Morphology, Transcriptomics and In Vitro Model of Skin from Polar Cod (Boreogadus Saida) and Atlantic Cod (Gadus Morhua)

 , , and
, , and
Abstract
:1. Introduction
2. Results
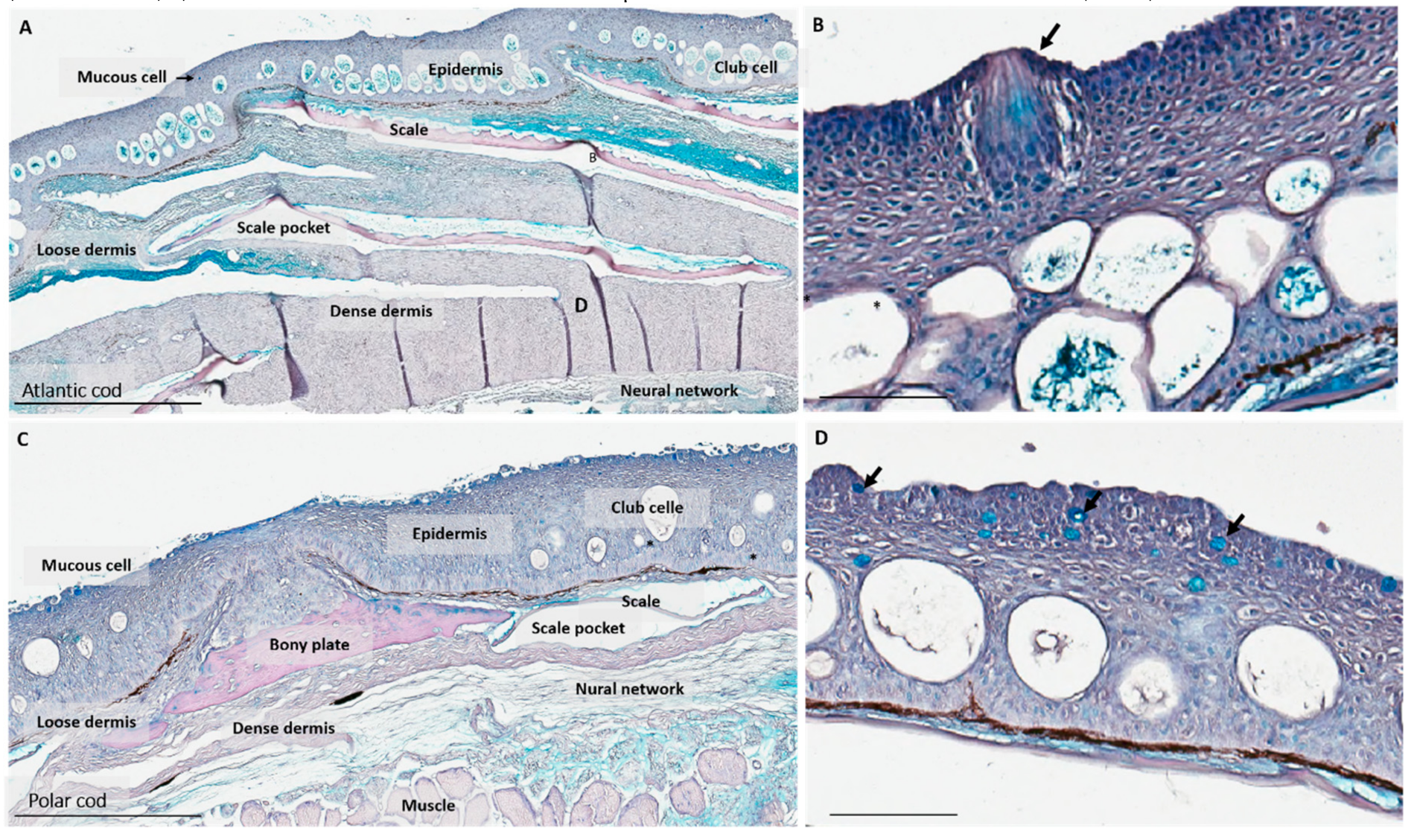
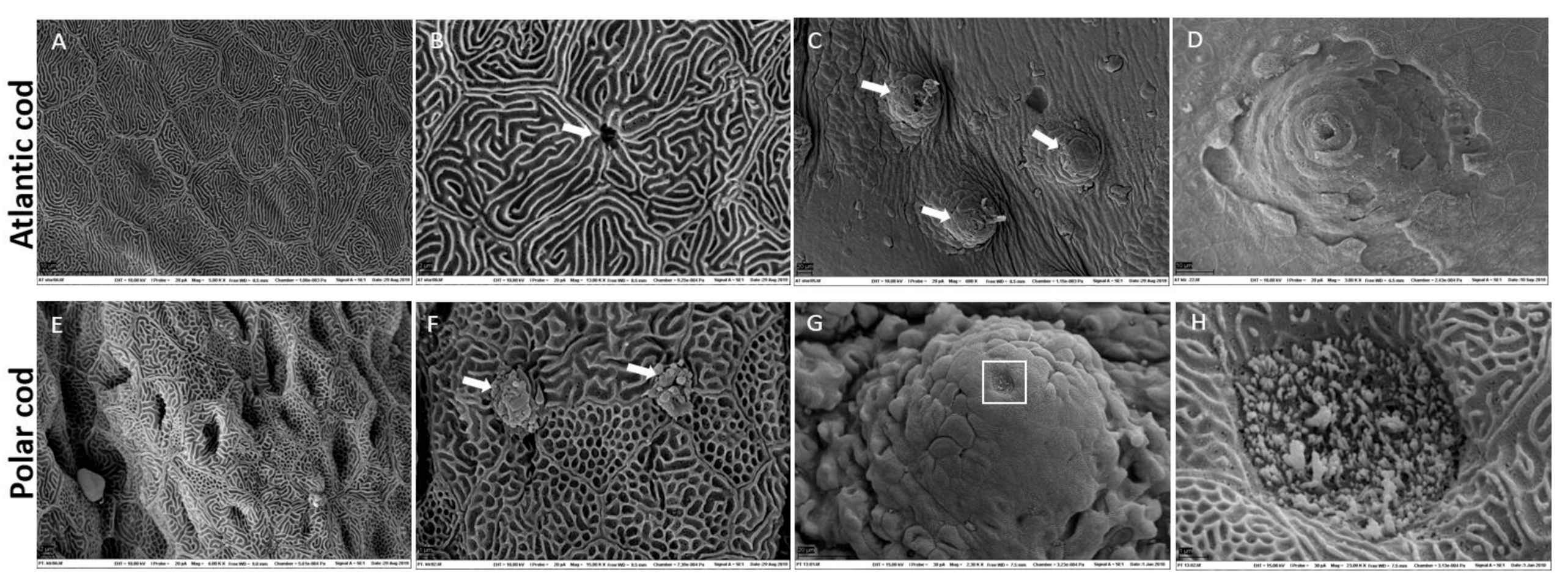
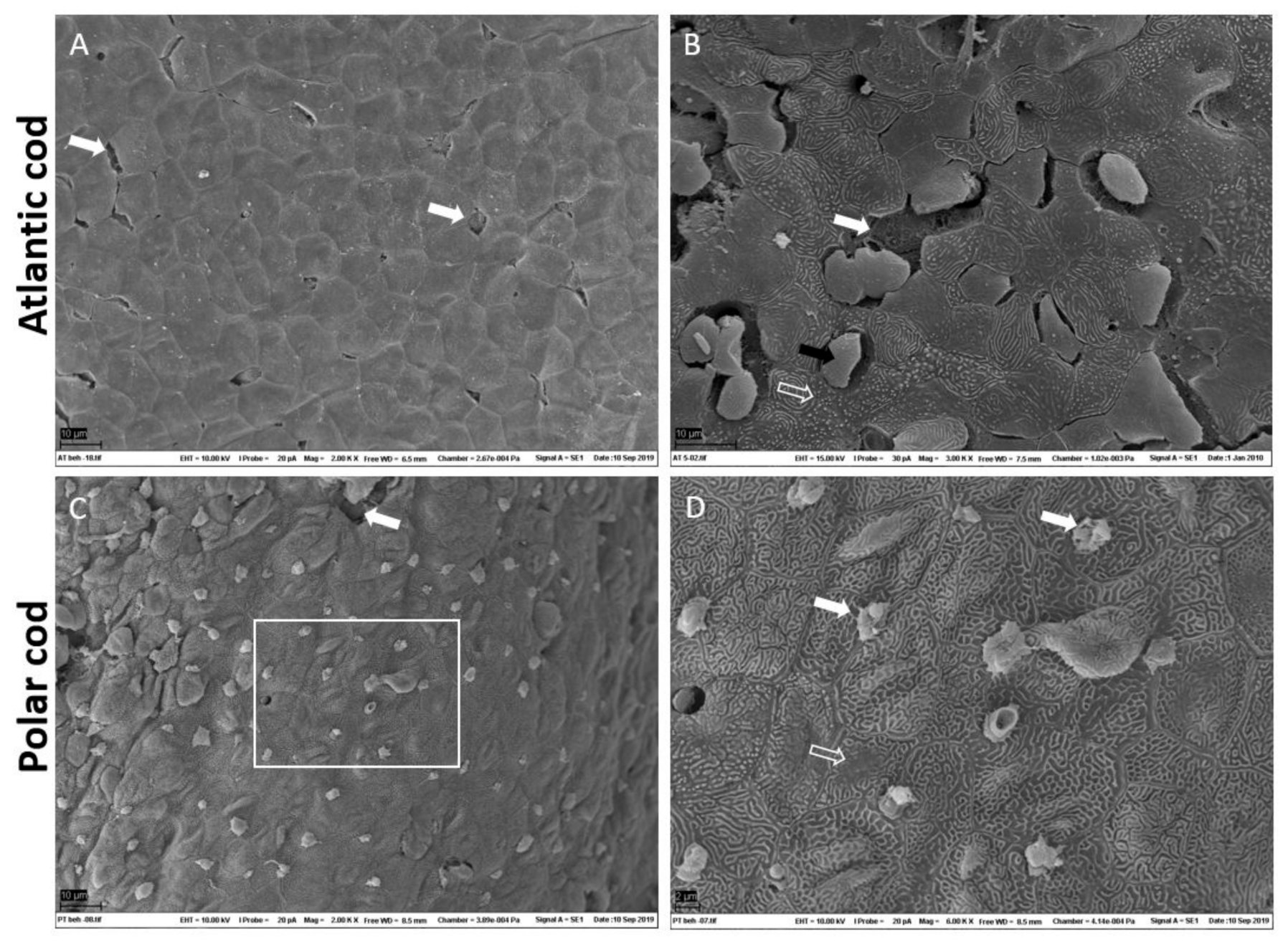
2.1. Characterization of Skin
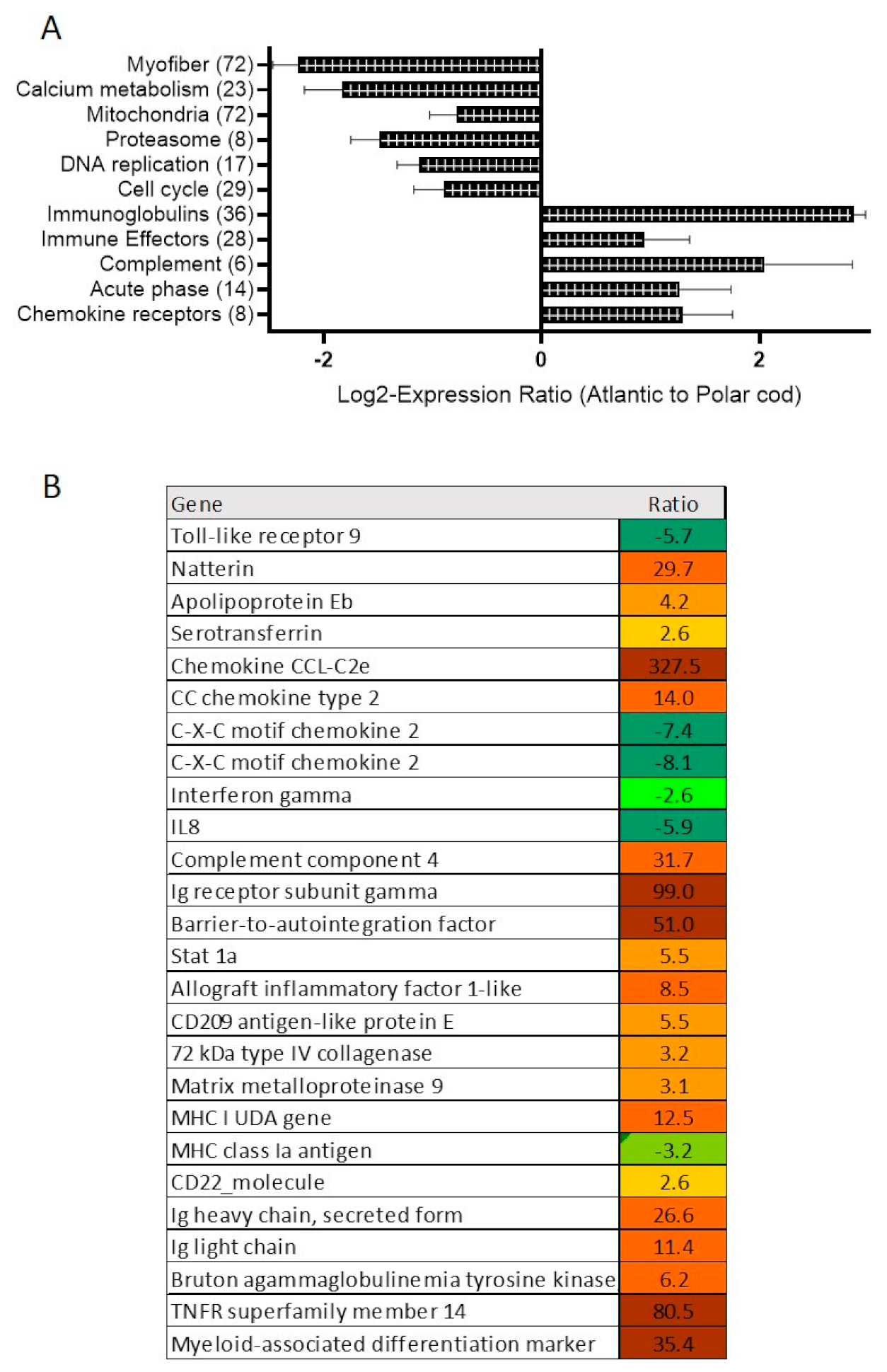
2.2. Skin Transcriptome
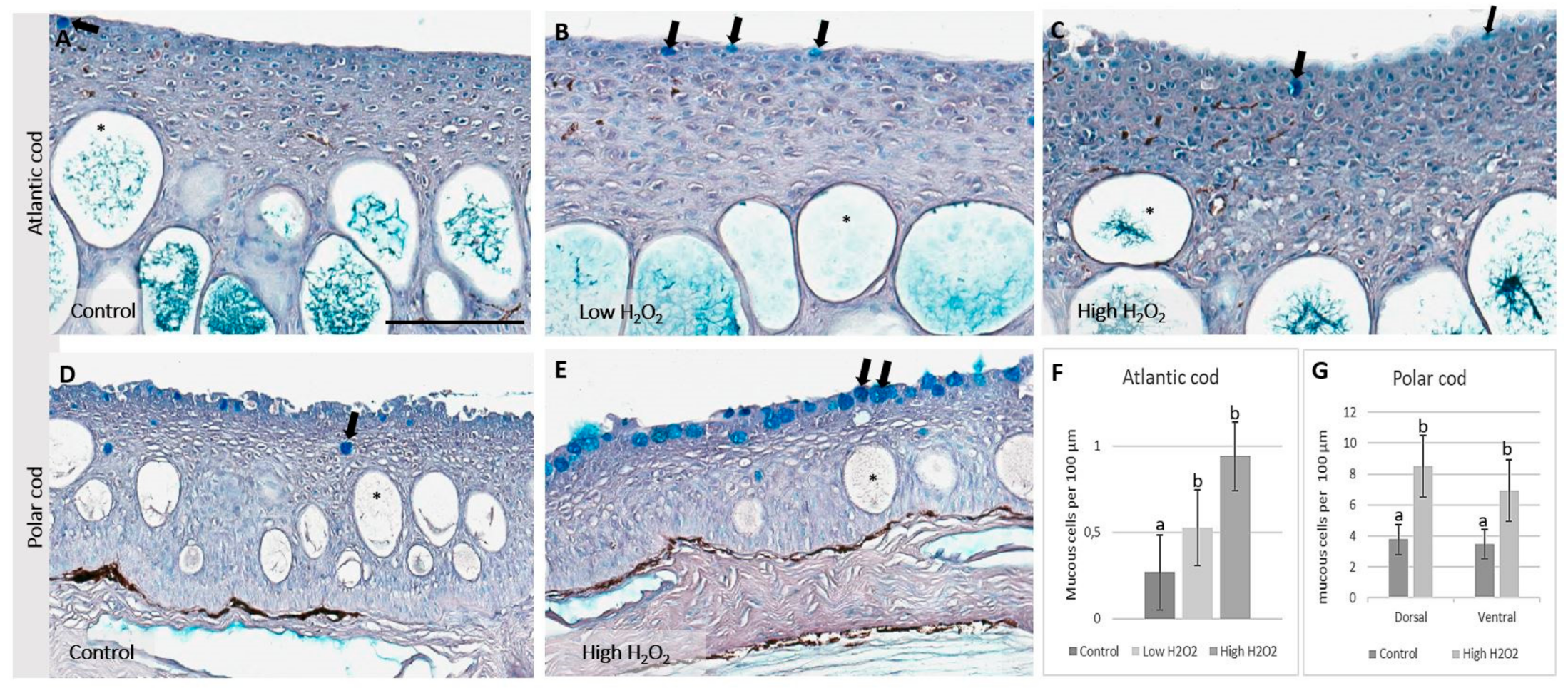
2.3. Effects of Hydrogen Peroxide Exposure on Skin
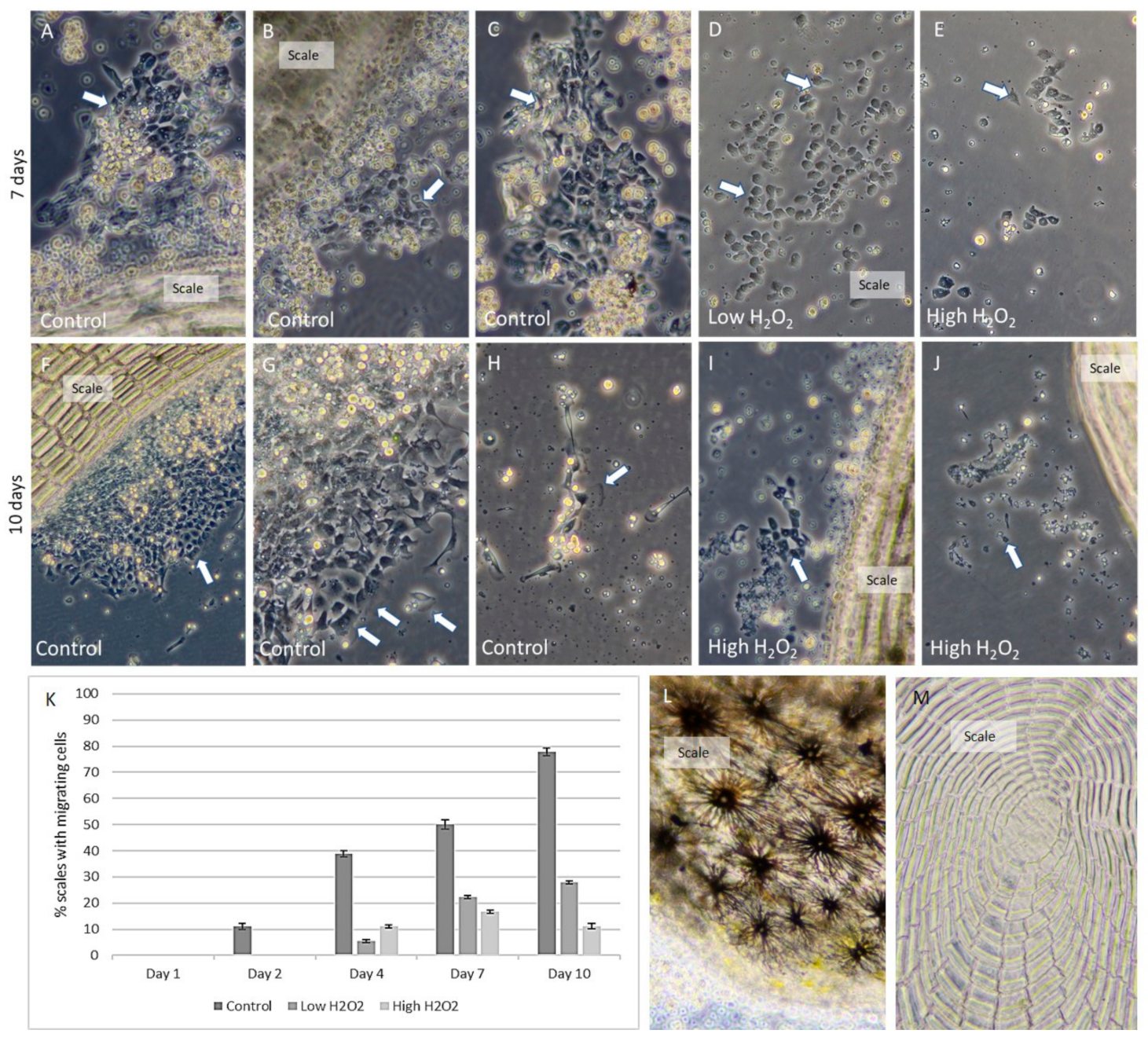
2.4. Primary Cell Culture of Atlantic Cod Keratocytes
3. Discussion
3.1. Polar Cod and Atlantic Skin Differ in Skin Micro-Architecture
3.2. Exposure to Hydrogen Peroxide
3.3. Keratocyte Cod Cell Culture is a Promising In Vitro Model
4. Materials and Methods
4.1. Experimental Setup and Sampling
4.2. Histological Staining
4.3. Scanning Electron Microscopy (SEM)
4.4. RNA Extraction
4.5. Microarray
4.6. In Vitro Primary Keratocyte Cell Culture
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hop, H.; Gjøsæter, H. Polar cod (Boreogadus saida) and capelin (Mallotus villosus) as key species in marine food webs of the Arctic and the Barents Sea. Mar. Biol. Res. 2013, 9, 878–894. [Google Scholar] [CrossRef]
- Welch, H.E.; Bergmann, M.A.; Siferd, T.D.; Martin, K.A.; Curtis, M.F.; Crawford, R.E.; Conover, R.J.; Hop, H. Energy-flow through the marine ecosystem of the Lancaster Sound Region, Arctic Canada. Arctic 1992, 45, 343–357. [Google Scholar] [CrossRef] [Green Version]
- Misund, O.A.; Heggland, K.; Skogseth, R.; Falck, E.; Gjøsæter, H.; Sundet, J.; Watne, J.; Lønne, O.J. Norwegian fisheries in the Svalbard zone since 1980. Regulations, profitability and warming waters affect landings. Polar Sci. 2016, 10, 312–322. [Google Scholar] [CrossRef] [Green Version]
- Carroll, J.; Vikebø, F.; Howell, D.; Broch, O.J.; Nepstad, R.; Augustine, S.; Skeie, G.M.; Bast, R.; Juselius, J. Assessing impacts of simulated oil spills on the Northeast Arctic cod fishery. Mar. Pollut. Bull. 2018, 126, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, E.; Hansen, H.M.; Øyvind, J.; Mortensen, A. Kunnskaps-Og Erfaringsgrunnlag for Torskeoppdrett; Research Report; Nofima 23: Tromsø, Norway, 2018. [Google Scholar]
