
Insights on the Advancements of In Silico Metabolic Studies of Succinic Acid Producing Microorganisms: A Review with Emphasis on Actinobacillus succinogenes



Abstract
:1. Introduction
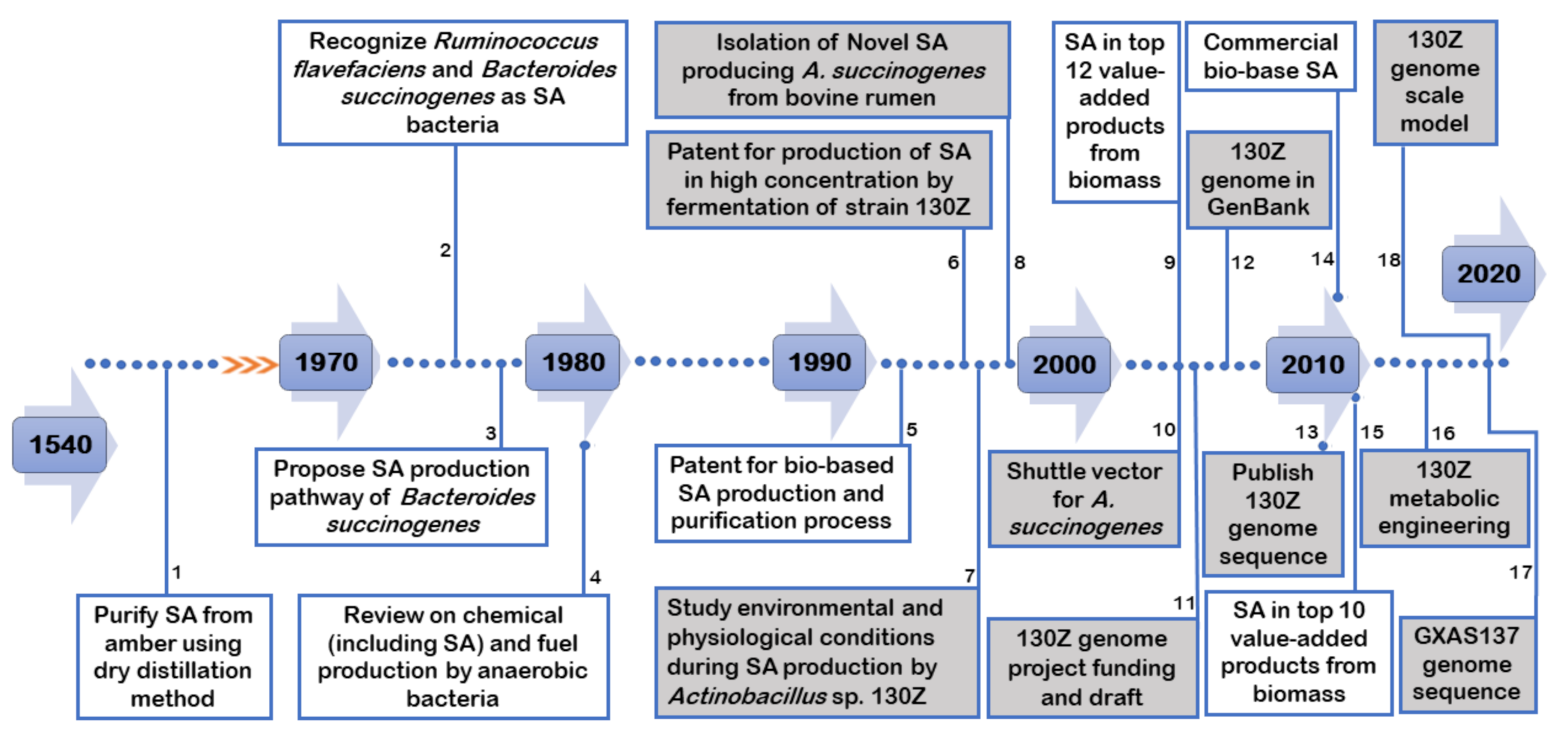
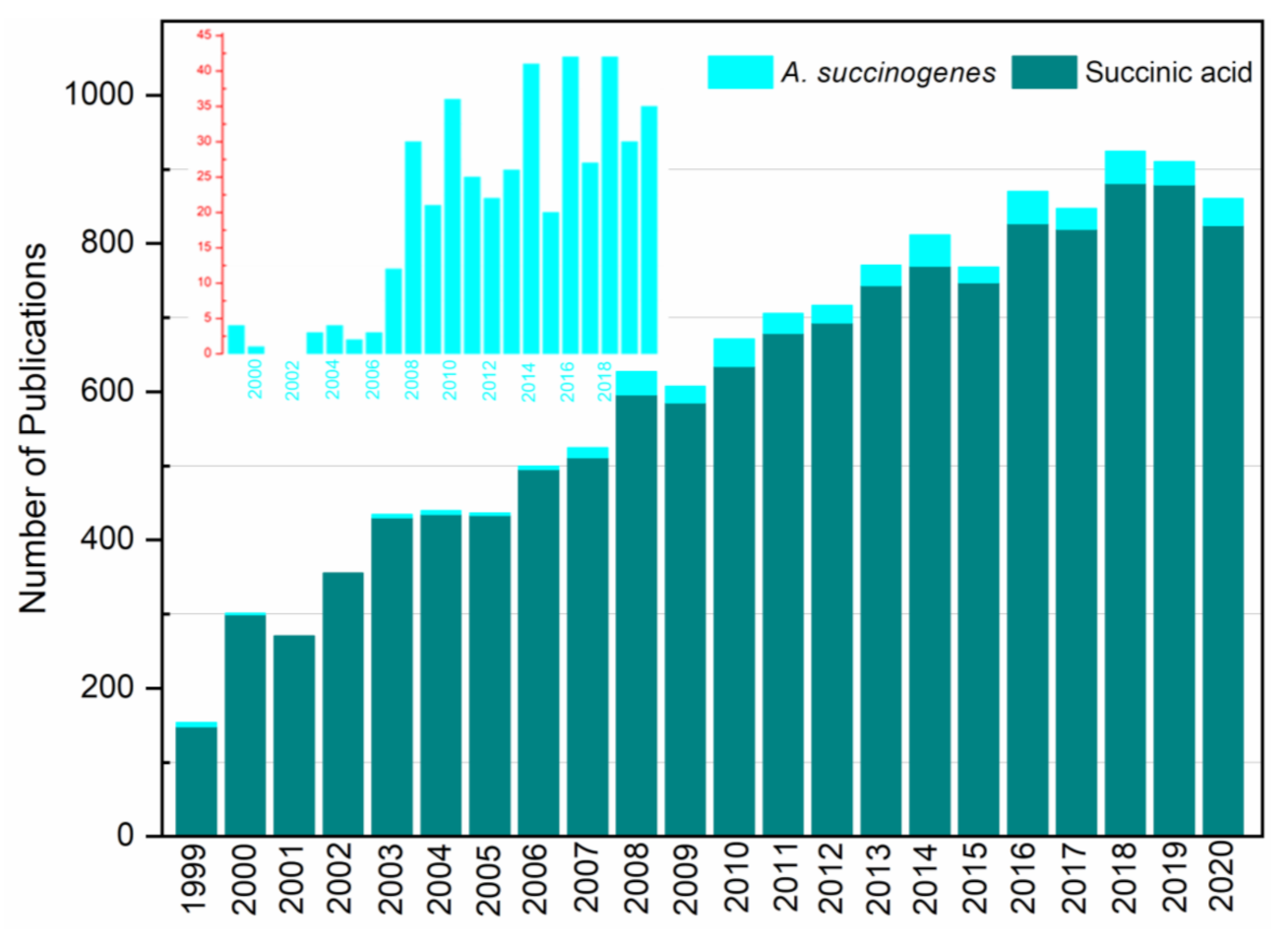
2. Major R&D Advancements on SA and A. succinogenes
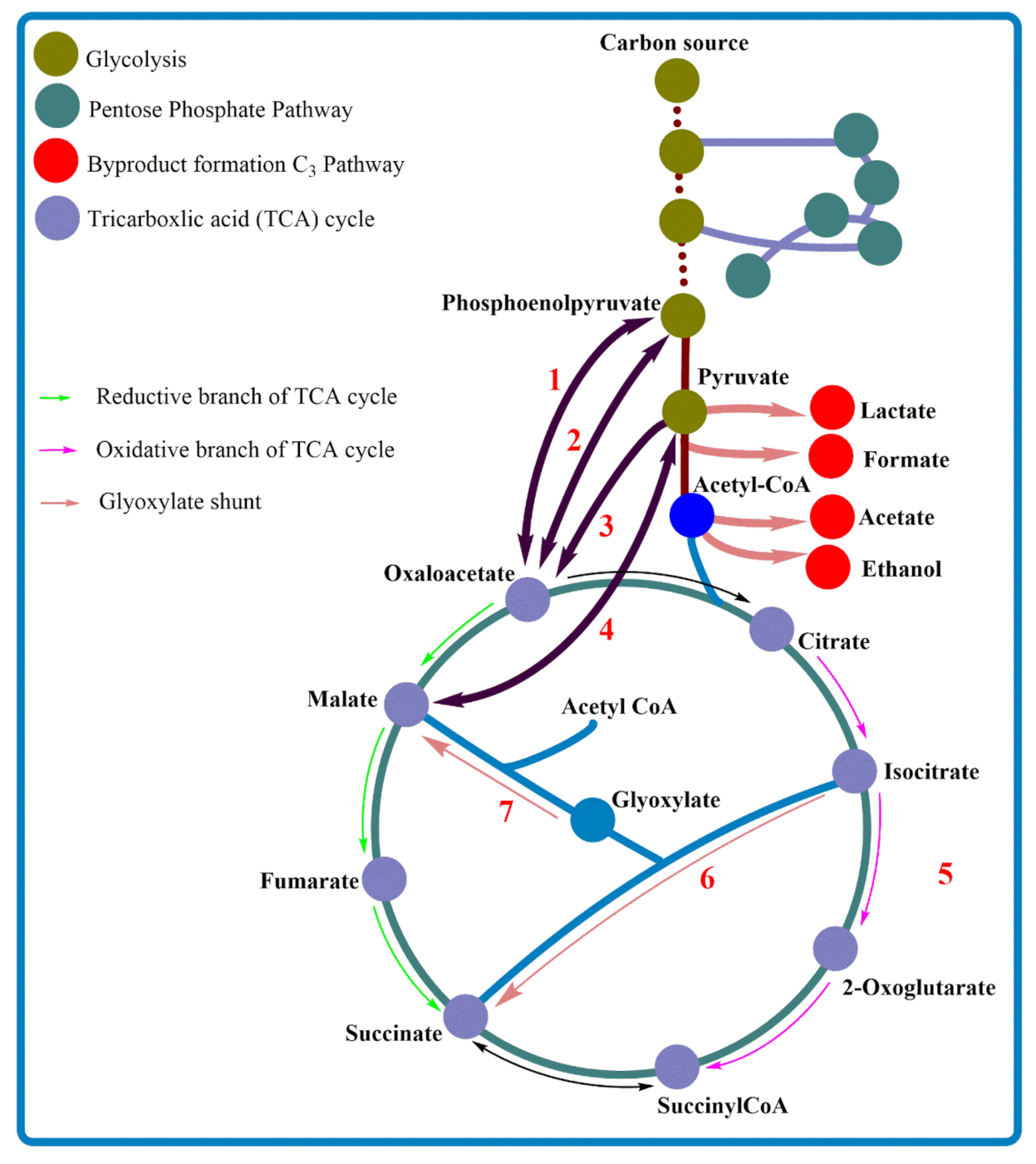
3. Succinic Acid Production Pathways
4. Metabolic Models of SA Producers
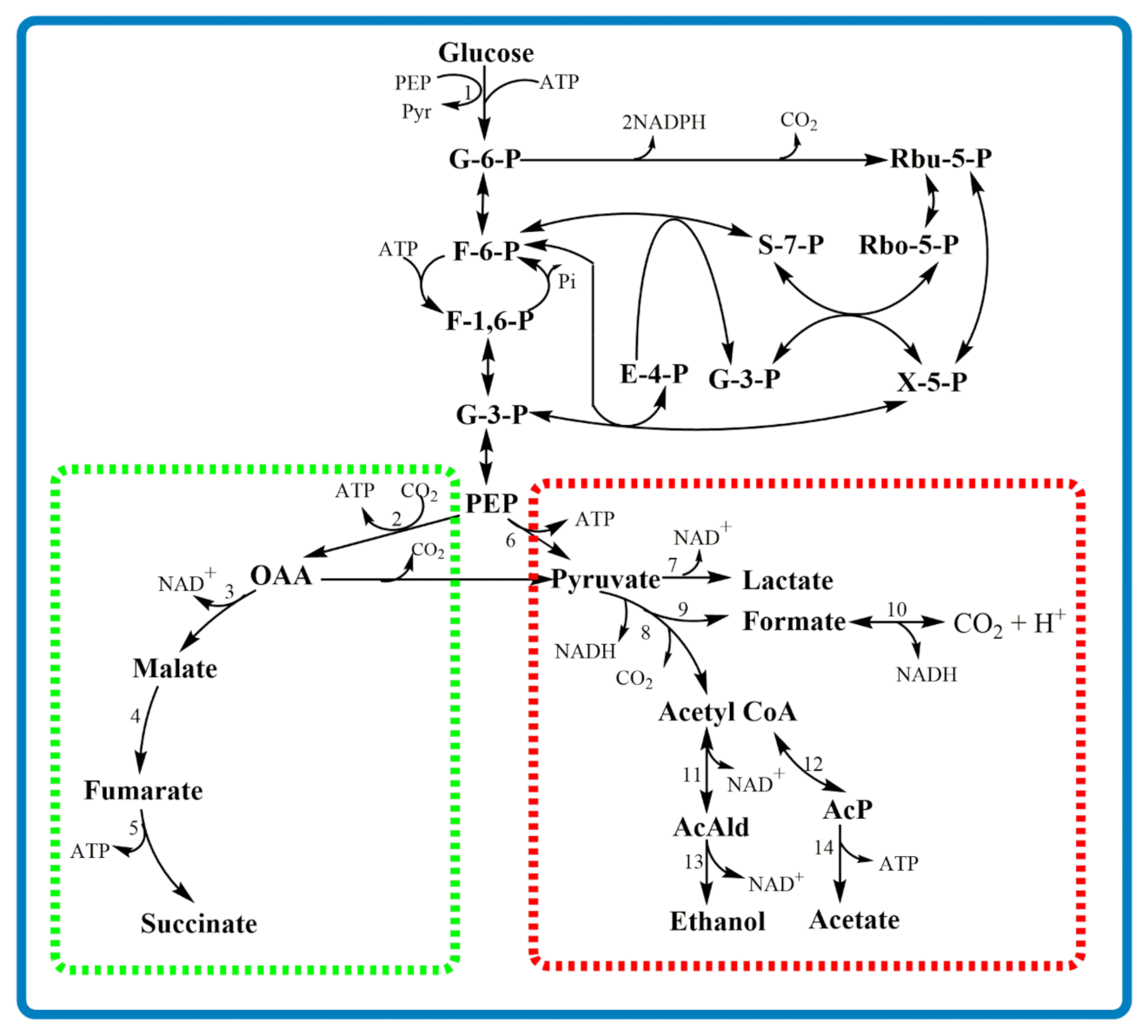
5. Attempts at Metabolic Modeling of A. succinogenes
6. Perspectives and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| AcAld | acetaldehyde |
| ATP | adenosine triphosphate |
| C4 | 4 carbon |
| e.g., | example |
| F-6-P | fructose-6-phosphate |
| G-3-P | glyceraldehyde-3-phosphate |
| GEM | genome-scale metabolic model |
| HPC | high-performance computing |
| IDH | isocitrate dehydrogenase |
| NADH | nicotinamide adenine dinucleotide |
| OAA | oxaloacetate |
| PEP | phosphoenolpyruvate |
| PPP | pentose phosphate pathway |
| pyc | pyruvate carboxylase gene |
| R&D | research and development |
| Ru-5-P | ribulose-5-phosphate |
| SA | succinic acid |
| TCA | tricarboxylic acid |
| AcP | acetylphosphate |
| C3 | 3 carbon |
| CO2 | carbon dioxide |
| F-1,6-P | fructose-1,6-bisphosphate |
| FBA | flux balance analysis |
| G-6-P | glucose-6-phosphate |
| GS | glyoxylate shunt |
| HT | high throughput |
| MFA | metabolic flux analysis |
| NADPH | nicotinamide adenine dinucleotide phosphate |
| OPR | open reading frame |
| Pi | inorganic phosphate |
| ptsG | PEP-dependent phosphotransferase system glucose-specific gene |
| Pyr | pyruvate |
| Rbo-5-P | ribose-5-phosphate |
| S-7-P | sedoheptulose-7-phosphate |
| SDH | succinate dehydrogenase |
| X-5-P | xylulose-5-phosphate |
References
- Dessie, W.; Luo, X.; Tang, J.; Tang, W.; Wang, M.; Qin, Z.; Tan, Y. Towards Full Utilization of Biomass Resources: A Case Study on Industrial Hemp Residue and Spent Mushroom Substrate. Processes 2021, 9, 1200. [Google Scholar] [CrossRef]
- Werpy, T.; Petersen, G. Top Value Added Chemicals from Biomass: Volume I—Results of Screening for Potential Candidates from Sugars and Synthesis Gas; National Renewable Energy Lab.: Golden, CO, USA, 2004. [Google Scholar]
- Bozell, J.J.; Petersen, G.R. Technology development for the production of biobased products from biorefinery carbohydrates—The US Department of Energy’s “Top 10” revisited. Green Chem. 2010, 12, 539–554. [Google Scholar] [CrossRef]
