
Fermented Myriophyllum aquaticum and Lactobacillus plantarum Affect the Distribution of Intestinal Microbial Communities and Metabolic Profile in Mice

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Cultures
2.2. Solid-State Fermentation and Conditions Optimization
2.3. Animal Experiment
2.4. 16S rDNA Sequencing
2.5. Metabolomics
2.6. KEGG Pathways
2.7. Statistical Analysis
3. Results and Discussion
3.1. Analysis of Solid-State Fermentation of M. aquaticum
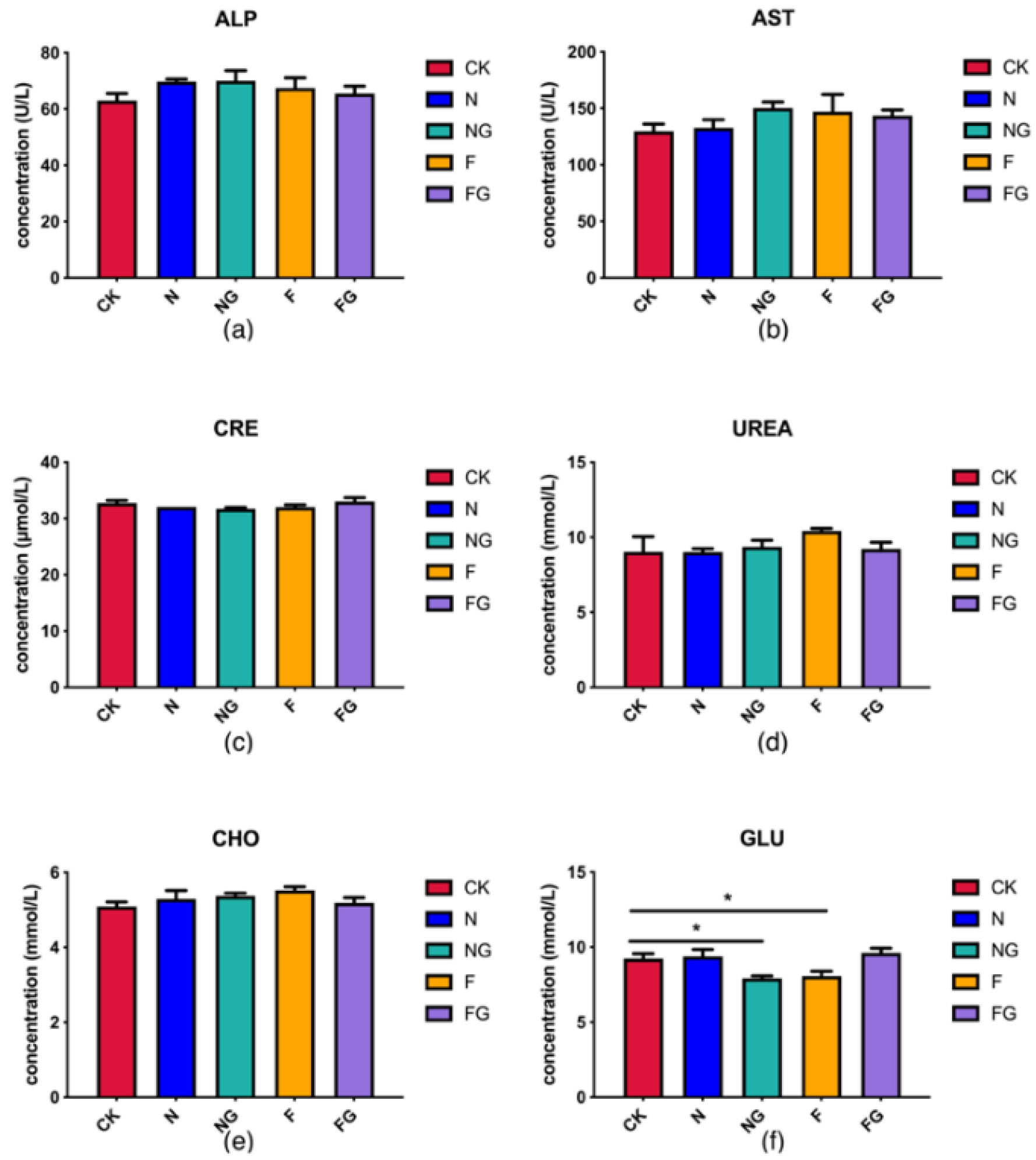
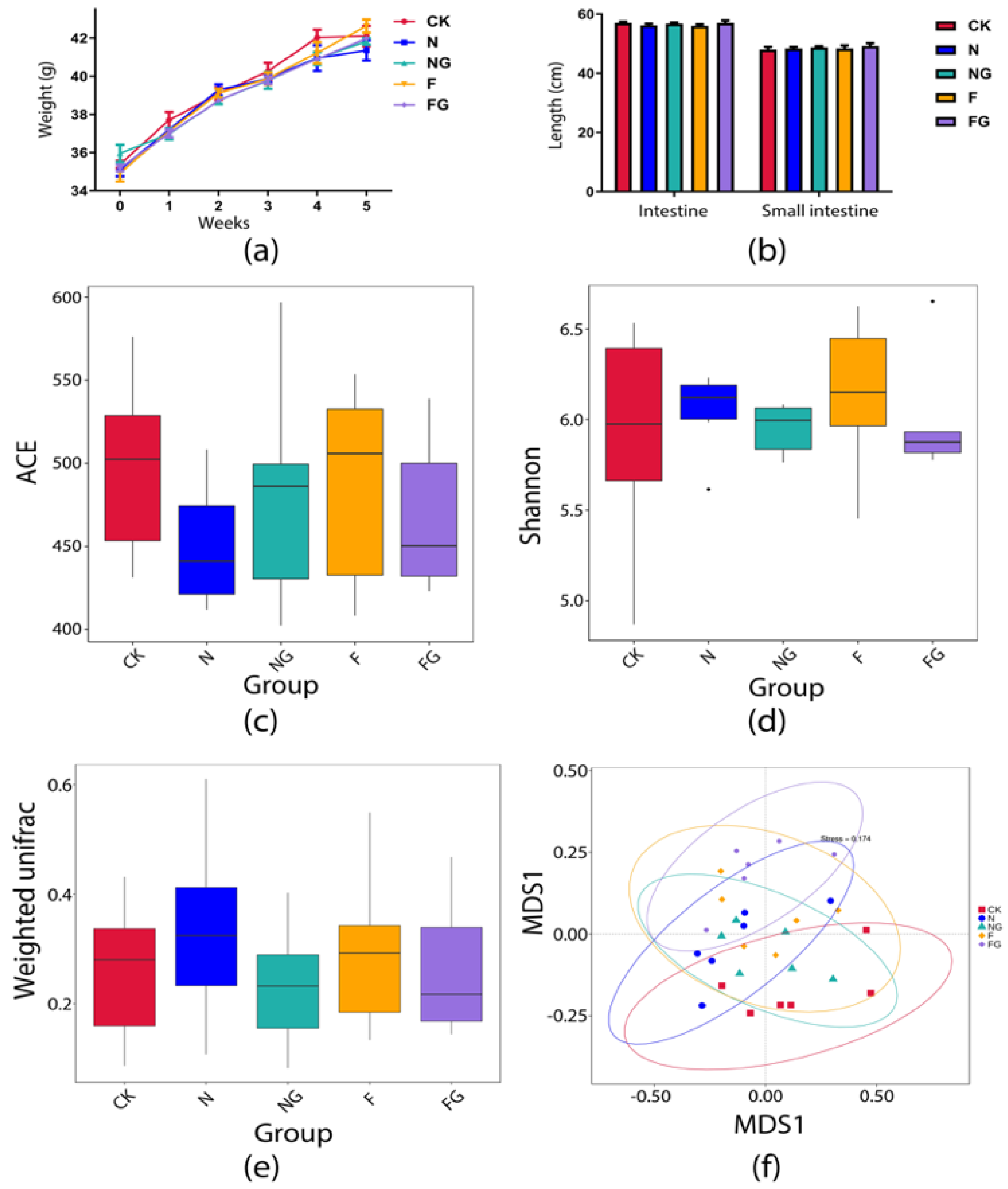
3.2. Effects of Dietary Intervention on Serum Markers, Body Weight (BW), and Intestinal Length
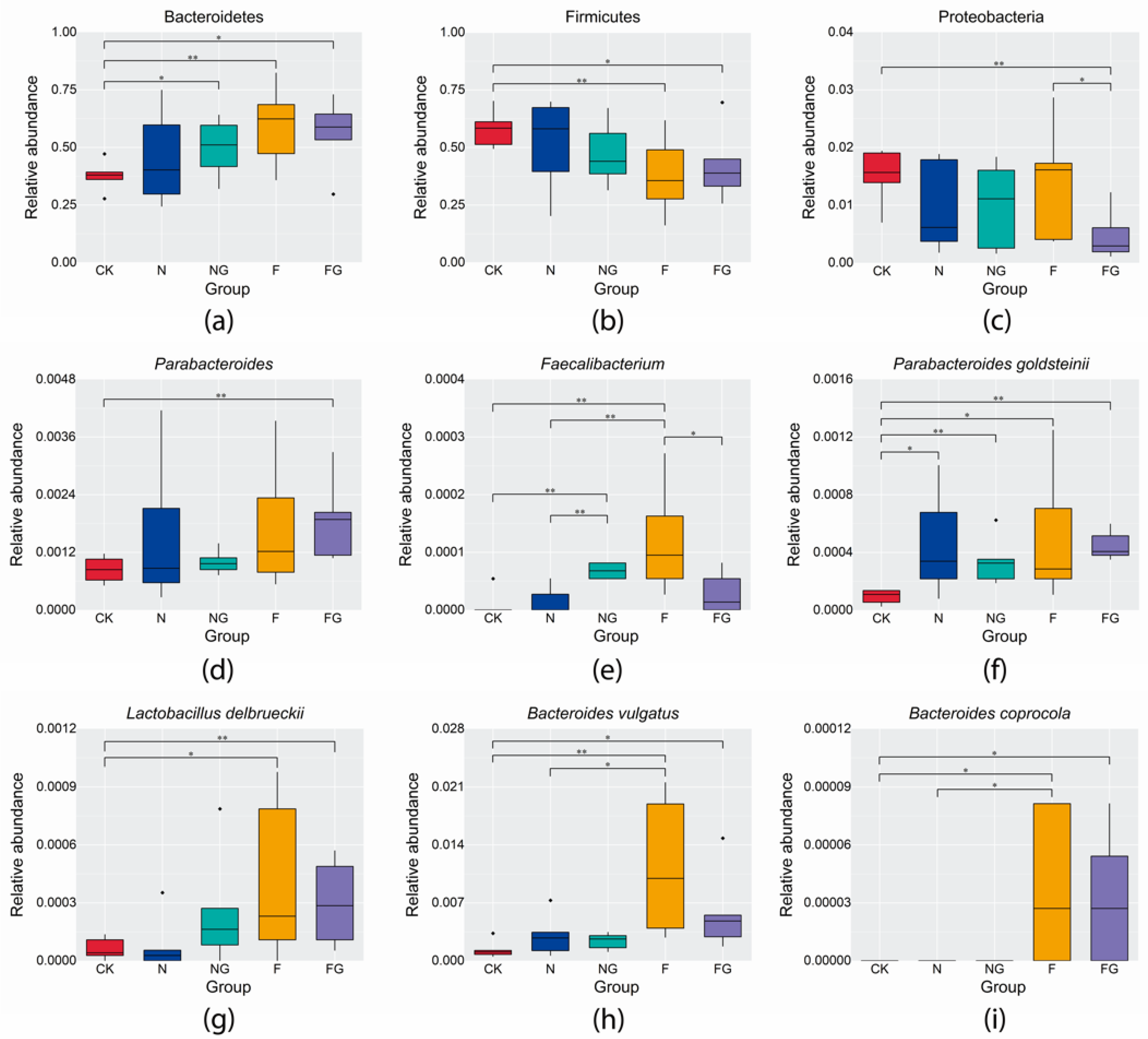
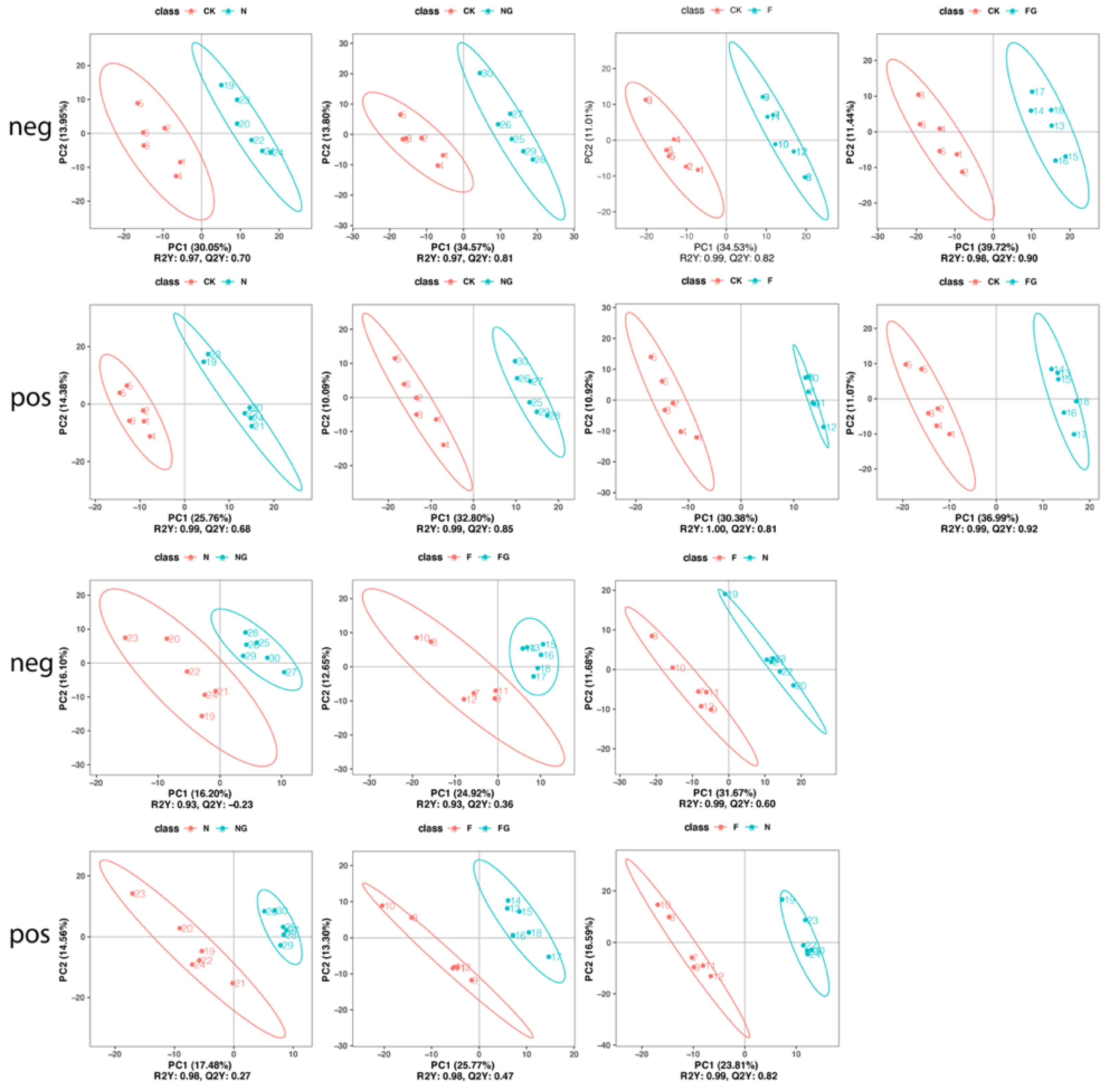
3.3. Effects of Dietary Intervention on Gut Microbiota in Mice
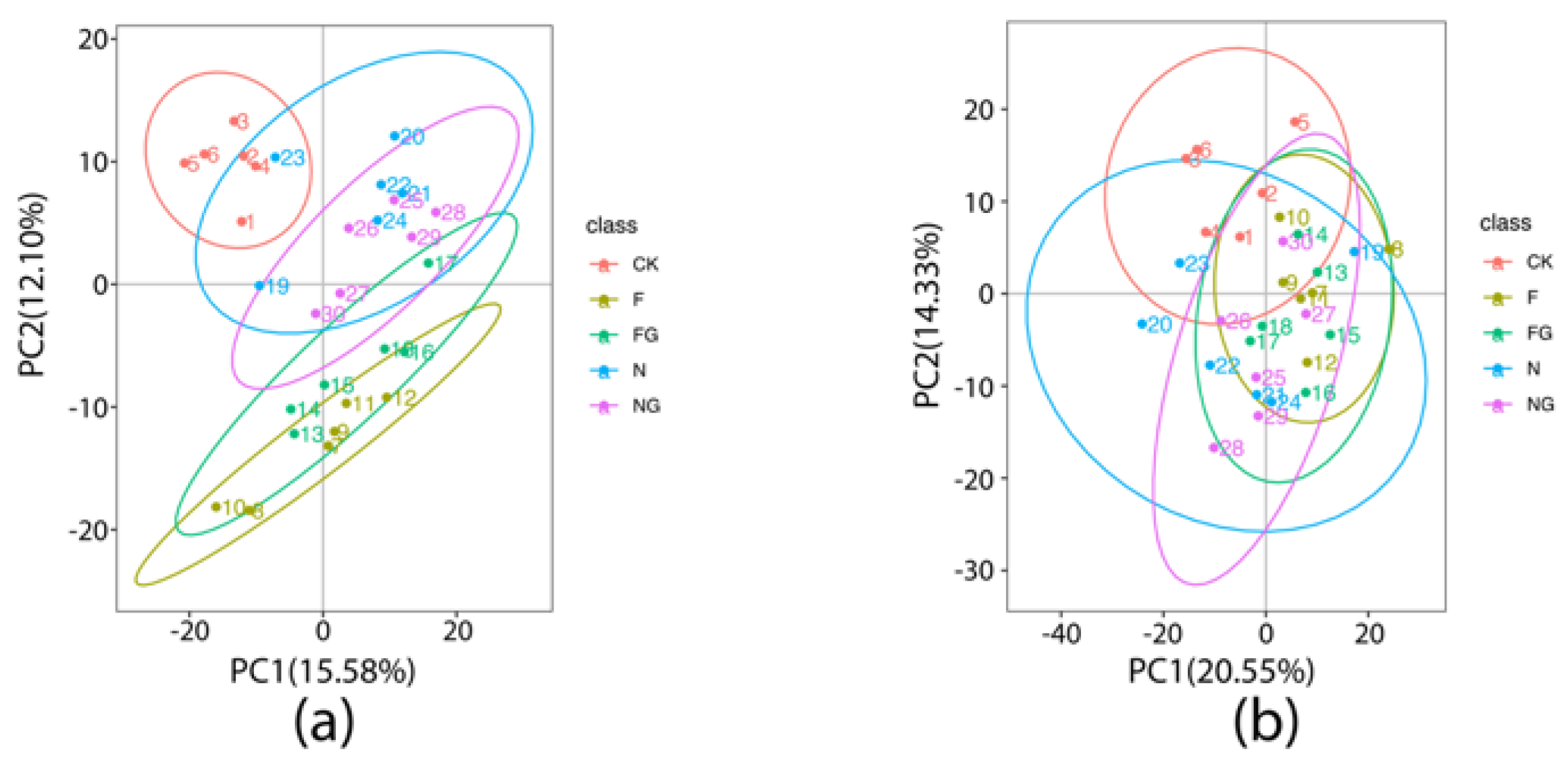
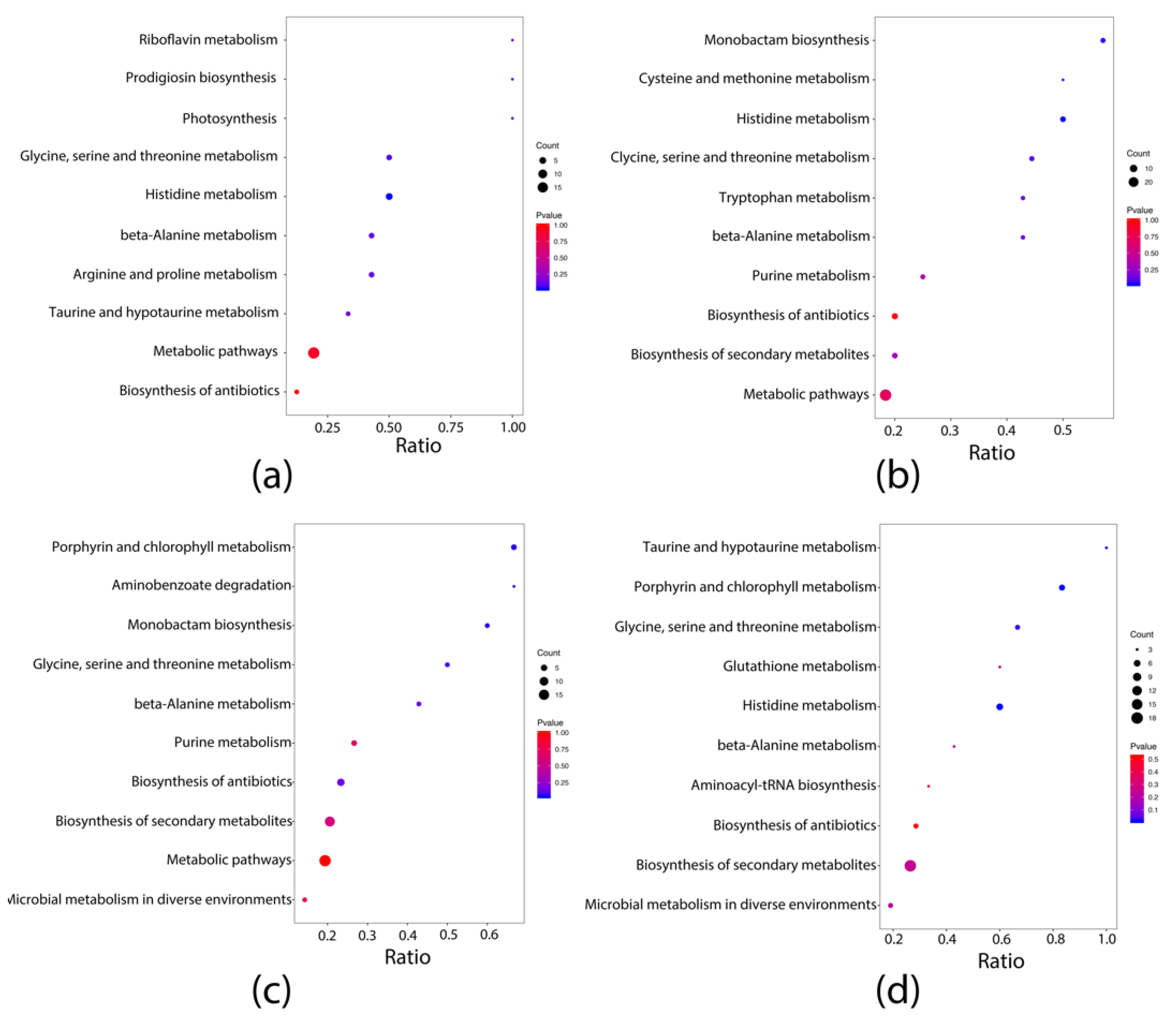
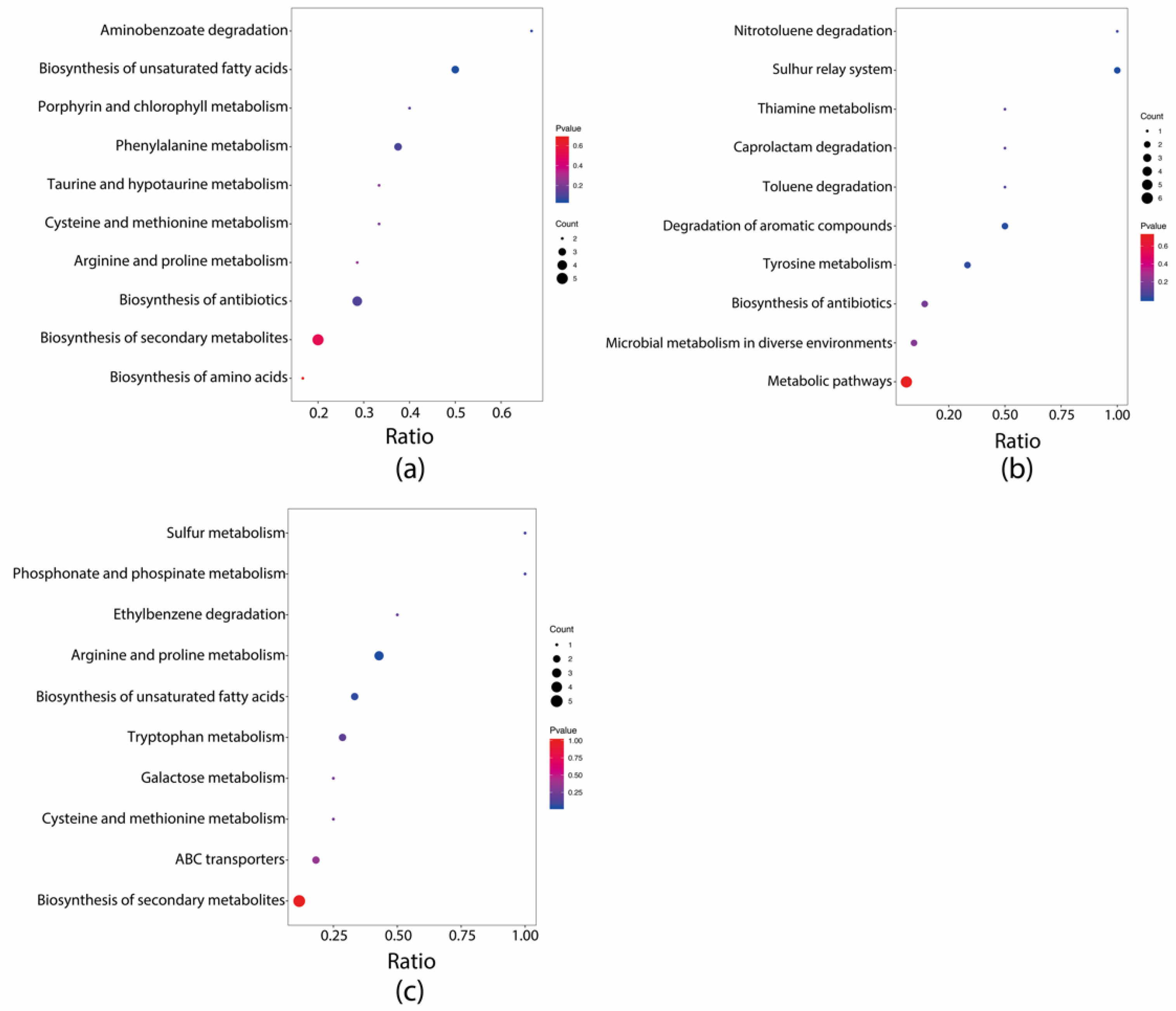
3.4. Effects of Dietary Intervention on Fecal Metabolites in Mice
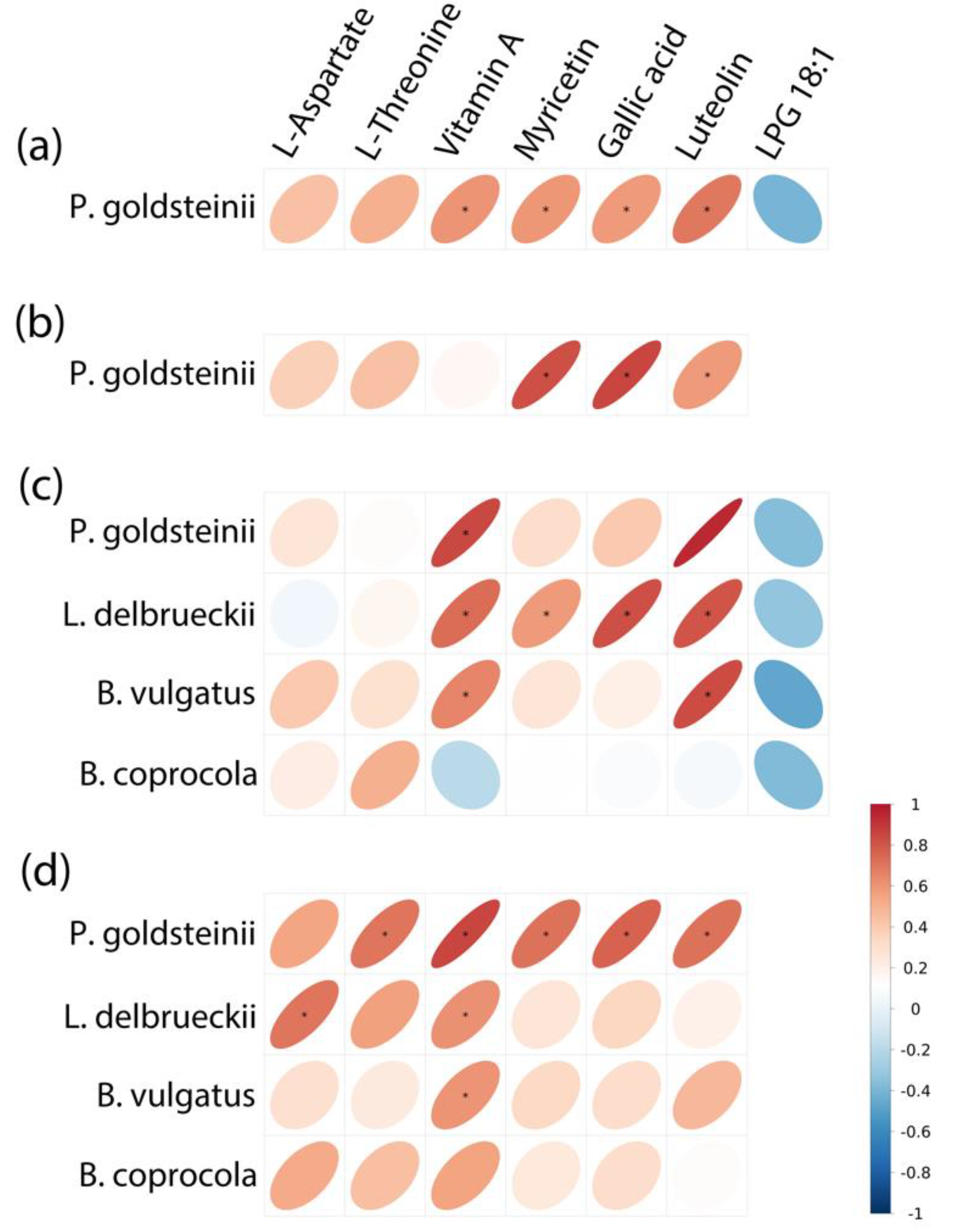
3.5. The Correlation of Gut Microbiota and Fecal Metabolites
4. Conclusions
5. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Colzi, I.; Lastrucci, L.; Rangoni, M.; Coppi, A.; Gonnelli, C. Using Myriophyllum aquaticum (Vell.) Verdc. to Remove Heavy Metals from Contaminated Water: Better Dead or Alive? J. Environ. Manag. 2018, 213, 320–328. [Google Scholar] [CrossRef] [PubMed]
- Luo, P.; Liu, F.; Zhang, S.; Li, H.; Chen, X.; Wu, L.; Jiang, Q.; Xiao, R.; Wu, J. Evaluating Organics Removal Performance from Lagoon-Pretreated Swine Wastewater in Pilot-Scale Three-Stage Surface Flow Constructed Wetlands. Chemosphere 2018, 211, 286–293. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zhang, S.; Luo, P.; Zhuang, X.; Chen, X.; Wu, J. Purification and Reuse of Non-Point Source Wastewater via Myriophyllum-Based Integrative Biotechnology: A Review. Bioresour. Technol. 2018, 248, 3–11. [Google Scholar] [CrossRef]
- Lastrucci, L.; Lazzaro, L.; Dell’Olmo, L.; Foggi, B.; Cianferoni, F. Impacts of Myriophyllum aquaticum Invasion in a Mediterranean Wetland on Plant and Macro-Arthropod Communities. Plant Biosyst. 2017, 152, 427–435. [Google Scholar] [CrossRef]
- Lastrucci, L.; Valentini, E.; Dell’Olmo, L.; Vietina, B.; Foggi, B. Hygrophilous Vegetation and Habitats of Conservation Interest in the Area of the Lake Porta (Tuscany, Central Italy). Atti Soc. Toscana Sci. Nat. Mem. Ser. B 2015, 122, 131–146. [Google Scholar]