- David, C.; Lange, B.; Krumpen, T.; Schaafsma, F.; van Franeker, J.A.; Flores, H. Under-ice distribution of polar cod Boreogadus saida in the central Arctic Ocean and their association with sea-ice habitat properties. Polar Biol. 2016, 39, 981–994. [Google Scholar] [CrossRef]
- Lønne, O.J.; Gulliksen, B. Size, age and diet of polar cod, Boreogadus saida (Lepechin 1773), in ice covered waters. Polar Biol. 1989, 9, 187–191. [Google Scholar] [CrossRef]
- Andersen, O.; Frantzen, M.; Rosland, M.; Timmerhaus, G.; Skugor, A.; Krasnov, A. Effects of crude oil exposure and elevated temperature on the liver transcriptome of polar cod (Boreogadus saida). Aquat. Toxicol. 2015, 165, 9–18. [Google Scholar] [CrossRef] [PubMed]
- Nahrgang, J.; Camus, L.; Gonzalez, P.; Goksoyr, A.; Christiansen, J.S.; Hop, H. PAH biomarker responses in polar cod (Boreogadus saida) exposed to benzo(a)pyrene. Aquat. Toxicol. 2009, 94, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Whiteley, N.M.; Christiansen, J.S.; Egginton, S. Polar cod, Boreogadus saida (Gadidae), show an intermediate stress response between Antarctic and temperate fishes. Comp Biochem. Physiol A Mol. Integr. Physiol 2006, 145, 493–501. [Google Scholar] [CrossRef]
- Sveen, L.; Karlsen, C.; Ytteborg, E. Mechanical induced wounds in fish–A review on models and healing mechanisms. Rev. Aquac. 2020, 1–20. [Google Scholar]
- Videler, J.J. Fish Swimming; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Glover, C.N.; Bucking, C.; Wood, C.M. The skin of fish as a transport epithelium: A review. J. Comp. Physiol. B 2013, 183, 877–891. [Google Scholar] [CrossRef] [PubMed]
- Groff, J.M. Cutaneous Biology and Diseases of Fish; Veterinary Clinics of North America: Exotic Animal Practice; Elsevier: Amsterdam, The Netherlands, 2001; Volume 4, pp. 321–411. [Google Scholar]
- Rubin, M.A. Thermal reception in fishes. J. Gen. Physiol. 1935, 18, 643–647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szewciw, L.; Barthelat, F. Mechanical properties of striped bass fish skin: Evidence of an exotendon function of the stratum compactum. J. Mech. Behav. Biomed. Mater. 2017, 73, 28–37. [Google Scholar] [CrossRef] [PubMed]
- Lane, E.B.; Whitear, M. Sensory structures at the surface of fish skin: I. Putative chemoreceptors. Zool. J. Linn. Soc. 2008, 75, 141–151. [Google Scholar] [CrossRef]
