Effect of Acute Seawater Temperature Increase on the Survival of a Fish Ectoparasite
by
 , and
, and
Mary O. Shodipo
1 ,
,
Berilin Duong
2,
Alexia Graba-Landry
3,
Alexandra S. Grutter
2 and
Paul C. Sikkel
4,5,* 1
Institute of Environmental and Marine Sciences, Silliman University, 6200 Dumaguete City, Negros Oriental, Philippines
2
School of Biological Sciences, The University of Queensland, St Lucia QLD 4072, Australia
3
ARC Centre of Excellence for Coral Reef Studies, James Cook University, Townsville QLD 4811, Australia
4
Department of Biological Sciences and Environmental Sciences Program, Arkansas State University, Jonesboro, AR 72467, USA
5
Water Research Group, Unit for Environmental Science and Management, North-West University, Potchefstroom 2520, South Africa
*
Author to whom correspondence should be addressed.
Oceans 2020, 1(4), 215-236; https://0-doi-org.brum.beds.ac.uk/10.3390/oceans1040016
Submission received: 17 July 2020
/
Revised: 25 September 2020
/
Accepted: 27 September 2020
/
Published: 4 October 2020
(This article belongs to the Special Issue The Future of Coral Reefs: Research Submitted to ICRS 2020, Bremen, Germany)
Abstract
:Extreme warming events that contribute to mass coral bleaching are occurring with increasing regularity, raising questions about their effect on coral reef ecological interactions. However, the effects of such events on parasite-host interactions are largely ignored. Gnathiid isopods are common, highly mobile, external parasites of coral reef fishes, that feed on blood during the juvenile stage. They have direct and indirect impacts on their fish hosts, and are the major food source for cleaner fishes. However, how these interactions might be impacted by increased temperatures is unknown. We examined the effects of acute temperature increases, similar to those observed during mass bleaching events, on survivorship of gnathiid isopod juveniles. Laboratory experiments were conducted using individuals from one species (Gnathia aureamaculosa) from the Great Barrier Reef (GBR), and multiple unknown species from the central Philippines. Fed and unfed GBR gnathiids were held in temperature treatments of 29 °C to 32 °C and fed Philippines gnathiids were held at 28 °C to 36 °C. Gnathiids from both locations showed rapid mortality when held in temperatures 2 °C to 3 °C above average seasonal sea surface temperature (32 °C). This suggests environmental changes in temperature can influence gnathiid survival, which could have significant ecological consequences for host-parasite-cleaner fish interactions during increased temperature events.
1. Introduction
Among the myriad anthropogenic impacts on the world’s oceans, perhaps the most significant is the increase in temperature associated with production of greenhouse gases [1]. This warming is responsible for large-scale changes in circulation and storm activity through melting of glaciers, warming of air masses, and increased evaporation and salinity [1], and as such, warming may have an indirect effect on marine organisms. However, the majority of marine organisms are ectothermic, and are therefore dependent on environmental temperature to gain adequate energy for their own biological functions. The relationship between the performance of an ectotherm and temperature is non-linear, where performance gradually increases with temperature until it reaches a thermal optimum after which it rapidly declines (Thermal Performance Curve: [2]). Hence, the effect of increasing temperature on marine ectotherms may be more direct, affecting physiology and metabolism [3,4,5,6,7], which may have implications for growth, motor function, development, reproduction and behaviour [6,7,8,9,10,11,12,13,14,15], which in turn may impact species’ abundance and distribution [16]. Therefore, increasing temperatures may be the most pervasive climate change factor influencing marine organisms [7,17,18]. Warming can subsequently impact entire ecological communities and ecosystems by differentially impacting individuals and functional traits [19,20,21].
Coral reefs are one of the most biodiverse ecosystems in the world [22,23]. Even though the rate of increase in sea surface temperature (SST) is 30% less in tropical oceans than the global average [24], coral reefs are also among the most sensitive ecosystems to changes in environmental conditions [22,25,26], and thus, are particularly at risk of thermal stress. Tropical ectotherms have a narrow thermal tolerance range, and their thermal optimum is close to their thermal maximum, as they have evolved under relatively stable thermal conditions [27,28]. As SSTs rise, corals and coral reef associated organisms are being subjected to higher temperatures (29–31 °C) for increasing periods of time [24]. As a consequence many tropical organisms are thought to be living at or close to their thermal limits [29]. SSTs are predicted to continue to rise over the coming years [1] and extreme warming events, resulting in global scale coral bleaching, are occurring with increasing regularity and severity [30,31,32,33,34], causing degradation of coral reef habitats [22,30,31,35,36].
