Magnetostratigraphy and Chronology of the Lower Pleistocene Primate Bearing Dafnero Fossil Site, N. Greece

 , ,
, ,
Abstract
:
1. Introduction
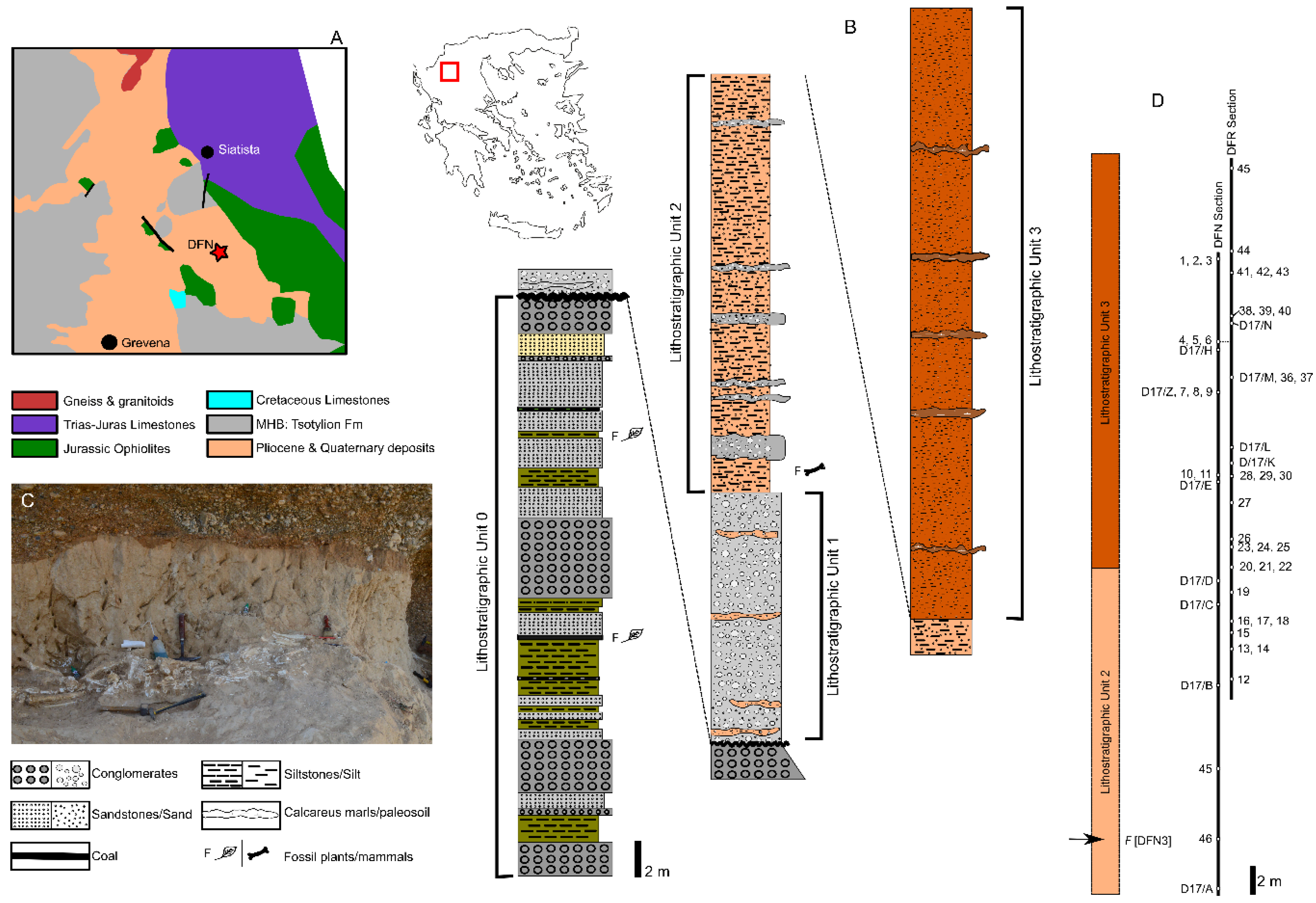
2. Geographical and Geological Settings
3. Paleomagnetic Sampling and Laboratory Procedures
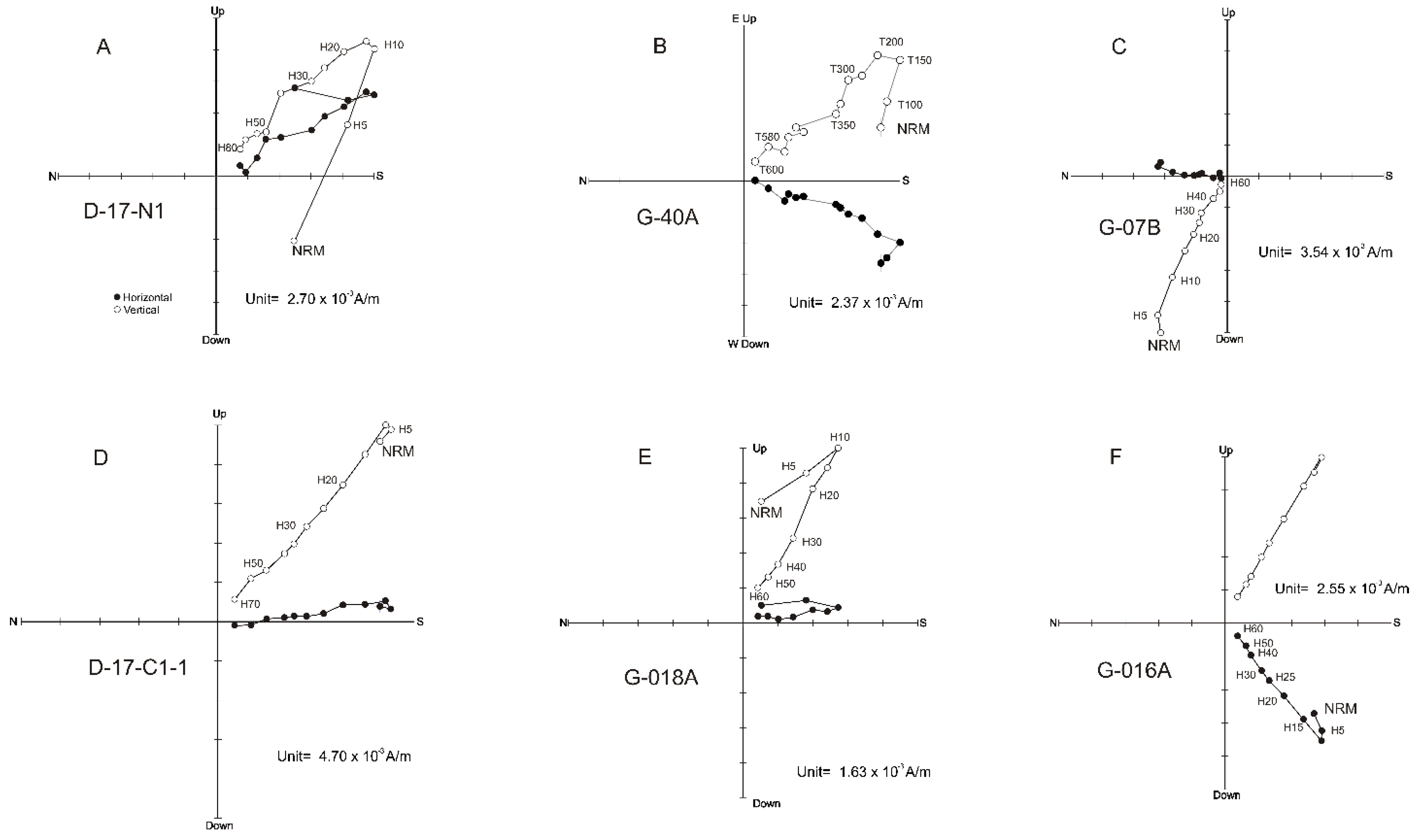
4. Paleomagnetic Results
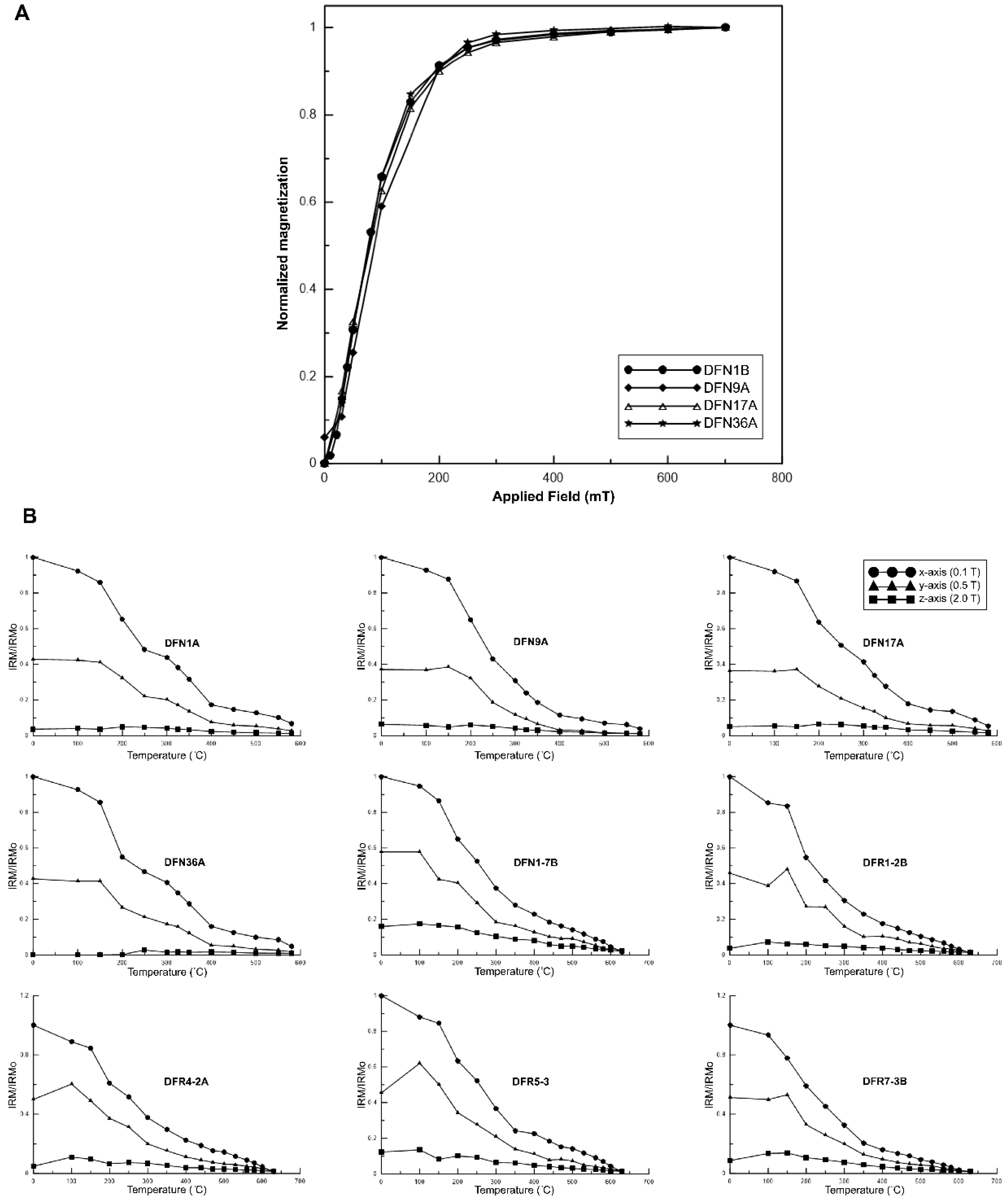
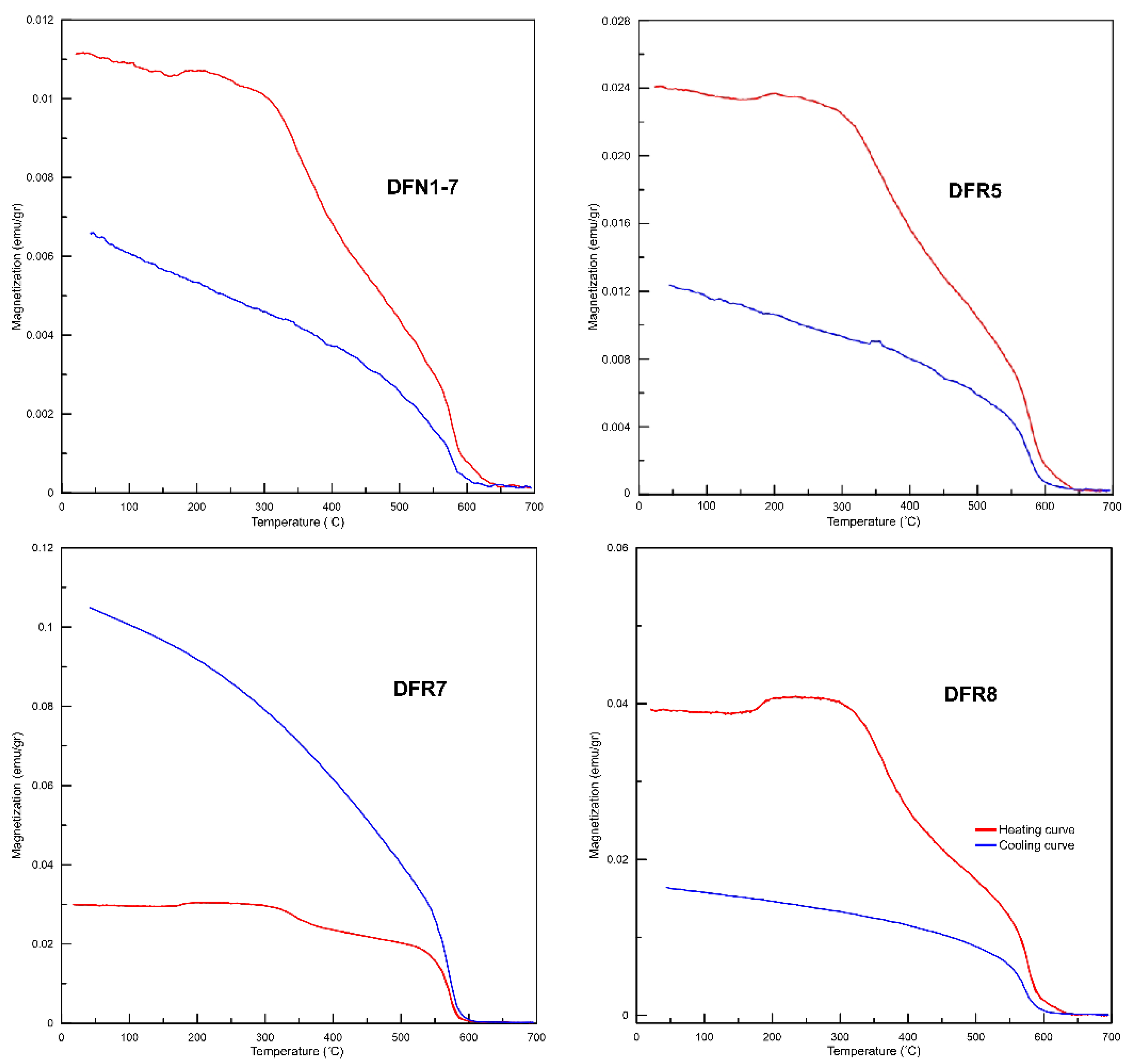
4.1. Magnetic Mineralogy
4.2. Curie Balance
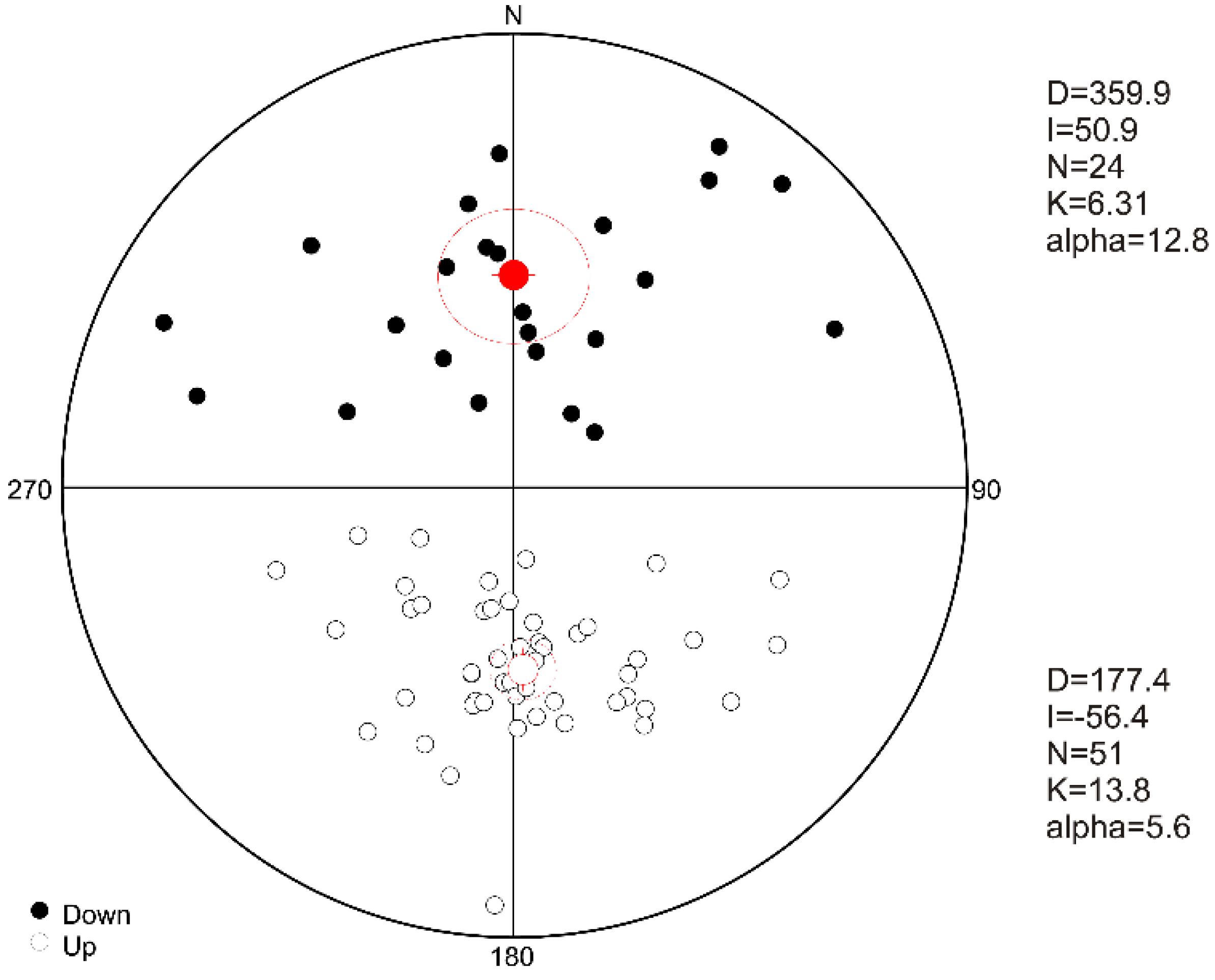
4.3. Natural Remanent Magnetization
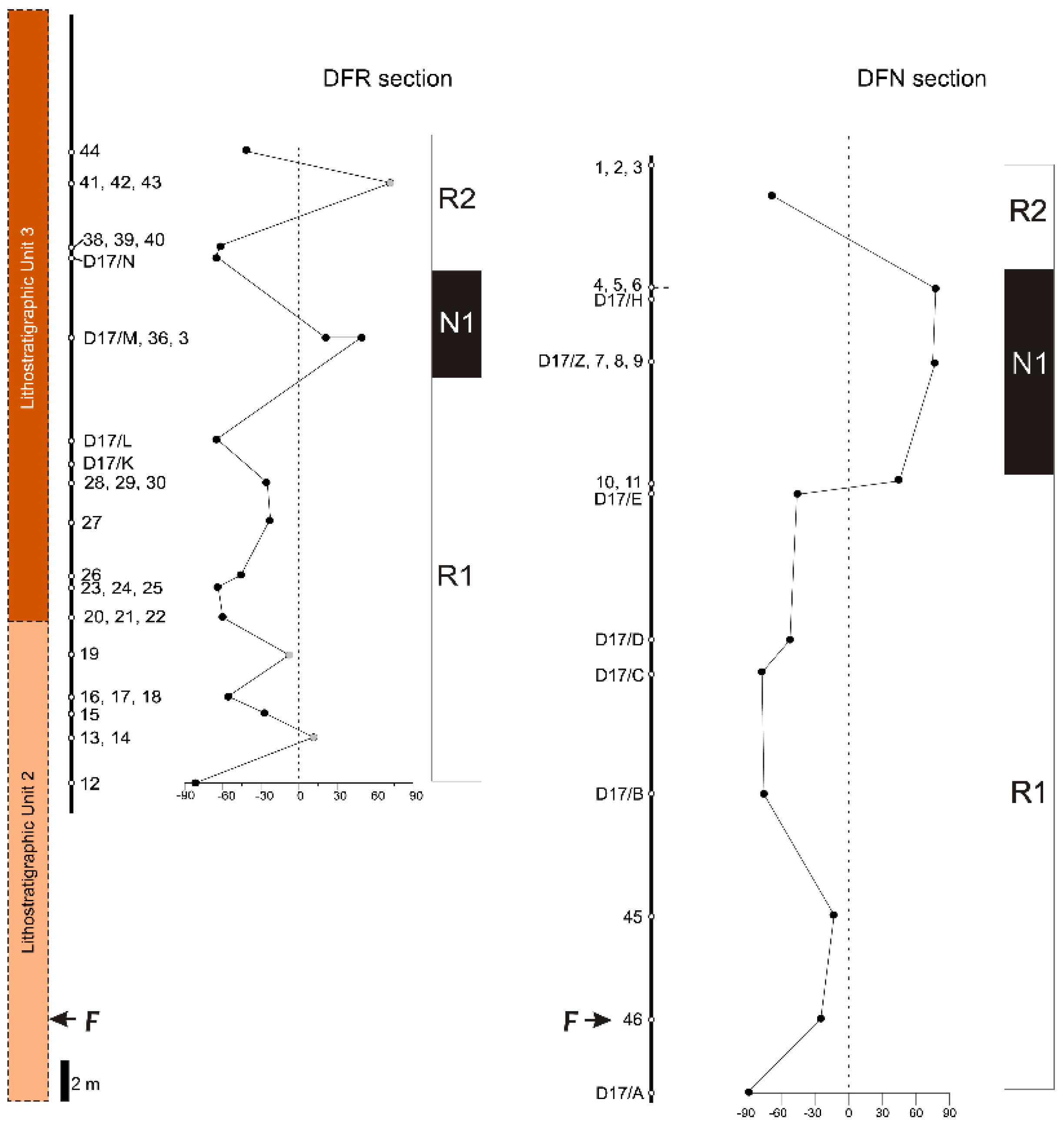
4.4. Magnetostratigraphic Interpetation
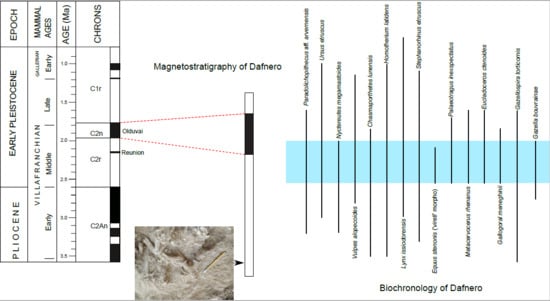
5. Age Estimation
5.1. Biochronological Frame
5.2. Correlation with GPTS
6. The Southern Balkan Middle Villafranchian Faunal Succession
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Torre, D.; Albianelli, A.; Bertini, A.; Ficcarelli, G.; Masini, F.; Napoleone, G. Paleomagnetic calibration of Plio-Pleistocene mammal localities in central Italy. Acta Zool. Cracov. 1996, 39, 559–570. [Google Scholar]
- Albianelli, A.; Azzaroli, A.; Beriini, A.; Ficcarelli, G.; Napoleone, G.; Torre, D. Paleomagnetic and palynologic investigations in the upper Valdarno basin (central Italy): Calibration of an early Villafranchian fauna. Riv. Ital. Paleontol. Stratigr. 1997, 103, 111–118. [Google Scholar]
- Sen, S. Magnetostratigraphic calibration of the Neogene mammal chronology. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1997, 133, 181–204. [Google Scholar] [CrossRef]
- Sen, S. Magnetostratigraphy of the Villafranchian mammal locality of Saint-Vallier (Drome). Geobios 2004, 37, 58–61. [Google Scholar] [CrossRef]
- Oms, O.; Dinares-Turell, J.; Agustı, J.; Pares, J. Refinements of the European Mammal Biochronology from the Magnetic Polarity Record of the Plio–Pleistocene Zujar section, Guadix-Baza Basin, SE Spain. Quat. Res. 1999, 51, 94–103. [Google Scholar] [CrossRef]
- Agusti, J.; Cabrera, L.; Garces, M.; Krijgsman, W.; Oms, O.; Pares, J.M. A calibrated mammal scale for the Neogene of Western Europe: State of the art. Earth Sci. Rev. 2001, 52, 247–260. [Google Scholar] [CrossRef] [Green Version]
- Scott, G.R.; Gibert, L.I.; Gibert, J. Magnetostratigraphy of the Orce region (Baza Basin), SE Spain: New chronologies for Early Pleistocene faunas and hominid occupation sites. Quat. Sci. Rev. 2007, 26, 415–435. [Google Scholar] [CrossRef]
