
Entomological Surveillance of Aedes Mosquitoes: Comparison of Different Collection Methods in an Endemic Area in RIO de Janeiro, Brazil

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
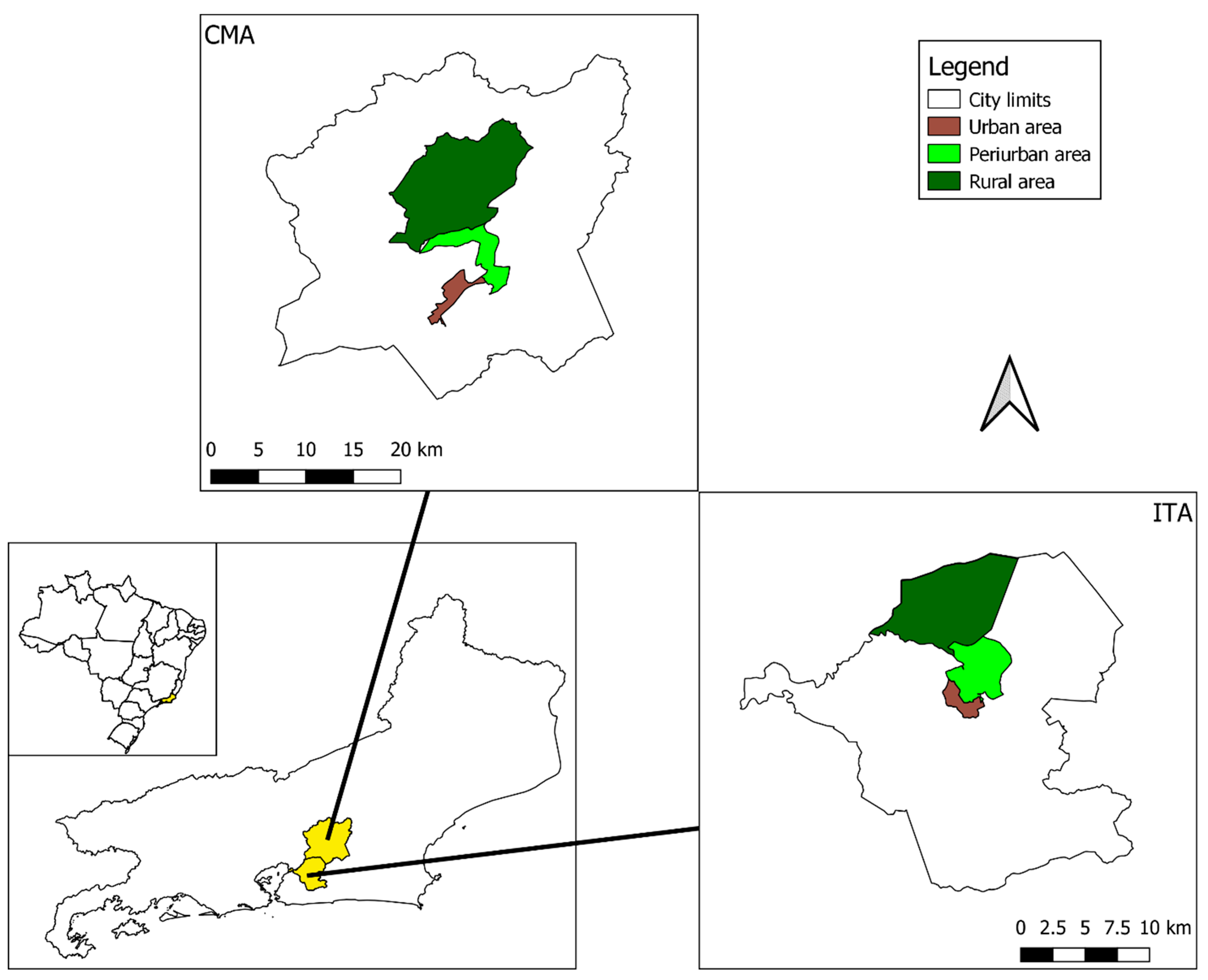
2.1. Study Area
2.2. Mosquito Sampling
2.3. Viral Detection
2.4. Data Analysis
2.4.1. Relationship between Positivity and Density Indices
2.4.2. Generalized Linear Mixed Models
3. Results
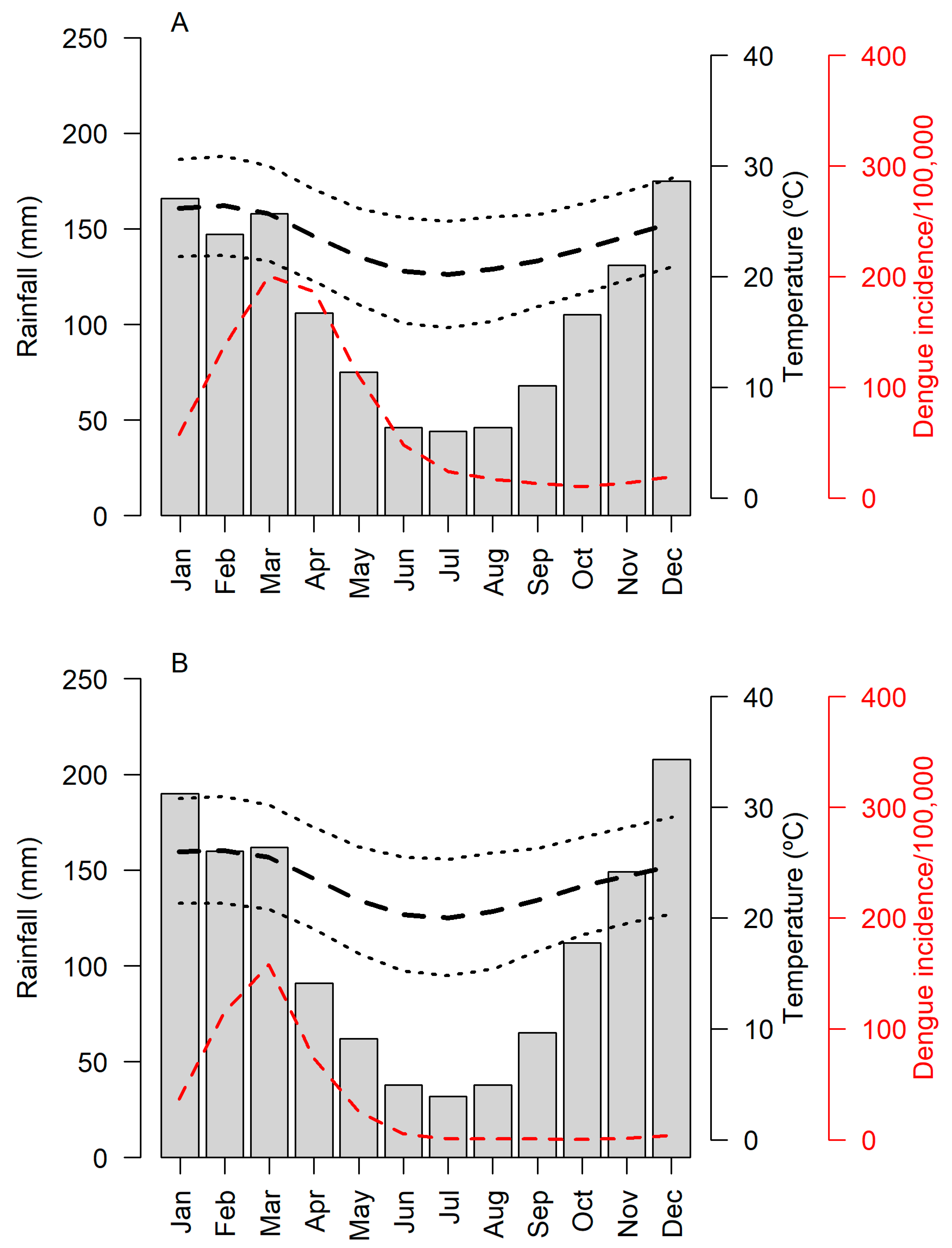
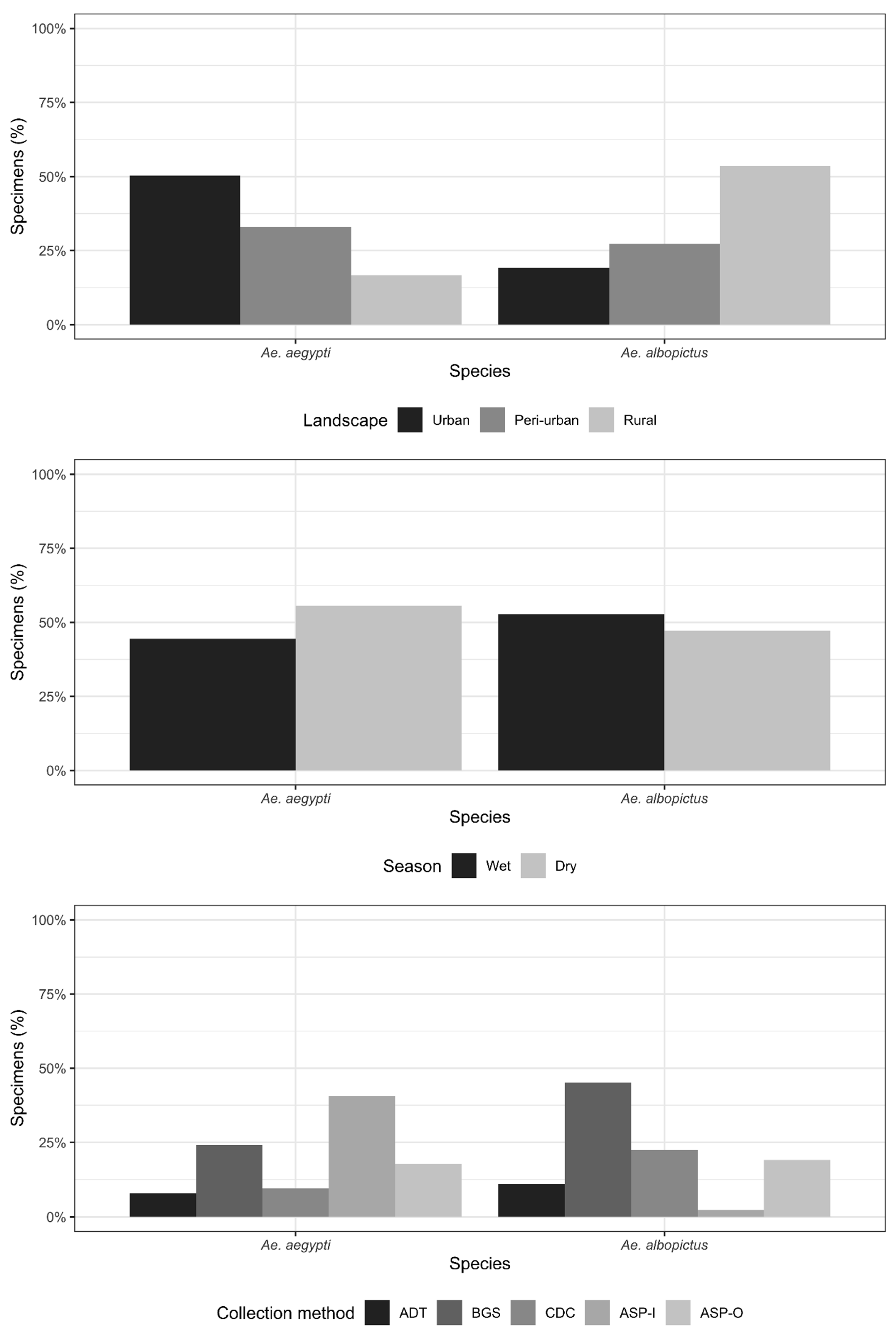
3.1. General Results and Viral Detection
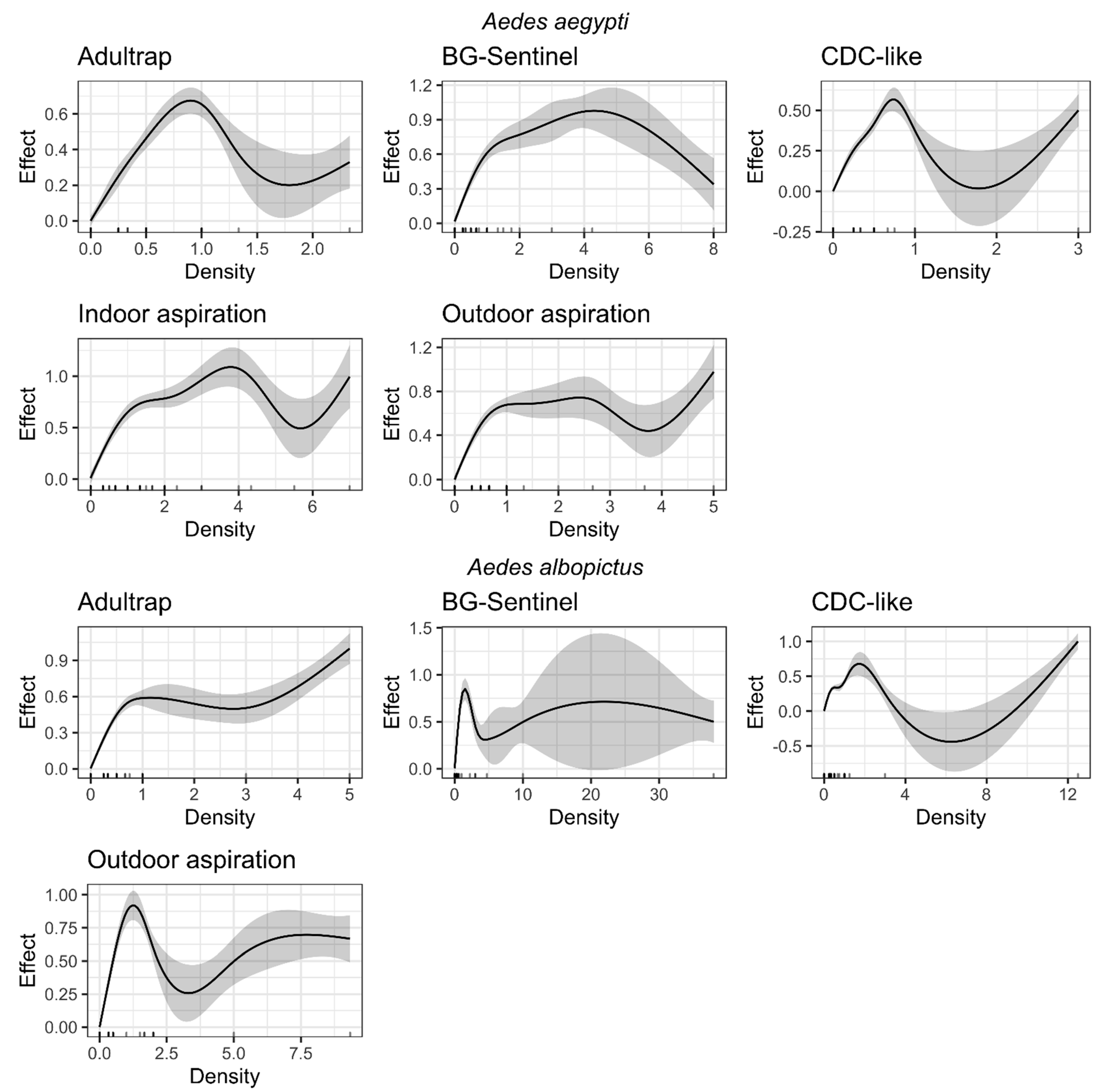
3.2. Relationship between Positivity and Density Indices
3.3. GLMM for Aedes Aegypti Abundance
3.4. GLMM for Aedes Albopictus Abundance
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Gubler, D.J. Dengue and dengue hemorrhagic fever. Clin. Microbiol. Rev. 1998, 11, 480–496. [Google Scholar] [CrossRef] [Green Version]
- Weaver, S.C. Arrival of Chikungunya Virus in the New World: Prospects for Spread and Impact on Public Health. PLoS Negl. Trop. Dis. 2014, 8, e2921. [Google Scholar] [CrossRef] [Green Version]
- Honório, N.A.; Câmara, D.C.P.; Calvet, G.A.; Brasil, P.; Honório, N.A.; Câmara, D.C.P.; Calvet, G.A.; Brasil, P. Chikungunya: An arbovirus infection in the process of establishment and expansion in Brazil. Cad. Saúde Pública 2015, 31, 906–908. [Google Scholar] [CrossRef] [Green Version]
- Musso, D.; Gubler, D.J. Zika Virus. Clin. Microbiol. Rev. 2016, 29, 487–524. [Google Scholar] [CrossRef] [Green Version]
- Lounibos, L.P. Invasions by insect vectors of human disease. Annu. Rev. Entomol. 2002, 47, 233–266. [Google Scholar] [CrossRef]
- Powell, J.R.; Tabachnick, W.J. History of domestication and spread of Aedes aegypti–a review. Mem. Inst. Oswaldo Cruz 2013, 18, 11–17. [Google Scholar] [CrossRef]
- Soper, F.L. The 1964 status of Aedes aegypti eradication and yellow fever in the Americas. Am. J. Trop. Med. Hyg. 1965, 14, 887–891. [Google Scholar] [CrossRef]
- Löwy, I. Representing and intervening in public health: Viruses, mosquitoes and Rockefeller Foundation experts in Brazil. Hist. Cienc. Saude-Manguinhos 1999, 5, 647–677. [Google Scholar] [CrossRef]
- Braga, I.A.; Valle, D. Aedes aegypti: Histórico do controle no Brasil. Epidemiol. Serv. Saúde 2007, 16, 113–118. [Google Scholar] [CrossRef]
- Forattini, O.P. Identificação de Aedes (Stegomyia) Albopictus (Skuse) no Brasil. Rev. Saúde Pública 1986, 20, 244–245. [Google Scholar] [CrossRef]
- Sprenger, D.; Wuithiranyagool, T. The discovery and distribution of Aedes albopictus in Harris County, Texas. J. Am. Mosq. Control. Assoc. 1986, 2, 217–219. [Google Scholar]
- Braga, I.A.; Valle, D. Aedes aegypti: Vigilância, monitoramento da resistência e alternativas de controle no Brasil. Epidemiol. Serv. Saúde 2007, 16, 295–302. [Google Scholar] [CrossRef]
- Codeço, C.T.; Lima, A.W.S.; Araújo, S.C.; Lima, J.B.P.; Maciel-de-Freitas, R.; Honório, N.A.; Galardo, A.K.R.; Braga, I.A.; Coelho, G.E.; Valle, D. Surveillance of Aedes aegypti: Comparison of House Index with Four Alternative Traps. PLoS Negl. Trop. Dis. 2015, 9, e0003475. [Google Scholar] [CrossRef]
- Kuno, G. Review of the factors modulating dengue transmission. Epidemiol. Rev. 1995, 17, 321–335. [Google Scholar] [CrossRef]
- Tun-Lin, W.; Kay, B.H.; Barnes, A.; Forsyth, S. Critical examination of Aedes aegypti indices: Correlations with abundance. Am. J. Trop. Med. Hyg. 1996, 54, 543–547. [Google Scholar] [CrossRef]
- Nathan, M.B.; Focks, D.A.; Kroeger, A. Pupal/demographic surveys to inform dengue-vector control. Ann. Trop. Med. Parasitol. 2006, 100, S1–S3. [Google Scholar] [CrossRef] [Green Version]
- Morrison, A.C.; Zielinski-Gutierrez, E.; Scott, T.W.; Rosenberg, R. Defining Challenges and Proposing Solutions for Control of the Virus Vector Aedes aegypti. PLoS Med. 2008, 5, e68. [Google Scholar] [CrossRef] [Green Version]