In addition to the corals themselves, research on the effects of marine heatwaves has also focused heavily on fishes [37,38,39,40,41,42,43,44,45,46,47,48], which are also typically included in coral reef monitoring efforts. However, studies have almost completely ignored the myriad of small, cryptic, species, which make up a disproportionate amount of coral reef biodiversity [49,50,51]. One such group are parasites, which make up the largest consumer strategy globally [52] and comprise an estimated 40% of global biodiversity [53,54,55,56]. In addition to host behavior, physiology, and population dynamics, parasitic organisms have been shown to have impacts on interspecific interactions, energy flow, and the structure, ecology, and biodiversity of communities [55,57,58,59,60]. Parasites are particularly diverse on coral reefs [61] with an estimate of over 20,000 species on the Great Barrier Reef (GBR) alone [62]. However even with such a large presence in coral reef communities, they are significantly underrepresented in ecological studies [10,55,63]. Coral reef parasites are also ectothermic, and as such, may be affected by changes to their environmental temperature [10,11,64,65,66,67]. Some parasites are ectoparasitic and would be highly vulnerable to increased temperatures. Ectoparasites are also likely to be directly impacted by the temperature itself, in addition to being indirectly affected through changes in community structure due to temperature impacts on hosts [68] and other organisms. For example, the life cycle of a monogenean ectoparasite (Neobenedinia) was faster and the life span of their larvae (oncomiracidia) decreased as temperatures increased from 22 °C to 34 °C [69].
Gnathiid isopods are one of the most common ectoparasites in coral reef habitats [70,71,72]. They are small crustaceans, typically 1–3 mm long, that do not permanently live on their fish hosts [73,74]. In fact, with few exceptions, they associate only long enough to extract a blood meal and may therefore also be referred to as “micropredators” [75,76]. After feeding on tissue and blood from their fish host they return to the benthos to molt and progress to the next developmental stage [73,74]. They are only parasitic during their three juvenile stages, and no longer feed once they metamorphose into an adult. Gnathiids can have significant impacts on their hosts [66]. Direct effects include influencing behavior [77,78,79,80,81,82,83], physiology [84], and mortality [85]. Indeed, as few as one gnathiid can kill a young juvenile fish [83,86,87,88,89,90]. Indirect effects include transmission of blood-borne parasites [91] and wounds that can facilitate infection [92]. Gnathiids are also the most common items in the diet of many cleaner fishes, including Labroides dimidiatus [93,94], a species with far-reaching ecosystem effects [95,96,97]. Indeed, environmental perturbations, including a coral bleaching event with water temperatures reaching up to 30 °C, resulted in an 80% decline in L. dimidiatus at Lizard Island, GBR [48]. However, the processes leading to this decline remain unknown.
A long-term monitoring study of gnathiid isopods off Lizard Island, GBR, revealed a significant decrease in gnathiid abundance during extreme warm-water periods associated with bleaching events, compared with cooler periods in the same year or during non-bleaching years [65]. However, the cause of this decline was unclear. Sikkel et al. (2019) [65] hypothesized that the direct effects of temperature on gnathiid mortality may have partly contributed to the decline in gnathiid abundance. The aim of this study therefore, was to assess the direct effect of a rapid increase in seawater temperatures on mortality of shallow-reef gnathiid isopods. By conducting laboratory experiments on gnathiids in two coral reef regions subject to bleaching, GBR, Australia [31,32,98], and Philippines [99,100,101], we show that a rapid increase in temperature causes significant increases in mortality.
2. Materials and Methods
2.1. Study Sites
This study was conducted between January and February 2018 at the Lizard Island Research Station (LIRS), northern GBR and between July and October 2017 at the Silliman University–Institute of Environmental and Marine Sciences (SU-IEMS), Dumaguete City, Negros Oriental, Visayas, Philippines.
2.2. Gnathiid Collection
For the GBR study, gnathiids were obtained from a culture maintained at LIRS since 2001, which uses the continual availability of wrasse Hemigymnus melapterus (Labridae) as hosts [102]. The culture is outdoors and uses a flow-through seawater system that obtains water directly from the nearby reefs. The previous exposures of the experimental (and previous generations) of gnathiids would have, therefore, reflected similar temperatures to the ocean and land ones [102].
Gnathiids for the Philippines study were collected from the shallow fringing coral reefs (<10 m) of Cangmating reef (9°21′18.38″ N, 123°17′58.91″ E) and Agan-an reef (9°20′2.6″ N, 123°18′41.5″ E) in Sibulan and from Bantayan reef (9°19′49.22″ N, 123°18′43.43″ E) in Dumaguete City, all within Negros Oriental Province. The Bantayan reef has small patch reefs with inshore seagrass beds. Cangmating and Agan-an have larger patch reefs and inshore seagrass beds. Gnathiids are common at all three sites [103,104]. Gnathiids were collected using light traps, adapted from Artim et al. (2015) [89] and Artim and Sikkel (2016) [105]. The traps were set at dusk and retrieved the following morning and then transported by boat to the SU-IEMS laboratory where they were emptied into individual 10 L plastic buckets with aerators. The contents of each trap were filtered with a funnel and 55 μm plankton mesh. The gnathiids were then sorted using a stereoscope and placed in an aquarium (27 L) with fresh, filtered, aerated seawater. The species of gnathiids collected were unknown due to difficulty with species identification of the juvenile stages [73], and the fact that no species have yet been formally described from our Philippines study region.