- Madurell-Malapeira, J.; Minwer-Barakat, R.; Alba, D.M.; Garcés, M.; Gómez, M.; Aurell-Garrido, J.; Ros-Montoya, R.; Moyà-Solà, S.; Berástegui, X. The Vallparadís section (Terrassa, Iberian Peninsula) and the latest Villafranchian faunas of Europe. Quat. Sci. Rev. 2010, 29, 3972–3982. [Google Scholar] [CrossRef]
- Muttoni, G.; Scardia, G.; Kent, D.V. Human migration into Europe during the late Early Pleistocene climate transition. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 296, 79–93. [Google Scholar] [CrossRef] [Green Version]
- Muttoni, G.; Scardia, G.; Kent, D.V. Early hominins in Europe: The Galerian migration hypothesis. Quat. Sci. Rev. 2018, 180, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Pueyo, E.; Muñoz, A.; Laplana, C.; Parés, J. The Last Appearance Datum of Hipparion in Western Europe: Magnetostratigraphy along the Pliocene–Pleistocene boundary in the Villarroya Basin (Northern Spain). Int. J. Earth Sci. (Geol. Rundsch) 2016, 105, 2203–2220. [Google Scholar] [CrossRef]
- Muttoni, G.; Kent, D.; Gaetani, M. Magnetostratigraphy of a Lower–Middle Triassic boundary section from Chios, Greece. Phys. Earth Planet. Inter. 1995, 92, 245–260. [Google Scholar] [CrossRef] [Green Version]
- Muttoni, G.; Kent, D.; Brack, P.; Nicora, A.; Balini, M. Middle Triassic magnetostratigraphy and biostratigraphy biostratigraphy from the Dolomites and Greece. Earth Planet. Sci. Lett. 1997, 146, 107–120. [Google Scholar] [CrossRef] [Green Version]
- Van Vugt, N.; Steenbrink, J.; Langereis, C.G.; Hilgen, F.J.; Meulenkamp, J.E. Magnetostratigraphy-based astronomical tuning of the early Pliocene lacustrine sediments of Ptolemais (NW Greece) and bed-to-bed correlation with the marine record. Earth Planet. Sci. Lett. 1998, 164, 535–551. [Google Scholar] [CrossRef] [Green Version]
- Steenbrink, J.; Hilgen, F.J.; Krijgsman, W.; Wijbrans, J.R.; Meulenkamp, J.E. Late Miocene to Early Pliocene depositional history of the intramontane Florina–Ptolemais–Servia Basin, NW Greece: Interplay between orbital forcing and tectonics. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2006, 238, 151–178. [Google Scholar] [CrossRef]