- Sivagnaname, N.; Gunasekaran, K. Need for an efficient adult trap for the surveillance of dengue vectors. Indian J. Med. Res. 2012, 136, 739–749. [Google Scholar]
- Honório, N.A.; Castro, M.G.; de Barros, F.S.M.; de Magalhães, M.A.F.M.; Sabroza, P.C. The spatial distribution of Aedes aegypti and Aedes albopictus in a transition zone, Rio de Janeiro, Brazil. Cad. Saúde Pública 2009, 25, 1203–1214. [Google Scholar] [CrossRef]
- Honório, N.A.; Codeço, C.T.; Alves, F.C.; a Magalhães, M.F.M.; Lourenço-De-Oliveira, R. Temporal distribution of Aedes aegypti in different districts of Rio de Janeiro, Brazil, measured by two types of traps. J. Med. Entomol 2009, 46, 1001–1014. [Google Scholar] [CrossRef] [Green Version]
- Silver, J.B. Mosquito Ecology: Field Sampling Methods, 3rd ed.; Springer: New York, NY, USA, 2008; ISBN 978-1-4020-6666-5. [Google Scholar]
- de Lima-Camara, T.N.; Honório, N.A.; Lourenço-de-Oliveira, R. Freqüência e distribuição espacial de Aedes aegypti e Aedes albopictus (Diptera, Culicidae) no Rio de Janeiro, Brasil. Cad. Saúde Pública 2006, 22, 2079–2084. [Google Scholar] [CrossRef] [PubMed]
- Reis, I.C.; Gibson, G.; Ayllón, T.; de Medeiros Tavares, A.; de Araújo, J.M.G.; da Silva Monteiro, E.; Rodrigues Aguiar, A.; de Oliveira, J.V.; de Paiva, A.A.P.; Wana Bezerra Pereira, H.; et al. Entomo-virological surveillance strategy for dengue, Zika and chikungunya arboviruses in field-caught Aedes mosquitoes in an endemic urban area of the Northeast of Brazil. Acta Trop. 2019, 197, 105061. [Google Scholar] [CrossRef] [PubMed]
- Lima-Camara, T.N.; Honório, N.A.; Lourenço-de-Oliveira, R. Parity and ovarian development of Aedes aegypti and Ae. albopictus (Diptera: Culicidae) in metropolitan Rio de Janeiro. J. Vector Ecol. 2007, 32, 34–40. [Google Scholar] [CrossRef]
- Chow, V.T.; Chan, Y.C.; Yong, R.; Lee, K.M.; Lim, L.K.; Chung, Y.K.; Lam-Phua, S.G.; Tan, B.T. Monitoring of dengue viruses in field-caught Aedes aegypti and Aedes albopictus mosquitoes by a type-specific polymerase chain reaction and cycle sequencing. Am. J. Trop. Med. Hyg. 1998, 58, 578–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lourenço de Oliveira, R.; Vazeille, M.; de Filippis, A.M.B.; Failloux, A.-B. Large genetic differentiation and low variation in vector competence for dengue and yellow fever viruses of Aedes albopictus from Brazil, the United States, and the Cayman Islands. Am. J. Trop. Med. Hyg. 2003, 69, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Guedes, D.R.D.; Cordeiro, M.T.; Melo-Santos, M.A.V.; Magalhaes, T.; Marques, E.; Regis, L.; Furtado, A.F.; Ayres, C.F.J. Patient-based dengue virus surveillance in Aedes aegypti from Recife, Brazil. J. Vector Borne Dis. 2010, 47, 67–75. [Google Scholar]
- Ayllón, T.; de Campos, R.M.; Brasil, P.; Morone, F.C.; Câmara, D.C.P.; Meira, G.L.S.; Tannich, E.; Yamamoto, K.A.; Carvalho, M.S.; Pedro, R.S.; et al. Early Evidence for Zika Virus Circulation among Aedes aegypti Mosquitoes, Rio de Janeiro, Brazil. Emerging Infect. Dis. 2017, 23, 1411–1412. [Google Scholar] [CrossRef] [Green Version]
- Andrade, E.H.P.; Figueiredo, L.B.; Vilela, A.P.P.; Rosa, J.C.C.; Oliveira, J.G.; Zibaoui, H.M.; Araújo, V.E.M.; Miranda, D.P.J.; Ferreira, P.C.P.; Abrahão, J.S.; et al. Spatial–Temporal Co-Circulation of Dengue Virus 1, 2, 3, and 4 Associated with Coinfection Cases in a Hyperendemic Area of Brazil: A 4-Week Survey. Am. J. Trop. Med. Hyg. 2016, 94, 1080–1084. [Google Scholar] [CrossRef] [Green Version]
- Câmara, D.C.P.; da Pinel, C.S.; Rocha, G.P.; Codeço, C.T.; Honório, N.A. Diversity of mosquito (Diptera: Culicidae) vectors in a heterogeneous landscape endemic for arboviruses. Acta. Tropica. 2020, 212, 105715. [Google Scholar] [CrossRef]
- Donatti, J.E.; de Gomes, A.C. Adultrap: Description of adult trap of Aedes aegypti (Diptera, Culicidae). Rev. Bras. Entomol. 2007, 51, 255–256. [Google Scholar] [CrossRef] [Green Version]
