Case Studies Demonstrate That Common Carp Can Be Sustainably Reduced by Exploiting Source-Sink Dynamics in Midwestern Lakes


Department of Fisheries, Wildlife, and Conservation Biology, University of Minnesota, St. Paul, MN 55108, USA
*
Author to whom correspondence should be addressed.
Fishes 2020, 5(4), 36; https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5040036
Submission received: 18 October 2020
/
Revised: 26 November 2020
/
Accepted: 30 November 2020
/
Published: 4 December 2020
(This article belongs to the Special Issue Biology and Control of Invasive Fishes)
Abstract
:The common carp has been highly problematic in North American ecosystems since its introduction over a century ago. In many watersheds, its abundance appears to be driven by source-sink dynamics in which carp reproduce successfully in peripheral ponds that lack egg/larva micro-predators which then serve as sources of recruits for deeper lakes. This manuscript describes how carp were sustainably reduced in two chains of lakes by disrupting source-sink dynamics in three steps. First, we ascertained whether lakes had problematic densities of carp that could be explained by source-sink dynamics. Second, ways to control recruitment were developed and implemented including: (i) aerating source ponds to reduce hypoxia and increase micro-predator abundance, (ii) blocking carp migration, and (iii) locating and removing adults from sinks using targeted netting guided by Judas fish. Third, we monitored and adapted. Using this strategy, the density of carp in 3 lakes in one chain was reduced from 177 kg/ha to ~100 kg/ha in 3 years and held constant for a decade. Similarly, adult density was reduced from 300 kg carp/ha in 2 lakes in the other chain to 25 kg/ha. Once carp densities were low, aluminum sulfate treatments became reasonable and once conducted, water quality improved.
1. Introduction
The common carp, Cyprinus carpio, (hereafter “carp”) is one of the world’s most invasive fishes and likely to become more as our climate warms [1]. Originally from Eurasia, it was initially moved across Europe by the Romans, and from there was exported across the globe in 1800s [2,3]. It is now found in all continents except Antarctica [2,3,4]. Although the carp is a valuable sport and food fish in many European countries as well as China [5], it has been stocked and become invasive in Africa, Australia, South America and North America [3]. Adult common carp are highly proficient at feeding in bottom sediments where they damage rooted vegetation, release nutrients and sediments into the water column, and degrade water quality (clarity) in shallow ecosystems [6,7]. This process also often causes dramatic reductions in biodiversity and waterfowl [6,8]. Carp control is a particularly onerous issue in North America [3] where control has traditionally focused on drawing-down and/or poisoning (rotenone) entire lakes and wetlands heavily infested with this species, and then installing barriers to prevent re-infestation [9,10,11,12]. However, this approach is extremely expensive and ecologically damaging, not applicable in many locales, and very often not sustainable as carp usually reinvade within 5–10 years [6,10]. New solutions to carp control are sought.
Although the common carp is generally considered to be a large river fish [13], it also does very well in many temperate lakes, especially systems of shallow eutrophic lakes with extensive littoral zones or lakes associated with such areas [6,14]. Adult carp are seasonal spawners and migrate, sometimes great distances, into shallow waters (floodplains, wetlands, ponds or shallow lakes) where females release up 3 million eggs each onto submersed vegetation [15,16]. After hatching, larval and juvenile carp forage on plankton in weed beds for 1–2 years but grow quickly [17,18,19,20,21]. Depending on the local environment, carp mature at 2–5 years of age and can live to be over 60 years of age, often reaching weights of up to 10 kg [2,15]. A single female carp might release 100 s of millions of eggs in her life. In shallow lakes in Midwestern North America, correlations between adult carp density, aquatic plant community composition, and water clarity, show that when adult carp densities reach about 100 kg/ha they drive about a 50% reduction in native plant cover, a value which many accept as a threshold for unacceptable ecological damage, and which almost always is associated with a shift in stable state and cyanobacteria blooms along with decreased in water quality [6]. When carp biomass exceeds 200 kg/ha, aquatic plants are typically almost absent [22,23].
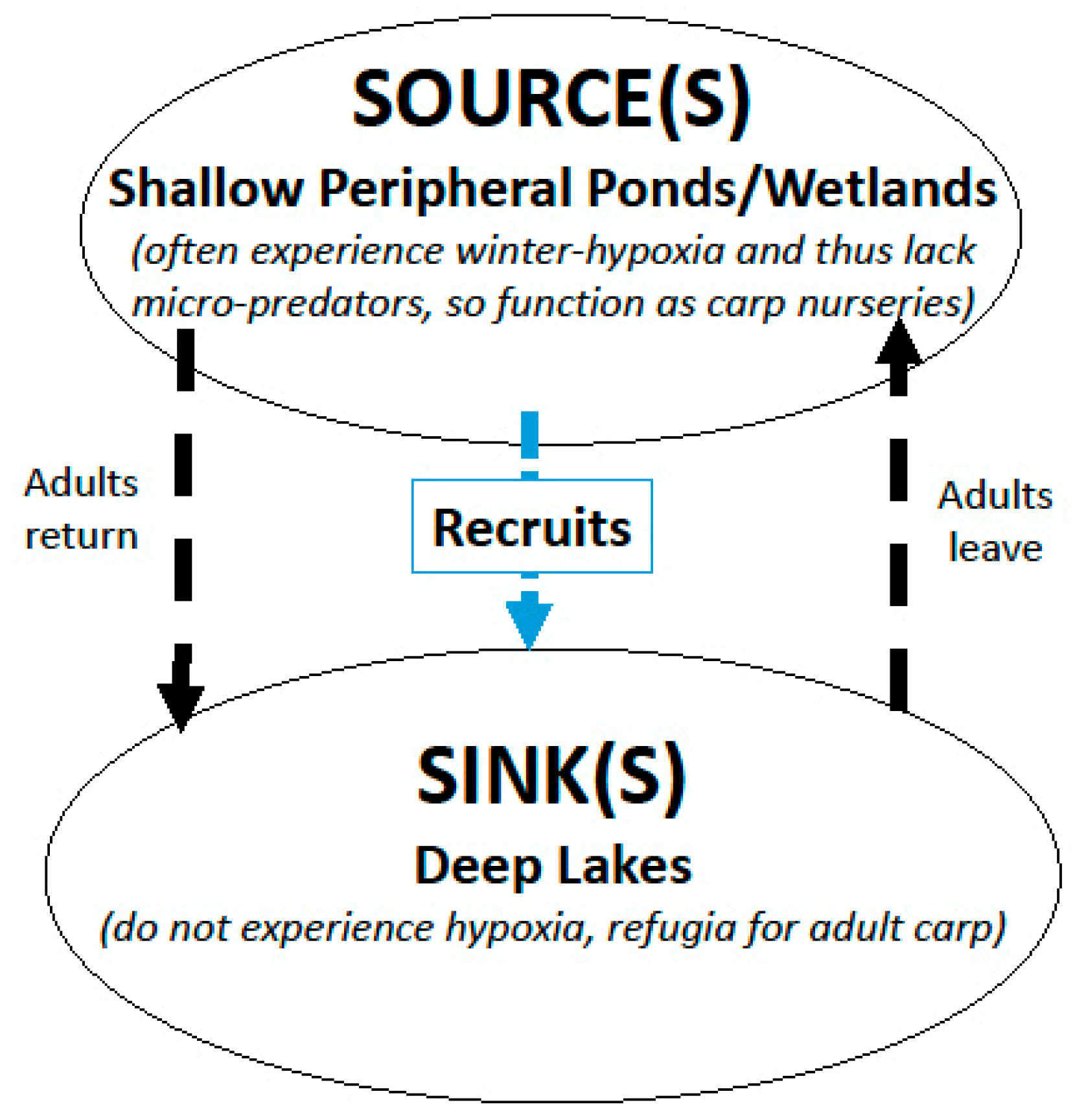
For a population of fish to become highly abundant and invasive, its recruitment rate must on average, exceed its mortality and emigration rates. This can happen in many ways. In many inter-connected glacial lakes in the Midwestern North America, a variety of evidence suggests this often happens because adult carp migrate in the spring into peripheral waters which lack egg and larva micro-predators (fish with small gapes that consume eggs and larvae) because they often “winterkill” (suffer from low oxygen in winter which leads to fish kills) [16]. In many of these situations, young carp are able to survive, grow quickly and leave these nurseries for deeper lakes that do not support self-sustaining populations themselves because they do not winterkill (and thus have native micro-predators). Some shallow sources may also support adults on occasion because they only sporadically winterkill, enabling them to also function as sources. This scenario results in watersheds in which peripheral shallow waterbodies including floodplains, wetlands, ponds and shallow lakes can serve as population “sources”, while deeper lakes function as “sinks”—a scenario known as a “source-sink” (Figure 1) [21,23,24,25]. Although not well studied, source-sink dynamics appear to be common in many watersheds in the North American Midwest, especially in its temperate forested zones where shallow lakes that frequently winterkill are commonly connected, sometimes in chains which flow to rivers or deep lakes [14,16,23]. Importantly, source-sink dynamics create special opportunities for controlling invasive species [25] because their recruitment is spatially restricted to a few locations so they can be easier to manage, as can removal of adults from population sinks.
Suspicions that adult carp might be superabundant in some lakes in the temperate forest ecoregion of the Midwest arose during the course of a research project conducted by the authors that sought to find the source of young carp in these systems while describing the damage they cause [16]. This work and the results of a dozen or so studies of the questions it addressed [16,18,21,26,27] eventually served as the foundation for long-term carp reduction and control efforts taken over by agency biologists and are described in this manuscript along with our studies. While some of this work has been published (and is referenced), much of it has not been previously published so it is presented here for the first time. The present manuscript describes two decade-long efforts to control carp based on the premise that source-sink dynamics can be sustainably exploited to control carp. In particular, we describe control programs in the two chains of lakes, the Phalen Chain of Lakes and the Riley Chain of Lakes, which have been studied and controlled for the longest period of time. First, we describe our overarching strategy (which we developed over the course of this study), and then how it was implemented, and finally our results.
2. Overarching Strategy and Approach
The overarching goal of Sorensen’s and more recently Bajer’s research on the common carp has been to develop an understanding of its biology and population dynamics in the Midwest so as to permit development of a sustainable way to reduce and control it. It has been an iterative process which required that we develop many new techniques including the ability to assess the abundance, distribution, and movement patterns of both juvenile and adult carp in our area and then explain them, and the level of ecological damaged caused by this species. During the course of conducting these studies, we developed a stepwise strategy and accompanying set of techniques to assess and control this species based on the observed source-sink dynamics. The present manuscript describes the results of this combined research and management effort in two chains of lakes. While research started in the Riley Chain of Lakes, control was eventually pursued more actively in the Phalen Chain of Lakes so we describe that system first. First, we provide a brief overview of the three steps, some of which were at times conducted concurrently and in an adaptive fashion.
- Step 1: Assessing the density and distribution of juvenile and adult carp. We found that before we could consider carp control, we first had to determine the location and number of adult and juvenile carp in the systems, the extent of their populations (carp movement patterns), and whether there were sources and sinks. We developed several techniques including different types of netting, electrofishing, genetic analysis, environmental DNA, and aging to conduct this process. These are described below in the two case studies and related publications.
- Step 2: Controlling and reducing the source(s) of juvenile carp while removing adult carp. While developing an understanding of how many carp there were, we realized that we could adopt these assessment techniques and approaches to affect control. Controlling sources required that we develop strategies different from those used for sinks. Many of these were adaptations of assessment strategies (ex. seining used for mark-recapture study were modified to include removal) and conducted as part of assessment itself. Different strategies had to be developed for different locales/carp populations. These are described in the case studies described below.
- Step 3. Monitoring and Adapting. Even after carp populations had been reduced and controlled, it was evident that continued systematic monitoring was necessary owing to the high fecundity of this species and how its reproductive success was influenced by variable winter conditions. If/as increases in carp numbers were noted, new techniques to address these increases had to be developed. This adaptive effort was strongly based on assessment techniques and is described below. These data are largely taken from agency records.
3. Case-Studies of Sustainable Carp Control
Since 2006, we have been studying carp biology and developing techniques to use this understanding to reduce carp numbers in several local watersheds. Of these watersheds, the Phalen Watershed and the Riley Watershed, have received the greatest attention so their stories are presented here. As is typical to our region, both watersheds are dominated by a single chain of lakes (inter-connected lakes that drain into rivers) but they do differ in complexity and the number of peripheral shallow waters they drain.
3.1. Case Study 1: The Phalen Chain of Lakes
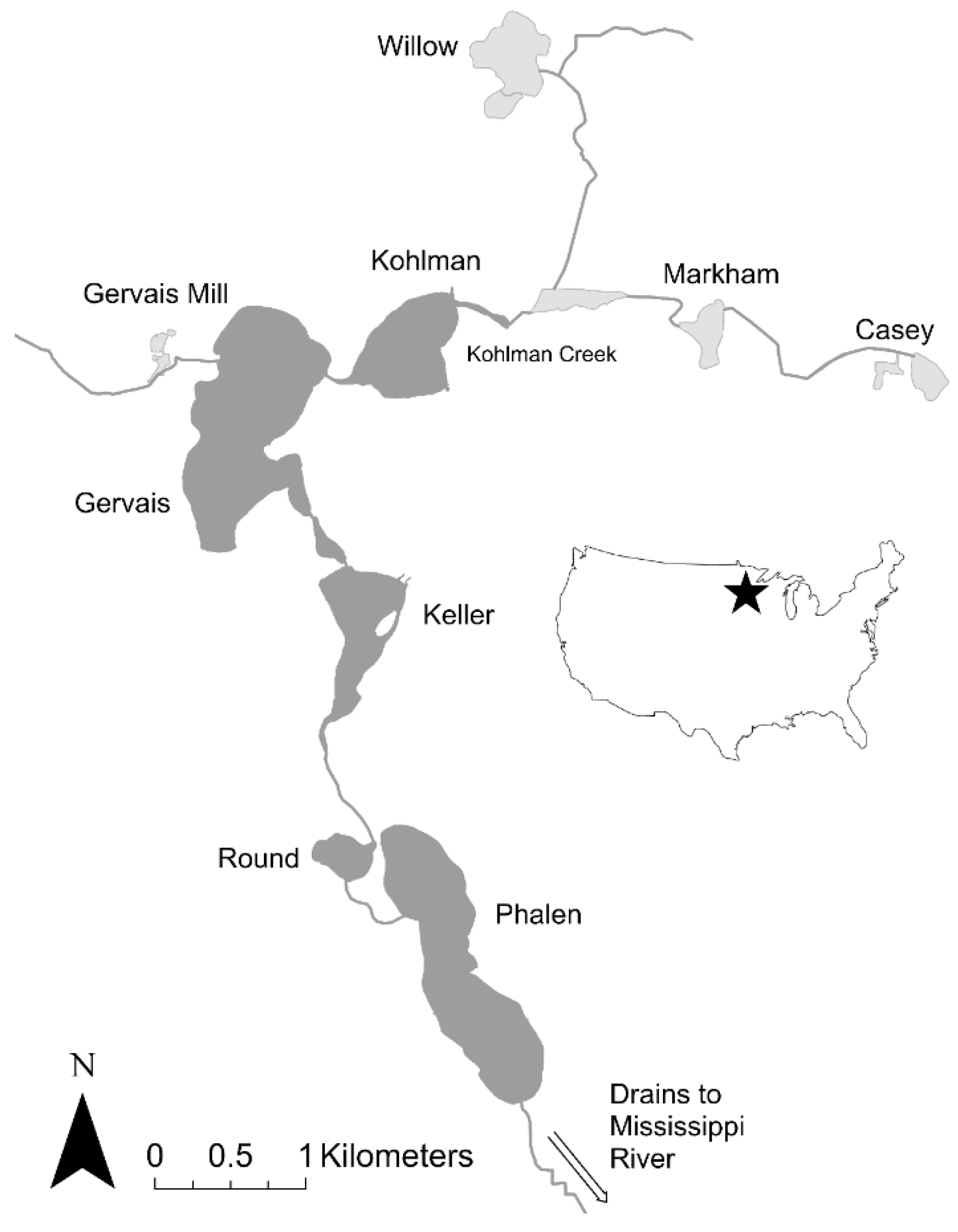
The Phalen Chain of Lakes is located in the Phalen Subwatershed in Ramsey County, Minnesota, USA (44.9884° N, 93.0545° W; Figure 2) and is similar in size and habitat complexity to many freshwater systems in other glaciated regions of the Upper Mississippi River Basin. It is densely populated and in the late 2000s was experiencing poor water quality, leading biologists in its watershed district (Ramsey Washington Metro Watershed District, RWMWD) to suspect that it might have an overabundance of common carp and to contact Sorensen. This watershed drains 6100 hectares of urban land which has been altered by human development including the draining of wetlands, ditching of creek segments, and construction of storm water ponds (Figure 2). This system contains four relatively deep lakes (surface area: 29–95 ha; maximum depth: 3–28 m) that do not winterkill, and are connected by navigable streams (Table 1). It also contains about half a dozen shallow lakes/ponds, of which Willow Lake, Casey Lake, Markham Pond, and Gervais Mill Pond are the largest (3–21 ha) and most directly connected. All of these shallow lakes/ponds are found in headwater regions of the watershed and are reported to experience periodic low winter oxygen levels and winterkills. At the time of this study, many were highly degraded and received a great deal of nutrient-enriched runoff, especially Casey Lake and Markham Pond.
