol. Biotechnol. 2016, 100, 2035–2046. [Google Scholar] [CrossRef]
- Englezos, V.; Rantsiou, K.; Giacosa, S.; Río Segade, S.; Rolle, L.; Cocolin, L. Cell-to-cell contact mechanism modulates Starmerella bacillaris death in mixed culture fermentations with Saccharomyces cerevisiae. Int. J. Food Microbiol. 2019, 289, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Gobbi, M.; Comitini, F.; Domizio, P.; Romani, C.; Lencioni, L.; Mannazzu, I.; Ciani, M. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: A strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 2013, 33, 271–281. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Torija, M.-J.; Mas, A.; Beltran, G.; Navarro, Y. Effect of a Multistarter Yeast Inoculum on Ethanol Reduction and Population Dynamics in Wine Fermentation. Foods 2021, 10, 623. [Google Scholar] [CrossRef] [PubMed]
- Puertas, B.; Jiménez, M.J.; Cantos-Villar, E.; Cantoral, J.M.; Rodríguez, M.E. Use of Torulaspora delbrueckii and Saccharomyces cerevisiae in semi-industrial sequential inoculation to improve quality of Palomino and Chardonnay wines in warm climates. J. Appl. Microbiol. 2016, 122, 733–746. [Google Scholar] [CrossRef] [PubMed]
- Puertas, B.; Jimenez-Hierro, M.J.; Cantos-Villar, E.; Marrufo-Curtido, A.; Carbú, M.; Cuevas, F.J.; Moreno-Rojas, J.M.; González-Rodríguez, V.E.; Cantoral, J.M.; Ruiz-Moreno, M.J. The influence of yeast on chemical composition and sensory properties of dry white wines. Food Chem. 2018, 253, 227–235. [Google Scholar] [CrossRef]
- Velázquez, R.; Zamora, E.; Álvarez, M.L.; Hernández, L.M.; Ramírez, M. Effects of new Torulaspora delbrueckii killer yeasts on the must fermentation kinetics and aroma compounds of white table wine. Front. Microbiol. 2015, 6, 1222. [Google Scholar] [CrossRef] [Green Version]
- Ramírez, M.; Velázquez, R.; Maqueda, M.; Zamora, E.; López-Piñeiro, A.; Hernández, L.M. Influence of the dominance of must fermentation by Torulaspora delbrueckii on the malolactic fermentation and organoleptic quality of red table wine. Int. J. Food Microbiol. 2016, 238, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Agarbati, A.; Canonico, L.; Comitini, F.; Ciani, M. Improved Saccharomyces cerevisiae Strain in Pure and Sequential Fermentation with Torulaspora delbrueckii for the Production of Verdicchio Wine with Reduced Sulfites. Appl. Sci. 2020, 10, 6722. [Google Scholar] [CrossRef]
- Dutraive, O.; Benito, S.; Fritsch, S.; Beisert, B.; Patz, C.-D.; Rauhut, D. Effect of Sequential Inoculation with Non-Saccharomyces and Saccharomyces Yeasts on Riesling Wine Chemical Composition. Fermentation 2019, 5, 79. [Google Scholar] [CrossRef] [Green Version]
- Englezos, V.; Rantsiou, K.; Cravero, F.; Torchio, F.; Ortiz-Julien, A.; Gerbi, V.; Rolle, L.; Cocolin, L. Starmerella bacillaris and Saccharomyces cerevisiae mixed fermentations to reduce ethanol content in wine. Appl. Microbiol. Biotechnol. 2016, 100, 5515–5526. [Google Scholar] [CrossRef] [PubMed]
- Nisiotou, A.; Sgouros, G.; Mallouchos, A.; Nisiotis, C.S.; Michaelidis, C.; Tassou, C.; Banilas, G. The use of indigenous Saccharomyces cerevisiae and Starmerella bacillaris strains as a tool to create chemical complexity in local wines. Food Res. Int. 2018, 111, 498–508. [Google Scholar] [CrossRef] [PubMed]