- Maciel-de-Freitas, R.; Lourenço-de-Oliveira, R. Presumed unconstrained dispersal of Aedes aegypti in the city of Rio de Janeiro, Brazil. Rev. Saúde Pública 2009, 43, 8–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leandro, A.S.; de Castro, W.A.C.; Lopes, R.D.; Delai, R.M.; Villela, D.A.M.; de-Freitas, R.M. Citywide Integrated Aedes aegypti Mosquito Surveillance as Early Warning System for Arbovirus Transmission, Brazil. Emerg. Infect. Dis. 2022, 28, 701–706. [Google Scholar] [CrossRef] [PubMed]
- Reiter, P.; Amador, M.A.; Colon, N. Enhancement of the CDC ovitrap with hay infusions for daily monitoring of Aedes aegypti populations. J. Am. Mosq. Control. Assoc. 1991, 7, 52–55. [Google Scholar]
- Maciel-de-Freitas, R.; Eiras, Á.E.; Lourenço-de-Oliveira, R. Field evaluation of effectiveness of the BG-Sentinel, a new trap for capturing adult Aedes aegypti (Diptera: Culicidae). Mem. Inst. Oswaldo Cruz 2006, 101, 321–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, C.R.; Long, S.A.; Russell, R.C.; Ritchie, S.A. Field efficacy of the BG-Sentinel compared with CDC Backpack Aspirators and CO2-baited EVS traps for collection of adult Aedes aegypti in Cairns, Queensland, Australia. J. Am. Mosq. Control. Assoc. 2006, 22, 296–300. [Google Scholar] [CrossRef]
- de Figueiredo, R.M.P.; Mourão, M.P.G.; Abi-Abib, Y.E.C.; de Oliveira, C.M.; Roque, R.; de Azara, T.; Ohly, J.; Degener, C.; Geier, M.; Eiras, Á.E. Identification of dengue viruses in naturally infected Aedes aegypti females captured with BioGents (BG)-Sentinel traps in Manaus, Amazonas, Brazil. Rev. Soc. Bras. Med. Trop. 2013, 46, 221–222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sudia, W.D.; Chamberlain, R.W. Battery-operated light trap, an improved model. Mosq. News 1962, 22, 126–129. [Google Scholar]
- Hoel, D.F.; Kline, D.L.; Allan, S.A. Evaluation of six mosquito traps for collection of Aedes albopictus and associated mosquito species in a suburban setting in north central Florida. J. Am. Mosq. Control. Assoc. 2009, 25, 47–57. [Google Scholar] [CrossRef]
- Amos, B.A.; Cardé, R.T. Efficiency of CO2-baited CDC miniature light traps under semi-field conditions and characterizing response behaviors of female Aedes aegypti (Diptera: Culicidae). J. Vector Ecol. 2020, 45, 180–187. [Google Scholar] [CrossRef]
- Newhouse, V.F.; Chamberlain, R.W.; Johnson, J.G.; Sudia, W.D. Use of dry ice to increase mosquito catches of the CDC miniature light trap. Mosq. News 1966, 26, 30–35. [Google Scholar]
- Clark, G.G.; Seda, H.; Gubler, D.J. Use of the “CDC backpack aspirator” for surveillance of Aedes aegypti in San Juan, Puerto Rico. J. Am. Mosq. Control. Assoc. 1994, 10, 119–124. [Google Scholar] [PubMed]
- Scott, T.W.; Morrison, A.C.; Lorenz, L.H.; Clark, G.G.; Strickman, D.; Kittayapong, P.; Zhou, H.; Edman, J.D. Longitudinal studies of Aedes aegypti (Diptera: Culicidae) in Thailand and Puerto Rico: Population dynamics. J. Med. Entomol. 2000, 37, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Dzul-Manzanilla, F.; Ibarra-López, J.; Bibiano Marín, W.; Martini-Jaimes, A.; Leyva, J.T.; Correa-Morales, F.; Huerta, H.; Manrique-Saide, P.; Vazquez-Prokopec, G.M. Indoor Resting Behavior of Aedes aegypti (Diptera: Culicidae) in Acapulco, Mexico. J. Med. Entomol. 2017, 54, 501–504. [Google Scholar] [CrossRef] [PubMed]
- Ayllón, T.; Câmara, D.C.P.; Morone, F.C.; da Gonçalves, L.S.; Saito Monteiro de Barros, F.; Brasil, P.; Carvalho, M.S.; Honório, N.A. Dispersion and oviposition of Aedes albopictus in a Brazilian slum: Initial evidence of Asian tiger mosquito domiciliation in urban environments. PLoS ONE 2018, 13, e0195014. [Google Scholar] [CrossRef] [PubMed]
- Consoli, R.A.G.B.; Lourenço-de-Oliveira, R. Principais Mosquitos de Importância Sanitária No Brasil; Editora FIOCRUZ: Rio de Janeiro, Brasil, 1994; ISBN 85-85676-03-5. [Google Scholar]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Velez, J.O.; Lambert, A.J.; Johnson, A.J.; Stanfield, S.M.; Duffy, M.R. Genetic and serologic properties of Zika virus associated with an epidemic, Yap State, Micronesia, 2007. Emerg. Infect. Dis. 2008, 14, 1232–1239. [Google Scholar] [CrossRef]