3.1.1. Step 1: Assessing the Density and Distribution of Adult and Juvenile Carp
In early 2008, we started a research project to develop long-term control of carp in the Phalen Chain of Lakes. We started by concurrently evaluating the relative abundance and distribution of both young-of year (YOY) and adult common carp across the entire chain of lakes. We knew from ongoing work in the Riley Chain of Lakes (see Section 3.2 below), that the easiest and quickest way to obtain an estimate of adult fish abundance, including carp, in our lakes was boat-electrofishing [27,28]. Each of the 4 deep lakes and all of the smaller lakes/ponds we could assess were evaluated in this manner at least once following a well-established protocol which used at least four 20-min transects conducted across each entire lake in 2009. This initial analysis discovered more than 1000 adult carp to be present in the two smallest lakes (<30 ha; lakes Keller and Kohlman), over 2000 individuals in 80 ha Lake Phalen, and finally over 4000 in 95 h Lake Gervais (Table 2). Markham Pond and Casey Lake also had modest populations of small adult carp whose numbers could not be estimated because they were too small to boat-electrofish. Notably, no adult carp were caught in other lakes and ponds. Next, to determine whether adult carp in the main lakes functioned as a single population (i.e., a self-contained group of animals), their movement patterns were described for two years using radio-telemetry following techniques being developed in the Riley Chain of Lakes [16]. Briefly, 10 adult carp were captured in each lake using boat electrofishing and surgically implanted with radio-transmitter tags (model F1850; Advanced Telemetry Systems; Isanti, MN, USA) in April and May 2009. An additional 10 carp were implanted with radio-transmitters and released in June 2009 in Lake Keller. The distribution of these radio-tagged carp was then monitored by surveying the entire system using a small boat and a low frequency loop antenna 1 to 4 times a month for 1.5 years. Monthly detections were then tabulated to assess the presence of fish and percent of all detections also calculated. We found that while those adult carp originally tagged in Lake Gervais tended to move to both Keller and Kohlman lakes in the summer, they returned to Lake Gervais to overwinter and never traveled to Lake Phalen. Carp tagged in Lake Keller tended to enter lakes Gervais and Kohlman but also overwintered in Lake Gervais, although four entered Lake Phalen (one eventually returned). Carp tagged in Lake Kohlman also tended to overwinter Lake Gervais. Tagged adult carp were observed trying to enter Gervais Mill Pond and Markham Pond in 2008–2009 but water levels were too low to permit passage that spring. Finally, only one carp tagged in Lake Phalen left that lake. Based on the results of this radio telemetry study, two populations of adult common carp were identified: a Kohlman–Gervais–Keller population and a Lake Phalen population (Table 2).
Once we knew the relative abundance and movement patterns of adult carp, we sought to determine precise population and density estimates. We focused on lakes Kohlman, Keller, and Gervais as these lakes both appeared to have the highest densities of adult carp while also functioning as a single population (almost all fish overwintered in a single location, Lake Gervais). To estimate the abundance of adult carp we used mark-recapture estimates, the details of which have already been published [28,29]. Briefly, we used radio-tagged carp (“Judas fish”) to help us locate aggregations of over-wintering carp in Lake Gervais [29]. A single large aggregation was seined under-ice using a 500 m net which was pulled through the ice after being positioned around the aggregation. A total of 3537 adult carp were captured, marked with numbered t-bar tags and released. The following summer, electrofishing surveys were performed in these lakes to evaluate the ratio of marked fish to unmarked fish in the population. In total, 160 adult carp were captured of which 20 were previously marked. The average weight of these carp was 3.4 ± 0.3 kg. Using a Chapman-modified Lincoln-Petersen mark-recapture equation, the carp population size (of the three lakes) was estimated at 8041 ± 1563 individuals with an average biomass of 177 ± 35 kg/ha, in excess of what we have previously calculated to be ecologically damaging [6]. Numbers and biomass of carp present in each of these deep lakes were then calculated based on the relative distribution of the radio-tagged carp found in each lake in the summer (Table 2 and Table 3).
While assessing adult carp, we commenced efforts to determine the source(s) of young-of-year (YOY) carp (<200 mm) [30]. Trap-netting was our primary tool as it was proving useful in the Riley Chain of Lakes [26] although we supplemented it with electrofishing in ponds. Trap-netting, a type of fyke net which is anchored to the shore and strung to trap, is commonly used by fisheries management agencies including the Minnesota Department of Natural Resources (MN DNR) to sample small fish (adopted by us for YOY carp in littoral zones [26,30]. Trap-netting is especially effective in shallow waters where boats cannot reach but it is a considerable amount of work so only a limited areas of lake shores can typically be sampled each year so it not highly quantitative. Trap-netting also does not sample large carp (>200 mm) well. Examining historic trap-netting data, we found that the MN DNR had sampled lakes Kohlman, Gervais, Keller and Phalen 10 times between 1999–2009 and only once caught more than 1 YOY carp per trap-net (Table 3), and Casey and Markham ponds a total of 4 times, twice capturing 25 or more YOY carp (or greater) per trap-net. Given the paucity of data, we started to collect our own trap-net data. Following established procedures [26], each lake in the Phalen Chain was sampled using 5 trap-nets (9 mm bar) set at evenly spaced intervals around the perimeter of each lake for a 24 h period in the falls of 2009 and 2010 (when YOYs were large enough to catch). In both 2009 and 2010, large numbers of YOY carp were captured in all three shallow ponds (Willow, Casey and Markham Ponds; see below), but no YOY carp were captured in any of the four deep lakes (lakes Kohlman, Gervais, Keller, and Phalen) which also had many bluegill sunfish (Lepomis macrochirus) (CPUE ranged up to 100), an important micro-predator of carp eggs and larvae in this region but also one susceptible to winterkill [26]. Markham Pond had especially high numbers of YOY carp in both 2009 and 2010 (Catch per unit effort (CPUEs) of 47 and 104) while Lake Willow had a CPUE of 174 in 2009. None of these shallow ponds had appreciable numbers of bluegill sunfish but some had goldfish (Carassius auratus) and black bullhead catfish (Ameiurus melas). Osborne [30] conducted a mark-recapture study of YOY carp in Lake Casey and Markham Pond in 2010 and estimated the former to contain 12,703 YOY carp, and the latter, 34,782 YOY [30]. Several dozen sexually-mature adult carp (<500 mm, 2–4 years in age) were also sampled in both Casey Lake and Markham Pond using boat electrofishing which was performed as part of the recapture process [30].
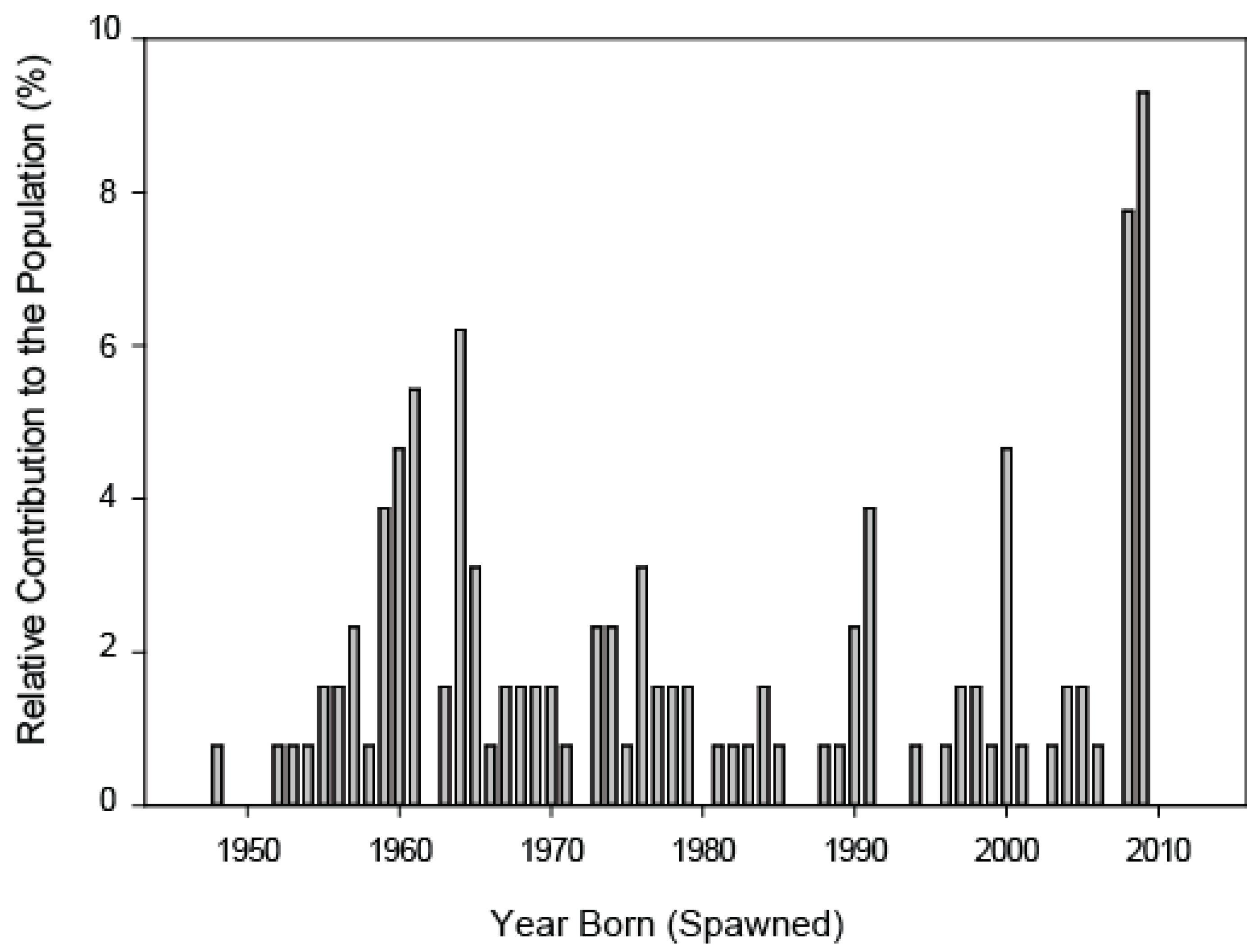
Next, to examine recruitment patterns across time we sampled and aged adult carp. A total of 127 adult carp were captured using boat-electrofishing in Lakes Kohlman, Gervais, and Keller, and then measured and weighed [21]. Following established procedures [16], their asterisci otoliths were removed and the number of annuli counted by three readers [16,31]. We found that recruitment has been very sporadic with distinct peaks in 1988–1962, 1965, 1991, 2000, 2009 (Figure 3). The oldest carp was 64 years old with the average carp being about half that age. Finally, to confirm the sources of these carp, fin samples from adults across the large lakes as well as YOY carp collected from Lake Casey and Markham Pond were analyzed for genetic variation at 12 microsatellite DNA loci (see [21] for full detail and data). This analysis described two genetic populations (strains “A” and “B”) in Lakes Kohlman, Gervais and Keller, and a third which was a hybrid of A and B. Over 80% of the adult carp that were less than 30 years old (the majority of the population), were either Strain A or the hybrid, while older fish were all strain B. The same ratio of strain A to hybrid was also found to characterize YOY captured in Casey Lake and Markham Lake, with strain B appearing to come from an unknown historical source at the western edge of the watershed which might have been Gervais Mill Pond [21]. Together, these data strongly suggested that over the past 30 years, YOY had been entering (recruiting to) lakes Kohlman, Gervais and Keller from the peripheral shallow systems as two separate genetic stocks, with Casey Lake and Markham Pond dominating recently. Notably, anecdotal observations suggested that adult carp could reach these two ponds in years when water levels were high from Lake Kohlman while young carp could almost always leave because the stream was nearly always open [21,27].
3.1.2. Step 2: Controlling the Number of Adult Common Carp while Controlling Recruitment/Sources
When we realized that there were only a few sources of carp in the Kohlman–Gervais–Keller population which functioned as a sink, we started to develop methods to reduce adults and reduce/control carp recruitment (see below). Adult carp removal commenced in 2009 as part of our effort to collect mark-recapture population estimates (see above). We took three approaches. First, as described above, we used Judas fish to find aggregating carp in the winter and then seined them using under-ice netting. We removed 3233 adult carp in the winter of 2010–2011 (3 hauls) from Lake Gervais where most carp aggregated (Table 3, [29]). Winter-netting was later repeated in the winters of 2011–2012 (825 carp removed) and 2012–2013 (no carp removed) (Table 3). Because under-ice netting is difficult / expensive and its success was waning as carp captures decreased (carp seemingly remembered and avoided the seine net), we shifted to another strategy also being developed in Riley Chain of Lakes [29,32,33] (see Section 3.2) in which adult carp are lured (trained) to food placed on top of a woven box-net (25 m x 25 m) placed on the bottom and pulled when carp are present. This versatile technique was especially successful in late summer when carp were feeding and 2233 adult carp were removed in this fashion from lakes Kohlman, Gervais and Keller in the summers of 2012–2016 (Table 3). In some lakes, a few (n = 110) adult carp were also removed using boat electrofishing in 2013 but this was not very efficient. Finally, we captured and removed 351 adult carp during their spring spawning runs into Kohlman Creek using a stream trap [27]. Annual capture patterns in the creek suggested that this creek was selected by adult carp to access their natal waters [27]. By the fall of 2016, a total of 5926 carp had been removed from this population, or about 80% of the population, and active removal efforts ceased, as values suggested densities were well below 100 kg/ha (Table 3).
While starting to remove adult carp, we undertook several efforts to eliminate carp recruitment in Markham, Casey and Willow. First, we blocked springtime access of adult carp to these ponds by building a fish barrier and trap in Kohlman Creek in the spring of 2010 (see section above). This effort was only moderately successful and its value seemed marginal, especially because we discovered a number of small adult carp overwintering in Markham, Casey and Willow [30]. Second, we attempted to drawdown Lake Casey and Markham Pond in late fall to force a winterkill and carp eradication. This process took three years, because these ponds had natural groundwater seepages, which made drawdowns difficult. Finally, in 2013 we succeeded achieving a complete drawdown and eradication of carp. We followed that with springtime stocking of native micro-predators (bluegill sunfish) and installed a small floating electric water-jet aeration system in both waterbodies to maintain the stocked predator fish. We did not have to address Willow Lake because an aeration system already in it failed, leading to the demise of all of its fish including carp. Trap-net surveys showed these efforts to be successful as YOY were never captured in these locations again (Table 4).