- Blanco, P.; Rabuñal, E.; Neira, N.; Castrillo, D. Dynamic of Lachancea thermotolerans Population in Monoculture and Mixed Fermentations: Impact on Wine Characteristics. Beverages 2020, 6, 36. [Google Scholar] [CrossRef]
- Fairbairn, S.; Engelbrecht, L.; Setati, M.E.; du Toit, M.; Bauer, F.F.; Divol, B.; Rossouw, D. Combinatorial analysis of population dynamics, metabolite levels and malolactic fermentation in Saccharomyces cerevisiae/Lachancea thermotolerans mixed fermentations. Food Microbiol. 2021, 96, 103712. [Google Scholar] [CrossRef] [PubMed]
- Roca-Mesa, H.; Sendra, S.; Mas, A.; Beltran, G.; Torija, M.-J. Nitrogen Preferences during Alcoholic Fermentation of Different Non-Saccharomyces Yeasts of Oenological Interest. Microorganisms 2020, 8, 157. [Google Scholar] [CrossRef] [Green Version]
- Luan, Y.; Zhang, B.-Q.; Duan, C.-Q.; Yan, G.-L. Effects of different pre-fermentation cold maceration time on aroma compounds of Saccharomyces cerevisiae co-fermentation with Hanseniaspora opuntiae or Pichia kudriavzevii. LWT 2018, 92, 177–186. [Google Scholar] [CrossRef]
- Zhu, X.; Navarro, Y.; Mas, A.; Torija, M.-J.; Beltran, G. A Rapid Method for Selecting Non-Saccharomyces Strains with a Low Ethanol Yield. Microorganisms 2020, 8, 658. [Google Scholar] [CrossRef] [PubMed]
- Lleixà, J.; Martín, V.; del Portillo, M.C.; Carrau, F.; Beltran, G.; Mas, A. Comparison of fermentation and wines produced by inoculation of Hanseniaspora vineae and Saccharomyces cerevisiae. Front. Microbiol. 2016, 7, 338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina, K.; Boido, E.; Fariña, L.; Gioia, O.; Gomez, M.E.; Barquet, M.; Gaggero, C.; Dellacassa, E.; Carrau, F. Increased flavour diversity of Chardonnay wines by spontaneous fermentation and co-fermentation with Hanseniaspora vineae. Food Chem. 2013, 141, 2513–2521. [Google Scholar] [CrossRef]
- Mestre, M.V.; Maturano, Y.P.; Gallardo, C.; Combina, M.; Mercado, L.; Toro, M.E.; Carrau, F.; Vazquez, F.; Dellacassa, E. Impact on Sensory and Aromatic Profile of Low Ethanol Malbec Wines Fermented by Sequential Culture of Hanseniaspora uvarum and Saccharomyces cerevisiae Native Yeasts. Fermentation 2019, 5, 65. [Google Scholar] [CrossRef] [Green Version]
- Fanzone, M.L.; Sari, S.E.; Mestre, M.V.; Catania, A.A.; Catelén, M.J.; Jofré, V.P.; González-Miret, M.L.; Combina, M.; Vazquez, F.; Maturano, Y.P. Combination of pre-fermentative and fermentative strategies to produce Malbec wines of lower alcohol and pH, with high chemical and sensory quality. OENO One 2020, 54. [Google Scholar] [CrossRef]
- Harlé, O.; Legrand, J.; Tesnière, C.; Pradal, M.; Mouret, J.-R.; Nidelet, T. Investigations of the mechanisms of interactions between four non-conventional species with Saccharomyces cerevisiae in oenological conditions. bioRxiv 2019, 363531. [Google Scholar] [CrossRef] [PubMed]
- Benito, S.; Hofmann, T.; Laier, M.; Lochbühler, B.; Schüttler, A.; Ebert, K.; Fritsch, S.; Röcker, J.; Rauhut, D. Effect on quality and composition of Riesling wines fermented by sequential inoculation with non-Saccharomyces and Saccharomyces cerevisiae. Eur. Food Res. Technol. 2015, 241, 707–717. [Google Scholar] [CrossRef]