2.3. Experimental Protocol
At both locations, gnathiid mortality was defined as the absence of detectable movement, even after disturbance (e.g., by moving the vial it was held in while viewing it under the microscope).
2.3.1. Great Barrier Reef
Gnathiids, all belonging to the species Gnathia aureamaculosa, were collected from the culture in the morning and afternoon, and placed together into 75 mL holding containers filled with seawater.
They were collected by moving a black tray (35 × 25 × 5 cm) up the side of the gnathiid culture tank and were removed using a pipette. From the holding containers, gnathiids were individually transferred into 5 mL vials that were half-filled with seawater. These vials were then individually labelled. Collecting and processing took approximately 2 to 4 h, depending on the catch size of the day (ranging from 9 to 226 gnathiids). The daily number varied as a result of fluctuations in the number that were active, most likely due to normal high variation in their population dynamics [102]. A mixture of fed and unfed gnathiids was used, and it was not known how much time elapsed since the last feeding. Gnathiids were not fed for practical reasons. After processing the gnathiids, the vials were randomly allocated, in a balanced way (approximate equal number), to a temperature treatment and aquarium replicate combination; there were three aquarium replicates per temperature treatment. Vials were labelled with a unique number across all replicates. Only the lids of the vials were labelled, reducing any potential bias when viewing them under the microscope. It also made it easier to monitor and return them to their respective treatment and aquarium daily. Vials were held underwater in plastic baskets (17 × 17 × 10 cm), one for each treatment and replicate (n = 9 baskets). Baskets had four mesh (1 mm2) windows (12 × 5 cm) on the sides and one on the lid (12 × 12 cm) to allow for flow of water. A dive weight was used to submerge the baskets. Aquaria were supplied with flow-through seawater, with seawater that was either chilled or heated in a sump under the aquarium benches and pumped up to the aquaria. Each bench had a different temperature treatment and held 10 aquaria (previously used for another experiment, see Graba-Landry et al. 2020 [106]). Three aquaria were randomly selected per bench and allocated to replicates.
We estimated the predicted ambient seawater temperature (29.25 °C ± 0.013 SE) based on the Australian Institute of Marine Science long-term average water temperature for February [107] (Figure S1). Actual average daily seawater temperature during the experiment was 29.0 °C ± 0.67 SE (February 1 to 23, 2018 available only). The temperature of the water that gnathiids had been maintained in throughout their lifetimes was not available. However, the temperature of incoming water from the station’s holding tanks was on average 1.4 °C warmer than the ocean, when sampled at two sources at three times of the day (09:00, 15:00 and 21:00 h from 15 to 20 October, 2018) relative to the same period in the ocean [108].
Temperature was manipulated in an outdoor seawater flow-through system at LIRS using purpose built 1KW steel bar heaters and chillers (Teco®) in a header or sump tank. Each sump, one per temperature treatment, fed replicate 40 L tanks with the appropriate experimental flow-through seawater using 1000 L hr−1 pumps (Eheim®) at a rate of approximately 1 L minute−1. Tanks were wrapped in Insulbreak® insulation to stabilize water temperatures. Temperature (±0.1 °C) was also measured at 12:00 h daily from each of the nine tanks housing the gnathid cultures/vials using a portable temperature probe (Comark®) calibrated to 26 °C, 28 °C and 30 °C (National Association of Testing Authorities certified) to ensure temperature remained stable across treatments. Experimental temperatures at 12:00 h per treatment, averaged across the means of the three replicate aquaria, were: 29 °C: 29.5 °C ± 0.07 SE, 31 °C: 31.4 °C ± 0.02 SE, and 32 °C: 32.6 °C ± 0.02 SE. Temperatures were very similar among replicates within a treatment, (Figure S2). One calibrated temperature logger (HOBO Pendant temperature/light logger, UA-002-08) per treatment also recorded the temperature every 2 h throughout the course of the study to account for diurnal fluctuations in temperature [mean (SE) per treatment: 29 °C: 28.9(0.042); 31 °C: 31.2(0.035): 32 °C: 31.9(0.06); Figure S3].