- Cornée, J.-J.; Moissette, P.; Joannin, S.; Suc, J.-P.; Quillévéré, F.; Krijgsman, W.; Hilgen, F.; Koskeridou, F.; Münch, P.H.; Lécuyer, C.H.; et al. Tectonic and climatic controls on coastal sedimentation: The Late Pliocene–Middle Pleistocene of Northeastern Rhodes, Greece. Sediment. Geol. 2006, 187, 159–181. [Google Scholar] [CrossRef]
- Snel, E.; Mărunţeanu, M.; Meulenkamp, J.E. Calcareous nannofossil biostratigraphy and magnetostratigraphy of the Upper Miocene and Lower Pliocene of the Northern Aegean (Orphanic Gulf–Strimon Basin areas), Greece. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2006, 238, 125–150. [Google Scholar] [CrossRef]
- Sen, S.; Valet, J.P. Magnetostratigraphy of late Miocene continental deposits in Samos, Greece. Earth Planet. Sci. Lett. 1986, 80, 167–174. [Google Scholar] [CrossRef]
- Sen, S.; Koufos, G.; Kondopoulou, D.; Bonis, L. Magnetostratigraphy of the late Miocene continental deposits of the lower Axios valley, Macedonia, Greece. Bull. Geol. Soc. Greece Spec. Issue 2000, 9, 197–206. [Google Scholar]
- Kostopoulos, D.S.; Sen, S.; Koufos, G.D. Magnetostratigraphy and revised chronology of the late Miocene mammal localities of Samos, Greece. Int. J. Earth Sci. 2003, 92, 779–794. [Google Scholar] [CrossRef]
- Kondopoulou, D.; Sen, S.; Aidona, E.; van Hinsbergen, D.J.J.; Koufos, G. Rotation history of Chios Island, Greece since the Middle Miocene. J. Geodyn. 2011, 51, 327–338. [Google Scholar] [CrossRef]
- Böhme, M.; Spassov, N.; Ebner, M.; Geraads, D.; Hristova, L.; Kirscher, U.; Kötter, S.; Linnemann, U.; Prieto, J.; Roussiakis, S.; et al. Messinian age and savannah environment of the possible hominin Graecopithecus from Europe. PLoS ONE 2017, 12, e0177347. [Google Scholar] [CrossRef]
- Tourloukis, V.; Muttoni, G.; Karkanas, P.; Monesi, E.; Scardia, G.; Panagopoulou, E.; Harvati, K. Magnetostratigraphic and chronostratigraphic constraints on the Marathousa 1 Lower Palaeolithic site and the Middle Pleistocene deposits of the Megalopolis basin, Greece. Quat. Int. 2018, 497, 154–169. [Google Scholar] [CrossRef]
- Koufos, G.D.; Kostopoulos, D.S.; Koliadimou, K.K. Un nouveau gisement de mammifères dans le Villafranchien de Macédoine occidentale (Grèce). CR Acad. Sci. Paris 1991, 313, 831–836. [Google Scholar]
- Koufos, G.D. Late Pliocene carnivores from Western Macedonia (Greece). Paläontol. Z. 1993, 67, 357–376. [Google Scholar] [CrossRef]