- Kotrschal, K. Ecomorphology of solitary chemosensory cell systems in fish: A review. In Ecomorphology of Fishes; Luczkovich, J.J., Motta, P.J., Norton, S.F., Liem, K.F., Eds.; Springer: Dordrecht, The Netherlands, 1995; pp. 143–155. [Google Scholar]
- Päkk, P.; Hussar, P.; Järveots, T.; Paaver, T. Club cells active role in epidermal regeneration after skin hyperplasia of koi carp Cyprinus carpio. AACL Bioflux 2011, 4, 455–462. [Google Scholar]
- Halbgewachs, C.F.; Marchant, T.A.; Kusch, R.C.; Chivers, D.P. Epidermal club cells and the innate immune system of minnows. Biol. J. Linn. Soc. 2009, 98, 891–897. [Google Scholar] [CrossRef]
- Ledy, K.; Giamberini, L.; Pihan, J.C. Mucous cell responses in gill and skin of brown trout (Salmo trutta) fario in acidic, aluminium-containing stream water. Dis. Aquat. Organ. 2003, 56, 235–240. [Google Scholar] [CrossRef]
- Holm, H.J.; Skugor, S.; Bjelland, A.K.; Radunovic, S.; Wadsworth, S.; Koppang, E.O.; Evensen, Ø. Contrasting expression of immune genes in scaled and scaleless skin of Atlantic salmon infected with young stages of Lepeophtheirus salmonis. Dev. Comp. Immunol. 2017, 67, 153–165. [Google Scholar] [CrossRef]
- Esteban, M. An overview of the immunological defenses in fish skin. ISRN Immunol. 2012, 2012, 29. [Google Scholar]
- Berntssen, M.H.G.; Kroglund, F.; Rosseland, B.O.; Wendelaar Bonga, S.E. Responses of skin mucous cells to aluminium exposure at low pH in Atlantic salmon (Salmo salar) smolts. Can. J. Fish. Aquat. Sci. 1997, 54, 1039–1045. [Google Scholar] [CrossRef] [Green Version]
- Zuchelkowski, E.M.; Lantz, R.C.; Hinton, D.E. Effects of acid-stress on epidermal mucous cells of the brown bullhead Ictalurus nebulosus (LeSeur): A morphometric study. Anat. Rec. 1981, 200, 33–39. [Google Scholar] [CrossRef]
- Sveen, L.R.; Timmerhaus, G.; Krasnov, A.; Takle, H.; Handeland, S.; Ytteborg, E. Wound healing in post-smolt Atlantic salmon (Salmo salar L.). Sci. Rep. 2019, 9, 3565. [Google Scholar] [CrossRef] [Green Version]