- Pateraki, C.; Patsalou, M.; Vlysidis, A.; Kopsahelis, N.; Webb, C.; Koutinas, A.A.; Koutinas, M. Actinobacillus succinogenes: Advances on succinic acid production and prospects for development of integrated biorefineries. Biochem. Eng. J. 2016, 112, 285–303. [Google Scholar] [CrossRef]
- Dessie, W.; Luo, X.; Wang, M.; Feng, L.; Liao, Y.; Wang, Z.; Yong, Z.; Qin, Z. Current advances on waste biomass transformation into value-added products. Appl. Microbiol. Biotechnol. 2020, 104, 4757–4770. [Google Scholar] [CrossRef]
- Bender, T.A.; Dabrowski, J.A.; Gagné, M.R. Homogeneous catalysis for the production of low-volume, high-value chemicals from biomass. Nat. Rev. Chem. 2018, 2, 35–46. [Google Scholar] [CrossRef]
- Zeikus, J.G.; Jain, M.K.; Elankovan, P. Biotechnology of succinic acid production and markets for derived industrial products. Appl. Microbiol. Biotechnol. 1999, 51, 545–552. [Google Scholar] [CrossRef]
- McKinlay, J.B.; Laivenieks, M.; Schindler, B.D.; McKinlay, A.A.; Siddaramappa, S.; Challacombe, J.F.; Lowry, S.R.; Clum, A.; Lapidus, A.L.; Burkhart, K.B.; et al. A genomic perspective on the potential of Actinobacillus succinogenes for industrial succinate production. BMC Genom. 2010, 11, 680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cvijovic, M.; Bordel, S.; Nielsen, J. Mathematical models of cell factories: Moving towards the core of industrial biotechnology. Microb. Biotechnol. 2011, 4, 572–584. [Google Scholar] [CrossRef] [Green Version]
- Badri, A.; Srinivasan, A.; Raman, K. In Silico Approaches to Metabolic Engineering. In Current Developments in Biotechnology and Bioengineering; Gunasekaran, P., Noronha, S., Pandey, A., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 161–200. [Google Scholar] [CrossRef]
- Dessie, W.; Xin, F.; Zhang, W.; Jiang, Y.; Wu, H.; Ma, J.; Jiang, M. Opportunities, challenges, and future perspectives of succinic acid production by Actinobacillus succinogenes. Appl. Microbiol. Biotechnol. 2018, 102, 9893–9910. [Google Scholar] [CrossRef]
- Lee, S.Y.; Kim, J.M.; Song, H.; Lee, J.W.; Kim, T.Y.; Jang, Y.-S. From genome sequence to integrated bioprocess for succinic acid production by Mannheimia succiniciproducens. Appl. Microbiol. Biotechnol. 2008, 79, 11–22. [Google Scholar] [CrossRef]
- Milne, C.B.; Kim, P.-J.; Eddy, J.A.; Price, N.D. Accomplishments in genome-scale in silico modeling for industrial and medical biotechnology. Biotechnol. J. 2009, 4, 1653–1670. [Google Scholar] [CrossRef]
- Valderrama-Gomez, M.A.; Kreitmayer, D.; Wolf, S.; Marin-Sanguino, A.; Kremling, A. Application of theoretical methods to increase succinate production in engineered strains. Bioprocess Biosyst. Eng. 2017, 40, 479–497. [Google Scholar] [CrossRef]