- Koufos, G.D.; Kostopoulos, D.S. A stenonoid horse (Equidae, Mammalia) from the Villafranchian of Western Macedonia (Greece). Bull. Geol. Soc. Greece 1993, 28, 131–143. [Google Scholar]
- Koufos, G.D.; Kostopoulos, D.S. New Carnivore material from the Plio-Pleistocene of Macedonia (Greece) with the description of a new canid. Münch. Geowiss. Abh. A Geol. Paläontol. 1997, 34, 33–63. [Google Scholar]
- Kostopoulos, D.S.; Koufos, G. The Plio-Pleistocene artiodactyls of Macedonia (Northern Greece) and their biostratigrphic significance; preliminary report. CR Acad. Sci. Paris 1994, 318, 1267–1272. [Google Scholar]
- Kostopoulos, D.S.; Guy, F.; Kynigopoulou, Z.; Koufos, G.D.; Valentin, X.; Merceron, G. A 2Ma old baboon-like monkey from Northern Greece and new evidence to support the Paradolichopithecus–Procynocephalus synonymy (Primates: Cercopithecidae). J. Hum. Evol. 2018, 121, 178–192. [Google Scholar] [CrossRef]
- Koufos, G.D.; Kostopoulos, D.S.; Merceron, G. The saber-toothed cat Homotherium latidens (Owen, 1864) from the lower Pleistocene locality Dafnero, Western Macedonia, Greece. Geodiversitas 2020, 42, 139–149. [Google Scholar] [CrossRef]
- Rook, L.; Martínez-Navarro, B. Villafranchian: The long story of a Plio-Pleistocene European large mammal biochronologic unit. Quat. Int. 2010, 219, 134–144. [Google Scholar] [CrossRef]
- Sinusía, C.; Pueyo, E.L.; Azanza, B.; Pocoví, A. Datación magnetoestratigráfica del yacimiento paleontológico de la Puebla de Valverde (Teruel). Geotemas 2006, 6, 329–342. [Google Scholar]
- Pla-Pueyo, S.; Viseras, C.; Soria, J.M.; Tent-Manclús, J.E.; Arribas, A. A stratigraphic framework for the Pliocene-Pleistocene continental sediments of the Guadix Basin (Betic Cordillera, S. Spain). Quat. Int. 2011, 243, 16–32. [Google Scholar] [CrossRef]
- Nomade, S.; Pastre, J.F.; Guillou, H.; Faure, H.M.; Guérin, C.; Delson, E.; Debard, E.; Voinchet, P.; Messager, E. 40Ar/39Ar constraints on some French landmark Late Pliocene to Early Pleistocene large mammalian paleofaunas: Paleoenvironmental and paleoecological implications. Quat. Geochronol. 2014, 21, 2–15. [Google Scholar] [CrossRef]
- Zelilidis, A.; Piper, D.J.W.; Kontopoulos, N. Sedimentation and basin evolution of the Oligocene–Miocene Mesohellenic basin, Greece. AAPG Bull. 2002, 86, 161–182. [Google Scholar]