- Gareau, M.G.; Sherman, P.M.; Walker, W.A. Probiotics and the Gut Microbiota in Intestinal Health and Disease. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 503–514. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Gao, Y.; Ma, F.; Sun, M.; Mu, G.; Tuo, Y. The Ameliorative Effect of Lactobacillus plantarum Y44 Oral Administration on Inflammation and Lipid Metabolism in Obese Mice Fed with a High Fat Diet. Food Funct. 2020, 11, 5024–5039. [Google Scholar] [CrossRef]
- Kim, J.S.; Lee, Y.H.; Kim, Y.I.; Ahmadi, F.; Oh, Y.K.; Park, J.M.; Kwak, W.S. Effect of Microbial Inoculant or Molasses on Fermentative Quality and Aerobic Stability of Sawdust-Based Spent Mushroom Substrate. Bioresour. Technol. 2016, 216, 188–195. [Google Scholar] [CrossRef]
- Ni, K.; Wang, F.; Zhu, B.; Yang, J.; Zhou, G.; Pan, Y.; Tao, Y.; Zhong, J. Effects of Lactic Acid Bacteria and Molasses Additives on the Microbial Community and Fermentation Quality of Soybean Silage. Bioresour. Technol. 2017, 238, 706–715. [Google Scholar] [CrossRef]
- Pan, T.; Xiang, H.; Diao, T.; Ma, W.; Shi, C.; Xu, Y.; Xie, Q. Effects of Probiotics and Nutrients Addition on the Microbial Community and Fermentation Quality of Peanut Hull. Bioresour. Technol. 2019, 273, 144–152. [Google Scholar] [CrossRef]
- Puntillo, M.; Gaggiotti, M.; Oteiza, J.M.; Binetti, A.; Massera, A.; Vinderola, G. Potential of Lactic Acid Bacteria Isolated From Different Forages as Silage Inoculants for Improving Fermentation Quality and Aerobic Stability. Front. Microbiol. 2020, 11, 3091. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Du, W.; Lei, K.; Wang, B.; Wang, Y.; Zhou, Y.; Li, W. Effects of Dietary Bacillus licheniformis on Gut Physical Barrier, Immunity, and Reproductive Hormones of Laying Hens. Probiotics Antimicrob. Proteins 2017, 9, 292–299. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Zeng, D.; Zhang, Y.; Ni, X.Q.; Wang, J.; Jian, P.; Zhou, Y.; Li, Y.; Yin, Z.Q.; Pan, K.C.; et al. Lactobacillus plantarum BS22 Promotes Gut Microbial Homeostasis in Broiler Chickens Exposed to Aflatoxin B1. J. Anim. Physiol. Anim. Nutr. 2018, 102, e449–e459. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Deng, L.; Zhao, M.; Tang, J.; Liu, T.; Zhang, H.; Feng, F. Probiotic-Fermented Blueberry Juice Prevents Obesity and Hyperglycemia in High Fat Diet-Fed Mice in Association with Modulating the Gut Microbiota. Food Funct. 2020, 11, 9192–9207. [Google Scholar] [CrossRef]