3.1.3. Step 3: Monitoring Carp Populations and Adapting New Management Approaches If/As Needed
While in the process of removing adult carp and controlling YOY, we commenced a monitoring program to confirm that the population was being reduced as expected. Water quality was also assessed each summer and an aluminum sulfate treatment (“alum”; a chemical treatment which binds phosphorous in the sediments) was performed in Lake Kohlman to increase water quality (see Section 3.1.4). Every year a boat electrofishing survey was conducted in all three problematic lakes following our established protocols, while trap-nets were also set each fall across all lakes following protocols to assess YOY carp abundance (Table 4). Because we did not know the precise movement patterns of adult carp between lakes on a weekly basis, and they functioned a single population, we averaged the CPUE for these lakes, weighting abundance estimates for lake size. During the course of monitoring we decided to remove additional adult carp using box-nets in 2014–2016 as well as with boat electrofishing, which was not very efficient as a removal tool (Table 4). Aeration in both outlying ponds continued along with trap-net surveys. These surveys failed to catch any YOY carp and confirmed that native micro-predators remained abundant.
3.1.4. Summary: Carp, Water Quality, and Current Status
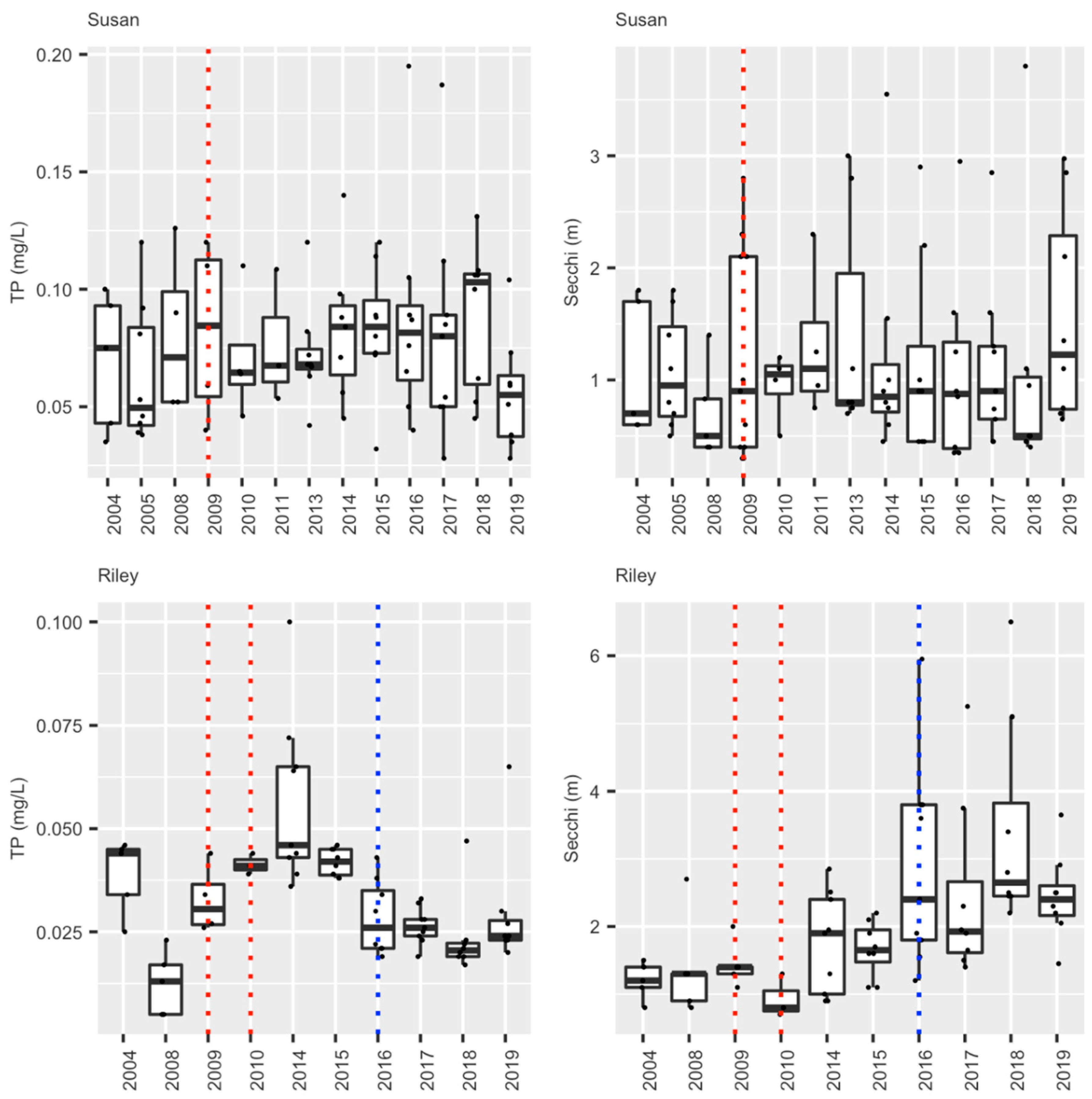
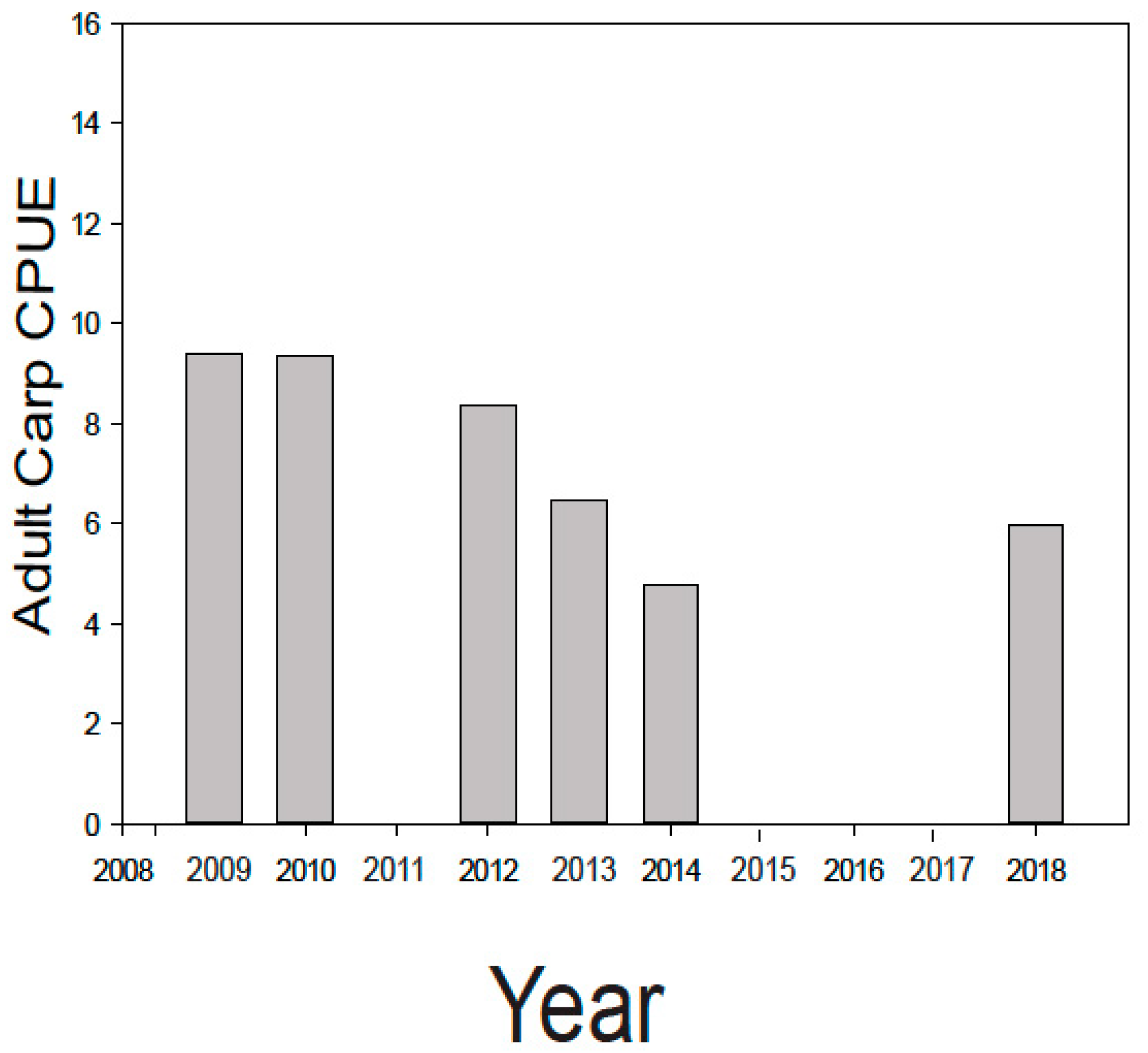
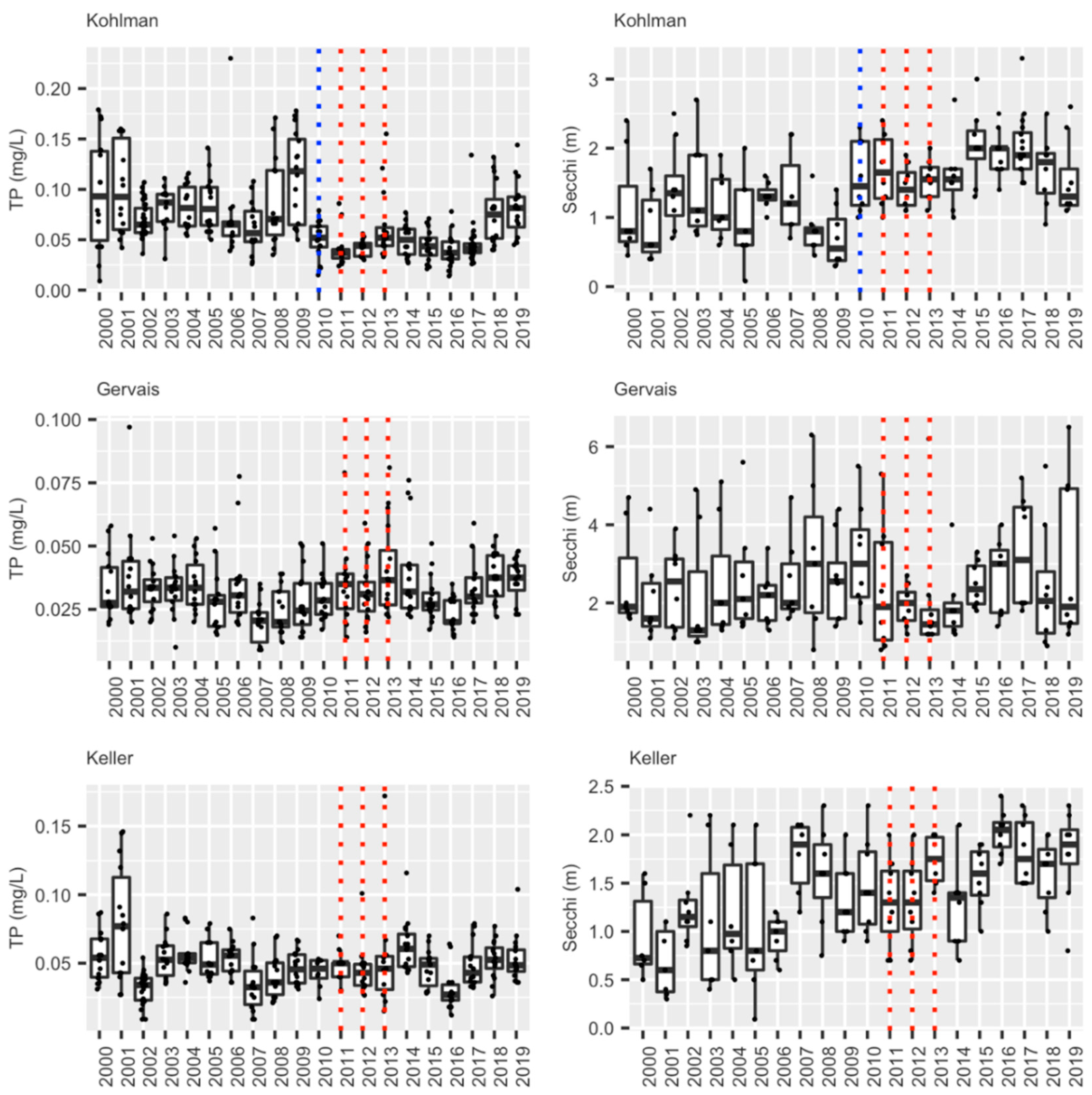
During the course of this 10 year study/ management effort, three major sources of young carp were controlled, and nearly 6000 adults removed. This integrated effort caused the carp biomass in lakes Kohlman–Gervais–Keller to fall from an average of about 177 kg/ha to a minimally-damaging level of about 40 kg/ha (mark-recapture) in 2016 from which it rose to ~100 kg/ha in 2018 (as estimated by a single electrofishing survey). In particular, electrofishing showed a drop in average adult carp CPUE (across the three lakes) from 9.41 in 2009 to 4.03 in 2014 (Figure 4). While CPUE estimates in 2018 suggested a possible slight increase in adult density, recent anecdotal reports of juvenile captures in Lake Gervais suggest this likely was caused by a few adult carp entering Gervais Mill Pond to spawn during high water events, (a possibility suggested by the genetic analyses [21]). This possible secondary source was not seemingly been identified earlier when water levels were lower, and in any case appears minor. In the future, measuring environmental DNA, or eDNA [34] might be a very useful tool for identifying possible nursery areas like this because water sampling is much easier than trapping fish. In any case, efforts are now underway to block carp from accessing Gervais Mill Pond. Meanwhile, fisheries surveys conducted by the MN DNR during this time have shown that Lake Gervais has witnessed a possible increase in gamefish from 29.2 CPUE by gill net in 2011 to 71.6 in 2017 (MN DNR). Importantly, an alum treatment was performed in Lake Kohlman to reduce benthic phosphorous release concurrent with carp removal effort while using carp exclosures to evaluate the effects of carp on the benthos and alum. It showed that carp foraging activity greatly disrupts sediments to a depth of over 13 cm [35], enough to greatly impair the benefits of alum treatment unless the carp are removed (as they were). Notably, a substantial (50%) reduction in total summer phosphorous (TP) was measured in Lake Kohlman after this joint alum treatment/carp removal. Furthermore, this lake’s water clarity doubled and remains high a decade later (Figure 5). Increased native submersed plant cover has also been measured in this shallow lake, as is common after carp removal [6,7,8]. Similar improvements in springtime plant cover were noted in Lake Gervais after carp removal, and while some improvement has also been noted in its springtime water clarity (data not shown), this improvement has not persisted into the summer in this lake which was also not yet been treated with alum because of high cost (Figure 5). Trends in Lakes Keller were similar to those in Lake Gervais (https://www.rwmwd.org/projects/keller-lake/) as it too was been treated with alum. In sum, our carp control project has been a success: most of watershed’s carp population has been sustainably and meaningfully reduced, many of its ponds are improved, as is fishing, as well as water quality especially where alum treatments occurred. Furthermore, the cost has been reasonable, and no permanent barrier or poisons were needed in this important urban system. While our carp control scheme will need to be maintained into the future and ideally a new effort made to find secondary source(s) of recruits, Ramsey-Washington Metro Watershed District is fully and officially committed to this effort as it has realized substantial benefits (personal communication, RWMWD).