- Ferrière, J.; Chanier, F.; Reynaud, J.-Y.; Pavlopoulos, A.; Ditbanjong, P.; Coutand, I. Evolution of the Mesohellenic Basin (Greece): A synthesis. In The Geology of Greece-Part II; Skourtsos, E., Ed.; The Virtual Explorer Pty Ltd.: Conder, Australia, 2013. [Google Scholar]
- Kilias, A.D.; Vamvaka, A.; Falalakis, G.; Sfeikos, A.; Papadimitriou, E.; Gkarlaouni, C.; Karakostas, B. The Mesohellenic Trough and the Paleogene Thrace Basin on the Rhodope Massif, their Structural Evolution and Geotectonic Significance in the Hellenides. J. Geol. Geosci. 2015, 4, 198. [Google Scholar] [CrossRef] [Green Version]
- Lowrie, W. Identification of ferrimagnetic minerals in rock by coercivity and unblocking temperature properties. Geophys. Res. Lett. 1990, 17, 159–162. [Google Scholar] [CrossRef]
- Zijderveld, J.D.A. AC demagnetization rocks: Analyses of results. In Methods in Paleomagnetism; Collinson, D.W., Creer, K.M., Runcorn, S.K., Eds.; Elsevier Scientific: Amsterdam, The Netherlands, 1967; pp. 254–286. [Google Scholar]
- Kirschvink, J.L. The least-square line and plane and analysis of palaeomagnetic data. Geophys. J. R. Astron. Soc. 1980, 62, 699–718. [Google Scholar] [CrossRef]
- Fisher, R.A. Dispersion on a sphere. Proc. R. Soc. Lond. 1953, 217, 295–305. [Google Scholar] [CrossRef]
- McFadden, P.L.; McElhinny, M.W. Classification of the reversal test in palaeomagnetism. Geophys. J. Int. 1990, 103, 725–729. [Google Scholar] [CrossRef] [Green Version]
- Opdyke, N.D.; Channel, J.E.T. Magnetic Stratigraphy; Academic Press: San Diego, CA, USA, 1996. [Google Scholar]
- Eltgen, H. Feinstratigraphisch-Fazielle Untersuchungen an Pliozän-Sedimenten im Tertiärbecken Südlich Neapolis/Kozani, Nordgriechenland; Institouton geōlogias kai ereunōn upedafous: Athens, Greece, 1986; pp. 107–115. [Google Scholar]
- Fountoulis, I.; Markopoulou-Diakantoni, A.; Mpakopoulou, A.; Moraiti, E.; Mikrou, M.-P.; Saroglou, X. The presence of marine Pliocene sediments in the Messohelenic trough (Pramoritsa banks, Grevena, Greece). Bull. Geol. Soc. Greece 2001, 24, 603–612. [Google Scholar]
- Guérin, C.; Tsoukala, E. The Tapiridae, Rhinocerotidae and Suidae (Mammalia) of the Early Villafranchian site of Milia (Grevena, Macedonia, Greece). Geodiversitas 2013, 35, 447–489. [Google Scholar] [CrossRef]