- Tadiso, T.M.; Krasnov, A.; Skugor, S.; Afanasyev, S.; Hordvik, I.; Nilsen, F. Gene expression analyses of immune responses in Atlantic salmon during early stages of infection by salmon louse (Lepeophtheirus salmonis) revealed bi-phasic responses coinciding with the copepod-chalimus transition. BMC Genom. 2011, 12, 141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karlsen, C.; Ytteborg, E.; Timmerhaus, G.; Høst, V.; Handeland, S.; Jørgensen, S.M.; Krasnov, A. Atlantic salmon skin barrier functions gradually enhance after seawater transfer. Sci. Rep. 2018, 8, 9510. [Google Scholar] [CrossRef]
- Kortner, T.M.; Skugor, S.; Penn, M.H.; Mydland, L.T.; Djordjevic, B.; Hillestad, M.; Krasnov, A.; Krogdahl, A. Dietary soyasaponin supplementation to pea protein concentrate reveals nutrigenomic interactions underlying enteropathy in Atlantic salmon (Salmo salar). BMC Vet. Res. 2012, 8, 101. [Google Scholar] [CrossRef] [Green Version]
- Karlsen, C.; Sorum, H.; Willassen, N.P.; Asbakk, K. Moritella viscosa bypasses Atlantic salmon epidermal keratocyte clearing activity and might use skin surfaces as a port of infection. Vet. Microbiol 2012, 154, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Elliott, D. Functional morphology of the integumentary system in fishes. In Encyclopedia of Fish Physiology: From Genome to Environment; Farrell, A.P., Ed.; Academic Press: San Diego, CA, USA, 2011; Volume 1, pp. 476–488. [Google Scholar]
- Fontenot, D.K.; Neiffer, D.L. Wound management in teleost fish: Biology of the healing process, evaluation, and treatment. Vet. Clin. North Am. Exot. Anim. Pract. 2004, 7, 57–86. [Google Scholar] [CrossRef]
- Lazado, C.C.; Caipang, C.M. Probiotics-pathogen interactions elicit differential regulation of cutaneous immune responses in epidermal cells of Atlantic cod Gadus morhua. Fish Shellfish. Immunol. 2014, 36, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Rajan, B.; Fernandes, J.M.; Caipang, C.M.; Kiron, V.; Rombout, J.H.; Brinchmann, M.F. Proteome reference map of the skin mucus of Atlantic cod (Gadus morhua) revealing immune competent molecules. Fish Shellfish. Immunol. 2011, 31, 224–231. [Google Scholar] [CrossRef]
- Torrissen, O.; Jones, S.; Asche, F.; Guttormsen, A.; Skilbrei, O.T.; Nilsen, F.; Horsberg, T.E.; Jackson, D. Salmon lice–Impact on wild salmonids and salmon aquaculture. J. Fish Dis. 2013, 36, 171–194. [Google Scholar] [CrossRef] [Green Version]