3.2. Study 2. The Riley Chain of Lakes
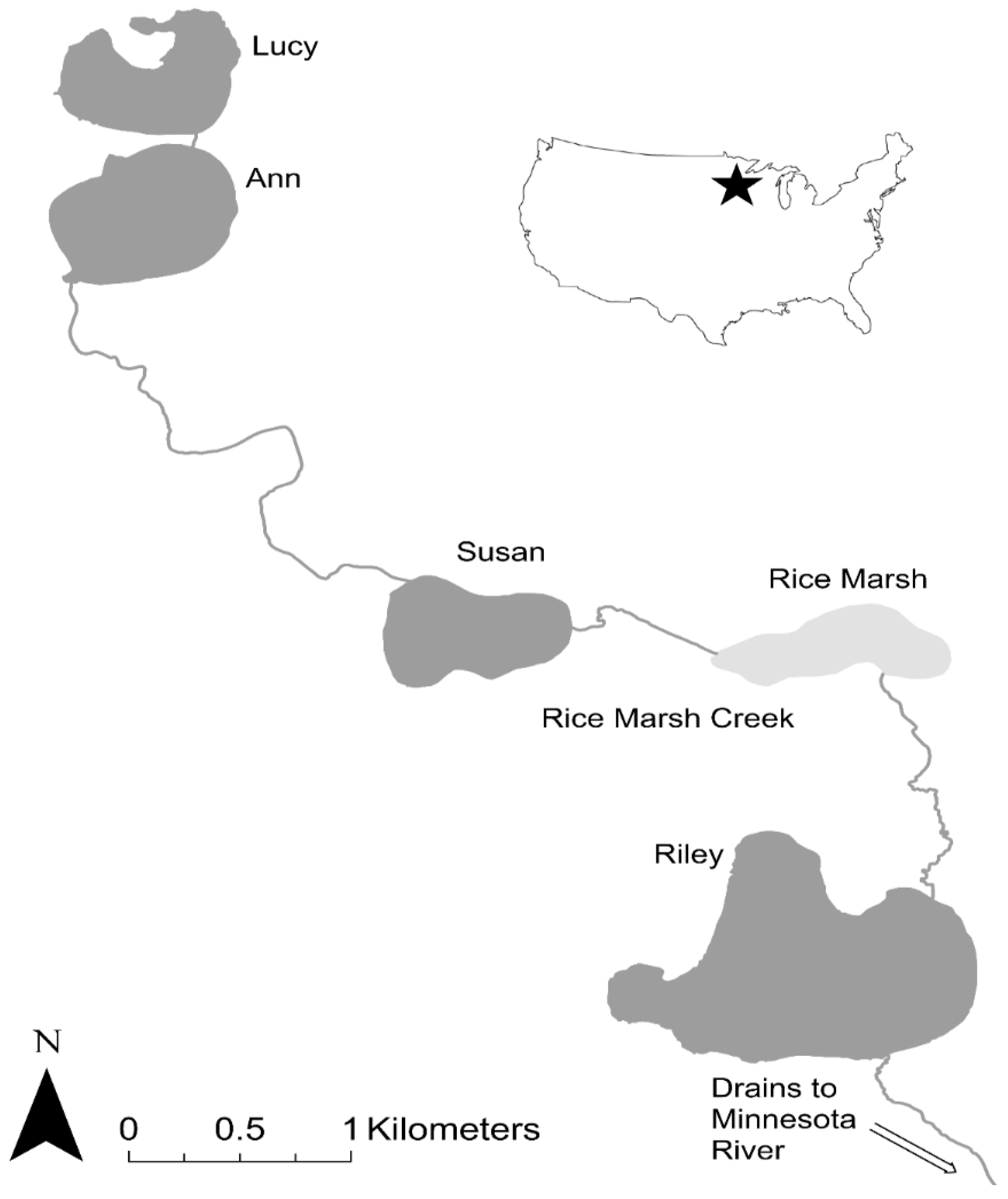
The Riley Chain of Lakes is located in Hennepin and Carver Counties, Minnesota, USA (44.8361° N, 93.5231° W; Figure 6). This watershed drains 3394 hectares of urban land through a chain of 4 interconnected lakes (surface area: 35–118 ha; maximum depth: 3–15 m) and a shallow marsh (Rice Marsh Lake) which are connected by a shallow stream (Riley Creek) and drain into the Minnesota River. All lakes except for Riley and Ann are known to occasionally winterkill, with Rice Marsh Lake being the most prone to this phenomenon, especially in recent decades (Lake Susan used to winterkill up until the early 1990s but an aeration system installed in 1992 has prevented winterkills since). The hydrology of this watershed has also been altered by human development (Table 5) and was suffering from impaired water quality in 2006 when we were approached by its watershed district, Riley Purgatory Bluff Creek Watershed District (RPBCWD), to determine if it had an overabundance of common carp and to develop solutions. Some of this work preceded that reported above in the Phalen Chain of Lakes which we showed first because it demonstrates the role of recruitment control more clearly. The work followed the same steps outlined above for the Phalen Chain of Lakes.
3.2.1. Step 1: Assessing the Density, Abundance, and Distribution of Adult and Juvenile Carp
We started this project in 2006 by evaluating the relative abundance, distribution, and movement patterns of adult common carp across the entire chain of lakes. First, we sought to determine whether adult carp in Riley and Susan (which pilot studies showed to have the most carp) functioned as a single population or several distinct populations, and to what extent carp from those lakes used the shallow lake/marsh in-between as spawning site. To determine this, we examined the movement patterns of approximately two dozen adult carp caught in each lake and also in Rice Marsh Lake, which we implanted with radio-transmitters ATS F1850; [16] and tracked for two years. We found that adult carp originally tagged in Lake Susan had a strong tendency to enter Rice Marsh Lake in the spring-summer to spawn and then return, while carp tagged in Lake Riley also tended to move into Rice Marsh Lake and then return and rarely entered Susan [37] (fully summarized in Table 6). A few fish appeared to be resident in Rice Marsh Lake, but they died when this marsh winterkilled in early 2007. No carp moved to lakes other than Lake Susan, Lake Riley, and Rice Marsh Lake while spawning was observed in all three lakes each spring. Based on both these movement data and boat electrofishing surveys, we concluded that this chain of lakes contained two major (and ecologically damaging) populations of adult carp: one centered in Lake Riley where some of the adult carp moved in and out of this system in spring to spawn in Rice Marsh Lake, and the other in Lake Susan which did the same. We then began studying both populations in more detail while conducting management efforts to reduce them below the management threshold. In the summer of 2008, we began mark-recapture analyses to precisely estimate carp abundance and biomass in lakes Susan and Riley. In Lake Susan, we used electrofishing and open water seining to mark and release 101 carp [28] in the summer 2008. We then conducted winter seining guided by radio-telemetry in March 2009. In that event, over 3000 carp were captured and removed, including 79 of the 101 marked carp [28]. This allowed us to estimate that Lake Susan had approximately 4181 adult carp with a biomass of 307 kg/ha in 2008 [28]. In Lake Riley, we also used radio-tagged carp to find aggregations of carp, seined them using an under-ice 500 m net, after which some carp were tagged and released, and then resampled the next summer and winter to calculate abundance using mark-recapture analyses [28,29]. A total of 4440 carp were captured in Lake Riley in January 2009, of which 600 were marked and released. Of the 600 marked carp, 388 were recaptured the following year among 2303 carp that were captured in another winter seine [29]. Overall, mark-recapture analyses showed that the carp population in Lake Riley in 2009 was estimated to be 6491 individuals with a biomass of 176 kg/ha [28,29], in excess of what we had previously calculated to be ecologically damaging (Table 5). The relationship between electrofishing CPUE and adult carp abundance and biomass was calculated, so boat electrofishing alone could be used here and elsewhere including the Phalen Chain to estimate carp abundance [28], see Section 3.1.
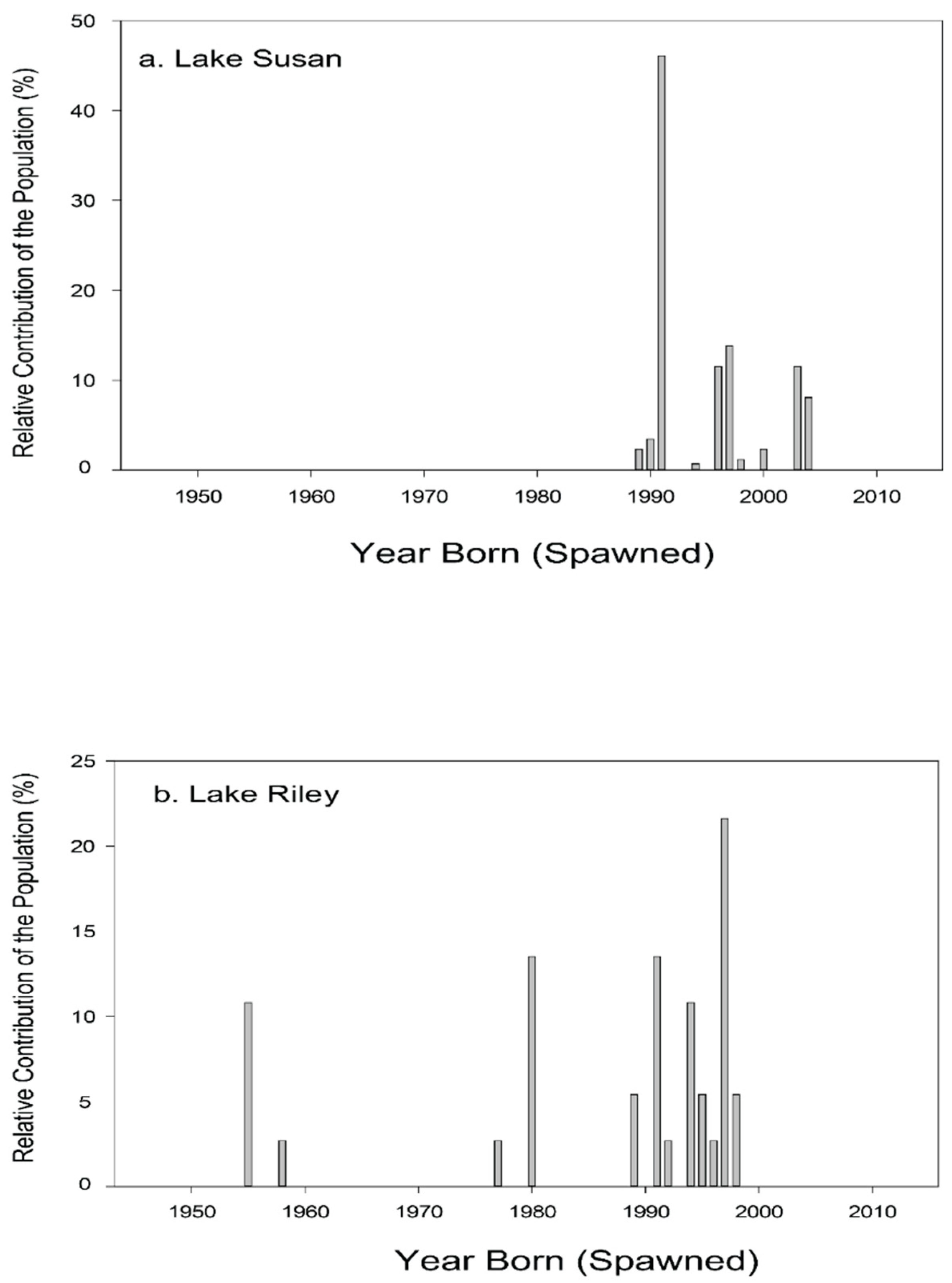
While assessing the population of adult carp, we started to assess the source(s) of these fish by examining trap-net data previously collected by the MN DNR. Although sparse (only six samples between 1997–2006), only Rice Marsh Lake was seen to have produced any YOY carp (Table 6). Starting in 2006, we began our own sampling program and set 5 trap-nets traps at evenly spaced intervals around the perimeter of each of the five lakes in the chain for 24-h every fall, similar to MN DNR protocol [26]. Very few (CPUE < 0.5) YOY carp were captured in any lake in 2006. However, on 28 March 2007, a winterkill was reported in Rice Marsh Lake and we found a large number (many hundreds) of juvenile carp, 18 of which were sampled and found to have an average length of 342 ± 10 mm (TL) and be 3 years old. Subsequent trap-netting in 2008, 2009, 2010 and 2011 showed that while Rice Marsh Lake produced a few YOY carp (CPUE = 1), the other lakes produced even smaller numbers of YOY (Table 7). No winterkill events were reported past 2007 in these lakes, but analysis of historical records showed that Lake Susan had winterkilled in 1990 and 1991 (after which it was aerated most winters), while Rice Marsh Lake had in 1997, 2000, and 2004 although these data were incomplete [16]. Lake Riley had no known records of winterkill, likely due to its depth (over 15 m). Ageing analyses and winterkill records then provided the key clue to solving the carp recruitment puzzle in the Susan and Riley systems. Examining the relationship between winterkill occurrence and carp recruitment, we collected 100 carp from Lake Susan and Lake Riley, and aged them using otoliths [16]. This analysis showed that recruitment in Lake Riley was sporadic and had not occurred since 1998 with large events in 1997, 1994, 1991, 1980 and 1955. The median age of carp in Lake Riley was 15 years (i.e., hatched in 1991) while the oldest carp was 51 years old (Figure 7a). In Lake Susan, the largest recruitment events occurred in 1989–1991, with other small events in 2004, 2003, and 1997; the median age was 15, maximum age was 17 (This lake shared large year classes with Riley in 1997 and 1991; Figure 7b). Historical records showed that winterkills had occurred in Lake Susan in 1989–91 (a winter aeration was installed in Susan in 1992 after this event which also explains the absence of older carp in this lake), and then in Rice Marsh Lake in 1997, 2000 and 2004 [16]. Together, these trends seemingly demonstrate that Lake Susan and Rice Marsh Lake had been the primary source of YOY carp in this chain of lakes for decades, and while the aeration system has prevented winterkills (and recruitment) in Lake Susan since 1991, Rice Marsh Lake has continued to serve as a source of carp.
3.2.2. Step 2—Reducing and Controlling Adult Carp while Controlling Recruitment
Efforts to control carp started in 2008 during the mark-recapture studies (Section 3.2.1) and focused on removing adults from sink populations in lakes Susan and Riley, while suppressing recruitment in Rice Marsh Lake, the putative source. (Lake Susan may have only have become a sink in early 1992 when it was first aerated and its internal source of recruits was likely removed.) We focused on winter-seining using radio-tagged Judas fish to remove carp in both Lake Susan and Lake Riley (while also conducting mark-recapture studies see above). A total of 3425 adult carp were removed from Lake Susan in this manner in single haul in March 2009 [28,29], realizing a reduction in biomass of over 80%. Later, a few hundred other adult carp were removed from Lake Susan using a barrier with a trap set in Rice Marsh Creek (Table 8). Meanwhile, a total of 5619 adult carp were removed from Lake Riley in the course of three successful winter-seines starting in 2008/2009, leaving only about 10% of the original population by number and biomass past 2011 (Table 8) [29].
Concurrent with adult removal, efforts to suppress recruitment focused on Rice Marsh Lake. In the spring of 2009 barriers with traps were installed at both the outflow of Lake Susan to Rice Marsh Lake and the inflow of Lake Riley to prevent Rice Marsh Lake from serving as a spawning site (this was also used for carp removal, see above). These barriers were maintained for 3 years and demonstrated that about 5–30% of the adult population of Lake Susan left it each spring for Rice Marsh Lake, while 5–10% left Lake Riley for Rice Marsh Lake, and fluctuated yearly with flow, suggesting this movement should be considered a type of a partial migration [27]. In addition, a winter aeration system was established in Rice Marsh Lake in 2011-2012 to prevent winterkills. This system is only operated in the winter and uses a bottom diffuser to aerate most of the lake while minimizing sediment disturbance. Winter oxygen and carp recruitment have been monitored every year since. Oxygen has not dropped below 2 ppm since its installation and no winterkills have been reported.
3.2.3. Step 3—Monitoring Carp Populations and Using Adaptive Management Approaches
While in the process of removing adult carp and suppressing recruitment, we commenced a monitoring program to confirm that the population was being reduced as expected in a sustainable and effective manner. A systematic (i.e., at least every-other year) boat electrofishing survey has been conducted in all lakes [28]. Water quality has also been monitored. An alum treatment was performed in Lake Riley once it was clear that its carp had been sustainably reduced and it was reasonable to expect lasting effects. Meanwhile, yearly autumnal trap-net surveys for YOY carp were initiated and these too showed no increase in recruitment so no new actions have been taken in this system. Native fishes are abundant in all lakes and bluegill sunfish average 53.4 sunfish/net in Rice Marsh Lake.