- Panning, M.; Grywna, K.; van Esbroeck, M.; Emmerich, P.; Drosten, C. Chikungunya fever in travelers returning to Europe from the Indian Ocean region, 2006. Emerg. Infect. Dis. 2008, 14, 416–422. [Google Scholar] [CrossRef]
- Leparc-Goffart, I.; Baragatti, M.; Temmam, S.; Tuiskunen, A.; Moureau, G.; Charrel, R.; de Lamballerie, X. Development and validation of real-time one-step reverse transcription-PCR for the detection and typing of dengue viruses. J. Clin. Virol. 2009, 45, 61–66. [Google Scholar] [CrossRef]
- Fischer, C.; Torres, M.C.; Patel, P.; Moreira-Soto, A.; Gould, E.A.; Charrel, R.N.; de Lamballerie, X.; Nogueira, R.M.R.; Sequeira, P.C.; Rodrigues, C.D.S.; et al. Lineage-specific real-time RT-PCR for yellow fever virus outbreak surveillance, Brazil. Emerg. Infect. Dis. 2017, 23, 1867. [Google Scholar] [CrossRef]
- Wood, S.N. Generalized Additive Models: An Introduction with R, 2nd ed.; Chapman and Hall/CRC: London, UK, 2017; ISBN 1-4987-2833-2. [Google Scholar]
- Zuur, A.F.; Ieno, E.N.; Walker, N.J.; Saveliev, A.A.; Smith, G.M. Mixed Effects Models and Extensions in Ecology with R; Springer: New York, NY, USA, 2009; ISBN 978-0-387-87457-9. [Google Scholar]
- Crepeau, T.N.; Healy, S.P.; Bartlett-Healy, K.; Unlu, I.; Farajollahi, A.; Fonseca, D.M. Effects of Biogents Sentinel Trap Field Placement on Capture Rates of Adult Asian Tiger Mosquitoes, Aedes albopictus. PLoS ONE 2013, 8, e60524. [Google Scholar] [CrossRef]
- Krockel, U.; Rose, A.; Eiras, A.E.; Geier, M. New tools for surveillance of adult yellow fever mosquitoes: Comparison of trap catches with human landing rates in an urban environment. J. Am. Mosq. Control. Assoc. 2006, 22, 229–238. [Google Scholar] [CrossRef]
- Vehtari, A.; Gelman, A.; Gabry, J. Practical Bayesian model evaluation using leave-one-out cross-validation and WAIC. Stat. Comput. 2017, 27, 1413–1432. [Google Scholar] [CrossRef] [Green Version]
- Tun-Lin, W.; Burkot, T.R.; Kay, B.H. Effects of temperature and larval diet on development rates and survival of the dengue vector Aedes aegypti in north Queensland, Australia. Med. Vet. Entomol. 2000, 14, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, V.C.S.; Tadei, W.P. Frequency, diversity, and productivity study on the Aedes aegypti most preferred containers in the city of Manaus, Amazonas, Brazil. Rev. Inst. Med. Trop. Sao Paulo 2002, 44, 245–250. [Google Scholar] [CrossRef] [PubMed]
- Morrison, A.C.; Gray, K.; Getis, A.; Astete, H.; Sihuincha, M.; Focks, D.; Watts, D.; Stancil, J.D.; Olson, J.G.; Blair, P.; et al. Temporal and Geographic Patterns of Aedes aegypti (Diptera: Culicidae) Production in Iquitos, Peru. J. Med. Entomol. 2004, 41, 1123–1142. [Google Scholar] [CrossRef] [Green Version]
- Vazquez-Prokopec, G.M.; Galvin, W.A.; Kelly, R.; Kitron, U. A new, cost-effective, battery-powered aspirator for adult mosquito collections. J. Med. Entomol. 2009, 46, 1256–1259. [Google Scholar] [CrossRef] [PubMed]
- de Freitas Barata, E.A.; da Costa, A.I.P.; Chiaravalloti Neto, F.; Glasser, C.M.; Barata, J.M.S.; Natal, D. População de Aedes aegypti (l.) em área endêmica de dengue, Sudeste do Brasil. Rev. Saúde Pública 2001, 35, 237–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoeler, G.B.; Schleich, S.S.; Manweiler, S.A.; Sifuentes, V.L. Evaluation of surveillance devices for monitoring Aedes aegypti in an urban area of northeastern Peru. J. Am. Mosq. Control. Assoc. 2004, 20, 6–11. [Google Scholar]
- Hawley, W.A. The biology of Aedes albopictus. J. Am. Mosq. Control. Assoc. Suppl. 1988, 1, 1–39. [Google Scholar]
- Delatte, H.; Desvars, A.; Bouétard, A.; Bord, S.; Gimonneau, G.; Vourc’h, G.; Fontenille, D. Blood-feeding behavior of Aedes albopictus, a vector of Chikungunya on La Réunion. Vector Borne Zoonotic Dis. 2010, 10, 249–258. [Google Scholar] [CrossRef] [Green Version]