- Treasurer, J.W.; Grant, A. The efficacy of hydrogen peroxide for the treatment of farmed Atlantic salmon, Salmo salar L. infested with sea lice (Copepoda: Caligidae). Aquaculture 1997, 148, 265–275. [Google Scholar] [CrossRef]
- Rapanan, J.L.; Pascual, A.S.; Uppalapati, C.K.; Cooper, K.E.; Leyva, K.J.; Hull, E.E. Zebrafish keratocyte explants to study collective cell migration and reepithelialization in cutaneous wound healing. J. Vis. Exp. JoVE 2015, 96, e52489. [Google Scholar] [CrossRef] [Green Version]
- Keren, K.; Pincus, Z.; Allen, G.M.; Barnhart, E.L.; Marriott, G.; Mogilner, A.; Theriot, J.A. Mechanism of shape determination in motile cells. Nature 2008, 453, 475–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sveen, L.R.; Timmerhaus, G.; Krasnov, A.; Takle, H.; Stefansson, S.O.; Handeland, S.O.; Ytteborg, E. High fish density delays wound healing in Atlantic salmon (Salmo salar). Sci. Rep. 2018, 8, 16907. [Google Scholar] [CrossRef] [PubMed]
- Mota, V.C.; Nilsen, T.O.; Gerwins, J.; Gallo, M.; Ytteborg, E.; Baeverfjord, G.; Kolarevic, J.; Summerfelt, S.T.; Terjesen, B.F. The effects of carbon dioxide on growth performance, welfare, and health of Atlantic salmon post-smolt (Salmo salar) in recirculating aquaculture systems. Aquaculture 2019, 498, 578–586. [Google Scholar] [CrossRef]
- Oeffner, J.; Lauder, G.V. The hydrodynamic function of shark skin and two biomimetic applications. J. Exp. Biol. 2012, 215, 785–795. [Google Scholar] [CrossRef] [Green Version]
- Wainwright, S.A.; Vosburgh, F.; Hebrank, J.H. Shark skin: Function in Locomotion. Science 1978, 202, 747–749. [Google Scholar] [CrossRef]
- De Vrieze, E.; Sharif, F.; Metz, J.R.; Flik, G.; Richardson, M.K. Matrix metalloproteinases in osteoclasts of ontogenetic and regenerating zebrafish scales. Bone 2011, 48, 704–712. [Google Scholar] [CrossRef] [Green Version]
- Harvey, R.; Batty, R.S. Cutaneous taste buds in cod. J. Fish Biol. 1998, 53, 138–149. [Google Scholar] [CrossRef]
- Hansen, A.; Reutter, K.; Zeiske, E. Taste bud development in the zebrafish, Danio rerio. Dev. Dyn. 2002, 223, 483–496. [Google Scholar] [CrossRef]
- Brown, G.; Wellings, S. Electron microscopy of the skin of the teleost, Hippoglossoides elassodon. Z. Zellforsch. Mikrosk. Anat. 1970, 103, 149–169. [Google Scholar] [CrossRef] [PubMed]