- Smyth, H.F.; Carpenter, C.P.; Weil, C.S. Range-finding toxicity data: List IV. AMA Arch. Indust. Hyg. Occup. Med. 1951, 4, 119–122. [Google Scholar]
- Saxena, R.K.; Saran, S.; Isar, J.; Kaushik, R. Production and Applications of Succinic Acid. In Current Developments in Biotechnology and Bioengineering; Pandey, A., Negi, S., Soccol, C.R., Eds.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 601–630. [Google Scholar] [CrossRef]
- Li, J.; Rong, L.; Zhao, Y.; Li, S.; Zhang, C.; Xiao, D.; Foo, J.L.; Yu, A. Next-generation metabolic engineering of non-conventional microbial cell factories for carboxylic acid platform chemicals. Biotechnol. Adv. 2020, 43, 107605. [Google Scholar] [CrossRef] [PubMed]
- Cok, B.; Tsiropoulos, I.; Roes, A.L.; Patel, M.K. Succinic acid production derived from carbohydrates: An energy and greenhouse gas assessment of a platform chemical toward a bio-based economy. Biofuels Bioprod. Biorefin. 2014, 8, 16–29. [Google Scholar] [CrossRef]
- Guettler, M.V.; Jain, M.K.; Rumler, D. Method for Making Succinic Acid, Bacterial Variants for Use in the Process, and Methods for Obtaining Variants. U.S. Patent 5,573,931, 12 November 1996. [Google Scholar]
- Guettler, M.V.; Rumler, D.; Jain, M.K. Actinobacillus succinogenes sp. nov., a novel succinic-acid-producing strain from the bovine rumen. Int. J. Syst. Evol. Microbiol. 1999, 49, 207–216. [Google Scholar] [CrossRef] [Green Version]
- Wolin, M.J. Metabolic interactions among intestinal microorganisms. Am. J. Clin. Nutr. 1974, 27, 1320–1328. [Google Scholar] [CrossRef] [PubMed]
- Miller, T.L. The pathway of formation of acetate and succinate from pyruvate by Bacteroides succinogenes. Arch. Microbiol. 1978, 117, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Zeikus, J.G. Chemical and fuel production by anaerobic bacteria. Annu. Rev. Microbiol. 1980, 34, 423–464. [Google Scholar] [CrossRef]
- Glassner, D.A.; Datta, R. Process for the Production and Purification of Succinic Acid. U.S. Patent 5,143,834, 1 September 1992. [Google Scholar]
- der Werf, M.J.V.; Guettler, M.V.; Jain, M.K.; Zeikus, J.G. Environmental and physiological factors affecting the succinate product ratio during carbohydrate fermentation by Actinobacillus sp. 130Z. Arch. Microbiol. 1997, 167, 332–342. [Google Scholar] [CrossRef]
- Kim, P.; Laivenieks, M.; McKinlay, J.; Vieille, C.; Gregory Zeikus, J. Construction of a shuttle vector for the overexpression of recombinant proteins in Actinobacillus succinogenes. Plasmid 2004, 51, 108–115. [Google Scholar] [CrossRef]
- JGI. Available online: https://genome.jgi.doe.gov/portal/actsu/actsu.info.html (accessed on 7 January 2021).