Each day, one random basket from each treatment was removed (to reduce time exposed to air temperature). The vials from each basket were rinsed in freshwater and placed in a large tub (all treatments were examined together to avoid bias). Each vial was then examined under a dissection microscope to check for gnathiid mortality. Vials with alive gnathiids were sorted back into their respective treatments/replicates and placed back into the aquaria. This was repeated for each of the remaining baskets from each treatment-replicate combination. Gnathiids were monitored until all had died (except for four survivors, see Results for details). Vials with dead gnathiids were preserved for later to undertake headwidth measurements, by adding a few drops of formalin into the seawater.
2.3.2. Philippines
Unfed juvenile gnathiids were given one day to acclimatize in an aquarium after collection, before host fish, Dascyllus trimaculatus (Pomacentridae) and various species of Labridae, were placed in the aquarium overnight to allow them to feed. The gnathiids did not feed again for the duration of the experiment. The following day, fed, mobile and healthy-looking individuals were selected. However, as the gnathiids’ species and therefore the consequent size range for each stage was not known, they could not be separated by juvenile stage as in the GBR study. Instead they were sorted into two size classes (<2 mm and >2 mm, to account for any effect of size of the gnathiid on its molting rate and survival.
Gnathiids were then placed in 270 mL plastic containers, 5–10 gnathiids per container, with plankton mesh (55 µm) secured on the top, and the containers were submerged in one of five 27 L aerated aquaria, each with a different set temperature. Each container was labelled with the size class, treatment, trial and replicate. The first trial consisted of 5 temperature treatments, ambient (28 °C), 30 °C, 32 °C, 34 °C and 36 °C. A second trial was also conducted to obtain a finer resolution of the temperature effect, with treatments of 30 °C, 32 °C, 33 °C, 34 °C and 35 °C. Aquaria were individually heated gradually with 100W and 200W aquarium electric heaters (Venusaaqua®) over a 10 h period to their desired temperature. Temperature readings were taken daily with an aquarium-mounted digital thermometer (Doutop®) to ensure the desired water temperatures were maintained and to calculate the average temperature for each treatment per trial (Table S1a,b). Containers from each treatment were inspected daily for evidence of changes in gnathiid development and mortality. Dead gnathiids were removed from the containers and molted adult males were preserved in ethanol for future species identification. The experiment was concluded for each treatment when all gnathiids were dead or when 20 d had passed. One third of the water in each aquarium was removed daily and replaced with ambient temperature, filtered, fresh seawater. Over the 20 d duration of each replicate the aquaria used for each temperature treatment was alternated every week (once per replicate) to ensure there were no confounding effects associated with individual aquaria. Three replicates of each temperature treatment were run for each trial. Trials 1 (n = 369 gnathiids) and 2 (n = 318 gnathiids) had a range of 60–76 and 59–68 gnathiids per temperature treatment respectively (Table S2a,b).
The baseline ambient temperatures for Trials 1 (28 °C) and 2 (30 °C) were similar to the SST in the Bohol Sea, Philippines, which fluctuated by about 3 °C (about 27–30 °C) during 2017; the SST during the experiment (July–October 2017) averaged at 30 °C ± 0.04 SE. [109].
2.4. Statistical Analyses
2.4.1. Great Barrier Reef
One live unfed gnathiid (stage three, 15 d), two adult males (alive, 15 d; dead, 29 d) and three adult females (alive, 15 and 24 d; dead, 16 d) were excluded from the data. These gnathiids were excluded because they were adults and thus their longevity would be different to that of the juveniles. The single juvenile that was still alive when we terminated the experiment was omitted for simplicity and consistency. We categorized the three juvenile stages based on their headwidth (stage one: 0.14–0.2, stage two: 0.21–0.24, stage three: 0.25–0.32 mm) [77]. Due to the large difference in sample size per unfed/fed status (based on the presence of an engorged gut), we conducted separate analyses for unfed (n = 1133) and fed gnathiids (n = 87).
To test whether survival of gnathiids differed among temperature treatments, we used a proportional hazards Cox mixed-effects model with temperature treatment and gnathiid juvenile stage as categorical fixed effects, aquarium as a random factor, and gnathiid headwidth as a covariate (Table 1 and Table S3). We used ambient temperature (29 °C) and juvenile stage one as the baselines for the analyses. We used the function “coxme” in the package “coxme” [110,111] and function “Anova” in the package “car” [112]. We tested the Cox model assumption of proportionality using the Global test statistic in the function “coxph” and “cox.zph” in the package “coxme” and graphically using a smoothed spline plot of the Shoenfeld residuals relative to time (see Tables S3 and S4 for results and Figures S4 and S5 for spline plots).