- Kim, H.Y.; Park, K.Y. Clinical Trials of Kimchi Intakes on the Regulation of Metabolic Parameters and Colon Health in Healthy Korean Young Adults. J. Funct. Foods 2018, 47, 325–333. [Google Scholar] [CrossRef]
- Nsenga Kumwimba, M.; Dzakpasu, M.; Li, X. Potential of Invasive Watermilfoil (Myriophyllum Spp.) to Remediate Eutrophic Waterbodies with Organic and Inorganic Pollutants. J. Environ. Manag. 2020, 270, 110919. [Google Scholar] [CrossRef]
- Rajoka, M.I.; Ahmed, S.; Hashmi, A.S.; Athar, M. Production of Microbial Biomass Protein from Mixed Substrates by Sequential Culture Fermentation of Candida utilis and Brevibacterium lactofermentum. Ann. Microbiol. 2012, 62, 1173–1179. [Google Scholar] [CrossRef]
- Wuyts, S.; van Beeck, W.; Allonsius, C.N.; van den Broek, M.F.; Lebeer, S. Applications of Plant-Based Fermented Foods and Their Microbes. Curr. Opin. Biotechnol. 2020, 61, 45–52. [Google Scholar] [CrossRef]
- Zha, M.; Li, K.; Zhang, W.; Sun, Z.; Kwok, L.Y.; Menghe, B.; Chen, Y. Untargeted Mass Spectrometry-Based Metabolomics Approach Unveils Molecular Changes in Milk Fermented by Lactobacillus plantarum P9. LWT Food Sci. Technol. 2021, 140, 110759. [Google Scholar] [CrossRef]
- Wen, J.J.; Li, M.Z.; Gao, H.; Hu, J.L.; Nie, Q.X.; Chen, H.H.; Zhang, Y.L.; Xie, M.Y.; Nie, S.P. Polysaccharides from Fermented Momordica charantia L. with Lactobacillus plantarum NCU116 Ameliorate Metabolic Disorders and Gut Microbiota Change in Obese Rats. Food Funct. 2021, 12, 2617–2630. [Google Scholar] [CrossRef]
- Guedes, C.M.; Gonçalves, D.; Rodrigues, M.A.M.; Dias-da-Silva, A. Effects of a Saccharomyces Cerevisiae Yeast on Ruminal Fermentation and Fibre Degradation of Maize Silages in Cows. Anim. Feed Sci. Technol. 2008, 145, 27–40. [Google Scholar] [CrossRef]
- Marcó, A.; Rubio, R.; Compañó, R.; Casals, I. Comparison of the Kjeldahl Method and a Combustion Method for Total Nitrogen Determination in Animal Feed. Talanta 2002, 57, 1019–1026. [Google Scholar] [CrossRef]
- Manirakiza, P.; Covaci, A.; Schepens, P. Comparative Study on Total Lipid Determination Using Soxhlet, Roese-Gottlieb, Bligh & Dyer, and Modified Bligh & Dyer Extraction Methods. J. Food Compos. Anal. 2001, 14, 93–100. [Google Scholar]
- Rodríguez, R.; Jiménez, A.; Fernández-Bolaños, J.; Guillén, R.; Heredia, A. Dietary Fibre from Vegetable Products as Source of Functional Ingredients. Trends Food Sci. Technol. 2006, 17, 3–15. [Google Scholar] [CrossRef]