- Ball, T.S.; Ritchie, S.R. Sampling biases of the BG-sentinel trap with respect to physiology, age, and body size of adult Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2010, 47, 649–656. [Google Scholar] [CrossRef]
- Degener, C.M.; Eiras, A.E.; Azara, T.M.F.; Roque, R.A.; Rösner, S.; Codeço, C.T.; Nobre, A.A.; Rocha, E.S.O.; Kroon, E.G.; Ohly, J.J.; et al. Evaluation of the effectiveness of mass trapping with BG-sentinel traps for dengue vector control: A cluster randomized controlled trial in Manaus, Brazil. J. Med. Entomol. 2014, 51, 408–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meeraus, W.H.; Armistead, J.S.; Arias, J.R. Field comparison of novel and gold standard traps for collecting Aedes albopictus in Northern Virginia. J. Am. Mosq. Control. Assoc. 2008, 24, 244–248. [Google Scholar] [CrossRef] [PubMed]
- Farajollahi, A.; Kesavaraju, B.; Price, D.C.; Williams, G.M.; Healy, S.P.; Gaugler, R.; Nelder, M.P. Field efficacy of BG-Sentinel and industry-standard traps for Aedes albopictus (Diptera: Culicidae) and West Nile virus surveillance. J. Med. Entomol. 2009, 46, 919–925. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drago, A.; Marini, F.; Caputo, B.; Coluzzi, M.; della Torre, A.; Pombi, M. Looking for the gold standard: Assessment of the effectiveness of four traps for monitoring mosquitoes in Italy. J. Vector Ecol. 2012, 37, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Englbrecht, C.; Gordon, S.; Venturelli, C.; Rose, A.; Geier, M. Evaluation of BG-Sentinel Trap as a Management Tool to Reduce Aedes albopictus Nuisance in an Urban Environment in Italy. J. Am. Mosq. Control. Assoc. 2015, 31, 16–25. [Google Scholar] [CrossRef]
- Ritchie, S.A.; Moore, P.; Carruthers, M.; Williams, C.; Montgomery, B.; Foley, P.; Ahboo, S.; van den Hurk, A.F.; Lindsay, M.D.; Cooper, B.; et al. Discovery of a widespread infestation of Aedes albopictus in the Torres Strait, Australia. J. Am. Mosq. Control. Assoc. 2006, 22, 358–365. [Google Scholar] [CrossRef]
- Smartt, C.T.; Stenn, T.M.S.; Chen, T.-Y.; Teixeira, M.G.; Queiroz, E.P.; Souza Dos Santos, L.; Queiroz, G.A.N.; Ribeiro Souza, K.; Kalabric Silva, L.; Shin, D.; et al. Evidence of Zika Virus RNA Fragments in Aedes albopictus (Diptera: Culicidae) Field-Collected Eggs From Camaçari, Bahia, Brazil. J. Med. Entomol. 2017, 54, 1085–1087. [Google Scholar] [CrossRef] [PubMed]
- de Gomes, A.C.; da Silva, N.N.; Bernal, R.T.I.; de Leandro, A.S.; de Camargo, N.J.; da Silva, A.M.; Ferreira, A.C.; Ogura, L.C.; de Oliveira, S.J.; de Moura, S.M. Especificidade da armadilha Adultrap para capturar fêmeas de Aedes aegypti (Diptera: Culicidae). Rev. Soc. Bras. Med. Trop. 2007, 40, 216–219. [Google Scholar] [CrossRef] [Green Version]
- Jensen, T.; Willis, O.R.; Fukuda, T.; Barnard, D.R. Comparison of bi-directional Fay, omni-directional, CDC, and duplex cone traps for sampling adult Aedes albopictus and Aedes aegypti in north Florida. J. Am. Mosq. Control. Assoc. 1994, 10, 74–78. [Google Scholar]
- Dennett, J.A.; Vessey, N.Y.; Parsons, R.E. A comparison of seven traps used for collection of Aedes albopictus and Aedes aegypti originating from a large tire repository in Harris County (Houston), Texas. J. Am. Mosq. Control. Assoc. 2004, 20, 342–349. [Google Scholar]
- Maciel-de-Freitas, R.; Peres, R.C.; Alves, F.; Brandolini, M.B. Mosquito traps designed to capture Aedes aegypti (Diptera: Culicidae) females: Preliminary comparison of Adultrap, MosquiTRAP and backpack aspirator efficiency in a dengue-endemic area of Brazil. Mem. Inst. Oswaldo Cruz 2008, 103, 602–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, N.S.; de Carvalho, G.C.; dos Santos, Y.G.A.; Monte-Alegre, A.F. Increased capture of Aedes aegypti (Linnaeus, 1762) (Diptera: Culicidae) by removing one ADULTRAP component. Rev. Soc. Bras. Med. Trop. 2020, 53, e20190043. [Google Scholar] [CrossRef]