2.4.2. Philippines
In the size class < 2 mm (n = 168 gnathiids) for Trial 1, five (3%) gnathiids molted into adult females and nine (5%) into males. In the size class < 2 mm (n = 153) for Trial 2, no gnathiids molted into adults. In contrast, for the larger size class > 2 mm for Trial 1 (n = 201), 20 (10%) gnathiids molted into females and 85 (42%) into males. In the size class > 2 mm (n = 165) for Trial 2, 37 (22%) gnathiids molted into females and 68 (41%) into males. In this study, newly metamorphosed males were first observed after day 1 in Trial 2 and day 2 in Trial 1, and no additional males appeared after day 5 in Trial 2 and day 7 in Trial 1. The mean number of days juvenile gnathiids molted into males for all treatments was 3.39 ± 0.75 and 3.23 ± 0.16 for Trials 1, and 2, respectively.
To test whether survival of gnathiids differed among temperature treatments, we used the same statistical methods as for the GBR data, with some modifications to the model. Temperature treatment was a fixed effect, and size class, life stage (male, female, or juvenile) and container (which the gnathiids were kept in) were treated as random effects. The ambient temperature of 28 °C was used as the baseline for analysis for Trial 1 and 30 °C for Trial 2. Assumptions of proportionality were met for both analyses (both Global tests: p > 0.05, see Tables S5 and S6 for results and Figures S6 and S7 for spline plots).
2.5. Ethics
All applicable international, national, and/or institutional guidelines for the care and use of animals were followed. All procedures performed in studies involving animals were in accordance with the ethical standards of Silliman University, Arkansas State University, The University of Queensland, and the Government of Australia and the Philippines.
3. Results
3.1. Great Barrier Reef
3.1.1. Unfed Gnathiids
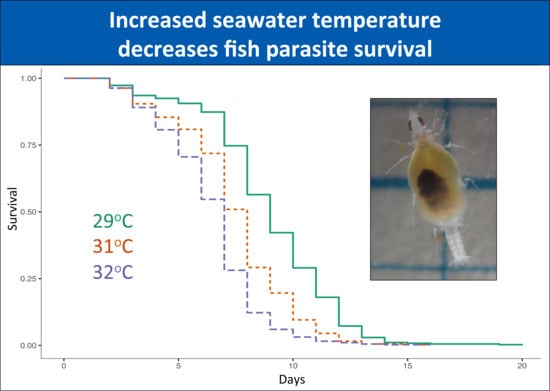
From the 1220 gnathiids whose survival was followed over time, 93% were unfed individuals. For unfed individuals, the numbers of gnathiids per stage and per temperature was relatively even within each of the 29 °C, 31 °C, and 32 °C temperature treatments (stage one: 186, 198, 210; stage two: 73, 66, 71; stage three: 113, 113, 103, respectively). There was a significant effect of temperature on gnathiid survival (p < 0.0001, Table 1, Figure 1), due to a significantly lower survival at 32 °C compared with the 29 °C baseline temperature (p = 0.016, Table S7). The interaction between gnathiid headwidth and juvenile stage was significant (p < 0.0001, Table 1); when further explored separately by stage, the association was largely due to a weakly positive relationship between gnathiid survival and gnathiid headwidth in stage one (p < 0.0001, Table S9a).
3.1.2. Fed Gnathiids
For fed individuals, the numbers of gnathiids per juvenile stage and per temperature were also relatively even between the 29 °C, 31 °C, and 32 °C temperature treatments (stage one: 8, 6, 6; stage two: 17, 15, 13; stage three: 8, 7, 7, respectively). Three (3.4%) contained more blood than clear material (i.e., plasma) in their gut, the remainder had a clear gut. Of the 87 individuals followed, 60% had molted during the course of the study. Survival differed according to an interaction between temperature and juvenile stage (p = 0.0085, Table 2); when further explored separately by stage (Figure 2), the effect of temperature was significant for stage two (p < 0.0001), and three (p = 0.0009, Figure 2b,c, Table S10b,c), with the strongest effect of temperature being that between the baseline (29 °C) and the 32 °C treatments for stage three (Table S10c, Figure 2c). Survival differed according to an interaction between temperature and headwidth (p = 0.0480, Table 2); when further explored separately by temperature treatment, the effect of headwidth was largely due to non-significant weakly positive relationships between survival and headwidth at 29 °C (p = 0.0758) and 32 °C (p = 0.0718, Table S11a,c).
3.2. Philippines
3.2.1. Trial 1
3.2.2. Trial 2
4. Discussion
With ocean temperatures predicted to rise 3 °C by the end of the century [113], the effects of ocean warming on coral reef organisms have received an increasing amount of attention. However, such studies largely ignore the cryptofauna that comprises most of coral reef’s biodiversity and biomass, including parasites [67]. In the only long-term monitoring study of any marine parasitic crustacean, Sikkel et al. (2019) [65] reported that during extreme warm-water events in the GBR parasitic gnathiid isopod populations crashed. The findings reported here are consistent with their hypothesis that this may be attributable, in part, to a direct effect of temperature on gnathiid mortality. Such an effect of temperature on the larvae of a tropical ectoparasite has been shown for monogeneans on farmed tropical fish [69].