- Quilhac, A.; Sire, J.Y. Spreading, proliferation, and differentiation of the epidermis after wounding a cichlid fish, Hemichromis bimaculatus. Anat. Rec. 1999, 254, 435–451. [Google Scholar] [CrossRef]
- Sire, J.-Y.; Donoghue, P.C.J.; Vickaryous, M.K. Origin and evolution of the integumentary skeleton in non-tetrapod vertebrates. J. Anat. 2009, 214, 409–440. [Google Scholar] [CrossRef]
- Sire, J.-Y.; Donoghue, P.C.J.; Vickaryous, M.K. Scale development in zebrafish (Danio rerio). J. Anat. 1997, 190, 545–561. [Google Scholar] [CrossRef]
- Rotllant, J.; Redruello, B.; Guerreiro, P.; Fernandes, H.; Canario, A.V.; Power, D. Calcium mobilization from fish scales is mediated by parathyroid hormone related protein via the parathyroid hormone type 1 receptor. Regul. Pept. 2005, 132, 33–40. [Google Scholar] [CrossRef]
- Persson, P.; Takagi, Y.; Björnsson, B.T. Tartrate resistant acid phosphatase as a marker for scale resorption in rainbow trout, Oncorhynchus mykiss: Effects of estradiol-17β treatment and refeeding. Fish Physiol. Biochem. 1995, 14, 329–339. [Google Scholar] [CrossRef]
- Yasuo, M. The source of calcium in regenerating scales of the goldfish Carassius auratus. Comp. Biochem. Physiol. Part A Physiol. 1980, 66, 521–524. [Google Scholar] [CrossRef]
- Richardson, R.; Slanchev, K.; Kraus, C.; Knyphausen, P.; Eming, S.; Hammerschmidt, M. Adult zebrafish as a model system for cutaneous wound healing research. J. Investig. Dermatol. 2013, 133, 1655–1665. [Google Scholar] [CrossRef] [Green Version]
- Rawlings, D.J.; Witte, O.N. Bruton’s tyrosine kinase is a key regulator in B-cell development. Immunol. Rev. 1994, 138, 105–119. [Google Scholar] [CrossRef]
- Watts, T.H. TNF/TNFR family members in costimulation of T cell responses. Annu. Rev. Immunol. 2005, 23, 23–68. [Google Scholar] [CrossRef] [PubMed]
- Jensen, L.B.; Wahli, T.; McGurk, C.; Eriksen, T.B.; Obach, A.; Waagbo, R.; Handler, A.; Tafalla, C. Effect of temperature and diet on wound healing in Atlantic salmon (Salmo salar L.). Fish Physiol Biochem. 2015, 41, 1527–1543. [Google Scholar] [CrossRef]
- Pittman, K.; Pittman, A.; Karlson, S.; Cieplinska, T.; Sourd, P.; Redmond, K.; Ravnøy, B.; Sweetman, E. Body site matters: An evaluation and application of a novel histological methodology on the quantification of mucous cells in the skin of Atlantic salmon, Salmo salar L. J. Fish Dis. 2013, 36, 115–127. [Google Scholar] [CrossRef]