- Joshi, R.V.; Schindler, B.D.; McPherson, N.R.; Tiwari, K.; Vieille, C. Development of a markerless knockout method for Actinobacillus succinogenes. Appl. Environ. Microbiol. 2014, 80, 3053–3061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Shen, N.; Qin, Y.; Zhu, J.; Li, Y.; Wu, J.; Jiang, M.-G. Complete Genome Sequence of Actinobacillus succinogenes GXAS137, a Highly Efficient Producer of Succinic Acid. Gen. Announc. 2018, 6, e01562-17. [Google Scholar] [CrossRef] [Green Version]
- Pereira, B.; Miguel, J.; Vilaça, P.; Soares, S.; Rocha, I.; Carneiro, S. Reconstruction of a genome-scale metabolic model for Actinobacillus succinogenes 130Z. BMC Syst. Biol. 2018, 12, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitkänen, E.; Jouhten, P.; Rousu, J. Inferring branching pathways in genome-scale metabolic networks. BMC Syst. Biol. 2009, 3, 103. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Cai, J.; Shang, X.; Wang, B.; Liu, S.; Chai, X.; Tan, T.; Zhang, Y.; Wen, T. A new genome-scale metabolic model of Corynebacterium glutamicum and its application. Biotechnol. Biofuels 2017, 10, 169. [Google Scholar] [CrossRef]
- Miklóssy, I.; Bodor, Z.; Sinkler, R.; Orbán, K.C.; Lányi, S.; Albert, B. In silico and in vivo stability analysis of a heterologous biosynthetic pathway for 1,4-butanediol production in metabolically engineered E. coli. J. Biomol. Str. Dyn. 2017, 35, 1874–1889. [Google Scholar] [CrossRef] [PubMed]
- Vuoristo, K.S.; Mars, A.E.; Sanders, J.P.M.; Eggink, G.; Weusthuis, R.A. Metabolic Engineering of TCA Cycle for Production of Chemicals. Trends Biotechnol. 2016, 34, 191–197. [Google Scholar] [CrossRef]
- Nghiem, N.P.; Kleff, S.; Schwegmann, S. Succinic Acid: Technology Development and Commercialization. Fermentation 2017, 3, 26. [Google Scholar] [CrossRef]
- Song, H.; Lee, S.Y. Production of succinic acid by bacterial fermentation. Enzym. Microb. Technol. 2006, 39, 352–361. [Google Scholar] [CrossRef]
- Cheng, K.-K.; Wang, G.-Y.; Zeng, J.; Zhang, J.-A. Improved Succinate Production by Metabolic Engineering. Biomed. Res. Int. 2013, 2013, 538790. [Google Scholar] [CrossRef] [Green Version]
- Vemuri, G.N.; Eiteman, M.A.; Altman, E. Effects of Growth Mode and Pyruvate Carboxylase on Succinic Acid Production by Metabolically Engineered Strains of Escherichia coli. Appl. Environ. Microbiol. 2002, 68, 1715–1727. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Wang, Z.; Zhu, N.; Wang, B.; Chen, T.; Zhao, X. Metabolic engineering of Escherichia coli and in silico comparing of carboxylation pathways for high succinate productivity under aerobic conditions. Microbiol. Res. 2014, 169, 432–440. [Google Scholar] [CrossRef]
- Kim, P.; Laivenieks, M.; Vieille, C.; Zeikus, J.G. Effect of Overexpression of Actinobacillus succinogenes Phosphoenolpyruvate Carboxykinase on Succinate Production in Escherichia coli. Appl. Environ. Microbiol. 2004, 70, 1238–1241. [Google Scholar] [CrossRef] [Green Version]
- Cui, Z.; Gao, C.; Li, J.; Hou, J.; Lin, C.S.K.; Qi, Q. Engineering of unconventional yeast Yarrowia lipolytica for efficient succinic acid production from glycerol at low pH. Metab. Eng. 2017, 42, 126–133. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Vadali, R.V.; Bennett, G.N.; San, K.-Y. Increasing the Acetyl-CoA Pool in the Presence of Overexpressed Phosphoenolpyruvate Carboxylase or Pyruvate Carboxylase Enhances Succinate Production in Escherichia coli. Biotechnol. Prog. 2004, 20, 1599–1604. [Google Scholar] [CrossRef]
- Li, Y.; Li, M.; Zhang, X.; Yang, P.; Liang, Q.; Qi, Q. A novel whole-phase succinate fermentation strategy with high volumetric productivity in engineered Escherichia coli. Bioresour. Technol. 2013, 149, 333–340. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Zheng, P.; Yu, F.; Yang, Q. A two-stage process for succinate production using genetically engineered Corynebacterium acetoacidophilum. Process Biochem. 2015, 50, 1692–1700. [Google Scholar] [CrossRef]
- Tsuji, A.; Okada, S.; Hols, P.; Satoh, E. Metabolic engineering of Lactobacillus plantarum for succinic acid production through activation of the reductive branch of the tricarboxylic acid cycle. Enzym. Microb. Technol. 2013, 53, 97–103. [Google Scholar] [CrossRef]
- Jojima, T.; Noburyu, R.; Suda, M.; Okino, S.; Yukawa, H.; Inui, M. Improving Process Yield in Succinic Acid Production by Cell Recycling of Recombinant Corynebacterium glutamicum. Fermentation 2016, 2, 5. [Google Scholar] [CrossRef] [Green Version]
- Yuzbashev, T.V.; Yuzbasheva, E.Y.; Sobolevskaya, T.I.; Laptev, I.A.; Vybornaya, T.V.; Larina, A.S.; Matsui, K.; Fukui, K.; Sineoky, S.P. Production of succinic acid at low pH by a recombinant strain of the aerobic yeast Yarrowia lipolytica. Biotechnol. Bioeng. 2010, 107, 673–682. [Google Scholar] [CrossRef] [PubMed]
- Prabhu, A.A.; Ledesma-Amaro, R.; Lin, C.S.K.; Coulon, F.; Thakur, V.K.; Kumar, V. Bioproduction of succinic acid from xylose by engineered Yarrowia lipolytica without pH control. Biotechnol. Biofuels 2020, 13, 113. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Zhang, B.; Chen, T.; Wang, Z.; Tang, Y.-J.; Zhao, X. Directed pathway evolution of the glyoxylate shunt in Escherichia coli for improved aerobic succinate production from glycerol. J. Ind. Microbiol. Biotechnol. 2013, 40, 1461–1475. [Google Scholar] [CrossRef]
- Zhu, L.-W.; Li, X.-H.; Zhang, L.; Li, H.-M.; Liu, J.-H.; Yuan, Z.-P.; Chen, T.; Tang, Y.-J. Activation of glyoxylate pathway without the activation of its related gene in succinate-producing engineered Escherichia coli. Metab. Eng. 2013, 20, 9–19. [Google Scholar] [CrossRef]
- Arikawa, Y.; Kobayashi, M.; Kodaira, R.; Shimosaka, M.; Muratsubaki, H.; Enomoto, K.; Okazaki, M. Isolation of sake yeast strains possessing various levels of succinate- and/or malate-producing abilities by gene disruption or mutation. J. Biosci. Bioeng. 1999, 87, 333–339. [Google Scholar] [CrossRef]
- Singh, A.; Cher Soh, K.; Hatzimanikatis, V.; Gill, R.T. Manipulating redox and ATP balancing for improved production of succinate in E. coli. Metab. Eng. 2011, 13, 76–81. [Google Scholar] [CrossRef]
- Mao, Y.; Li, G.; Chang, Z.; Tao, R.; Cui, Z.; Wang, Z.; Tang, Y.-J.; Chen, T.; Zhao, X. Metabolic engineering of Corynebacterium glutamicum for efficient production of succinate from lignocellulosic hydrolysate. Biotechnol. Biofuels 2018, 11, 95. [Google Scholar] [CrossRef]