- Gillies, M.T. The role of carbon dioxide in host-finding by mosquitoes (Diptera: Culicidae): A review. Bull. Entomol. Res. 1980, 70, 525–532. [Google Scholar] [CrossRef] [Green Version]
- Reeves, W.C. Quantitative field studies on a carbon dioxide chemotropism of mosquitoes. 1953. J. Am. Mosq Control. Assoc. 1990, 6, 708–712. [Google Scholar]
- Dekker, T.; Geier, M.; Cardé, R.T. Carbon dioxide instantly sensitizes female yellow fever mosquitoes to human skin odours. J. Exp. Biol 2005, 208, 2963–2972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McMeniman, C.J.; Corfas, R.A.; Matthews, B.J.; Ritchie, S.A.; Vosshall, L.B. Multimodal integration of carbon dioxide and other sensory cues drives mosquito attraction to humans. Cell 2014, 156, 1060–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Ázara, T.M.F.; Degener, C.M.; Roque, R.A.; Ohly, J.J.; Geier, M.; Eiras, Á.E. The impact of CO2 on collection of Aedes aegypti (Linnaeus) and Culex quinquefasciatus Say by BG-Sentinel® traps in Manaus, Brazil. Mem Inst. Oswaldo Cruz 2013, 108, 229–232. [Google Scholar] [CrossRef] [Green Version]
- Roiz, D.; Duperier, S.; Roussel, M.; Boussès, P.; Fontenille, D.; Simard, F.; Paupy, C. Trapping the Tiger: Efficacy of the Novel BG-Sentinel 2 With Several Attractants and Carbon Dioxide for Collecting Aedes albopictus (Diptera: Culicidae) in Southern France. J. Med. Entomol. 2016, 53, 460–465. [Google Scholar] [CrossRef]
- Wilke, A.B.B.; Carvajal, A.; Medina, J.; Anderson, M.; Nieves, V.J.; Ramirez, M.; Vasquez, C.; Petrie, W.; Cardenas, G.; Beier, J.C. Assessment of the effectiveness of BG-Sentinel traps baited with CO2 and BG-Lure for the surveillance of vector mosquitoes in Miami-Dade County, Florida. PLoS ONE 2019, 14, e0212688. [Google Scholar] [CrossRef]
- Russell, R.C. The relative attractiveness of carbon dioxide and octenol in CDC- and EVS-type light traps for sampling the mosquitoes Aedes aegypti (L.), Aedes polynesiensis Marks, and Culex quinquefasciatus say in Moorea, French Polynesia. J. Vector Ecol. 2004, 29, 309–314. [Google Scholar]
- Connor, M.E.; Monroe, W.M. Stegomyia Indices and Their Value in Yellow Fever Control1. Am. J. Trop. Med. Hyg. 1923, 3, 9–19. [Google Scholar] [CrossRef]
- Breteau, H. Yellow fever in French West Africa; an aspect of mass-preventive medicine. Bull. World Health Organ. 1954, 11, 453–481. [Google Scholar] [PubMed]
- Tun-Lin, W.; Kay, B.H.; Barnes, A. The Premise Condition Index: A tool for streamlining surveys of Aedes aegypti. Am. J. Trop. Med. Hyg. 1995, 53, 591–594. [Google Scholar] [CrossRef]
- Gu, W.; Novak, R.J. Short report: Detection probability of arbovirus infection in mosquito populations. Am. J. Trop. Med. Hyg. 2004, 71, 636–638. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Unnasch, T.R.; Katholi, C.R.; Lampman, R.; Novak, R.J. Fundamental issues in mosquito surveillance for arboviral transmission. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 817–822. [Google Scholar] [CrossRef] [Green Version]
- Nogueira, R.M.; Schatzmayr, H.G.; Miagostovich, M.P.; Farias, M.F.; Farias Filho, J.D. Virological study of a dengue type 1 epidemic at Rio de Janeiro. Mem. Inst. Oswaldo Cruz 1988, 83, 219–225. [Google Scholar] [CrossRef]
- Dégallier, N.; Teixeira, J.M.S.; Vilarinhos, P.d.T.R.; Pinto, S.C.F.; Pereira, R.D. First isolation of dengue 1 virus from Aedes aegypti in Federal District, Brazil. Rev. Soc. Bras. Med. Trop. 2000, 33, 95–96. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, V.C.S.; Tadei, W.P.; Barros, P.M.S.S.; Vasconcelos, P.F.C.; Cruz, A.C.R. Detection of dengue virus serotype 3 by reverse transcription-polymerase chain reaction in Aedes aegypti (Diptera, Culicidae) captured in Manaus, Amazonas. Mem. Inst. Oswaldo Cruz 2005, 100, 833–839. [Google Scholar] [CrossRef] [Green Version]