The present study is the first to examine effects of acute temperature increases on this common reef fish ectoparasite. In our study, gnathiids from both the GBR in Australia and Negros Oriental in the Philippines demonstrated rapid mortality in temperatures raised to above average SST, suggesting that environmental changes in temperature can influence gnathiid survival. In the Philippines, temperatures as little as 2 °C (i.e., 32 °C) above average seasonal SST (30 °C) caused significantly lower survival, with increasingly steep survival curves at 35 °C, with the steepest at 36 °C, where no gnathiids survived past five days. Unfed gnathiids on the GBR had lower survival at 32 °C compared with 29 °C, an effect which was consistent across all three juvenile stages. For fed gnathiids on the GBR, the effect of temperature was significant for juvenile stages two and three, with the strongest effect of temperature on stage three, also between the 32 °C and 29 °C treatments. It is, therefore, likely that gnathiids from both the Philippines and Australia may be living near their thermal limit, as small increases in temperature from the annual seasonal mean have resulted in increased mortality in organisms from both regions. These results indicate not only that an acute change of temperature to just 32 °C decreases the survival of gnathiids, but that the effect of increased temperature is greater on the larger juvenile stages.
In the GBR we found evidence that greater gnathiid headwidth, not just juvenile stage, increased gnathiid survival. For unfed gnathiids, there was a weakly positive relationship between gnathiid survival and gnathiid headwidth, but only in juvenile stage one. For fed gnathiids, there was also a weakly positive relationship between survival and headwidth, but only at 29 °C and 32 °C. Gnathiid length is correlated with headwidth [95,105] and thus likely with mass also. Both results suggest that even small increases in gnathiid size within a juvenile stage can increase gnathiid survival; these findings also supported our decision to include both headwidth and stage in the statistical model as being important factors to consider when modelling gnathiid juvenile survival. Such a difference in the thermal response related to size may be due to the increased metabolic demand caused by the increase in temperature, an effect which may lead to an energetic deficit for smaller individuals if enough food cannot be obtained, therefore, creating a metabolic mismatch between energy obtained versus energy required [114]. Alternatively, there may possibly be a higher baseline metabolism or higher growth rate at smaller sizes, which then slows down as they reach the maximum size for that stage, resulting in smaller sizes using up their reserves faster than larger sizes. Furthermore, the energetic demands of development may also differ among juvenile life-history stages. Thus increased metabolic demand for basal processes (such as cell maintenance) as a result of increases in temperature, coupled with differential energetic requirements for development may also explain the variation in the thermal response among juvenile stages in our study. Therefore, understanding the effect of increasing temperature on individual metabolism and survival also requires an understanding of food resources and availability [114].
Overall, these results suggest that even with a small increase of 2–3 °C above the normal ambient mean, raised temperature can ultimately lead to increased gnathiid mortality. However, there were some key differences in the experimental protocol between the GBR and Philippines studies that should be considered. First, only one species of gnathiid was used in the GBR experiment, compared to at least three (all undescribed, M.O.S. personal observation) species present in the pool of gnathiids used for the Philippines experiment. Second, because these species were unknown, we were unable to confidently separate juvenile gnathiids into their different stages, and so used size class as a proxy. Therefore, we cannot discount the possibility of some among-species and life-stage variation in thermal tolerance. Finally, in contrast to the Philippines, in the GBR study the time the gnathiids were last fed was unknown. This would account for much of the unexplained variation in survivorship in the analysis of the GBR data, as the variation in resources available to the gnathiid, in the form of a blood meal, would decrease over time since their last feeding event. In addition, it should be noted that, in both studies, the gnathiids were not fed for the duration of the experiment, and thus starvation may have been a contributing cause of mortality. While starvation may have influenced mortality of gnathiids among the treatments (as suggested by increased mortality over time in the ambient temperatures), the rates of mortality at higher temperatures were greater, with rapid mortality taking place very early on in the experiments (e.g., one to five days in the Phillipines). This supports the interpretation that increased temperatures influence gnathiid survival directly. It is of relevance that marine “heatwaves” (which are categorised as periods of abnormally high SST lasting for longer than five days [115]) have been predicted to become more frequent, longer and more severe [116,117]. Our observations of rapid gnathiid mortality even after just one day suggests there may be a decline in gnathiid survival from early on in a heatwave, so that gnathiid populations may be heavily impacted if there are more frequent and severe heatwaves in the future.
Our findings appear consistent with data for other tropical marine invertebrates, which have an upper thermal tolerance that is not far above normal sea temperature (reviewed in [4,118]). For example, in a meta-analysis on bivalves, and a study on porcelain crabs, tropical species were found to have upper thermal limits that were closer to the maximum temperature of their habitat than temperate species [118,119]. Tropical species of bivalves have also been shown to have a smaller thermal tolerance window than temperate species [118]. This is thought to be due to tropical marine organisms being more sensitive to changes in temperature as they have evolved under relatively invariable thermal conditions [27]. Other studies on marine invertebrates have also shown increased mortality with high SST [120,121], with hermatypic corals being particularly sensitive to increases, with SSTs needing to rise only a few degrees for bleaching to occur [25,26,31,122].