- Fuentes-Soriano, P.; Bellido-Milla, D.; García-Guzmán, J.J.; Hernández-Artiga, M.P.; Gallardo-Bernal, J.J.; Palacios-Santander, J.M.; Espada-Bellido, E. A Simple Phosphorus Determination in Walnuts and Assessment of the Assimilable Fraction. Talanta 2019, 204, 57–62. [Google Scholar] [CrossRef]
- Chahdoura, H.; Morales, P.; Barreira, J.C.M.; Barros, L.; Fernández-Ruiz, V.; Ferreira, I.C.F.R.; Achour, L. Dietary Fiber, Mineral Elements Profile and Macronutrients Composition in Different Edible Parts of Opuntia microdasys (Lehm.) Pfeiff and Opuntia macrorhiza (Engelm.). LWT Food Sci. Technol. 2015, 64, 446–451. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly Accurate OTU Sequences from Microbial Amplicon Reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA Ribosomal RNA Gene Database Project: Improved Data Processing and Web-Based Tools. Nucleic Acids Res. 2013, 41, 590–596. [Google Scholar] [CrossRef]
- Cao, Y.; Liu, Y.; Dong, Q.; Wang, T.; Niu, C. Alterations in the Gut Microbiome and Metabolic Profile in Rats Acclimated to High Environmental Temperature. Microb. Biotechnol. 2022, 15, 276–288. [Google Scholar] [CrossRef]
- Wang, J.; Cao, F.; Su, E.; Zhao, L.; Qin, W. Improvement of Animal Feed Additives of Ginkgo Leaves through Solid-State Fermentation Using Aspergillus niger. Int. J. Biol. Sci. 2018, 14, 736. [Google Scholar] [CrossRef] [Green Version]
- Shin, N.R.; Whon, T.W.; Bae, J.W. Proteobacteria: Microbial Signature of Dysbiosis in Gut Microbiota. Trends Biotechnol. 2015, 33, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Zhang, M.; Wang, Y.; Dorfman, R.G.; Liu, H.; Yu, T.; Chen, X.; Tang, D.; Xu, L.; Yin, Y.; et al. Faecalibacterium prausnitzii Produces Butyrate to Maintain Th17/Treg Balance and to Ameliorate Colorectal Colitis by Inhibiting Histone Deacetylase 1. Inflamm. Bowel Dis. 2018, 24, 1926–1940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staley, C.; Kaiser, T.; Vaughn, B.P.; Graiziger, C.; Hamilton, M.J.; Kabage, A.J.; Khoruts, A.; Sadowsky, M.J. Durable Long-Term Bacterial Engraftment Following Encapsulated Fecal Microbiota Transplantation to Treat Clostridium difficile Infection. mBio 2019, 10, e01586-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, T.R.; Lin, C.S.; Chang, C.J.; Lin, T.L.; Martel, J.; Ko, Y.F.; Ojcius, D.M.; Lu, C.C.; Young, J.D.; Lai, H.C. Gut Commensal Parabacteroides goldsteinii Plays a Predominant Role in the Anti-Obesity Effects of Polysaccharides Isolated from Hirsutella sinensis. Gut 2019, 68, 248–262. [Google Scholar] [CrossRef] [PubMed]