- Kow, C.Y.; Koon, L.L.; Yin, P.F. Detection of dengue viruses in field caught male Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in Singapore by type-specific PCR. J. Med. Entomol. 2001, 38, 475–479. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.-F.; Shu, P.-Y.; Teng, H.-J.; Su, C.-L.; Wu, J.-W.; Wang, J.-H.; Lin, T.-H.; Huang, J.-H.; Wu, H.-S. Screening of dengue virus in field-caught Aedes aegypti and Aedes albopictus (Diptera: Culicidae) by one-step SYBR green-based reverse transcriptase-polymerase chain reaction assay during 2004-2007 in Southern Taiwan. Vector Borne Zoonotic Dis. 2010, 10, 1017–1025. [Google Scholar] [CrossRef]
- De Castro, M.G.; Nogueira, R.M.R.; de Filippis, A.M.B.; Ferreira, A.A.; da Lima, M.R.Q.; da Faria, N.R.C.; de Nogueira, F.B.; Simões, J.B.S.; Nunes, P.C.G.; Sampaio, S.A.; et al. Dengue virus type 4 in Niterói, Rio de Janeiro: The role of molecular techniques in laboratory diagnosis and entomological surveillance. Mem. Inst. Oswaldo Cruz 2012, 107, 940–945. [Google Scholar] [CrossRef]
- Elizondo-Quiroga, D.; Medina-Sánchez, A.; Sánchez-González, J.M.; Eckert, K.A.; Villalobos-Sánchez, E.; Navarro-Zúñiga, A.R.; Sánchez-Tejeda, G.; Correa-Morales, F.; González-Acosta, C.; Arias, C.F.; et al. Zika Virus in Salivary Glands of Five Different Species of Wild-Caught Mosquitoes from Mexico. Sci. Rep. 2018, 8, 809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maciel-de-Freitas, R.; Neto, R.B.; Gonçalves, J.M.; Codeço, C.T.; Lourenço-de-Oliveira, R. Movement of dengue vectors between the human modified environment and an urban forest in Rio de Janeiro. J. Med. Entomol. 2006, 43, 1112–1120. [Google Scholar] [CrossRef]
- Dalpadado, R.; Amarasinghe, D.; Gunathilaka, N.; Ariyarathna, N. Bionomic aspects of dengue vectors Aedes aegypti and Aedes albopictus at domestic settings in urban, suburban and rural areas in Gampaha District, Western Province of Sri Lanka. Parasites Vectors 2022, 15, 148. [Google Scholar] [CrossRef] [PubMed]
- Chadee, D.D. Resting behaviour of Aedes aegypti in Trinidad: With evidence for the re-introduction of indoor residual spraying (IRS) for dengue control. Parasites Vectors 2013, 6, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perich, M.J.; Davila, G.; Turner, A.; Garcia, A.; Nelson, M. Behavior of resting Aedes aegypti (Culicidae: Diptera) and its relation to ultra-low volume adulticide efficacy in Panama City, Panama. J. Med. Entomol. 2000, 37, 541–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xia, S.; Dweck, H.K.M.; Lutomiah, J.; Sang, R.; McBride, C.S.; Rose, N.H.; Ayala, D.; Powell, J.R. Larval sites of the mosquito Aedes aegypti formosus in forest and domestic habitats in Africa and the potential association with oviposition evolution. Ecol. Evol. 2021, 11, 16327–16343. [Google Scholar] [CrossRef]
- Juliano, S.A.; Lounibos, L.P.; O’Meara, G.F. A field test for competitive effects of Aedes albopictus on A. aegypti in South Florida: Differences between sites of coexistence and exclusion? Oecologia 2004, 139, 583–593. [Google Scholar] [CrossRef] [Green Version]
- Camara, D.C.P.; Codeço, C.T.; Juliano, S.A.; Lounibos, L.P.; Riback, T.I.S.; Pereira, G.R.; Honorio, N.A. Seasonal Differences in Density But Similar Competitive Impact of Aedes albopictus (Skuse) on Aedes aegypti (L.) in Rio de Janeiro, Brazil. PLoS ONE 2016, 11, e0157120. [Google Scholar] [CrossRef]
- Juliano, S.A.; O’Meara, G.F.; Morrill, J.R.; Cutwa, M.M. Desiccation and thermal tolerance of eggs and the coexistence of competing mosquitoes. Oecologia 2002, 130, 458–469. [Google Scholar] [CrossRef] [Green Version]