- Lazaridis, G.; Tsoukala, E.; Maul, L.C.H. The earliest Hystrix refossa: A new Early Villafranchian record from Milia (Grevena, Macedonia, Greece). Hystrix 2019, 30, 12–18. [Google Scholar]
- Koufos, G.D. The Villafranchian mammalian faunas and biochronology of Greece. Boll. Soc. Paleontol. Ital. 2001, 40, 217–223. [Google Scholar]
- Sotnikova, M.; Rook, L. Dispersal of the Canini (Mammalia, Canidae: Caninae) across Eurasia during the Late Miocene to Early Pleistocene. Quat. Int. 2010, 212, 86–97. [Google Scholar] [CrossRef]
- Madurell-Malapeira, J.; Ros-Montoya, S.; Espigares, M.P.; Alba, D.M.; Aurell-Garrido, J. Villafranchian large mammals from the Iberian Peninsula: Paleobiogeography, paleoecology and dispersal events. J. Iber. Geol. 2014, 40, 167–178. [Google Scholar] [CrossRef] [Green Version]
- Bellucci, L.; Bona, F.; Corrado, P.; Magri, D.; Mazzini, I.; Parenti, F.; Scardia, G.; Sardella, R. Evidence of late Gelasian dispersal of African fauna at Coste San Giacomo (Anagni Basin, central Italy): Early Pleistocene environments and the background of early human occupation in Europe. Quat. Sci. Rev. 2014, 96, 72–85. [Google Scholar] [CrossRef]
- Cohen, K.M.; Gibbard, P.L. Global chronostratigraphical correlation table for the last 2.7 million years, version 2019 QI-500. Quat. Int. 2019, 500, 20–31. [Google Scholar] [CrossRef]
- Ferring, C.R. Rates of Fluvial Sedimentation: Implications for Archaeological Variability. Geoarchaeology 1986, 1, 259–274. [Google Scholar] [CrossRef]
- Sen, S. Magnetostratigraphie et taux de sédimentation: Quelques données sur les dépôts fluviatiles, lacustres et marins du Néogène mediterranéen. Bull. Société Géol. France 1988, 4, 161–166. [Google Scholar] [CrossRef]
- Sen, S.; de Bonis, L.; Dalfes, N.; Geraads, D.; Koufos, G.D. Les gisements de mammifères du Miocène supérieur de Kemiklitepe, Turquie: Stratigraphie et magnetostratigraphie. Bull. Mus. Nat. Hist. Nat. Paris 1994, 16, 5–17. [Google Scholar]
- Arribas, A.; Garrido, G.; Viseras, C.; Soria, J.M.; Pla, S.; Solano, J.G.; Garcés, M.; Beamud, E.; Carrió, J.S. A Mammalian Lost World in Southwest Europe during the Late Pliocene. PLoS ONE 2009, 4, e7127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Navarro, B. Early Pleistocene faunas of Eurasia and hominid dispersals. In Out of Africa I: The First Hominin Colonization of Eurasia; Vertebrate Paleobiology and Paleoanthropology Series; Fleagle, J.G., Shea, J.J., Grine, F.E., Baden, A.L., Leakey, R.E., Eds.; Springer Press: Amsterdam, The Netherlands, 2010; pp. 207–224. [Google Scholar]