- Pevzner, R.A.; Hernádi, L.; Salánki, J. Effect of mercury on the fish (Alburnus alburnus) chemoreceptor taste buds. A scanning electron microscopic study. Acta Biol. Hung. 1986, 37, 159–167. [Google Scholar]
- Iger, Y.; Bonga, S.E.W. Cellular responses of the skin of carp (Cyprinus carpio) exposed to acidified water. Cell Tissue Res. 1994, 275, 481–492. [Google Scholar] [CrossRef] [Green Version]
- Peterson, L.L.; Zettergren, J.G.; Wuepper, K.D. Biochemistry of transglutaminases and cross-linking in the skin. J. Investig. Dermatol. 1983, 81 (Suppl. S1), 95s–100s. [Google Scholar] [CrossRef] [Green Version]
- Sveen, L.R.; Timmerhaus, G.; Torgersen, J.S.; Ytteborg, E.; Jørgensen, S.M.; Handeland, S.; Stefansson, S.O.; Nilsen, T.O.; Calabrese, S.; Ebbesson, L.; et al. Impact of fish density and specific water flow on skin properties in Atlantic salmon (Salmo salar L.) post-smolts. Aquaculture 2016, 464, 629–637. [Google Scholar] [CrossRef] [Green Version]
- Christiansen, J.; Jobling, M.; Amin, A.; Ribe, H. Histomorphological changes in epidermal structure of juvenile Arctic charr (Salvelinus alpinus L.) subjected to sustained exercise. Can. J. Zool. Rev. Can. De Zool. 1991, 69, 2450–2455. [Google Scholar] [CrossRef]
- Vatsos, I.N.; Kotzamanis, Y.; Henry, M.; Angelidis, P.; Alexis, M. Monitoring stress in fish by applying image analysis to their skin mucous cells. Eur. J. Histochem. 2010, 54, e22. [Google Scholar] [CrossRef] [Green Version]
- Vera, L.M.; Migaud, H. Hydrogen peroxide treatment in Atlantic salmon induces stress and detoxification response in a daily manner. Chronobiol Int. 2016, 33, 530–542. [Google Scholar] [CrossRef] [Green Version]
- Valenzuela-Muñoz, V.; Gallardo-Escárate, A.; Sáez-Vera, C.; Garcés, F.; Bonfatti, J.; Gallardo-Escárate, C. More than bubbles: In vivo assessment and transcriptome modulation of Caligus rogercresseyi and Atlantic salmon exposed to hydrogen peroxide (PARAMOVE®). Aquaculture 2020, 522, 735170. [Google Scholar] [CrossRef]
- Overton, K.; Samsing, F.; Oppedal, F.; Dalvin, S.; Stien, L.H.; Dempster, T. The use and effects of hydrogen peroxide on salmon lice and post-smolt Atlantic salmon. Aquaculture 2018, 486, 246–252. [Google Scholar] [CrossRef]