- Costanzo, K.S.; Kesavaraju, B.; Juliano, S.A. Condition-specific competition in container mosquitoes: The role of noncompeting life-history stages. Ecology 2005, 86, 3289–3295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lounibos, L.P.; O’Meara, G.F.; Juliano, S.A.; Nishimura, N.; Escher, R.L.; Reiskind, M.H.; Cutwa, M.; Greene, K. Differential Survivorship of Invasive Mosquito Species in South Florida Cemeteries: Do Site-Specific Microclimates Explain Patterns of Coexistence and Exclusion? Ann. Entomol. Soc. Am. 2010, 103, 757–770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiskind, M.H.; Lounibos, L.P. Spatial and temporal p.patterns of abundance of Aedes aegypti L. (Stegomyia aegypti) and Aedes albopictus (Skuse) [Stegomyia albopictus (Skuse)] in southern Florida. Med. Vet. Entomol. 2013, 27, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Braks, M.A.H.; Honório, N.A.; Lourenço-De-Oliveira, R.; Juliano, S.A.; Lounibos, L.P. Convergent habitat segregation of Aedes aegypti and Aedes albopictus (Diptera: Culicidae) in southeastern Brazil and Florida. J. Med. Entomol. 2003, 40, 785–794. [Google Scholar] [CrossRef] [Green Version]
- Ferreira-de-Lima, V.H.; Câmara, D.C.P.; Honório, N.A.; Lima-Camara, T.N. The Asian tiger mosquito in Brazil: Observations on biology and ecological interactions since its first detection in 1986. Acta Trop. 2020, 205, 105386. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Aedes Aegypti | Aedes Albopictus | |||||
|---|---|---|---|---|---|---|
| Method | Urban | Peri-Urban | Rural | Urban | Peri-Urban | Rural |
| ADT | 19/19 (2) | 11/10 (1) | 5/5 (0) | 8/7 (0) | 25/25 (0) | 10/10 (2) |
| BGS | 78/36 (3) | 10/6 (3) | 18/11 (4) | 22/21 (0) | 46/43 (2) | 109/84 (0) |
| CDC | 15/11 (0) | 5/3 (0) | 22/17 (0) | 27/26 (0) | 19/19 (0) | 43/42 (0) |
| ASP-I | 81/42 (42) | 85/44 (34) | 12/7 (6) | 4/2 (2) | 3/3 (3) | 2/2 (2) |
| ASP-O | 28/9 (6) | 34/23 (17) | 11/0 (0) | 14/5 (1) | 15/9 (4) | 46/16 (0) |
| Total | 221/117 (53) | 145/86 (55) | 68/40 (10) | 75/61 (3) | 108/99 (9) | 210/156 (4) |
| Effects | Estimates | Aedes Aegypti | Aedes Albopictus | ||||
|---|---|---|---|---|---|---|---|
| Mean | CI95 | SD | Mean | CI95 | SD | ||
| Fixed | Intercept | −1.653 | (−2.147, −1.156) | 0.250 | −1.689 | (−2.351, −0.909) | 0.370 |
| Period: Dry | −0.094 | (−0.456, 0.279) | 0.187 | −0.827 | (−1.456, −0.219) | 0.318 | |
| Landscape: Periurban | −0.570 | (−0.992, −0.140) | 0.217 | 0.562 | (−0.141, 1.266) | 0.358 | |
| Landscape: Rural | −1.069 | (−1.510, −0.628) | 0.225 | 1.189 | (0.413, 1.960) | 0.383 | |
| ADT | −1.055 | (−1.637, −0.453) | 0.304 | −1.471 | (−2.349, −0.673) | 0.421 | |
| CDC | −0.803 | (−1.412, −0.200) | 0.306 | −0.829 | (−1.630, −0.066) | 0.406 | |
| ASP-I | 0.471 | (−0.046, 1.000) | 0.264 | −3.185 | (−4.339, −2.132) | 0.561 | |
| ASP-O | −0.273 | (−0.803, 0.263) | 0.278 | −1.003 | (−1.867, −0.222) | 0.411 | |
| Random | Site | 0.179 | (0.014, 0.479) | 0.121 | 0.016 | (0, 0.114) | 0.035 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Câmara, D.C.P.; Codeço, C.T.; Ayllón, T.; Nobre, A.A.; Azevedo, R.C.; Ferreira, D.F.; da Silva Pinel, C.; Rocha, G.P.; Honório, N.A. Entomological Surveillance of Aedes Mosquitoes: Comparison of Different Collection Methods in an Endemic Area in RIO de Janeiro, Brazil. Trop. Med. Infect. Dis. 2022, 7, 114. https://0-doi-org.brum.beds.ac.uk/10.3390/tropicalmed7070114
Câmara DCP, Codeço CT, Ayllón T, Nobre AA, Azevedo RC, Ferreira DF, da Silva Pinel C, Rocha GP, Honório NA. Entomological Surveillance of Aedes Mosquitoes: Comparison of Different Collection Methods in an Endemic Area in RIO de Janeiro, Brazil. Tropical Medicine and Infectious Disease. 2022; 7(7):114. https://0-doi-org.brum.beds.ac.uk/10.3390/tropicalmed7070114
Chicago/Turabian StyleCâmara, Daniel Cardoso Portela, Claudia Torres Codeço, Tania Ayllón, Aline Araújo Nobre, Renata Campos Azevedo, Davis Fernandes Ferreira, Célio da Silva Pinel, Gláucio Pereira Rocha, and Nildimar Alves Honório. 2022. "Entomological Surveillance of Aedes Mosquitoes: Comparison of Different Collection Methods in an Endemic Area in RIO de Janeiro, Brazil" Tropical Medicine and Infectious Disease 7, no. 7: 114. https://0-doi-org.brum.beds.ac.uk/10.3390/tropicalmed7070114