3.2.4. Summary: Carp, Water Quality, and Current Status
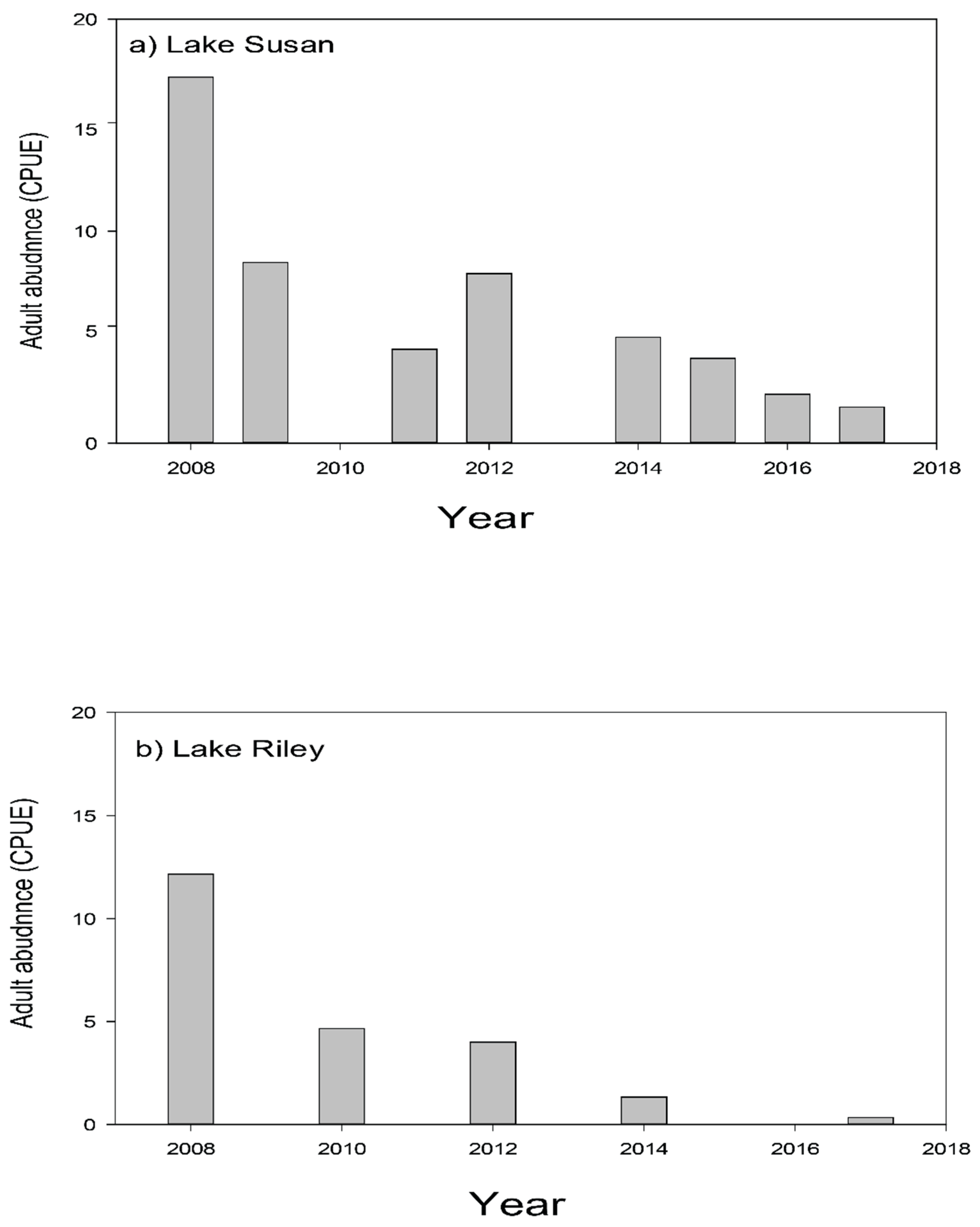
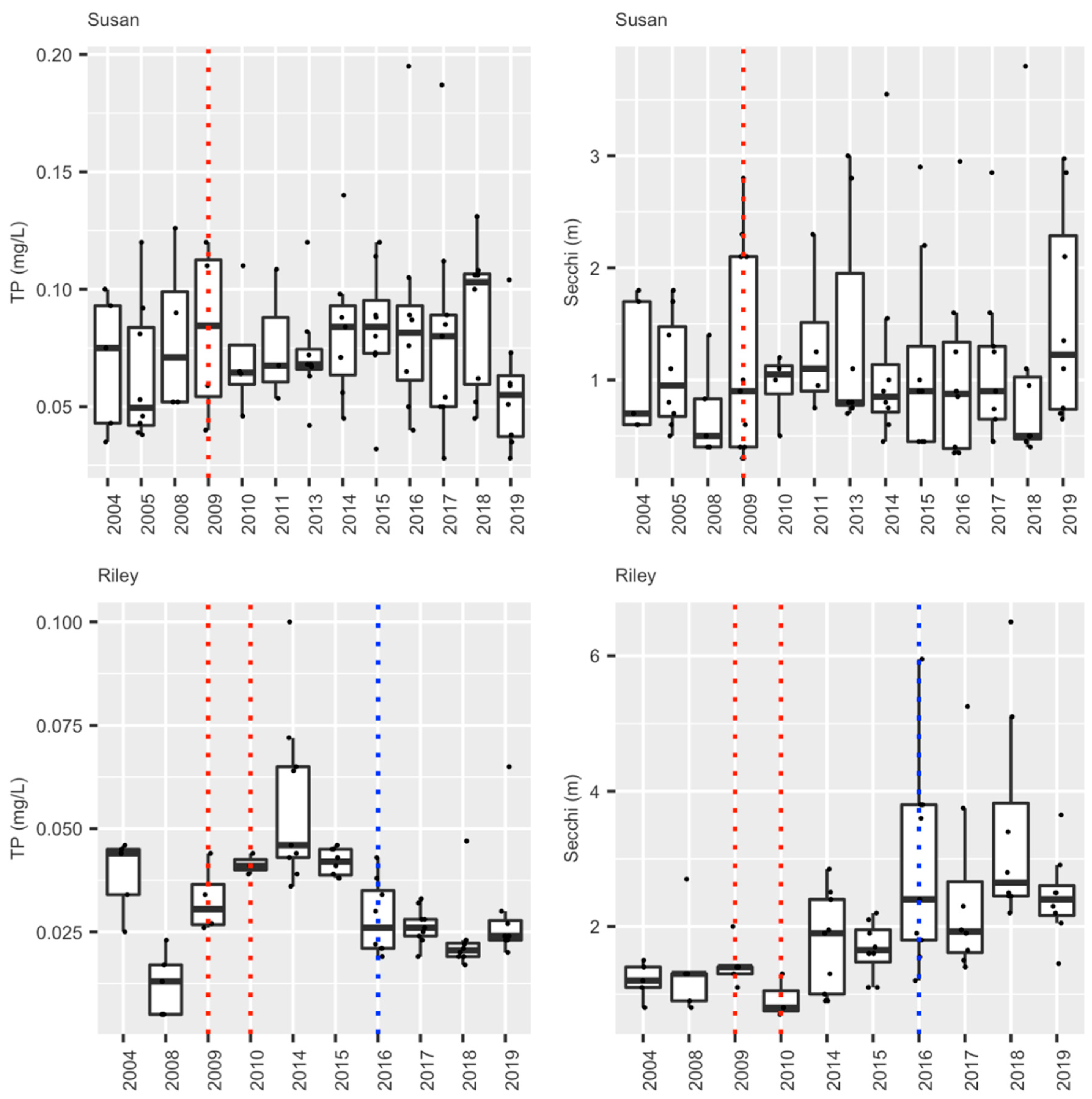
We have been able to sustainably bring carp levels down to non-damaging levels in two lakes in this watershed within several years after removing Rice Marsh Lake as its source of young, and targeting adult carp removal. Briefly, average adult CPUE across Lake Susan has fallen from over 16 carp/h in 2007 to below 2.0 carp/h in 2017, corresponding with a biomass reduction from ~307 kg/ha in 2008 to ~24 kg/ha in 2017 (Figure 8a). Similar results have been seen in Lake Riley where CPUE dropped from just over 12 (76 kg/ha) to under 1, suggesting a density of only ~15 kg/ha in 2017 (Figure 8b). It is possible that this continuous fall has been driven by natural mortality (annual mortality is estimated at 5–15% annually; [16]. Meanwhile only a handful of YOY carp have been captured during this entire time in either Lake Susan, Rice Marsh Lake, or Lake Riley (CPUE < 1; Table 7). Early springtime water clarity has increased by at least two-fold in Lake Susan, while plant cover and species richness has also increased [36]. However, while there have been small decreases in springtime total phosphorous and water clarity in this lake, this improvement has not persisted through the summer [36] (Figure 9), almost certainly because of benthic loading and hypolimnetic mixing unrelated to carp; see [35,36]. Since carp removal, submersed aquatic plant abundance and species richness have also increased greatly in Lake Riley [38,39]. Furthermore, the improvements associated with carp removal and reduction in their foraging activity have inspired the transplantation of submersed native plants into lakes Susan and Riley which have been moderately successful see [38,39]. These improvements enabled an alum treatment in Lake Riley in 2016, which led to a doubling in summer water clarity which has persisted as well a small decrease in total dissolved phosphorous (Figure 9). In sum, we have achieved successful, sustainable carp control by exploiting source-sink dynamics, and this, in turn, has enabled improvements in water quality and plant biodiversity throughout the entire watershed, especially where followed by alum treatment—the effects of which have been sustainable. The administration of RPBCWD remains committed to maintaining carp control indefinitely and is now planning an alum treatment for Lake Susan.
4. Discussion
This manuscript describes decade-long case studies of common carp management in seemingly typical chains of lakes in the North American Midwest. Both studies demonstrate that carp abundance was explained by source-sink dynamics and thus exploited in a sustainable manner to reduce carp abundance, enabling lasting improvements in water quality with alum. Both chains experienced substantial declines in carp biomass to levels that are no longer ecologically damaging as result of a combination of targeted removal and recruitment suppression at identified sources. While these systems are similar in size, control has been especially effective in the Riley Chain of Lakes. This is likely because the Phalen Chain of Lakes has many more shallow ponds (over half a dozen vs. a single pond) and thus more sources of carp including some secondary ones. Nevertheless, in both instances carp reduction has been sustainable and adequate to permit successful alum treatment which has provided long-term improvements in submersed plant cover as well as water quality. These case studies appear to be the only known examples of successful invasive fish control that do not involve expensive and damaging use of poisons and barriers. An added benefit of carp removal/control was improved springtime plant communities and some apparent improvement in some game fisheries. It seems likely that our three-step approach based on source-sink dynamics could work in other chains of lakes in the Midwest and perhaps elsewhere to control populations of carp and other invasive fishes whose abundance might also be explained by source-sink dynamics.
The key to our strategy was likely our ability to identify sources of young carp and then suppress them for extended periods. Because of the high fecundity of adult carp and their longevity in Midwestern waters, these efforts needed (and still need) to continue for many years. As mentioned, this was a simpler task in the Lake Riley Chain of Lakes which seemingly had a single source (Rice Marsh Lake) than the Phalen Chain of Lakes which had many (Casey, Markham, Gervais Mill Pond). Nevertheless, because the solution involved restoring ponds in the Phalen Chain of Lakes that had previously been considered problematic, the effort is considered justifiable by the watershed district. It is notable that winter aeration alone was sufficient to control carp recruitment and native fisheries quickly rebounded albeit with some stocking in lakes Casey and Markham. In fact, Lake Casey is now a popular urban fishing lake. Increasingly strong evidence that adult carp return (home) to the source waters also made the task somewhat easier because carp do not appear to readily move to different ponds for spawning if blocked so efforts can be targeted at just a few locations [21,27,40]. In spite of our success, a significant challenge remains in these watersheds because of the longevity of adult carp which requires these efforts and monitoring continue for decades, but similar efforts are already spent for game fish. Lake systems with discrete and discontinuous patterns of recruitment such as found in many lakes of the temperate forest ecoregion of the Midwest are likely easier to control than other systems, such as shallow prairie lakes where recruitment might occur within many regions of highly interconnected main lakes [14,20].
Efficient removal of adult carp was another key to our success. In both our two case studies this involved several factors. One very important factor was our being able to develop good population estimates and couple them with an understanding of how much ecological damage different densities of carp cause in the glacial lakes so that removal targets could be established 100 kg/ha [6]. A second factor contributing to our success was our ability to develop a good understanding of adult carp distributions and movement patterns through the use of radio-tagged Judas carp. Not only did radio-tagged Judas carp provide essential information on where and how adult carp were aggregating under the ice in the winter [29], but also how and when they found food in baited box-nets and likewise how they could be specifically targeted for removal using baited traps [32,33]. Judas fish were also very helpful for locating and targeting carp spawning areas in the spring [6]. A third factor was our development of several different removal techniques that could be applied at different times of year to different types of carp aggregations. Notably, carp removal became progressively more difficult as their numbers declined so under-ice netting which worked very well initially [29], became less useful as carp densities dropped and required that we remove carp using food-induced aggregations [33] as well as spawning movements, the latter of which are unfortunately not highly predictable [27]. Boat electrofishing did not work well for removal because it was labor intensive although it proved to be a reliable method for rapidly and relatively accurately assessing carp abundance and biomass [27]. Finally, monitoring adult population size and distribution during removal efforts was essential. While boat electrofishing worked well in deeper navigable waters, it does not work well in shallow areas and it is possible the environmental DNA (eDNA) could supplement it as it has the advantage of being rapid and easy to use at almost any location and time [34]. Tracking sexually active adult carp could also be improved by the simultaneous deployment of sex pheromone measurement [32] or archival tag receivers. The success of commercial fishing schemes might also be improved with highly targeted approaches which do not rely on selling fish at market price, but reducing propagule pressure for a fee [41]. There is little doubt that all of our monitoring and removal techniques could be improved to drive an even more effective integrated control program for this species.
Several factors likely played into why we were able to eliminate carp recruitment in source ponds and are relevant to how this approach might be pursued by others in other locations. Our reliance on native egg and larval micro-predators was critical. There is little doubt, based on several laboratory and field studies [16,18,42], that bluegill sunfish, a voracious and agile predator with a small gape but high oxygen demand, played a key role eliminating any eggs and surviving larvae in restored aerated ponds and marshes. Nevertheless, several ponds (ex. Markham Pond) had few bluegill sunfish but many bullhead catfish, suggesting that they too, might have been important predators on larval and juvenile carp. A laboratory experiment also supports this possibility [18]. Likely, other species of native fish also serve this function in other locations and habitats. In addition, our efforts to control carp recruitment were likely strongly enhanced by our ability to both block adults from entering these ponds and to then remove the small number of resident adults they contained (i.e., Markham and Casey Ponds). Integrated, multi-faceted responses to invasive species control such as this are warranted. Especially remarkable about our study was how effectively our relatively low-cost strategy using native micro-predators worked: no carp recruitment was measured after winter aeration was installed in any pond. Carp lay their eggs on aquatic vegetation which also serves as habitat for larvae and young [18], and they are extremely vulnerable. In contrast, efforts to control carp by creating fishless wetlands using poisons (rotenone) and then protecting them with barriers [9] may, if not conducted with great care, enhance the long-term success of adults, because if migratory carp do manage to enter these systems (flood, power failure to electric barrier), reproductive success is all but assured. It thus makes sense that if possible (not all systems can support fish year-around) to manage native fishes together with carp, as we did in Lake Casey and Markham Pond. Notably, all carp nursery ponds restored by this study had been previously neglected and had poor water quality because of nutrient loading (poor water clarity, low oxygen, few resident fish), so their restoration was welcomed for its own sake. While it appears from our studies that most juvenile carp generally do not leave nursey ponds until they at least a year old [14,21,43], this issue remains an unknown and warrants study because it could be exploited. In sum, restoration of shallow degraded ponds so that they can support native fishes is clearly a concept worth exploring to control carp.