There are a number of studies investigating the potential impact of temperature increase associated with climate change on parasite communities and aquatic parasite-host interactions. The majority of these studies have been on endoparasitic trematodes from temperate regions [123,124,125]. Temperature was consistently observed to have a significant effect on the survival times of trematodes in their free-living juvenile stage, with survival rates decreasing as temperature increased (e.g., [126,127,128,129,130,131,132,133]). Similarly, temperature has been reported to have an effect on parasitic barnacles (rhizocephalans), with their prevalence decreasing at higher temperatures [68].
In one of the few other studies on ectoparasites, Conley and Curtis (1993) [134] found that, in temperatures of 8–20 °C, survival of copepodids was also inversely proportional to temperature. This same trend was observed in the survival rates of monogeneans, and isopods (Cymothoidae) in two studies in sub-tropical regions [135,136] and one study of monogeneans in a tropical region [69]. In all three studies, temperature treatments of 30 °C and above had the lowest survival rates [69,135,136]. Similar results were also observed with trematode cercariae from sub-tropical regions [137,138,139]. Summer temperatures for these lower latitudes parasites are in the range of 30–31 °C, which suggests that like gnathiids in the warmer months they are living close to their thermal limits.
Although, this study focused on effects of temperature on mortality, increased temperature can also have sub-lethal effects on marine organisms, impacting their ability to perform essential tasks [140,141,142]. Based on a review of the literature, Lough (2012) [24] suggested that temperatures between 30–32 °C may reflect a potential temperature threshold where a proportion of reef organisms’ physiological processes are negatively impacted. Higher temperatures may also affect the ability of parasites with mobile life history stages (such as gnathiids) to successfully detect and associate with a host. To our knowledge there are no studies that specifically examine this. However, the ability to physically reach a host by swimming does appear to be influenced by temperature. For temperate parasitic copepods in their free-living stage, the duration of swimming activity was found to be inversely related to water temperature [134]. For newly emerged cercariae, swimming speed increased in higher temperatures (19–36 °C). However, the speed declined over time, with rate of decline increasing with temperature. This resulted in higher swimming speeds, but for shorter durations in water of 30 °C and above [143]. In a sub-tropical study, cercariae infectivity also increased with temperature with maximum infectivity occurring at 30 °C before declining at 36 °C and 40 °C [137]. This could be attributable to greater cercariae swimming activity [137]. Although, we did not quantify the effects of temperature on movement, in the Philippines study it was apparent that gnathiids moved more slowly and less frequently at temperatures of 32 °C and above, with movement decreasing further as temperature increased, and also with apparent effects greater for the larger size class (M.O.S. personal observation).
Elevated temperatures may also impact host physiology, behavior and survival in ways that impact the balance between parasite and host. For parasitic barnacles (rhizocephalans), the effects of temperature on infected host mortality (and consequent transmission) could threaten their survival, with models showing that just an increase of 2 °C in ambient temperature could cause local parasite eradication [68]. In contrast, reef fishes can live further away from their thermal limits than gnathiids were observed to do in this study and in some cases can tolerate temperatures of up to 34–40 °C, [3,15,37,47]. However, they can still experience sub-lethal effects with smaller increases in temperatures [3,6,12,14,15,142,144,145,146], which could also impact host-parasite interactions.
Large hosts, like many reef fishes, can also leave areas of warm water for cooler water, or leave habitat impacted by coral bleaching for other habitats [39,40,43,44,45,46,147], depriving gnathiids and other similar ectoparasites, like natatory-stage cymothoid isopods of hosts [148]. The potentially impaired physiological and swimming ability of the parasite, combined with direct effects on mortality and host availability, could result in a decline in parasite populations. However, the ability of some gnathiids to feed on invertebrate hosts [149,150], combined with weakened immune response for the smaller, less mobile, fish species could leave fish more susceptible to ectoparasites, and thus, compensate for the loss of larger hosts. Indeed, during the 2016–2017 mass bleaching event on the GBR, there was a significant decrease in the numbers of larger, more mobile host fishes in shallow areas, with only smaller, site-attached species remaining [48]. This could have also contributed significantly to the crash in gnathiid populations observed by Sikkel et al. (2019) [65]. However, it should also be noted that as gnathiids are mostly free-living and have a temporary association with their hosts, they too can potentially avoid higher water temperatures. This might happen passively by the gnathiid “hitching a ride” whilst feeding on their host, a process which can last from a few minutes up to a few hours [73]. The gnathiids may, thereby, be transferred to different locations [151,152]. However, as knowledge of the dispersal mechanisms of gnathiids, the infection rates of host fish, and fish movements after disturbances is limited, the proportion of the gnathiid populations that could transfer location with their hosts remains unknown.