- Anfang, N.; Brajkovich, M.; Goddard, M.R. Co-fermentation with Pichia kluyveri increases varietal thiol concentrations in Sauvignon Blanc. Aust. J. Grape Wine Res. 2009, 15, 1–8. [Google Scholar] [CrossRef]
- Vicente, J.; Calderón, F.; Santos, A.; Marquina, D.; Benito, S. High Potential of Pichia kluyveri and Other Pichia Species in Wine Technology. Int. J. Mol. Sci. 2021, 22, 1196. [Google Scholar] [CrossRef] [PubMed]
- Tofalo, R.; Chaves-López, C.; Di Fabio, F.; Schirone, M.; Felis, G.E.; Torriani, S.; Paparella, A.; Suzzi, G. Molecular identification and osmotolerant profile of wine yeasts that ferment a high sugar grape must. Int. J. Food Microbiol. 2009, 130, 179–187. [Google Scholar] [CrossRef]
- Castrillo, D.; Rabuñal, E.; Neira, N.; Blanco, P. Oenological potential of non-Saccharomyces yeasts to mitigate effects of climate change in winemaking: Impact on aroma and sensory profiles of Treixadura wines. FEMS Yeast Res. 2019, 19, 1–11. [Google Scholar] [CrossRef]
- Varela, C.; Barker, A.; Tran, T.; Borneman, A.; Curtin, C. Sensory profile and volatile aroma composition of reduced alcohol Merlot wines fermented with Metschnikowia pulcherrima and Saccharomyces uvarum. Int. J. Food Microbiol. 2017, 252, 1–9. [Google Scholar] [CrossRef]
- Morales, P.; Rojas, V.; Quirós, M.; Gonzalez, R. The impact of oxygen on the final alcohol content of wine fermented by a mixed starter culture. Appl. Microbiol. Biotechnol. 2015, 99, 3993–4003. [Google Scholar] [CrossRef] [Green Version]
- Escribano, R.; González-Arenzana, L.; Portu, J.; Garijo, P.; López-Alfaro, I.; López, R.; Santamaría, P.; Gutiérrez, A.R. Wine aromatic compound production and fermentative behaviour within different non-Saccharomyces species and clones. J. Appl. Microbiol. 2018, 124, 1521–1531. [Google Scholar] [CrossRef] [PubMed]
- García, M.; Arroyo, T.; Crespo García, J.; Cabellos, J.M.; Esteve-Zarzoso, B. Use of native non-Saccharomyces strain: A new strategy in D.O. “Vinos de Madrid” (Spain) wines elaboration. Eur. J. Food Sci. Technol. 2017, 5, 1–26. [Google Scholar]
- Ivit, N.N.; Longo, R.; Kemp, B. The Effect of Non-Saccharomyces and Saccharomyces Non-cerevisiae Yeasts on Ethanol and Glycerol Levels in Wine. Fermentation 2020, 6, 77. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Pretorius, I.S. Yeast modulation of wine flavor. Adv. Appl. Microbiol. 2005, 57, 131–175. [Google Scholar] [CrossRef]
- Englezos, V.; Rantsiou, K.; Cravero, F.; Torchio, F.; Pollon, M.; Fracassetti, D.; Ortiz-Julien, A.; Gerbi, V.; Rolle, L.; Cocolin, L. Volatile profile of white wines fermented with sequential inoculation of Starmerella bacillaris and Saccharomyces cerevisiae. Food Chem. 2018, 257, 350–360. [Google Scholar] [CrossRef]
- Belda, I.; Ruiz, J.; Beisert, B.; Navascués, E.; Marquina, D.; Calderón, F.; Rauhut, D.; Benito, S.; Santos, A. Influence of Torulaspora delbrueckii in varietal thiol (3-SH and 4-MSP) release in wine sequential fermentations. Int. J. Food Microbiol. 2017, 257, 183–191. [Google Scholar] [CrossRef]
- Lambrechts, M.G.; Pretorius, I.S. Yeast and its Importance to Wine Aroma—A Review. S. Afr. J. Enol. Vitic. 2000, 21, 97–129. [Google Scholar] [CrossRef] [Green Version]