- Raab, A.M.; Gebhardt, G.; Bolotina, N.; Weuster-Botz, D.; Lang, C. Metabolic engineering of Saccharomyces cerevisiae for the biotechnological production of succinic acid. Metab. Eng. 2010, 12, 518–525. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Bennett, G.N.; San, K.-Y. Metabolic engineering of aerobic succinate production systems in Escherichia coli to improve process productivity and achieve the maximum theoretical succinate yield. Metab. Eng. 2005, 7, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Litsanov, B.; Kabus, A.; Brocker, M.; Bott, M. Efficient aerobic succinate production from glucose in minimal medium with Corynebacterium glutamicum. Microb. Biotechnol. 2012, 5, 116–128. [Google Scholar] [CrossRef] [Green Version]
- Khodayari, A.; Chowdhury, A.; Maranas, C.D. Succinate Overproduction: A Case Study of Computational Strain Design Using a Comprehensive Escherichia coli Kinetic Model. Front. Bioeng. Biotechnol. 2015, 2, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raab, A.M.; Lang, C. Oxidative versus reductive succinic acid production in the yeast Saccharomyces cerevisiae. Bioeng. Bugs 2011, 2, 120–123. [Google Scholar] [CrossRef]
- Fang, X.; Lloyd, C.J.; Palsson, B.O. Reconstructing organisms in silico: Genome-scale models and their emerging applications. Nat. Rev. Microbiol. 2020, 18, 731–743. [Google Scholar] [CrossRef]
- Sarkar, D.; Maranas, C.D. Engineering microbial chemical factories using metabolic models. BMC Chem. Eng. 2019, 1, 22. [Google Scholar] [CrossRef]
- Landon, S.; Rees-Garbutt, J.; Marucci, L.; Grierson, C. Genome-driven cell engineering review: In vivo and in silico metabolic and genome engineering. Essays Biochem. 2019, 63, 267–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jouhten, P. Metabolic modelling in the development of cell factories by synthetic biology. Comput. Struct. Biotechnol. J. 2012, 3, e201210009. [Google Scholar] [CrossRef] [Green Version]
- Bordbar, A.; Monk, J.M.; King, Z.A.; Palsson, B.O. Constraint-based models predict metabolic and associated cellular functions. Nat. Rev. Genet. 2014, 15, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Fleming, R.M.T.; Thiele, I.; Provan, G.; Nasheuer, H.P. Integrated stoichiometric, thermodynamic and kinetic modelling of steady state metabolism. J. Theor. Biol. 2010, 264, 683–692. [Google Scholar] [CrossRef] [Green Version]
- Copeland, W.B.; Bartley, B.A.; Chandran, D.; Galdzicki, M.; Kim, K.H.; Sleight, S.C.; Maranas, C.D.; Sauro, H.M. Computational tools for metabolic engineering. Metab. Eng. 2012, 14, 270–280. [Google Scholar] [CrossRef] [Green Version]
- Jing, L.S.; Shah, F.F.M.; Mohamad, M.S.; Hamran, N.L.; Salleh, A.H.M.; Deris, S.; Alashwal, H. Database and tools for metabolic network analysis. Biotechnol. Bioprocess Eng. 2014, 19, 568–585. [Google Scholar] [CrossRef]
- O’Shea, K.; Misra, B.B. Software tools, databases and resources in metabolomics: Updates from 2018 to 2019. Metabolomics 2020, 16, 36. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.M.; Saha, R. Computational Approaches on Stoichiometric and Kinetic Modeling for Efficient Strain Design. In Synthetic Metabolic Pathways: Methods and Protocols; Jensen, M.K., Keasling, J.D., Eds.; Springer: New York, NY, USA, 2018; pp. 63–82. [Google Scholar] [CrossRef]
- Orth, J.D.; Thiele, I.; Palsson, B.Ø. What is flux balance analysis? Nat. Biotechnol. 2010, 28, 245–248. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Hong, S.H.; Moon, S.Y. In silico metabolic pathway analysis and design: Succinic acid production by metabolically engineered Escherichia coli as an example. Genome Inform. 2002, 13, 214–223. [Google Scholar]
- Hong, S.H.; Kim, J.S.; Lee, S.Y.; In, Y.H.; Choi, S.S.; Rih, J.-K.; Kim, C.H.; Jeong, H.; Hur, C.G.; Kim, J.J. The genome sequence of the capnophilic rumen bacterium Mannheimia succiniciproducens. Nat. Biotechnol. 2004, 22, 1275–1281. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.H.; Lee, S.Y. Enhanced production of succinic acid by metabolically engineered Escherichia coli with amplified activities of malic enzyme and fumarase. Biotechnol. Bioprocess Eng. 2004, 9, 252. [Google Scholar] [CrossRef]
- Lee, S.J.; Lee, D.-Y.; Kim, T.Y.; Kim, B.H.; Lee, J.; Lee, S.Y. Metabolic Engineering of Escherichia coli for Enhanced Production of Succinic Acid, Based on Genome Comparison and In Silico Gene Knockout Simulation. Appl. Environ. Microbiol. 2005, 71, 7880–7887. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Chen, X.; Yang, Y.; Zhao, X. Genome-scale in silico aided metabolic analysis and flux comparisons of Escherichia coli to improve succinate production. Appl. Microbiol. Biotechnol. 2006, 73, 887–894. [Google Scholar] [CrossRef]
- Kim, T.Y.; Kim, H.U.; Park, J.M.; Song, H.; Kim, J.S.; Lee, S.Y. Genome-scale analysis of Mannheimia succiniciproducens metabolism. Biotechnol. Bioeng. 2007, 97, 657–671. [Google Scholar] [CrossRef]
- Kim, T.Y.; Kim, H.U.; Song, H.; Lee, S.Y. In silico analysis of the effects of H2 and CO2 on the metabolism of a capnophilic bacterium Mannheimia succiniciproducens. J. Biotechnol. 2009, 144, 184–189. [Google Scholar] [CrossRef]
- Meijer, S.; Nielsen, M.L.; Olsson, L.; Nielsen, J. Gene deletion of cytosolic ATP: Citrate lyase leads to altered organic acid production in Aspergillus niger. J. Ind. Microbiol. Biotechnol. 2009, 36, 1275–1280. [Google Scholar] [CrossRef] [PubMed]
- Agren, R.; Otero, J.M.; Nielsen, J. Genome-scale modeling enables metabolic engineering of Saccharomyces cerevisiae for succinic acid production. J. Ind. Microbiol. Biotechnol. 2013, 40, 735–747. [Google Scholar] [CrossRef] [PubMed]
- Otero, J.M.; Cimini, D.; Patil, K.R.; Poulsen, S.G.; Olsson, L.; Nielsen, J. Industrial systems biology of Saccharomyces cerevisiae enables novel succinic acid cell factory. PLoS ONE 2013, 8, e54144. [Google Scholar] [CrossRef] [Green Version]
- Becker, J.; Reinefeld, J.; Stellmacher, R.; Schäfer, R.; Lange, A.; Meyer, H.; Lalk, M.; Zelder, O.; von Abendroth, G.; Schröder, H.; et al. Systems-wide analysis and engineering of metabolic pathway fluxes in bio-succinate producing Basfia succiniciproducens. Biotechnol. Bioeng. 2013, 110, 3013–3023. [Google Scholar] [CrossRef] [PubMed]
- Rafieenia, R. Metabolic capabilities of Actinobacillus succinogenes for succinic acid production. Braz. J. Chem. Eng. 2014, 31, 859–865. [Google Scholar] [CrossRef]