- Spassov, N. Zorillas (Carnivora, Mustelidae, Ictonychini) from the Villafranchian of Bulgaria with a description of a new species of Baranogale Kormos, 1934. Geodiversitas 2001, 23, 87–104. [Google Scholar]
- Spassov, N. The Plio-Pleistocene vertebrate fauna in South-Eastern Europe and the megafaunal migratory waves from the east to Europe. Revue Paleobiol. 2003, 22, 197–229. [Google Scholar]
- Popescu, S.-M.; Biltekin, D.; Winter, H.; Suc, J.-P.; Melinte-Dobrinescu, M.C.; Klotz, S.; Rabineau, M.; Combourieu-Nebout, N.; Clauzon, G.; Deaconu, F. Pliocene and Lower Pleistocene vegetation and climate changes at the European scale: Long pollen records and climatostratigraphy. Quat. Int. 2010, 219, 152–167. [Google Scholar] [CrossRef]
- Rivals, F.; Athanassiou, A. Dietary adaptations in an ungulate community from the late Pliocene of Greece. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2008, 265, 134–139. [Google Scholar] [CrossRef]
- Berlioz, E.; Kostopoulos, D.S.; Blondel, C.; Merceron, G. Feeding ecology of Eucladoceros ctenoides as a proxy to track regional environmental variations in Europe during the early Pleistocene. CR Palevol. 2018, 17, 320–332. [Google Scholar] [CrossRef]
- Hermier, R.; Merceron, G.; Kostopoulos, D.S. The emblematic Eurasian Villafranchian antelope Gazellospira (Mammalia: Bovidae): New insights from the Lower Pleistocene Dafnero fossil sites (Northern Greece). Geobios 2020, in press. [Google Scholar] [CrossRef]
- De Vos, J.; van der Made, J.; Athanassiou, A.; Lyras, G.; Sondaar, P.Y.; Dermitzakis, M.D. Preliminay note on the late Pliocene fauna from Vatera (Lesvos, Greece). Ann. Geol. Pays Hell. 2002, 39, 37–70. [Google Scholar]
- Athanassiou, A. A Villafranchian Hipparion-bearing mammal fauna from Sésklo (E. Thessaly, Greece): Implications for the question of Hipparion–Equus sympatry in Europe. Quaternary 2018, 1, 12. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primates | Perissodactyla |
|---|---|
| Paradolichopithecus aff. arvernensis | Stephanorhinus ex gr. etruscus |
| Carnivora | Equus stenonis (‘vireti’ morphotype) |