- Elean, M.; Albarracín, L.; Cataldo, P.G.; Londero, A.; Kitazawa, H.; Saavedra, L.; Villena, J.; Hebert, E.M. New Immunobiotics from Highly Proteolytic Lactobacillus delbrueckii Strains: Their Impact on Intestinal Antiviral Innate Immune Response. Benef. Microbes 2020, 11, 375–390. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.Y.; Joung, H.C.; Kim, B.K.; Kim, B.Y.; Park, T.S.; Suk, K.T. Lactobacillus lactis CKDB001 Ameliorate Progression of Nonalcoholic Fatty Liver Disease through of Gut Microbiome: Addendum. Gut Microbes 2020, 12, 1829449. [Google Scholar] [CrossRef]
- Yoshida, N.; Emoto, T.; Yamashita, T.; Watanabe, H.; Hayashi, T.; Tabata, T.; Hoshi, N.; Hatano, N.; Ozawa, G.; Sasaki, N.; et al. Bacteroides vulgatus and Bacteroides dorei Reduce Gut Microbial Lipopolysaccharide Production and Inhibit Atherosclerosis. Circulation 2018, 138, 2486–2498. [Google Scholar] [CrossRef]
- Makki, K.; Deehan, E.C.; Walter, J.; Bäckhed, F. The Impact of Dietary Fiber on Gut Microbiota in Host Health and Disease. Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Jiang, T.; Gao, X.; Wu, C.; Tian, F.; Lei, Q.; Bi, J.; Xie, B.; Wang, H.Y.; Chen, S.; Wang, X. Apple-Derived Pectin Modulates Gut Microbiota, Improves Gut Barrier Function, and Attenuates Metabolic Endotoxemia in Rats with Diet-Induced Obesity. Nutrients 2016, 8, 126. [Google Scholar] [CrossRef] [Green Version]
- Wexler, H.M. Bacteroides: The Good, the Bad, and the Nitty-Gritty. Clin. Microbiol. Rev. 2007, 20, 593–621. [Google Scholar] [CrossRef] [Green Version]
- Kitahara, M.; Sakamoto, M.; Ike, M.; Sakata, S.; Benno, Y. Bacteroides plebeius Sp. Nov. and Bacteroides coprocola Sp. Nov., Isolated from Human Faeces. Int. J. Syst. Evol. Microbiol. 2005, 55, 2143–2147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Ji, H.; Wang, S.; Liu, H.; Zhang, W.; Zhang, D.; Wang, Y. Probiotic Lactobacillus plantarum Promotes Intestinal Barrier Function by Strengthening the Epithelium and Modulating Gut Microbiota. Front. Microbiol. 2018, 9, 1953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macfarlane, G.T.; Cummings, J.H.; Allison, C. Protein Degradation by Human Intestinal Bacteria. Microbiology 1986, 132, 1647–1656. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, P.; Liu, Y.; Zhu, H.; Li, S.; Shi, H.; Chen, F.; Leng, W.; Pi, D.; Hou, Y.; Yi, D. The Effect of Aspartate on the Energy Metabolism in the Liver of Weanling Pigs Challenged with Lipopolysaccharide. Eur. J. Nutr. 2014, 54, 581–588. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Wang, X.; Zhang, T.; Si, H.; Nan, W.; Xu, C.; Guan, L.; Wright, A.D.G.; Li, G. The Development of Microbiota and Metabolome in Small Intestine of Sika Deer (Cervus nippon) from Birth to Weaning. Front. Microbiol. 2018, 9, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, S.; Li, Q.; Hu, K.; He, Y.; Ai, Q.; Hu, L.; Yu, J. Vitamin A and Retinoic Acid Exhibit Protective Effects on Necrotizing Enterocolitis by Regulating Intestinal Flora and Enhancing the Intestinal Epithelial Barrier. Arch. Med. Res. 2018, 49, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Tan, L.; Wang, M.; Ren, C.; Guo, C.; Yang, B.; Ren, Y.; Cao, Z.; Li, Y.; Pei, J. Myricetin: A Review of the Most Recent Research. Biomed. Pharmacother. 2021, 134, 111017. [Google Scholar] [CrossRef]
- Li, D.; Lv, B.; Wang, D.; Xu, D.; Qin, S.; Zhang, Y.; Chen, J.; Zhang, W.; Zhang, Z.; Xu, F. Network Pharmacology and Bioactive Equivalence Assessment Integrated Strategy Driven Q-Markers Discovery for Da-Cheng-Qi Decoction to Attenuate Intestinal Obstruction. Phytomedicine 2020, 72, 153236. [Google Scholar] [CrossRef]
- Wilsher, N.E.; Arroo, R.R.; Matsoukas, M.T.; Tsatsakis, A.M.; Spandidos, D.A.; Androutsopoulos, V.P. Cytochrome P450 CYP1 Metabolism of Hydroxylated Flavones and Flavonols: Selective Bioactivation of Luteolin in Breast Cancer Cells. Food Chem. Toxicol. 2017, 110, 383–394. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Zhang, G.; Pan, J.; Wang, Y. α-Glucosidase Inhibition by Luteolin: Kinetics, Interaction and Molecular Docking. Int. J. Biol. Macromol. 2014, 64, 213–223. [Google Scholar] [CrossRef]
- Shen, J.; Li, P.; Liu, S.; Liu, Q.; Li, Y.; Zhang, Z.; Yang, C.; Hu, M.; Sun, Y.; He, C.; et al. The Chemopreventive Effects of Huangqin-Tea against AOM-Induced Preneoplastic Colonic Aberrant Crypt Foci in Rats and Omics Analysis. Food Funct. 2020, 11, 9634–9650. [Google Scholar] [CrossRef] [PubMed]
- Ye, G.; Yang, B.C.; Gao, H.; Wu, Z.; Chen, J.; Ai, X.Y.; Huang, Q. Metabolomics Insights into Oleate-Induced Disorders of Phospholipid Metabolism in Macrophages. J. Nutr. 2021, 151, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Shama, S.; Liu, W. Omega-3 Fatty Acids and Gut Microbiota: A Reciprocal Interaction in Nonalcoholic Fatty Liver Disease. Dig. Dis. Sci. 2020, 65, 906–910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Chen, Y.; Wang, Y.; Li, Y.; Zhang, X.; Zheng, H.; Ma, F.; Ma, C.W.; Lu, B.; Xie, Z.; et al. Beneficial Changes of Gut Microbiota and Metabolism in Weaned Rats with Lactobacillus acidophilus NCFM and Bifidobacterium lactis Bi-07 Supplementation. J. Funct. Foods 2018, 48, 252–265. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fermentation Conditions | pH | Lactic Acid (μg/mL) | Acetic Acid (μg/mL) | Propionic Acid (μg/mL) | Dry Matter (g/30 g) | Crude Protein (%) | NH3-N/N (%) | Score | |
|---|---|---|---|---|---|---|---|---|---|
| 0 d | 30 °C | 5.56 ± 0.04 a | 0.11 ± 0.01 b | 43.7 ± 2.78 e | 1.84 ± 0.12 b | 8.44 ± 0.15 ab | 21.6 ± 0.38 ab | 3.30 ± 0.17 c | 00.0 |
| 35 °C | 5.56 ± 0.04 a | 0.11 ± 0.01 b | 43.7 ± 2.78 e | 1.84 ± 0.12 b | 8.44 ± 0.15 ab | 21.6 ± 0.38 ab | 3.30 ± 0.17 c | 00.0 | |
| 1 d | 30 °C | 5.05 ± 0.02 b | 0.42 ± 0.05 b | 61.5 ± 9.70 cde | 2.06 ± 0.17 ab | 8.53 ± 0.12 ab | 21.8 ± 0.51 ab | 4.56 ± 0.32 bc | 40.0 |
| 35 °C | 5.08 ± 0.03 b | 0.42 ± 0.01 b | 51.9 ± 2.06 de | 2.20 ± 0.31 ab | 9.71 ± 0.94 a | 22.2 ± 0.90 ab | 4.68 ± 0.56 bc | 53.0 | |
| 3 d | 30 °C | 4.34 ± 0.03 d | 1.09 ±0.35 a | 85.4 ± 23.7 bcd | 2.53 ± 0.87 ab | 8.26 ± 0.49 b | 21.0 ± 0.40 ab | 5.38 ± 0.12 ab | 56.9 |
| 35 °C | 4.26 ± 0.03 d | 1.13 ± 0.10 a | 92.0 ± 17.4 bc | 2.31 ± 0.13 ab | 8.48 ± 0.34 ab | 21.8 ± 0.90 ab | 5.69 ± 0.61 ab | 59.1 | |
| 5 d | 30 °C | 4.11 ± 0.03 e | 1.11 ± 0.10 a | 110 ± 16.8 ab | 2.37 ± 0.16 ab | 9.18 ± 0.52 ab | 22.3 ± 0.40 a | 5.60 ± 0.37 ab | 76.4 |
| 35 °C | 4.22 ± 0.03 d | 1.16 ± 0.10 a | 94.8 ± 6.20 bc | 2.44 ± 0.47 ab | 8.40 ± 0.25 ab | 20.0 ± 0.75 b | 6.70 ± 0.33 a | 55.3 | |
| 7 d | 30 °C | 4.43 ± 0.03 c | 1.24 ± 0.10 a | 114 ± 17.1 ab | 2.57 ± 0.35 ab | 8.70 ± 0.33 ab | 22.3 ± 0.78 a | 5.53 ± 1.41 ab | 73.0 |
| 35 °C | 4.42 ± 0.04 c | 1.22 ± 0.10 a | 117 ± 12.2 ab | 2.98 ± 0.48 a | 8.44 ± 0.34 ab | 21.3 ± 0.97 ab | 6.96 ± 0.52 a | 60.3 | |
| 10 d | 30 °C | 4.45 ± 0.03 c | 1.08 ± 0.04 a | 135 ± 14.0 a | 2.82 ± 0.30 ab | 8.73 ± 0.78 ab | 21.5 ± 0.42 ab | 6.90 ± 0.07 a | 60.4 |
| 35 °C | 4.46 ± 0.03 c | 1.24 ± 0.04 a | 134 ± 12.0 a | 2.52 ± 0.23 ab | 8.99 ± 0.60 ab | 21.2 ± 1.22 ab | 6.91 ± 1.14 a | 62.3 | |
| Analysis | NFM | FM | Trends | p-Value |
|---|---|---|---|---|
| Values Shown Per 100 g | ||||
| Crude protein | 18.1 ± 1.20 g | 20.5 ± 0.80 g | ↑ 13.3% | 0.045 * |
| Lipids | 3.00 ± 0.14 g | 4.60 ± 0.22 g | ↑ 53.3% | 0.000 ** |
| Ash | 12.4 ± 0.90 g | 15.9 ± 0.72 g | ↑ 28.2% | 0.006 ** |