- Chua, P.S.; Salleh, A.H.M.; Mohamad, M.S.; Deris, S.; Omatu, S.; Yoshioka, M. Identifying a gene knockout strategy using a hybrid of the bat algorithm and flux balance analysis to enhance the production of succinate and lactate in Escherichia coli. Biotechnol. Bioprocess Eng. 2015, 20, 349–357. [Google Scholar] [CrossRef]
- Jian, X.; Li, N.; Zhang, C.; Hua, Q. In silico profiling of cell growth and succinate production in Escherichia coli NZN111. Bioresour. Bioprocess 2016, 3, 48. [Google Scholar] [CrossRef] [Green Version]
- Mienda, B.S.; Shamsir, M.S.; Illias, R.M. Model-aided atpE gene knockout strategy in Escherichia coli for enhanced succinic acid production from glycerol. J. Biomol. Str. Dyn. 2016, 34, 1705–1716. [Google Scholar] [CrossRef] [PubMed]
- Mienda, B.S.; Shamsir, M.S.; Illias, R.M. Model-guided metabolic gene knockout of gnd for enhanced succinate production in Escherichia coli from glucose and glycerol substrates. Comput. Biol. Chem. 2016, 61, 130–137. [Google Scholar] [CrossRef]
- Choi, S.; Song, H.; Lim, S.W.; Kim, T.Y.; Ahn, J.H.; Lee, J.W.; Lee, M.-H.; Lee, S.Y. Highly selective production of succinic acid by metabolically engineered Mannheimia succiniciproducens and its efficient purification. Biotechnol. Bioeng. 2016, 113, 2168–2177. [Google Scholar] [CrossRef]
- Lee, J.W.; Yi, J.; Kim, T.Y.; Choi, S.; Ahn, J.H.; Song, H.; Lee, M.-H.; Lee, S.Y. Homo-succinic acid production by metabolically engineered Mannheimia succiniciproducens. Metab. Eng. 2016, 38, 409–417. [Google Scholar] [CrossRef] [PubMed]
- Angeles-Martinez, L.; Theodoropoulos, C. Estimation of flux distribution in metabolic networks accounting for thermodynamic constraints: The effect of equilibrium vs. blocked reactions. Biochem. Eng. J. 2016, 105, 347–357. [Google Scholar] [CrossRef]
- Jian, X.; Li, N.; Chen, Q.; Hua, Q. Model-guided identification of novel gene amplification targets for improving succinate production in Escherichia coli NZN111. Integr. Biol. 2017, 9, 830–835. [Google Scholar] [CrossRef]
- St. John, P.C.; Crowley, M.F.; Bomble, Y.J. Efficient estimation of the maximum metabolic productivity of batch systems. Biotechnol. Biofuels 2017, 10, 28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tafur Rangel, A.E.; Camelo Valera, L.C.; Gómez Ramírez, J.M.; González Barrios, A.F. Effects of metabolic engineering on downstream processing operational cost and energy consumption: The case of Escherichia coli’s glycerol conversion to succinic acid. J. Chem. Technol. Biotechnol. 2018, 93, 2011–2020. [Google Scholar] [CrossRef]
- Mohd Daud, K.; Zakaria, Z.; Shah, Z.A.; Mohamad, M.; Deris, S.; Omatu, S.; Corchado Rodríguez, J. A hybrid of differential search algorithm and flux balance analysis to: Identify knockout strategies for in silico optimization of metabolites production. Int. J. Adv. Soft Comput. Appl. 2018, 10, 84–107. [Google Scholar]
- Mienda, B.S. Escherichia coli genome-scale metabolic gene knockout of lactate dehydrogenase (ldhA), increases succinate production from glycerol. J. Biomol. Str. Dyn. 2018, 36, 3680–3686. [Google Scholar] [CrossRef]
- Arif, M.A.; Mohamad, M.S.; Abd Latif, M.S.; Deris, S.; Remli, M.A.; Mohd Daud, K.; Ibrahim, Z.; Omatu, S.; Corchado, J.M. A hybrid of Cuckoo Search and Minimization of Metabolic Adjustment to optimize metabolites production in genome-scale models. Comput. Biol. Med. 2018, 102, 112–119. [Google Scholar] [CrossRef]
- Nag, A.; St. John, P.C.; Crowley, M.F.; Bomble, Y.J. Prediction of reaction knockouts to maximize succinate production by Actinobacillus succinogenes. PLoS ONE 2018, 13, e0189144. [Google Scholar] [CrossRef] [Green Version]
- Widiastuti, H.; Lee, N.-R.; Karimi, I.A.; Lee, D.-Y. Genome-Scale In Silico Analysis for Enhanced Production of Succinic Acid in Zymomonas mobilis. Processes 2018, 6, 30. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.K.; Mohamad, M.S.; Choon, Y.W.; Daud, K.M.; Nasarudin, N.A.; Ismail, M.A.; Ibrahim, Z.; Napis, S.; Sinnott, R.O. Comparison of Optimization-Modelling Methods for Metabolites Production in Escherichia coli. J. Integr. Bioinform. 2020, 17, 20190073. [Google Scholar] [CrossRef] [PubMed]
- Upton, D.J.; McQueen-Mason, S.J.; Wood, A.J. In silico evolution of Aspergillus niger organic acid production suggests strategies for switching acid output. Biotechnol. Biofuels 2020, 13, 27. [Google Scholar] [CrossRef]
- Ahn, J.H.; Seo, H.; Park, W.; Seok, J.; Lee, J.A.; Kim, W.J.; Kim, G.B.; Kim, K.-J.; Lee, S.Y. Enhanced succinic acid production by Mannheimia employing optimal malate dehydrogenase. Nat. Commun. 2020, 11, 1970. [Google Scholar] [CrossRef] [Green Version]
- Mienda, B.S. Genome-scale metabolic models as platforms for strain design and biological discovery. J. Biomol. Str. Dyn. 2017, 35, 1863–1873. [Google Scholar] [CrossRef] [PubMed]
- Famili, I.; Forster, J.; Nielsen, J.; Palsson, B.O. Saccharomyces cerevisiae phenotypes can be predicted by using constraint-based analysis of a genome-scale reconstructed metabolic network. Proc. Natl. Acad. Sci. USA 2003, 100, 13134–13139. [Google Scholar] [CrossRef] [Green Version]
- Kjeldsen, K.R.; Nielsen, J. In silico genome-scale reconstruction and validation of the Corynebacterium glutamicum metabolic network. Biotechnol. Bioeng. 2009, 102, 583–597. [Google Scholar] [CrossRef] [PubMed]
- Shinfuku, Y.; Sorpitiporn, N.; Sono, M.; Furusawa, C.; Hirasawa, T.; Shimizu, H. Development and experimental verification of a genome-scale metabolic model for Corynebacterium glutamicum. Microb. Cell Fact. 2009, 8, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mei, J.; Xu, N.; Ye, C.; Liu, L.; Wu, J. Reconstruction and analysis of a genome-scale metabolic network of Corynebacterium glutamicum S9114. Gene 2016, 575, 615–622. [Google Scholar] [CrossRef]
- Reed, J.L.; Vo, T.D.; Schilling, C.H.; Palsson, B.O. An expanded genome-scale model of Escherichia coli K-12 (iJR904 GSM/GPR). Genome Biol. 2003, 4, R54. [Google Scholar] [CrossRef] [Green Version]
- Feist, A.M.; Henry, C.S.; Reed, J.L.; Krummenacker, M.; Joyce, A.R.; Karp, P.D.; Broadbelt, L.J.; Hatzimanikatis, V.; Palsson, B.Ø. A genome-scale metabolic reconstruction for Escherichia coli K-12 MG1655 that accounts for 1260 ORFs and thermodynamic information. Mol. Syst. Biol. 2007, 3, 121. [Google Scholar] [CrossRef]