- Varela, J.; Varela, C. Microbiological strategies to produce beer and wine with reduced ethanol concentration. Curr. Opin. Biotechnol. 2019, 56, 88–96. [Google Scholar] [CrossRef]
- Escribano, R.; González-Arenzana, L.; Garijo, P.; Berlanas, C.; López-Alfaro, I.; López, R.; Gutiérrez, A.R.; Santamaría, P. Screening of enzymatic activities within different enological non-Saccharomyces yeasts. J. Food Sci. Technol. 2017, 54, 1555–1564. [Google Scholar] [CrossRef] [Green Version]
- Canonico, L.; Solomon, M.; Comitini, F.; Ciani, M.; Varela, C. Volatile profile of reduced alcohol wines fermented with selected non-Saccharomyces yeasts under different aeration conditions. Food Microbiol. 2019, 84, 103247. [Google Scholar] [CrossRef] [PubMed]
- Garavaglia, J.; de Souza Schneider, R.D.C.; Camargo Mendes, S.D.; Welke, J.E.; Zini, C.A.; Caramão, E.B.; Valente, P. Evaluation of Zygosaccharomyces bailii BCV 08 as a co-starter in wine fermentation for the improvement of ethyl esters production. Microbiol. Res. 2015, 173, 59–65. [Google Scholar] [CrossRef]
- Saerens, S.M.G.; Delvaux, F.R.; Verstrepen, K.J.; Van Dijck, P.; Thevelein, J.M.; Delvaux, F.R. Parameters affecting ethyl ester production by Saccharomyces cerevisiae during fermentation. Appl. Environ. Microbiol. 2008, 74, 454–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shinohara, T. Gas Chromatographic Analysis of Volatile Fatty Acids in Wines. Agric. Biol. Chem. 1985, 49, 2211–2212. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Yeast Species | Strain | Code |
|---|---|---|
| Hanseniaspora vineae | 129 | Hv129 |
| Hanseniaspora uvarum | 95 | Hu95 |
| Starmerella bacillaris | 474 | Sb474 |
| Metschnikowia fructicola | 278 | Mf278 |
| Lachancea thermotolerans | 93 | Lt93 |
| Torulaspora delbrueckii | 315 | Td315 |
| Metschnikowiapulcherrima | 176 | Mp176 |
| Pichia kudriavzevii | 158 | Pk158 |
| Pichia kluyveri | 88 | Pkl88 |
| Kluyveromyces dobzhanskii | 231 | Kd231 |
| Zygosaccharomycesbailii | 314 | Zba314 |
| Zygosaccharomycesparabailii | 181 | Zp181 |
| Candidaapicola | 31 | Ca31 |
| Saccharomyces cerevisiae | EC1118 | EC1118 |
| Fermentation | Fi | Ft * | Ff |
|---|---|---|---|
| Hv129 + Sc | 7.50 | 8.19 a | 6.44 |
| Sb474 + Sc | 8.26 | 9.06 b | 7.74 |
| Mf278 + Sc | 8.02 | 8.71 ab | 6.96 |
| Lt93 + Sc | 7.91 | 7.61 a | 6.68 |
| Td315 + Sc | 7.96 | 8.76 ab | 5.02 |
| Hu95 + Sc | 7.66 | 8.78 ab | 6.48 |
| Mp176 + Sc | 7.98 | 8.68 ab | 6.75 |
| Pk158 + Sc | 7.83 | 8.80 ab | 7.49 |
| Kd231 + Sc | 8.30 | 8.97 ab | 7.32 |
| Zba314 + Sc | 7.73 | 8.49 ab | 5.97 |
| Zp181 + Sc | 7.51 | 8.36 ab | 6.39 |
| Ca31 + Sc | 7.74 | 8.90 ab | 6.74 |
| Pkl88 + Sc | 7.60 | 8.57 ab | 7.73 |
| EC1118 (Control-Sc) | 7.86 | 8.38 ab | 6.60 |
| Fermentation | Ethanol (% v/v) | Glucose + Fructose (g/L) | Ethanol Yield (g/g) | Total Acidity (g Tartaric/L) | Volatile Acidity (g Acetic/L) | Glycerol (g/L) | Sensory Evaluation * |
|---|---|---|---|---|---|---|---|
| Hv129 + Sc | 16.9 ± 0.12 a | 6.8 ± 0.4 bc | 0.52 ± 0.01 ab | 6.4 ± 0.4 abc | 0.61 ± 0.13 abc | 6.7 ± 0.8 fg | 3 |
| Sb474 + Sc | 16.9 ± 0.03 a | 3.4 ± 0.2 c | 0.52 ± 0.01 ab | 7.5 ± 0.1 ab | 0.79 ± 0.01 ab | 11.1 ± 0.5 a | 1− |