| Raw Fiber | 13.2 ± 0.80% | 11.2 ± 0.70% | ↓ 15.2% | 0.031 * |
| Phosphorus | 653 ± 6.00 mg | 657 ± 4.00 mg | ↑ <0.01% | 0.391 |
| Calcium | 2.07 ± 0.03 g | 2.09 ± 0.04 g | ↑ <0.01% | 0.510 |
| No. | Metabolites | Group N-CK | Group NG-CK | Group F-CK | Group FG-CK | Group F-N | Group NG-N | Group FG-F | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| FC | Sig. | FC | Sig. | FC | Sig. | FC | Sig. | FC | Sig. | FC | Sig. | FC | Sig. | ||
| 1 | L-Aspartate | 2.01 | 0.04 | 2.10 | 0.04 | 2.06 | 0.04 | 2.09 | 0.03 | 1.02 | ns | 1.04 | ns | 1.02 | ns |
| 2 | L-Threonine | 2.45 | 0.01 | 1.96 | 0.03 | 2.28 | 0.02 | 2.74 | 0.01 | 0.93 | ns | 0.80 | ns | 0.83 | ns |
| 3 | Vitamin A | 3.50 | <0.01 | 4.53 | <0.01 | 86.4 | <0.01 | 10.35 | <0.01 | 24.7 | 0.03 | 1.30 | ns | 8.35 | ns |
| 4 | Myricetin | 3.43 | 0.01 | 6.39 | <0.01 | 73.2 | <0.01 | 97.5 | <0.01 | 21.3 | <0.01 | 1.86 | ns | 0.75 | ns |
| 5 | Gallic acid | 3.23 | <0.01 | 5.56 | <0.01 | 16.7 | <0.01 | 9.09 | <0.01 | 5.16 | 0.03 | 1.72 | ns | 0.54 | ns |
| 6 | Luteolin | 4.21 | <0.01 | 5.03 | <0.01 | 9.49 | <0.01 | 8.70 | <0.01 | 2.26 | 0.03 | 1.19 | ns | 1.09 | ns |
| 7 | LPG 18:1 | 0.31 | 0.02 | 0.52 | ns | 0.18 | <0.01 | 0.81 | ns | 0.60 | ns | 1.70 | 0.02 | 4.46 | <0.01 |
| 8 | 9-oxo-ODA | 1.01 | ns | 0.70 | ns | 0.72 | ns | 1.12 | ns | 0.71 | ns | 0.69 | ns | 1.56 | 0.049 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Ling, Y.; Liu, J.; Zhang, M.; Li, Z.; Bai, Z.; Wu, Z.; Xia, R.; Wu, Z.; Wan, Y.; et al. Fermented Myriophyllum aquaticum and Lactobacillus plantarum Affect the Distribution of Intestinal Microbial Communities and Metabolic Profile in Mice. Fermentation 2022, 8, 210. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8050210
Li Y, Ling Y, Liu J, Zhang M, Li Z, Bai Z, Wu Z, Xia R, Wu Z, Wan Y, et al. Fermented Myriophyllum aquaticum and Lactobacillus plantarum Affect the Distribution of Intestinal Microbial Communities and Metabolic Profile in Mice. Fermentation. 2022; 8(5):210. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8050210
Chicago/Turabian StyleLi, Yueyang, Yuxi Ling, Jia Liu, Michael Zhang, Zuming Li, Zhihui Bai, Zhenlong Wu, Ran Xia, Zhichao Wu, Yingxin Wan, and et al. 2022. "Fermented Myriophyllum aquaticum and Lactobacillus plantarum Affect the Distribution of Intestinal Microbial Communities and Metabolic Profile in Mice" Fermentation 8, no. 5: 210. https://0-doi-org.brum.beds.ac.uk/10.3390/fermentation8050210