- Orth, J.D.; Conrad, T.M.; Na, J.; Lerman, J.A.; Nam, H.; Feist, A.M.; Palsson, B.Ø. A comprehensive genome-scale reconstruction of Escherichia coli metabolism—2011. Mol. Syst. Biol. 2011, 7, 535. [Google Scholar] [CrossRef]
- O’Brien, E.J.; Lerman, J.A.; Chang, R.L.; Hyduke, D.R.; Palsson, B.Ø. Genome-scale models of metabolism and gene expression extend and refine growth phenotype prediction. Mol. Syst. Biol. 2013, 9, 693. [Google Scholar] [CrossRef]
- Nookaew, I.; Jewett, M.C.; Meechai, A.; Thammarongtham, C.; Laoteng, K.; Cheevadhanarak, S.; Nielsen, J.; Bhumiratana, S. The genome-scale metabolic model iIN800 of Saccharomyces cerevisiae and its validation: A scaffold to query lipid metabolism. BMC Syst. Biol. 2008, 2, 71. [Google Scholar] [CrossRef] [Green Version]
- Mo, M.L.; Palsson, B.Ø.; Herrgård, M.J. Connecting extracellular metabolomic measurements to intracellular flux states in yeast. BMC Syst. Biol. 2009, 3, 37. [Google Scholar] [CrossRef] [Green Version]
- Österlund, T.; Nookaew, I.; Bordel, S.; Nielsen, J. Mapping condition-dependent regulation of metabolism in yeast through genome-scale modeling. BMC Syst. Biol. 2013, 7, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradfield, M.F.A.; Nicol, W. The pentose phosphate pathway leads to enhanced succinic acid flux in biofilms of wild-type Actinobacillus succinogenes. Appl. Microbiol. Biotechnol. 2016, 100, 9641–9652. [Google Scholar] [CrossRef] [Green Version]
- Pateraki, C.; Almqvist, H.; Ladakis, D.; Lidén, G.; Koutinas, A.A.; Vlysidis, A. Modelling succinic acid fermentation using a xylose based substrate. Biochem. Eng. J. 2016, 114, 26–41. [Google Scholar] [CrossRef]
- Vlysidis, A.; Du, C.; Webb, C.; Theodoropoulos, C. Experimental and Modelling Studies of the Bioconversion of Glycerol to Succinic Acid by Actinobacillus Succinogenes. In Proceedings of the AIChE Annual Meeting, Philadelphia, PA, USA, 16–21 November 2008. [Google Scholar]
- McKinlay, J.B.; Zeikus, J.G.; Vieille, C. Insights into Actinobacillus succinogenes fermentative metabolism in a chemically defined growth medium. Appl. Environ. Microbiol. 2005, 71, 6651–6656. [Google Scholar] [CrossRef] [Green Version]
- McKinlay, J.B.; Shachar-Hill, Y.; Zeikus, J.G.; Vieille, C. Determining Actinobacillus succinogenes metabolic pathways and fluxes by NMR and GC-MS analyses of 13C-labeled metabolic product isotopomers. Metab. Eng. 2007, 9, 177–192. [Google Scholar] [CrossRef] [PubMed]
- McKinlay, J.B.; Vieille, C. 13C-metabolic flux analysis of Actinobacillus succinogenes fermentative metabolism at different NaHCO3 and H2 concentrations. Metab. Eng. 2008, 10, 55–68. [Google Scholar] [CrossRef]
- Edwards, J.S.; Palsson, B.O. Systems properties of the Haemophilus influenzae Rd metabolic genotype. J. Biol. Chem. 1999, 274, 17410–17416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tibocha-Bonilla, J.D.; Zuñiga, C.; Godoy-Silva, R.D.; Zengler, K. Advances in metabolic modeling of oleaginous microalgae. Biotechnol. Biofuels 2018, 11, 241. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Production Route | Pathway Construction | Microorganism | Reference |
|---|---|---|---|
| Reductive TCA cycle (RT) | Inactivate lactate and acetate formation pathways and overexpress pyruvate carboxylase gene (pyc) | Corynebacterium acetoacidophilum | [44] |
| Activate enzymes of RT | Lactobacillus plantarum | [45] | |
| Overexpress pyc | C. glutamicum | [46] | |
| Oxidative TCA cycle (OT) | Delete succinate dehydrogenase (SDH) gene (sdh) | Yarrowia lipolytica | [47] |
| Overexpress genes in the PPP for xylose utilization and delete sdh | Y. lipolytica | [48] | |
| Glyoxylate shunt (GS) | Reverse PEP carboxylase via deletion of sdh and overexpress genes involved in GS | E. coli | [49] |
| Activate GS via inactivation of SA biosynthetic byproduct (lactate, acetate, formate and ethanol) formation-encoding genes | E. coli | [50] | |
| RT-OT (TCA) | Disrupt genes of aconitase, fumarate reductase, alpha ketoglutarate dehydrogenase, SDH, fumarase, isocitrate lyase and fumarate reductase | S. cerevisiae | [51] |
| Delete ptsG and genes of SA biosynthetic byproducts and overexpress PEP carboxykinase | E. coli | [52] | |
| RT-GS | Overexpress genes in the PPP for xylose utilization, pyc, citrate synthase and succinate exporter | C. glutamicum | [53] |
| Delete genes of SDH (sdh1 and sdh2) and isocitrate dehydrogenase (idh1 and idh2) | S. cerevisiae | [54] | |
| OT-GS | Delete genes of SDH, IDH and acetate-producing pathway | E. coli | [55] |
| Delete genes of SDH and acetate-producing pathway and overexpression of pyc and PEP carboxylase | C. glutamicum | [56] | |
| TCA-GS (RT-OT-GS) | Delete genes of SDH and acetyl-CoA transferase and overexpression of key enzymes of RT, OT and GS | Y. lipolytica | [41] |
| Kinetic study including RT, OT, GS and other pathways | E. coli | [57] |
| Organism | In Silico Operation and Purpose | Year | Reference |
|---|---|---|---|
| E. coli | Metabolic flux analysis (301 reactions and 294 metabolites) to attain the highest in silico SA yield | 2002 | [70] |
| M. succiniciproducens | Genome-scale flux analysis (373 reactions and 352 metabolites) to determine the general genome-scale metabolic characteristics | 2004 | [71] |
| E. coli | Metabolic flux analysis (310 reactions and 295 metabolites) to predict volumetric rates of intracellular metabolites | 2004 | [72] |
| E. coli | Comparative genomic analysis to estimate the relationship between the maximum biomass and SA production in metabolically modified strains | 2005 | [73] |
| E. coli | Genome-scale in silico aided metabolic analysis and flux comparisons to determine the in silico optimal SA production pathway | 2006 | [74] |
| M. succiniciproducens | Genome-scale metabolic analysis (686 reactions and 519 metabolites) for genome-scale analysis and designing efficient metabolic engineering studies | 2007 | [75] |
| M. succiniciproducens | Constraints-based flux analysis of genome-scale metabolic model to evaluate the production patterns of various organic acids against variable rates of glucose, CO2 and H2 | 2009 | [76] |
| A. niger | Genome-scale stoichiometric metabolic model to identify target genes for metabolic manipulation and redirect the pathway towards SA production route | 2009 | [77] |
| E. coli | Genome-scale thermodynamics-based flux balance analysis to predict the maximum biomass and SA flux | 2011 | [52] |