| Mf278 + Sc | 16.8 ± 0.05 ab | 3.8 ± 0.4 bc | 0.52 ± 0.01 abc | 7.2 ± 0.0 abc | 0.66 ± 0.01 abc | 8.7 ± 0.2 bcd | 5+ |
| Lt93 + Sc | 15.8 ± 0.74 bc | 17.0 ± 11.4 bc | 0.49 ± 0.02 abc | 8.8 ± 0.3 abc | 0.62 ± 0.01 abc | 7.2 ± 0.3 cdefg | 5+ |
| Td315 + Sc | 14.2 ± 0.21 d | 42.2 ± 4.7 a | 0.44 ± 0.01 d | 6.9 ± 0.3 gh | 0.47 ± 0.09 c | 6.3 ± 0.5 g | 3 |
| Hu95 + Sc | 16.8 ± 0.04 ab | 5.5 ± 0.5 bc | 0.51 ± 0.01 abc | 7.8 ± 0.1 cde | 0.63 ± 0.07 abc | 7.1 ± 0.3 efg | 3 |
| Mp176 + Sc | 16.9 ± 0.06 a | 3.2 ± 0.8 c | 0.53 ± 0.00 a | 7.2 ± 0.1 fg | 0.58 ± 0.01 bc | 8.8 ± 0.2 bc | 5+ |
| Pk158 + Sc | 16.8 ± 0.12 ab | 4.5 ± 1.7 bc | 0.53 ± 0.00 ab | 8.2 ± 0.2 abc | 0.80 ± 0.14 ab | 7.2 ± 0.1 defg | 3 |
| Kd231 + Sc | 15.6 ± 1.05 c | 18.4 ± 14.6 bc | 0.48 ± 0.03 bc | 8.5 ± 0.4 ab | 0.46 ± 0.08 c | 9.7 ± 0.7 ab | 0− |
| Zba314 + Sc | 15.8 ± 0.41 bc | 15.2 ± 6.7 bc | 0.49 ± 0.01 abc | 8.4 ± 0.1 bcd | 0.59 ± 0.14 bc | 8.4 ± 1.1 bcde | 1− |
| Zp181 + Sc | 15.4 ± 0.30 c | 20.3 ± 4.4 b | 0.48 ± 0.01 cd | 8.2 ± 0.2 bcd | 0.68 ± 0.05 abc | 8.2 ± 0.8 bcdef | 2− |
| Ca31 + Sc | 16.9 ± 0.05 a | 2.9 ± 0.3 c | 0.52 ± 0.01 ab | 7.7 ± 0.1 cdef | 0.67 ± 0.03 abc | 7.8 ± 0.2 cdefg | 3 |
| Pkl88 + Sc | 16.8 ± 0.04 ab | 4.5 ± 0.2 bc | 0.52 ± 0.01 abc | 7.7 ± 0.1 cdef | 0.82 ± 0.03 a | 7.1 ± 0.4 efg | 5+ |
| EC1118 (Sc control) | 16.9 ± 0.04 ab | 2.2 ± 0.3 c | 0.53 ± 0.00 ab | 7.9 ± 0.0 bcd | 0.56 ± 0.02 c | 6.4 ± 0.2 g | 3 |
| Fermentation | Higher Alcohols | Esters | C3–C5 Fatty Acids | C6–C10 Fatty Acids |
|---|---|---|---|---|
| Hv129 + Sc | 195.824 | 0.442 | 3.309 | 1.640 |
| Sb474 + Sc | 185.974 | 0.347 | 4.465 | 2.654 |
| Mf278 + Sc | 260.457 | 0.942 | 2.015 | 5.411 |
| Lt93 + Sc | 246.885 | 0.415 | 2.810 | 1.565 |
| Td315 + Sc | 272.385 | 0.173 | 4.156 | 0.598 |
| Hu95 + Sc | 212.950 | 0.770 | 2.584 | 4.342 |
| Mp176 + Sc | 235.175 | 0.792 | 2.990 | 4.599 |
| Pk158 + Sc | 233.540 | 0.977 | 3.969 | 4.521 |
| Kd231 + Sc | 591.286 | 0.355 | 6.028 | 0.976 |
| Zba314 + Sc | 191.697 | 0.303 | 5.657 | 0.863 |
| Zp181 + Sc | 183.328 | 0.408 | 4.613 | 1.013 |
| Ca31 + Sc | 212.528 | 0.773 | 5.256 | 4.196 |
| Pkl88 + Sc | 209.564 | 1.746 | 3.869 | 5.312 |
| EC1118 (Sc control) | 229.637 | 0.806 | 3.453 | 3.794 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanco, P.; Castrillo, D.; Graña, M.J.; Lorenzo, M.J.; Soto, E. Evaluation of Autochthonous Non-Saccharomyces Yeasts by Sequential Fermentation for Wine Differentiation in Galicia (NW Spain). Fermentation 2021, 7, 183. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7030183
Blanco P, Castrillo D, Graña MJ, Lorenzo MJ, Soto E. Evaluation of Autochthonous Non-Saccharomyces Yeasts by Sequential Fermentation for Wine Differentiation in Galicia (NW Spain). Fermentation. 2021; 7(3):183. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7030183
Chicago/Turabian StyleBlanco, Pilar, David Castrillo, María José Graña, María José Lorenzo, and Elvira Soto. 2021. "Evaluation of Autochthonous Non-Saccharomyces Yeasts by Sequential Fermentation for Wine Differentiation in Galicia (NW Spain)" Fermentation 7, no. 3: 183. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7030183