Our study also sheds new light on the life history of common carp and how it may have evolved to use source-sink dynamics. Like the large rivers in which carp evolved, the chains of lakes we studied are complex and possess large interconnected ponds/wetlands (which resemble flood plains) that are subject to fluctuating physical conditions (winter oxygen, flows and water levels) and thus not amenable to supporting stable populations of resident native micro-predators. It is fascinating that common carp have evolved a life history which simultaneously includes high fecundity, longevity and directed yet flexible movement strategies that allow adults to exploit peripheral wetland habitats which often lack micro-predators in the spring. Their ability to find these spawning/ nursery habitats is also remarkable, as is their ability to exhibit these migratory behaviors when appropriate (high flow years). Being long-lived, they only need to be successful on rare occasions. We hypothesize that these movement patterns may reflect a type of homing, or ability to remember and preferentially return to natal areas when appropriate (at times of high water), thus explaining how and why they select certain ponds and not others [27,37,40]. We are not aware of any other species that has coupled a source-sink life history with partial migration as well as homing [44], and it may do much to explain the invasiveness of this species in many regions.
Our study will hopefully provide guidance to others hoping to control carp. Our case studies emphasize the need to develop an understanding of the population dynamics of carp in local areas, especially the sources of young, and to appreciate the need that a long-term commitment is needed given the longevity of this species. We recommend the same three-step process we employed which includes assessment, control, and monitoring. However, the highly adaptive and flexible nature of the carp’s life history means that its population dynamics are likely to take different forms in different locations and different approaches to this control scheme may be required elsewhere. Nevertheless, targeting the production of young is always likely to be key. As shown by others [35], we too show that if improved water quality is the objective of carp removal, alum treatment may often be required after carp removal because nutrient input from eutrophic sediments may persist even in the absence of carp, but that if alum is then applied it can work well and in a sustainable manner. Added benefits of carp control include increased aquatic plant cover [39] and likely aquatic biodiversity, but we did not assess the later possibility.
Finally, a fascinating and important question is whether source-sink dynamics might commonly explain high densities of common carp and perhaps other invasive fish species, and thus be commonly exploitable in ways similar to those used by us. Many hundreds of species of fish employ migratory life histories which include occasional seasonal use of spawning areas and nurseries [44], and thus could also be considered to be employing a type of source-sink dynamics. In addition to common carp, several of these species are highly invasive including rainbow trout (Oncorhynchys mykiss), brown trout (Salmo trutta), and sea lamprey (Petromyzon marinus). Aside from the sea lamprey, which uses migratory pheromones to select spawning/nursery streams [45], all of these fish home so they have only a few sources. Indeed, recent efforts to control lake trout in western lakes by targeting embryos on their home spawning grounds are similar in concept [46]. Carp reproductive success seems to vary greatly with region and local ecosystem, perhaps because the presence of native micro-predators for their eggs/ larvae likely varies for several reasons including: native fish biodiversity (ex. Australia), flooding (river floodplains in Eurasia), and/or local/seasonal hypoxia (Midwestern lakes). This variation will make this species easier to control in some regions/ ecosystems than others. The question of what ecological factors determine the causes and sources of common carp recruitment in different ecosystems is thus one that must be actively explored, especially because it is now clear that common carp may be controlled once this question has been answered.
Author Contributions
Conceptualization: P.W.S.; methodology: P.G.B. and P.W.S.; formal analysis: P.W.S. and P.G.B.; writing—original draft: P.W.S.; writing—review and editing: P.W.S. and P.G.B.; project administration: P.W.S.; Funding: P.W.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by The Minnesota Environment and Natural Resources Trust Fund; The Riley Purgatory Bluff Creek Watershed District and The Ramsey Washington Metro Watershed District.
Acknowledgments
This study is the culmination of the work of many over a decade. The late Bill Oemichen kick-started this research with a personal gift of USD 10,000 to Sorensen with no strings attached. This paper is dedicated to him. Ray Norrgard from the MN DNR was also very encouraging and helped us get initial funding. Since that time, additional funding has been provided from many sources (see above) which have supported many (without promise of practical results) who have dedicated themselves to understanding carp biology and carp control including the following graduate students: Mario Travaline, Jake Osborne, Justin Silbernagel, Nate Banet, Reid Swanson, Justine Dauphinais, Joey Lechelt, Josh Poole; Undergraduate students: Keith Phillips; Technicians: Nathan Berg, Mary Haedrick Newman, Brett Miller; and Postdoctoral fellows: Przemek Bajer Haude Levesque, Hangkyo Lim, Chris Chizinski, Ratna Ghosal, Jessica Eichmiller. Bill Bartodziej, Matt Kocian, Paul Haik, Mike Casanova and Ken Wentzel were also very encouraging. Dave and Anne Florenzano supported and encouraged our early work on Lake Riley. Daryl Ellison from the MN DNR was also very helpful and pointed out the importance of winterkill to local fisheries while also helping us select the Riley Chain of Lakes for our study and providing the permits we needed. Sorensen’s visit with the Tasmanian Fisheries Service and the inspiration they offered in the mid-2000s was extremely influential and appreciated. Justine Dauphinais, Ray Newman, Jeff Whitty, and Bill Bartodziej all read initial drafts and provided feedback. Sorensen is deeply grateful to all of these people.
Conflicts of Interest
P.W.S. Sorensen has no conflict. P.G.B. Bajer is the founder and owner of Carp Solutions LLC, a company involved in the development of management strategies for invasive fish. These interests have been reviewed and managed by the University of Minnesota in accordance with its Conflict of Interest policies.
References
- Britton, J.R.; Cucherousset, J.; Davies, G.D.; Godard, M.J.; Copp, G.H. Non-native fishes and climate change: Predicting species responses to warming temperatures in a temperate region. Freshw. Biol. 2010, 55, 1130–1141. [Google Scholar] [CrossRef]
- Balon, E.K. Origin and domestication of the wild carp, Cyprinus carpio: From Roman gourmets to the swimming flowers. Aquaculture 1995, 129, 3–48. [Google Scholar] [CrossRef]
- Sorensen, P.W.; Bajer, P.G. Carp, common. In Encyclopedia of Biological Invasions; Simberloff, D., Rejmanek, M., Eds.; University of California Press: Berkeley, CA, USA, 2011. [Google Scholar]
- Kohlmann, K.; Kersten, P. Deeper insight into the origin and spread of European common carp (Cyprinus carpio carpio) based on mitochondrial D-loop sequence polymorphisms. Aquaculture 2013, 376, 97–104. [Google Scholar] [CrossRef]
- Funge-Smith, S. Review of the State of the World Fisheries Resources: Inland Fisheries; FAO Fisheries and Aquaculture Circular: Rome, Italy, 2018; 253p. [Google Scholar]
- Bajer, P.G.; Sullivan, G.; Sorensen, P.W. Effects of a rapidly increasing population of common carp on vegetative cover and waterfowl in a recently restored Midwestern shallow lake. Hydrobiologia 2009, 632, 235–245. [Google Scholar] [CrossRef]
- Weber, M.J.; Brown, M.L. Effects of Common Carp on Aquatic Ecosystems 80 Years after “Carp as a Dominant”: Ecological Insights for Fisheries Management. Rev. Fish. Sci. 2009, 17, 524–537. [Google Scholar] [CrossRef]
- Matsuzaki, S.S.; Usio, N.; Takamura, N.; Washitani, I. Contrasting impacts of invasive engineers on freshwater ecosystems: An experiment and meta-analysis. Oecologia 2008, 158, 673–686. [Google Scholar] [CrossRef]
- Verrill, C.R.; Berry, B.R., Jr. Effectiveness of a lake drawdown and electrical barrier for reducing common carp and bigmouth buffalo abudances. N. Am. J. Fish. Manag. 1995, 5, 137–141. [Google Scholar] [CrossRef]
- Meronek, T.G.; Bouchard, P.M.; Buckner, E.R.; Burri, T.M.; Demmerly, K.K.; Hatleli, D.C.; Klumb, R.A.; Schmidt, S.H.; Coble, D.W. A Review of Fish Control Projects. N. Am. J. Fish. Manag. 1996, 16, 63–74. [Google Scholar] [CrossRef]
- Roberts, J.; Tilzey, R. Controlling Carp: Options for Australia; CSIRO Press: Griffith, Australia, 1997; 141p. [Google Scholar]
- Weber, M.J.; Hennen, M.J.; Brown, M.L.; Lucchesi, D.O.; Sauver, T.R.S. Compensatory response of invasive common carp Cyprinus carpio to harvest. Fish. Res. 2016, 179, 168–178. [Google Scholar] [CrossRef]
- Balon, E.K. The common carp, Cyprinus carpio: Its wild origin, domestication in aquaculture, and selection as colored nishikigoi. Guelph Ichthyol. Rev. 1995, 3, 1–55. [Google Scholar]
- Bajer, P.G.; Cross, T.K.; Lechelt, J.D.; Chizinski, C.J.; Weber, M.J.; Sorensen, P.W. Across-ecoregion analysis suggests a hierarchy of ecological filters that regulate recruitment of a globally invasive fish. Divers. Distrib. 2015, 21, 500–510. [Google Scholar] [CrossRef] [Green Version]
- McCrimmon, H.R. Carp in Canada. In Bulletin Fisheries Research; Fisheries Research Board of Canada: Ottawa, ON, Canada, 1968; 165p. [Google Scholar]
- Bajer, P.G.; Sorensen, P.W. Recruitment and abundance of an invasive fish, the common carp, is driven by its propensity to invade and reproduce in basins that experience winter-time hypoxia in interconnected lakes. Biol. Invasions 2009, 12, 1101–1112. [Google Scholar] [CrossRef]
- Billard, R. Carp: Biology and Culture; Praxis Publishing: Chichester, UK, 1999. [Google Scholar]
- Silbernagel, J.J.; Sorensen, P.W. Direct Field and Laboratory Evidence that a Combination of Egg and Larval Predation Controls Recruitment of Invasive Common Carp in Many Lakes of the Upper Mississippi River Basin. Trans. Am. Fish. Soc. 2013, 142, 1134–1140. [Google Scholar] [CrossRef]
- Koehn, J.D. Carp (Cyprinus carpio) as a powerful invader in Australian waterways. Freshw. Biol. 2004, 49, 882–894. [Google Scholar] [CrossRef]
- Lechelt, J.; Bajer, P. Elucidating the mechanism underlying the productivity-recruitment hypothesis in the invasive common carp. Aquat. Invasions 2016, 11, 469–482. [Google Scholar] [CrossRef]
- Dauphinais, J.D.; Miller, L.M.; Swanson, R.G.; Sorensen, P.W. Source-sink dynamics explain the distribution and persistence of an invasive population of common carp across a model Midwestern watershed. Biol. Invasions 2018, 20, 1961–1976. [Google Scholar] [CrossRef]
- Vilizzi, L.; Tarkan, A.S.; Copp, G.S. Experimental evidence from causal criteria analysis for the effects of common carp Cyprinus carpio on freshwater ecosystems: A global perspective. Rev. Fish. Sci. Aquac. 2015, 23, 253–290. [Google Scholar] [CrossRef]
- Bajer, P.G.; Beck, M.W.; Cross, T.K.; Koch, J.; Bartodziej, B.; Sorensen, P.W. Biological invasion by a benthivorous fish reduced the cover and species richness of aquatic plants in most lakes of a large North American ecoregion. Glob. Chang. Biol. 2016, 22, 3937–3947. [Google Scholar] [CrossRef] [PubMed]
- Pulliam, H.R. Sources, Sinks, and Population Regulation. Am. Nat. 1988, 132, 652–661. [Google Scholar] [CrossRef]
- Travis, J.M.J.; Park, K. Spatial structure and the control of invasive alien species. Anim. Conserv. 2004, 7, 321–330. [Google Scholar] [CrossRef]
- Bajer, P.G.; Chizinski, C.J.; Silbernagel, J.J.; Sorensen, P.W. Variation in native micro-predator abundance explains recruitment of a mobile invasive fish, the common carp, in a naturally unstable environment. Biol. Invasions 2012, 14, 1919–1929. [Google Scholar] [CrossRef] [Green Version]
- Chizinski, C.J.; Bajer, P.G.; Headrick, M.E.; Sorensen, P.W. Different migratory strategies of invasive Common Carp and native Northern Pike in the American Midwest suggest an opportunity for selective management strategies. N. Am. J. Fish. Manag. 2016, 36, 769–779. [Google Scholar] [CrossRef]
- Bajer, P.G.; Sorensen, P.W. Using boat electrofishing to estimate the abundance of invasive Common Carp in small Midwestern lakes. N. Am. J. Fish. Manag. 2012, 32, 817–822. [Google Scholar] [CrossRef]
- Bajer, P.G.; Chizinski, C.J.; Sorensen, P.W. Using the Judas technique to locate and remove wintertime aggregations of invasive common carp. Fish. Manag. Ecol. 2011, 18, 497–505. [Google Scholar] [CrossRef] [Green Version]
- Osborne, J. Distribution, Abundance and Overwinter Survival of Young-of-the-Year Common Carp in a Midwestern Watershed. Master’s Thesis, University of Minnesota, Minneapolis, MN, USA, 2012, (unpublished). [Google Scholar]
- Brown, P.; Green, C.; Sivakumaran, K.P.; Giles, A.; Stoessel, D. Validating otolith annuli for use in age-determination of carp (Cyprinus carpio L.) from Victoria, Australia. Trans. Am. Fish. Soc. 2004, 133, 190–196. [Google Scholar] [CrossRef]
- Ghosal, R.; Eichmiller, J.J.; Witthuhn, B.A.; Sorensen, P.W. Attracting Common Carp to a bait site with food reveals strong positive relationships between fish density, feeding activity, environmental DNA, and sex pheromone release that could be used in invasive fish management. Ecol. Evol. 2018, 8, 6714–6727. [Google Scholar] [CrossRef]
- Bajer, P.G.; Lim, H.; Travaline, M.J.; Miller, B.D.; Sorensen, P.W. Cognitive aspects of food searching behavior in free-ranging wild Common Carp. Environ. Boil. Fishes 2010, 88, 295–300. [Google Scholar] [CrossRef]
- Eichmiller, J.J.; Bajer, P.G.; Sorensen, P.W. The Relationship between the Distribution of Common Carp and Their Environmental DNA in a Small Lake. PLoS ONE 2014, 9, e112611. [Google Scholar] [CrossRef] [Green Version]
- Huser, B.J.; Bajer, P.G.; Chizinski, C.J.; Sorensen, P.W. Effects of common carp (Cyprinus carpio) on sediment mixing depth and mobile phosphorus mass in the active sediment layer of a shallow lake. Hydrobiologia 2015, 763, 23–33. [Google Scholar] [CrossRef]
- Bajer, P.G.; Sorensen, P.W. Effects of common carp on phosphorus concentrations, water clarity, and vegetation density: A whole system experiment in a thermally stratified lake. Hydrobiologia 2014, 746, 303–311. [Google Scholar] [CrossRef]
- Bajer, P.G.; Parker, J.E.; Cross, T.K.; Venturelli, P.A.; Sorensen, P.W. Partial migration to seasonally-unstable habitat facilitates biological invasions in a predator-dominated system. Oikos 2015, 124, 1520–1526. [Google Scholar] [CrossRef]
- Knopik, J.M.; Newman, R.M. Transplanting aquatic macrophytes to restore the littoral community of a eutrophic lake after the removal of common carp. Lake Reserv. Manag. 2018, 34, 365–375. [Google Scholar] [CrossRef]
- Dunne, M.A.; Newman, R.M. Effect of light on macrophyte sprouting and assessment of viable seedbank to predict community composition. J. Aquat. Plant Manag. 2019, 57, 90–98. [Google Scholar]
- Banet, N. Partial Migration, Homing, Diel Activity, and Distribution of Adult Common Carp across a Large, Model Watershed in the North American Midwest. Master’s Thesis, University of Minnesota, Saint Paul, MN, USA, 2016, (unpublished). [Google Scholar]
- Colvin, M.E.; Pierce, C.L.; Stewart, T.W.; Grummer, S.E. Strategies to control a common carp population by pulsed commercial harvest. N. Am. J. Fish. Manag. 2012, 32, 1251–1264. [Google Scholar] [CrossRef] [Green Version]
- Poole, J.R.; Bajer, P.G. A small native predator reduces reproductive success of a large invasive fish as revealed by whole-lake experiments. PLoS ONE 2019, 14, e0214009. [Google Scholar] [CrossRef] [Green Version]
- Lechelt, J.; Kocian, M.; Bajer, P. Low downstream dispersal of young-of-year common carp from marshes into lakes in the Upper Mississippi Region and its implications for integrated pest management strategies. Manag. Biol. Invasions 2017, 8, 485–495. [Google Scholar] [CrossRef] [Green Version]
- Chapman, B.B.; Hulthén, K.; Brodersen, J.; Nilsson, P.A.; Skov, C.; Hansson, L.-A.; Brönmark, C. Partial migration in fishes: Causes and consequences. J. Fish Biol. 2012, 81, 456–478. [Google Scholar] [CrossRef] [Green Version]
- Sorensen, P.W.; Fine, J.M.; Dvornikovs, V.; Jeffrey, C.S.; Shao, F.; Wang, J.; Vrieze, L.A.; Anderson, K.R.; Hoye, T.R. Mixture of new sulfated steroids functions as a migratory pheromone in the sea lamprey. Nat. Chem. Biol. 2005, 1, 324–328. [Google Scholar] [CrossRef]
- Koel, T.M.; Arnold, J.L.; Bigelow, P.E.; Brenden, T.O.; Davis, J.D.; Detjens, C.R.; Doepke, P.D.; Ertel, B.D.; Glassic, H.C.; Gresswell, R.E.; et al. Yellowstone Lake ecosystem restoration: A case study for invasive fish management. Fishes 2020, 5, 18. [Google Scholar] [CrossRef]
Figure 1.