| S. cerevisiae | Genome-scale metabolic model and flux balance analysis to establish SA overproduction strategies | 2013 | [78] |
| S. cerevisiae | Genome-scale metabolic network reconstruction to predict gene deletions that can couple enhanced biomass and SA production | 2013 | [79] |
| Basfia succiniciproducens | Metabolic flux analysis to identify undesired fluxes and improve SA yield | 2013 | [80] |
| E. coli | Metabolic network construction (65 reactions and 44 metabolites) to evaluate the effect of the carboxylation reactions on SA production | 2014 | [39] |
| A. succinogenes | Metabolic model (27 reactions and 28 metabolites) for SA production using a mixture of glucose and xylose substrates | 2014 | [81] |
| E. coli | Kinetic model prediction to predict response to multiple environmental perturbations and overproduction of SA | 2015 | [57] |
| E. coli | Optimization algorithm and flux balance analysis to identify a set of genes for deletion to improve SA and lactic acid productions | 2015 | [82] |
| E. coli | Genome-scale metabolic core model to reconstruct the metabolic fluxes and evaluate the characteristics so as to improve SA production and reduce byproduct formation | 2016 | [83] |
| E. coli | Genome-scale metabolic model and Minimization of Metabolic Adjustment algorithm to improve the strain and increase SA production using glucose and glycerol substrates | 2016 | [84] |
| E. coli | Genome-scale metabolic model to evaluate the effect of gene deletion for enhanced SA production | 2016 | [85] |
| M. succiniciproducens | Genome-scale metabolic simulations to identify gene targets to be engineered for enhanced nearly homo-SA production | 2016 | [86] |
| M. succiniciproducens | Genome-scale metabolic flux analysis, omics analyses and metabolic reconstruction to develop a high-yield homo-SA-producing strain by metabolic engineering and carbon source optimization | 2016 | [87] |
| A. succinogenes | Thermodynamically constrained metabolic flux analysis to demonstrate the effect of environmental conditions on metabolic fluxes | 2016 | [88] |
| E. coli | Simulation and reaction expression analysis to identify genetic strategies for overproduction of SA | 2017 | [89] |
| E. coli and A. succinogenes | Dynamic flux balance analysis to estimate the maximum theoretical productivity of a batch culture system | 2017 | [90] |
| E. coli | Metabolism–downstream coupled model for metabolic engineering of the strain to produce SA using glycerol substate | 2018 | [91] |
| E. coli and Z. mobilis | Hybrid of differential search algorithm and flux balance analysis to identify knockout relations for enhanced SA production | 2018 | [92] |
| E. coli | Genome-scale metabolic model to predict gene deletion for enhanced SA production using glycerol substrate | 2018 | [93] |
| E. coli and S. cerevisiae | Hybrid of optimization algorithm and genome-scale metabolic models to predict the near-optimal set of gene deletions for overproduction of SA | 2018 | [94] |
| A. succinogenes | Comprehensive carbon metabolism model (375 reactions) to analyze the metabolism and predict knockout strategies for maximum SA production with maintaining the cell growth | 2018 | [95] |
| A. succinogenes | Genome-scale metabolic model to evaluate the metabolic capability of the strain to produce SA under various conditions | 2018 | [30] |
| Zymomonas mobilis | Genome-scale metabolic model to characterize SA-producing capability and comparatively identify gene deletions for enhanced SA production | 2018 | [96] |
| E. coli | Optimization modeling to identify near-optimal knockout genes for the maximum production of SA | 2020 | [97] |
| Aspergillus niger | Integration of genome-scale metabolic model with dynamic modeling and genetic algorithm to provide simpified gene deletion strategies for the complex evolutionary goals containing multiple targets | 2020 | [98] |
| M. succiniciproducens | Flux variability scanning using genome-scale metabolic model to identify amplification target genes for improved SA production | 2020 | [99] |
| Genome Size (bp) | Total # Genes | Model Version | Year | # Genes in the Model | # Metabolites | # Reactions | ORF Coverage (%) | Reference | |
|---|---|---|---|---|---|---|---|---|---|
| A. succinogenes | 2,319,663 | 2210 | iBP722 | 2018 | 722 | 713 | 1072 | 35.00 | [30] |
| M. succiniciproducens | 2,314,078 | 2384 | - | 2004 | 335 | 352 | 373 | 14.05 | [71] |
| - | 2007 | 425 | 519 | 686 | 17.83 | [75] | |||
| C. glutamicum | 3,282,708 | 3002 | ModelCg1 | 2008 | 247 | 411 | 446 | 8.23 | [102] |
| 3,282,708 | 3002 | ModelCg2 | 2009 | 277 | 423 | 502 | 9.23 | [103] | |
| 3,292,392 | 3015 | iJM658 | 2015 | 658 | 984 | 1065 | 21.82 | [104] | |
| 3,282,708 | 3002 | iC773 | 2017 | 773 | 950 | 1207 | 25.57 | [32] | |
| E. coli | 4,641,652 | 4453 | iJR904 | 2003 | 904 | 625 | 931 | 20.30 | [105] |
| 4,641,652 | 4453 | iAF1260 | 2007 | 1260 | 1039 | 2077 | 28.30 | [106] | |
| 4,639,675 | 4325 | iJO1366 | 2011 | 1366 | 1136 | 2251 | 31.58 | [107] | |
| 4,639,675 | 4420 | iOL1650-ME | 2013 | 1541 | 6563 | 12,009 | 34.86 | [108] | |
| S. cerevisiae | 12,261,038 | 6183 | iFF708 | 2003 | 708 | 733 | 1175 | 16.00 | [101] |
| iIN800 | 2008 | 800 | 1013 | 1446 | 17.20 | [109] | |||
| iMM904 | 2009 | 904 | 1228 | 1412 | 19.65 | [110] | |||
| iTO977 | 2013 | 977 | 1353 | 1566 | 21.24 | [111] |
| Opportunities/Advancements | Gaps/Challenges | Perspectives/Recommendations |
|---|---|---|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dessie, W.; Wang, Z.; Luo, X.; Wang, M.; Qin, Z. Insights on the Advancements of In Silico Metabolic Studies of Succinic Acid Producing Microorganisms: A Review with Emphasis on Actinobacillus succinogenes. Fermentation 2021, 7, 220. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7040220
Dessie W, Wang Z, Luo X, Wang M, Qin Z. Insights on the Advancements of In Silico Metabolic Studies of Succinic Acid Producing Microorganisms: A Review with Emphasis on Actinobacillus succinogenes. Fermentation. 2021; 7(4):220. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7040220
Chicago/Turabian StyleDessie, Wubliker, Zongcheng Wang, Xiaofang Luo, Meifeng Wang, and Zuodong Qin. 2021. "Insights on the Advancements of In Silico Metabolic Studies of Succinic Acid Producing Microorganisms: A Review with Emphasis on Actinobacillus succinogenes" Fermentation 7, no. 4: 220. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation7040220