| Ursus etruscus | Giraffidae |
| Nyctereutes megamastoides | Palaeotragus inexspectatus |
| Vulpes alopecoides | Cervidae |
| Baranogale aff. helbingi | Metacervocerus rhenanus |
| Chasmaporthetes lunensis | Eucladoceros ctenoides |
| Homotherium latidens | Bovidae |
| Lynx issiodorensis | Gallogoral meneghinii |
| Rodentia | Gazellospira torticornis |
| Castor sp. | Gazella bouvrainae |
| Insectivora | Capra? sp. |
| Erinaceus sp. |
| Main Large Mammal Taxa | DFN | SES | VTR | VOL |
|---|---|---|---|---|
| Paradolichopithecus arvernensis | aff | |||
| Anancus arvernensis | ||||
| Mammuthus meridionalis | cf. | |||
| Nyctereutes megamastoides | ||||
| Vulpes alopecoides/praecorsac | cf. | |||
| Ursus etruscus | cf. | |||
| Baranogale helbingi | cf. | |||
| Chasmaporthetes lunensis | ||||
| Pliocrocuta perrieri | ? | |||
| Homotherium latidens | sp. | |||
| Megantereon megantereon | ||||
| Lynx issiodorensis | ||||
| Meles thorali | ||||
| Stephanorhinus ex gr. etruscus | sp. | cf. | indet. | |
| Equus stenonis ‘vireti group’ | ||||
| Palaeotragus inexspectatus | cf. | |||
| Eucladoceros ctenoides | ||||
| Metacervocerus rhenanus | aff. | cf. | ||
| Croizetoceros ramosus | ||||
| Leptobos sp. | ||||
| Gallogoral meneghinii | ||||
| Gazellospira torticornis | cf. | cf. | ||
| Gazella borbonica | aff. | |||
| Gazella bouvrainae | cf. | |||
| Gazella aegea | ||||
| Euthyceros thessalicus | ||||
| Procamptoceras | ? |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benammi, M.; Aidona, E.; Merceron, G.; Koufos, G.D.; Kostopoulos, D.S. Magnetostratigraphy and Chronology of the Lower Pleistocene Primate Bearing Dafnero Fossil Site, N. Greece. Quaternary 2020, 3, 22. https://0-doi-org.brum.beds.ac.uk/10.3390/quat3030022
Benammi M, Aidona E, Merceron G, Koufos GD, Kostopoulos DS. Magnetostratigraphy and Chronology of the Lower Pleistocene Primate Bearing Dafnero Fossil Site, N. Greece. Quaternary. 2020; 3(3):22. https://0-doi-org.brum.beds.ac.uk/10.3390/quat3030022
Chicago/Turabian StyleBenammi, Mouloud, Elina Aidona, Gildas Merceron, George D. Koufos, and Dimitris S. Kostopoulos. 2020. "Magnetostratigraphy and Chronology of the Lower Pleistocene Primate Bearing Dafnero Fossil Site, N. Greece" Quaternary 3, no. 3: 22. https://0-doi-org.brum.beds.ac.uk/10.3390/quat3030022