Schematic of the source-sink process hypothesized to determine the abundance of common carp in many Midwestern chains of lakes. The dashed arrows show that most, but not all, adults move between deep lakes and peripheral shallow water bodies to spawn and then return to lakes after the spawning season. Some adults may also occasionally overwinter in shallow waterbodies. Being long-lived, this flexible movement strategy, also known as partial migration, seems to work to the carp’s advantage, especially in dynamic and complex temperate ecosystems that experience occasional flooding and winterkill.
Figure 1.
Schematic of the source-sink process hypothesized to determine the abundance of common carp in many Midwestern chains of lakes. The dashed arrows show that most, but not all, adults move between deep lakes and peripheral shallow water bodies to spawn and then return to lakes after the spawning season. Some adults may also occasionally overwinter in shallow waterbodies. Being long-lived, this flexible movement strategy, also known as partial migration, seems to work to the carp’s advantage, especially in dynamic and complex temperate ecosystems that experience occasional flooding and winterkill.

Figure 2.
The Phalen Chain of Lakes.

Figure 3.
Age structure of adult carp collected from lakes Kohlman, Gervais and Keller. Data from [21] which were re-analyzed for this figure to show their percent contribution to the population. Fish were captured and aged in 2011.
Figure 3.
Age structure of adult carp collected from lakes Kohlman, Gervais and Keller. Data from [21] which were re-analyzed for this figure to show their percent contribution to the population. Fish were captured and aged in 2011.

Figure 4.
Average number of adult carp caught electrofishing per hour (CPUE) across lakes Kohlman, Gervais and Keller (data are combined and averaged). The study started in 2009.
Figure 4.
Average number of adult carp caught electrofishing per hour (CPUE) across lakes Kohlman, Gervais and Keller (data are combined and averaged). The study started in 2009.
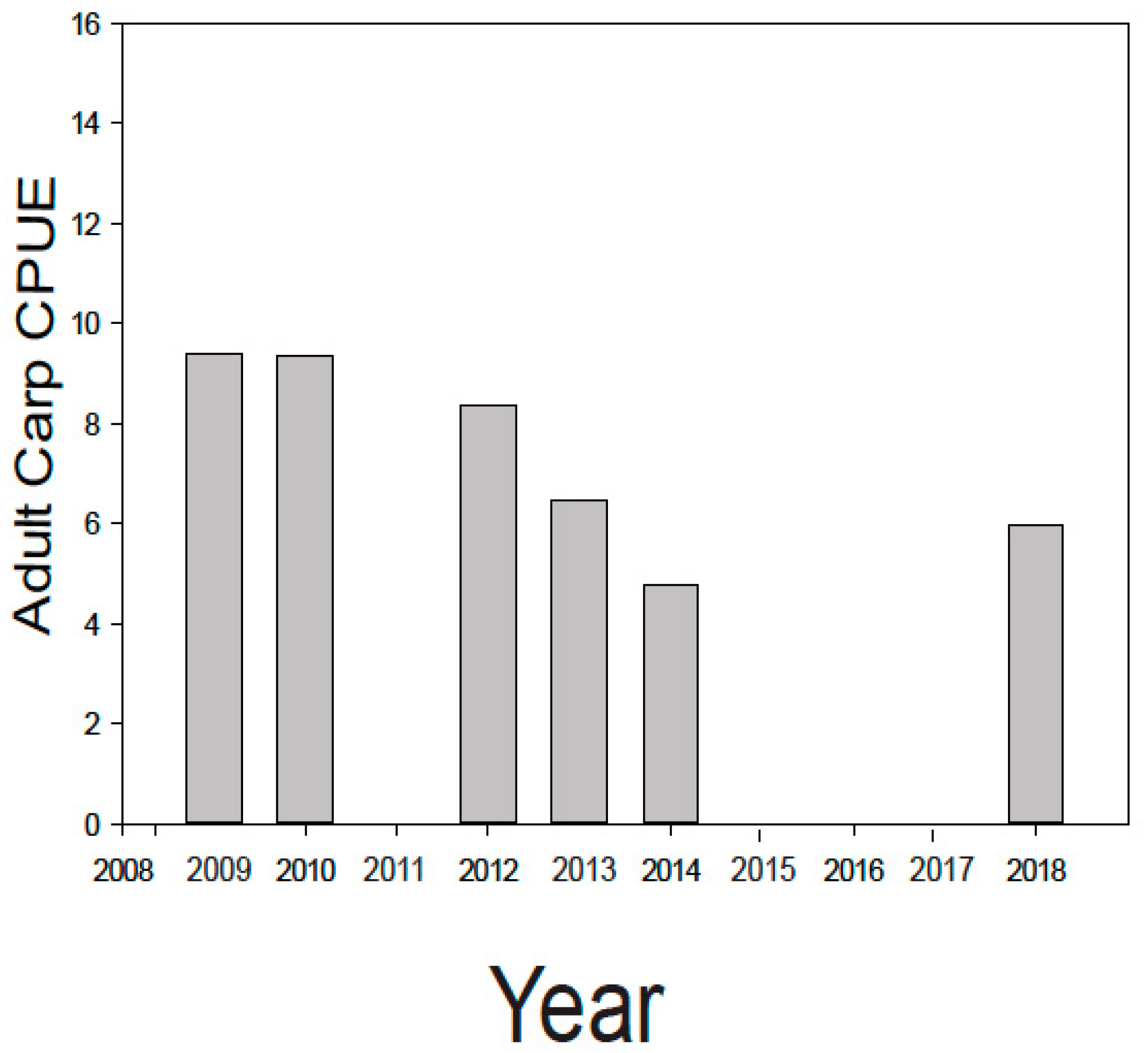
Figure 5.
Summer (June–August) epilimnion water quality values (TP, or Total Phosphorous, and Secchi Depth (clarity in m)) for lakes Kohlman, Gervais and Keller. The blue dotted line shows when the alum treatment occurred [35] while the dotted red lines show years of carp removal in the Kohlman- Gervais-Keller system of lakes (see text).
Figure 5.
Summer (June–August) epilimnion water quality values (TP, or Total Phosphorous, and Secchi Depth (clarity in m)) for lakes Kohlman, Gervais and Keller. The blue dotted line shows when the alum treatment occurred [35] while the dotted red lines show years of carp removal in the Kohlman- Gervais-Keller system of lakes (see text).

Figure 6.
The Riley Chain of Lakes.
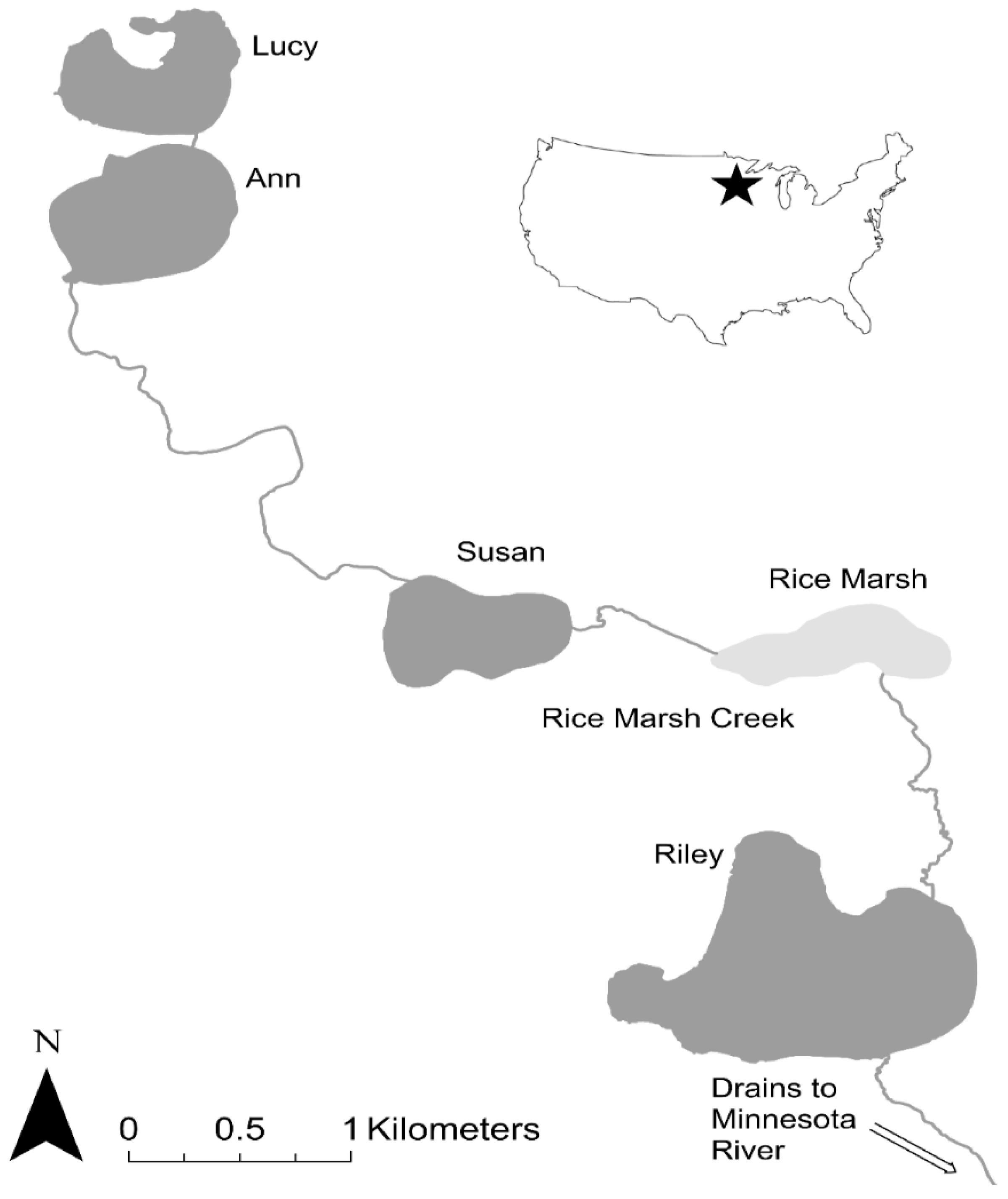
Figure 7.
Age structure of adult carp collected from lakes: (a) Susan, and (b) Riley. Fish were aged in 2009. The Lake Susan data are published [16].
Figure 7.
Age structure of adult carp collected from lakes: (a) Susan, and (b) Riley. Fish were aged in 2009. The Lake Susan data are published [16].

Figure 8.
Adult carp CPUE (carp/h) as monitored by boat electrofishing in: (a) Lake Susan, and (b) Lake Riley.
Figure 8.
Adult carp CPUE (carp/h) as monitored by boat electrofishing in: (a) Lake Susan, and (b) Lake Riley.
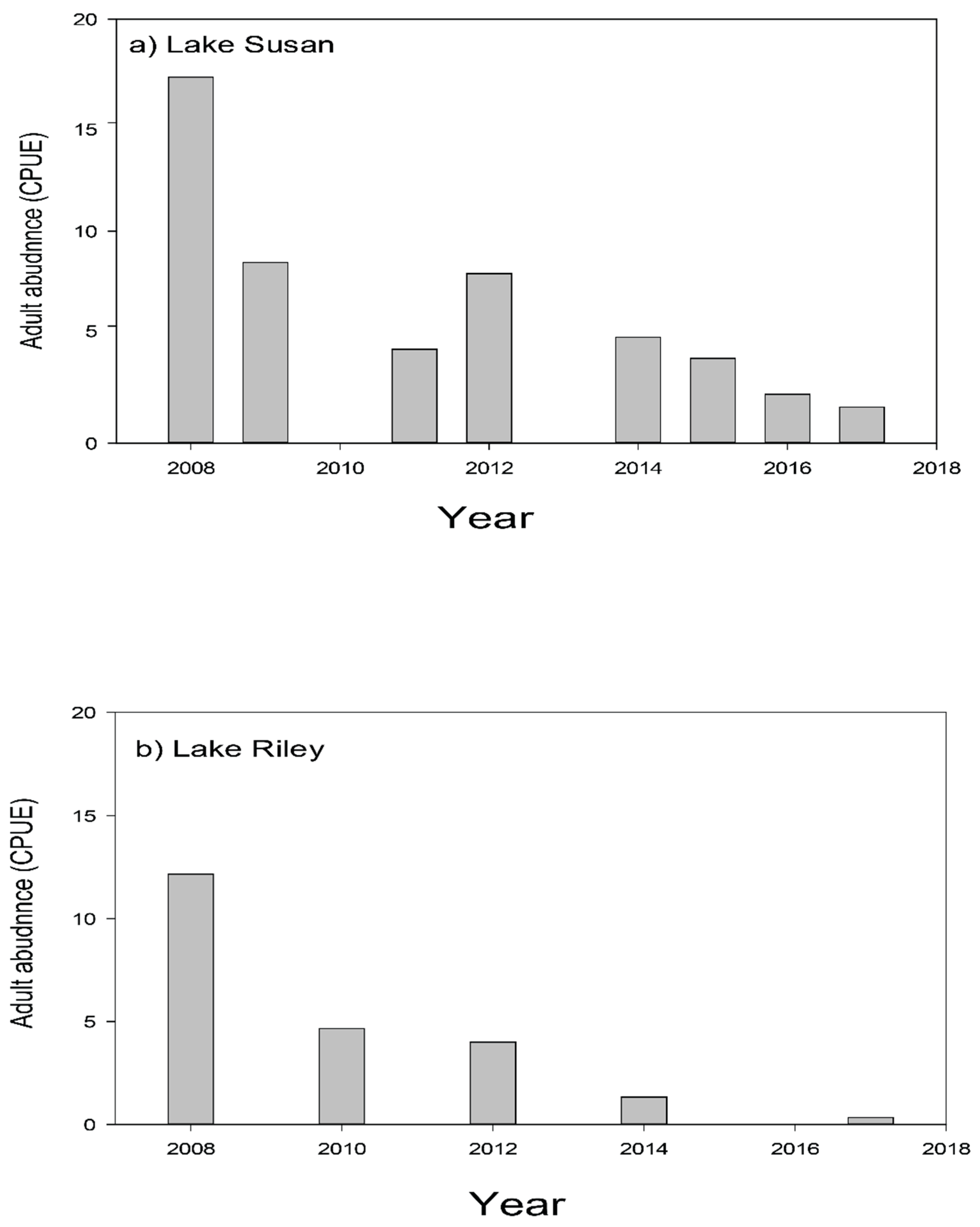
Figure 9.