- Ream, R.A.; Theriot, J.A.; Somero, G.N. Influences of thermal acclimation and acute temperature change on the motility of epithelial wound-healing cells (keratocytes) of tropical, temperate and Antarctic fish. J. Exp. Biol. 2003, 206, 4539–4551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rapanan, J.L.; Cooper, K.E.; Leyva, K.J.; Hull, E.E. Collective cell migration of primary zebrafish keratocytes. Exp. Cell Res. 2014, 326, 155–165. [Google Scholar] [CrossRef]
- Skugor, A.; Krasnov, A.; Andersen, O. Genome-wide microarray analysis of Atlantic cod (Gadus morhua) oocyte and embryo. BMC Genom. 2014, 15, 594. [Google Scholar] [CrossRef] [Green Version]
- Krasnov, A.; Timmerhaus, G.; Afanasyev, S.; Jorgensen, S.M. Development and assessment of oligonucleotide microarrays for Atlantic salmon (Salmo salar L.). Comp. Biochem. Physiol. Part D Genom. Proteom. 2011, 6, 31–38. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Fold |
|---|---|
| Natterin | −2.8 |
| Leukocyte cell-derived chemotaxin 2 | −1.8 |
| Myeloid-specific peroxidase | −2.0 |
| Matrix metalloproteinase 13 | −3.2 |
| Matrix metalloproteinase 13a | −4.6 |
| Claudin g | −4.8 |
| Transglutaminase 2, like | −18.7 |
| Laminin gamma_3 | −7.3 |
| Laminin subunit beta-1 | −13.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ytteborg, E.; Hansen, Ø.J.; Høst, V.; Afanasyev, S.; Vieweg, I.; Nahrgang, J.; Krasnov, A. Morphology, Transcriptomics and In Vitro Model of Skin from Polar Cod (Boreogadus Saida) and Atlantic Cod (Gadus Morhua). Fishes 2020, 5, 34. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5040034
Ytteborg E, Hansen ØJ, Høst V, Afanasyev S, Vieweg I, Nahrgang J, Krasnov A. Morphology, Transcriptomics and In Vitro Model of Skin from Polar Cod (Boreogadus Saida) and Atlantic Cod (Gadus Morhua). Fishes. 2020; 5(4):34. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5040034
Chicago/Turabian StyleYtteborg, Elisabeth, Øyvind Johannes Hansen, Vibeke Høst, Sergey Afanasyev, Ireen Vieweg, Jasmine Nahrgang, and Aleksei Krasnov. 2020. "Morphology, Transcriptomics and In Vitro Model of Skin from Polar Cod (Boreogadus Saida) and Atlantic Cod (Gadus Morhua)" Fishes 5, no. 4: 34. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5040034