Summer (June–August) epilimnion water quality values (TP or Total Phosphorous, and Secchi depth (clarity)) for lakes Susan and Riley. The blue dotted line shows when the alum treatment occurred while the dotted red lines show years of carp removal in this system of lakes (see text).
Figure 9.
Summer (June–August) epilimnion water quality values (TP or Total Phosphorous, and Secchi depth (clarity)) for lakes Susan and Riley. The blue dotted line shows when the alum treatment occurred while the dotted red lines show years of carp removal in this system of lakes (see text).

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Lakes and ponds in the Phalen Chain of Lakes and their carp as initially assessed in 2009–2010. CPUE (catch per unit effort) for adults is the number of carp caught per hour by electrofishing, while CPUE for juveniles is the number of carp caught per trap-net (see text). Emboldened values are ecologically problematic (i.e., >100 kg/ha). YOY = young-of–the-year carp.
Table 1.
Lakes and ponds in the Phalen Chain of Lakes and their carp as initially assessed in 2009–2010. CPUE (catch per unit effort) for adults is the number of carp caught per hour by electrofishing, while CPUE for juveniles is the number of carp caught per trap-net (see text). Emboldened values are ecologically problematic (i.e., >100 kg/ha). YOY = young-of–the-year carp.
| Lake | Size (ha) | Max Depth (m) | Adult Carp (CPUE) | Adult Carp (#) | Adult Carp Density | YOY Carp (CPUE) | Notes |
|---|---|---|---|---|---|---|---|
| Casey | 4.7 | 6.1 | 8.5 | 2585 | n.a. | 6.2 b | Winterkills |
| Markham | 7.0 | 2.0 | 8.2 | 211 | n.a. | 104 b | Winterkills |
| Willow | 21.1 | 2.5 | 0 | 0 | n.a. | 173 b | Winterkills |
| Kohlman | 29.9 | 2.7 | 11.2 a | 1668 a | 177 kg/ha c | 0 b | - |
| Gervais | 94.5 | 12.5 | 7.2 a | 3499 a | 177 kg/ha c | 0 b | - |
| G Mill Pond | 3.0 | 4.0 | n.s. | n.s. | n.s. | n.s. | - |
| Keller | 29.0 | 2.4 | 14.7 | 2103 | 177 kg/ha c | 0 b | - |
| Round | 14.3 | 2.5 | n.s. | n.s. | n.s. | n.s. | - |
| Phalen | 79.6 | 53 | 6.0 | 2504 | 106 kg/ha d | 0 b |
a. Previously published [26]. b. Previously published [21]. c. Biomass data show the average combined biomass/ha of carp in Kohlman, Gervais and Keller lakes, reflecting our finding that carp readily moved between these three lakes and functioned a single population (Table 2). d from 2018. n.s. = not sampled.
Table 2.
The absolute number and percentage of total detections of 50 adult carp originally radio-tagged in lakes Gervais, Keller, Kohlman, and Phalen found in 2009 by lake and by season. Emboldened lake names in the left column denote the lake where the carp were originally captured.
Table 2.
The absolute number and percentage of total detections of 50 adult carp originally radio-tagged in lakes Gervais, Keller, Kohlman, and Phalen found in 2009 by lake and by season. Emboldened lake names in the left column denote the lake where the carp were originally captured.
| Origin Lake | Summer 2009 | Fall 2009 | Winter 2009–2010 | Spring 2010 | Summer 2010 |
|---|---|---|---|---|---|
| Gervais Lake (Lake Where Detected) | |||||
| Kohlman | 1 (2%) | 0 (0%) | 19 (26%) | 6 (18%) | 4 (15%) |
| Gervais | 35 (71%) | 36 (61%) | 18 (25%) | 16 (47%) | 21 (78%) |
| Keller | 13 (27%) | 23 (39%) | 35 (49%) | 12 (35%) | 2 (7%) |
| Phalen | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Lake Keller | |||||
| Kohlman | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Gervais | 1 (9%) | 9 (11%) | 3 (33%) | 9 (17%) | 5 (8%) |
| Keller | 110 (91%) | 73 (89%) | 6 (67%) | 43 (83%) | 55 (92%) |
| Phalen | 0 (0) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Kohlman Lake | |||||
| Kohlman | 67 (70%) | 56 (84%) | 5 (83%) | 20 (77%) | 36 (68%) |
| Gervais | 21 (22%) | 11 (16%) | 1 (17%) | 5 (19%) | 7 (13%) |
| Keller | 8 (8%) | 0 (0%) | 0 (0%) | 1 (4%) | 10 (19%) |
| Phalen | 1 (1%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Lake Phalen | |||||
| Kohlman | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Gervais | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) | 0 (0%) |
| Keller | 1 (1%) | 26 (30%) | 4 (21%) | 3 (14%) | 12 (20%) |
| Phalen | 82 (99%) | 62 (70%) | 15 (79%) | 18 (86%) | 49 (80%) |
Table 3.
Common carp control efforts and results in the Lake Phalen Chain of Lakes (previously published data is referenced, the rest is from various data reports to the RWMWD).
Table 3.
Common carp control efforts and results in the Lake Phalen Chain of Lakes (previously published data is referenced, the rest is from various data reports to the RWMWD).
| Date | Removal Method | # Carp Caught | # Carp Removed |
|---|---|---|---|
| Winter 2010–11 | Gervais Winter Seine-1 a | 3537 b | 100 |
| Winter 2010–11 | Gervais Winter Seine-2 | 1509 | 1505 |
| Winter 2010–11 | Gervais Winter Seine-3 | 930 | 732 |
| Winter 2011–12 | Gervais Winter Seine | 73 | 71 |
| Fall 2012 | Kohlman Boxnet | 630 | 628 |
| Winter 2012–13 | Gervais Winter Seine | 827 | 825 |
| Spring 2011–2013 | Spawning Block/Trap | 381 | 351 |
| Summer 2013 | Keller Electrofishing | 8 | 8 |
| Summer 2013 | Gervais Electrofishing | 97 | 97 |
| Summer 2013 | Kohlman Electrofishing | 5 | 5 |
| Fall 2013 | Keller Boxnet | 446 | 446 |
| Fall 2013 | Gervais Boxnet | 192 | 192 |
| Summer 2013 | Drawdown Casey, Markham | - | 100+ |
| Summer 2014 | Drawdown Casey, Markham | - | 100+ |
| Winter 2014–2015 | Aerate Casey, Markham | - | - |
| Fall 2015 | Gervais Boxnet | 302 | 302 |
| Fall 2016 | Gervais Boxnet * | 324 | 324 * |
a: The lake was seined three times in 2011 (1, 2, 3). b: Previously published [29]. * By the fall of 2016, a grand total of 5926 adult carp had been removed of the 7270 estimated present in lakes Kohlman, Gervais, and Keller (81%). The carp biomass was then 34 kg/ha across the 3 lakes, lakes Kohlman, Gervais and Keller (when combined and averaged).
Table 4.
Average number of Young-of-Year (YOY) carp caught per trap-net (CPUE) in the Phalen Chain of Lakes.
Table 4.
Average number of Young-of-Year (YOY) carp caught per trap-net (CPUE) in the Phalen Chain of Lakes.
| Year | Lakes | ||||||
|---|---|---|---|---|---|---|---|
| - | Casey | Markham | Willow | Kohlman | Gervais | Keller | Phalen |
| 1999 | 0 D | 0 | - | 0 D | 0 D | 0 D | 0 D |
| 2000 | - | - | - | - | - | - | - |
| 2001 | 25 D | - | - | - | - | - | - |
| 2002 | - | - | - | - | - | - | - |
| 2003 | - | - | - | - | - | - | - |
| 2004 | - | - | - | 0.33 D | - | - | 0 D |
| 2005 | - | - | - | 1.3 D | 0.56 D | 0.5 D | - |
| 2006 | - | - | - | - | - | - | - |
| 2007 | 28 D | - | - | - | - | 0 D | - |
| 2008 | - | - | - | - | - | - | - |
| 2009 S | 6.2 * | 104 * | 173 * | 0 * | 0 * | 0 * | 0 * |
| 2010 | 3.8 | 47 | 5.4 W | 0 | 0 | - | - |
| 2011 | 2 | 0.8 | 0 | 0 | 0 | - | - |
| 2012 | 8 | 0.7 | 0 | 0 | 0 | - | - |
| 2013 | 0 | 68.8 DD | 0 | 0 | 0 | - | - |
| 2014 | -DD | -DD | - | - | - | - | - |
| 2015 | 0 | 0 | - | 1 | - | - | 0 |
| 2016 | 0 | 0 | - | 0 | - | - | - |
| 2017 | - | - | - | 0 | - | - | - |
| 2018 | 0 | 0 | - | 0 | - | 0 | 0 |
Table 5.
Lakes in the Riley Chain of Lakes and their carp in 2008–2009. Data on carp densities are described in the text (emboldened values are for adult densities that are ecologically problematic).
Table 5.
Lakes in the Riley Chain of Lakes and their carp in 2008–2009. Data on carp densities are described in the text (emboldened values are for adult densities that are ecologically problematic).
| Lake | Size (ha) | Max Depth(m) | Adult Carp (CPUE) | Adult Carp (#) | Adult Carp (kg/ha) | YOY Carp (CPUE) | Notes |
|---|---|---|---|---|---|---|---|
| Lucy a | 34.6 | 6.0 | 8.15 b | 808 c | 70 kg/ha d | n.s. | winterkills |
| Ann | 44.1 | 13.7 | 3.57 e | 662 c | 66 kg/ha d | 0 | - |
| Susan | 35.1 | 5.1 | 17.26 b | 4181 c | 307/kg/h e | 0.4 | aerated |
| RiceMarshL | 40.0 | 3.0 | 3.0 | 178 c | 16 kg/h | 0 | winterkills |
| Riley | 118.8 | 14.7 | 12.16 b | 6419 c | 176 kg/ha d | 0 | - |
a: Lake Lucy winterkilled in 2012 and most fish died and population temporarily dropped to 20 kg/ha, before returning to 60–70 kg/ha seemingly because of adult immigration. b: Boat electrofishing 2008 and 2010 (Lucy), some of these data are published [28,29]. c: Published, population size extrapolated from mark-recapture study [28]. d: Biomass densities extrapolated from population size [28] and fish size. e: Previously reported [36].
Table 6.
The number of adult carp radio-tagged in lakes Susan, Rice Marsh, and Riley and their locations in these lakes by season (% = portion of the population they represent; some of these data have been published [37]). Lakes were carp were captured is emboldened.
Table 6.
The number of adult carp radio-tagged in lakes Susan, Rice Marsh, and Riley and their locations in these lakes by season (% = portion of the population they represent; some of these data have been published [37]). Lakes were carp were captured is emboldened.
| Original Lake | Spring 2006 | Summer 2006 | Winter 2006–7 | Summer 2007 | Winter 2007–8 |
|---|---|---|---|---|---|
| Lake Susan (Lake Where Detected) | |||||
| Susan | 16 (100%) | 10 (62%) | 15 (88%) | 24 (100%) | 26 (79%) |
| RiceM | - | 6 (38%) | 1 (6%) | 0 (0%) | 7 (21%) |
| Riley | - | 0 (0%) | 1 (6%) | 0 (0%) | 0 (0%) |
| Rice Marsh Lake | |||||
| Susan | - | 6 | 1 | 7 (0%) | 0 (0%) |
| RiceM | 15 (100%) | 15 (63%) | D | 0 (0%) | 11 (61%) |
| Riley | - | 3 | D | 7 (0%) | 7 (39%) |
| Lake Riley | |||||
| Susan | - | 0 (%) | 0 (0%) | 0 (0%) | 0 (0%) |
| RiceM | - | 3 (15%) | 0 (0%) | 0 (0%) | 1 (6%) |
| Riley | 19 (100%) | 16 (85%) | 18 (100%) | 24 (100%) | 17 (94%) |
D = died in Rice Marsh Lake due to winterkill.
Table 7.
Trap-net captures of YOY carp (CPUE) in the Riley Chain of Lakes.
| Year | Lakes (CPUE) | ||||
|---|---|---|---|---|---|
| - | Lucy | Ann | Susan | RiceMarsh | Riley |
| 1999 | - | - | - | - | 0 D |
| 2000 | 0 D | 0 D | - | - | - |
| 2001 | - | - | - | - | - |
| 2002 | - | - | - | - | - |
| 2003 | - | - | 0 D | - | - |
| 2004 | - | - | - | - | - |
| 2005 | - | - | - | - | 0 D |
| 2006 S | 0 D | 0 D | 0 | 0 | 0 |
| 2007 | - | - | 0 | T | 0 |
| 2008 | - | - | 0 | 0 | 0 |
| 2009 | 0 | 0 | 1.3 | 0 | 0 |
| 2010 | 0.8 | 0.6 | 1 | 1 | 1 |
| 2011 | 0.2 | 0 | 0.5 | 1 | 1 |
| 2012 | 0.2 | 0.2 | 0.8 | 1 | 0.8 |
| 2013 | 0.6 | 0.4 | 0.8 | 1 | 1 |
| 2014 | 0.6 | 0.4 | 0.6 | 0.8 | 0.8 |
| 2015 | - | - | 0 | 0 | 0 |
| 2016 | - | - | 0 W | 0.2 W | 0 |
| 2017 | - | 0 D | 0 D | - | - |
S = start of this study; D = MN DNR sampling in previous years; W = Watershed District data. T = hundreds of juvenile carp were seen leaving this pond during a winterkill event, 18 of which were later aged (see text).
Table 8.
Common carp control efforts and results in the Lake Riley Chain of Lakes.
| Date | Removal Method | # Carp Caught | # Carp Removed |
|---|---|---|---|
| Winter 2008–2009 | Susan Winter Seine | 3981 | 3425 |
| Spring 2009–2011 | Spawning block/trap | 242 | 242 a |
| Winter 2008–2009 | Riley Winter Seine | 4040 | 2940 |
| Winter 2009–2010 | Riley Winter Seine | 376 | 376 |
| Winter 2009–2010 | Riley Winter Seine | 2303 | 2303 |
| Spring 2009–2011 | Spawning block/trap | 21 | 21 b |
| Fall 2011 | Rice Marsh Lake aerated | - | - |
| Winter 2010–2011 | Lucy Winterkills | 630 | 500 c |
a: A total of 3667 adult carp were captured and removed from Lake Susan, leaving in theory 514 of the original 4181 (13%) or 40.8 kg/ha in 2011. b: A total of 5619 adult carp were captured and removed from Lake Riley, leaving, in theory, 800 of the original 6419 (12%) or 21 kg/ha in 2011. c: Lake Lucy was seined in 2010 and then winterkills in 2011, most carp appeared to die (we examined this lake as a side project intended to benefit lakes upstream of Lake Susan).
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Sorensen, P.W.; Bajer, P.G. Case Studies Demonstrate That Common Carp Can Be Sustainably Reduced by Exploiting Source-Sink Dynamics in Midwestern Lakes. Fishes 2020, 5, 36. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5040036
AMA Style
Sorensen PW, Bajer PG. Case Studies Demonstrate That Common Carp Can Be Sustainably Reduced by Exploiting Source-Sink Dynamics in Midwestern Lakes. Fishes. 2020; 5(4):36. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5040036
Chicago/Turabian StyleSorensen, Peter W., and Przemyslaw G. Bajer. 2020. "Case Studies Demonstrate That Common Carp Can Be Sustainably Reduced by Exploiting Source-Sink Dynamics in Midwestern Lakes" Fishes 5, no. 4: 36. https://0-doi-org.brum.beds.ac.uk/10.3390/fishes5040036