Another indirect effect of increased SST may be effects of warming on predators of gnathiids and other ectoparasites’ free-living stages. In particular, coral polyps are a major source of predation on juvenile gnathiids [153,154], and thus high coral mortality associated with warm-water events, combined with the loss of cleaner fish [48], which prey on ectoparasites [93], might increase living space and decrease predation on gnathiids. Indeed, once water cools following a bleaching event and most corals are dead, gnathiid populations appear to recover rapidly [65]. While, oceans are also experiencing increased acidification [1,155], Paula et al. (2020) [156] found no effect of acidification on the mortality of the same GBR gnathiid isopod as that studied here.
As parasites have a significant role in ecosystem function, changes in parasite abundance may pose consequences for ecological communities [157,158,159]. Therefore, while the diversity of coral reef parasites and their hosts makes it difficult to draw general conclusions on how warming events will impact parasite-host interactions, it remains important to further investigate parasite responces to both the direct and indirect effects of warming [67]. Future studies on gnathiids examining sublethal thermal effects on molting, physiology, locomotion, host-detecting mechanisms and reproductive performance will provide a more comprehensive understanding of effects of temperature on host-parasite interactions in coral reef systems.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/2673-1924/1/4/16/s1,. Table S1: Philippines; Average water temperature of aquaria for five treatments over three aquarium replicates, Table S2: Philippines; Sample size of larval gnathiid isopods < 2 mm and >2 mm in length in five different temperature treatments over three aquarium replicates, Table S3: Great Barrier Reef; Tests of proportionality, using function “cox.zph” in library “coxme” in R 3.2.5, for full model for unfed gnathiid survival among temperature treatments and juvenile stages, Table S4: Great Barrier Reef; Tests of proportionality, using function “cox.zph” in library “coxme”, for full model of fed gnathiid survival among temperature treatments and juvenile stages, Table S5: Philippines; Tests of proportionality, using function “cox.zph” in library “coxme” in R 3.2.5, for full model for Trial 1 gnathiid survival among temperature treatments, Table S6: Philippines; Tests of proportionality, using function “cox.zph” in library “coxme” in R 3.2.5, for full model for Trial 2 gnathiid survival among temperature treatments, Table S7: Great Barrier Reef; Summary output for full model for unfed gnathiid survival among temperature treatments and juvenile stages for Cox model, Table S8: Great Barrier Reef; Summary output for full model for fed gnathiid survival among temperature treatments and juvenile stages for Cox model, Table S9: Great Barrier Reef; Analysis of deviance tables (Type II tests) and summary outputs for unfed gnathiid survival for separate Cox models for each juvenile stage, Table S10: Great Barrier Reef; Analysis of deviance tables (Type II tests) and summary outputs for fed gnathiid survival for separate Cox models for each juvenile stage, Table S11: Great Barrier Reef; Analysis of deviance tables (Type II tests) and summary outputs for fed gnathiid survival for separate Cox models for each temperature treatment, Table S12: Philippines; Summary output for full model for Trial 1gnathiid survival among temperature treatments for Cox model, Table S13: Philippines; Summary output for full model for Trial 2 gnathiid survival among temperature treatments for Cox model. Bolded values are ones mentioned in main text, Figure S1: Seawater temperature for Great Barrier Reef data, Figure S2: Great Barrier Reef; Temperatures, measured using a handheld device at 12:00 h, for three replicate aquaria per temperature treatment, Figure S3: Great Barrier Reef; Water temperatures in an aquarium over duration of study for each of the temperature treatments between February 1 and March 2 2018, Figure S4: Great Barrier Reef; Scaled Shoenfeld residual plot for full model testing unfed gnathiid survival relative to time (days), Figure S5: Great Barrier Reef; Scaled Shoenfeld residual plot for full model testing fed gnathiid survival relative to time (days), Figure S6: Philippines; Scaled Shoenfeld residual plot for full model testing gnathiid survival relative to time (days) for Trial 1, Figure S7: Philippines; Scaled Shoenfeld residual plot for full model testing gnathiid survival relative to time (days), for Trial 2.
Author Contributions
Conceptualization, P.C.S. and A.S.G.; methodology, P.C.S., A.S.G., M.O.S. and B.D.; validation, A.S.G. and P.C.S.; formal analysis, A.S.G., M.O.S. and A.G.-L.; investigation, M.O.S. and B.D.; resources, P.C.S., A.S.G., M.O.S. and A.G.-L.; data curation, M.O.S. and B.D.; writing—original draft preparation, M.O.S., B.D., P.C.S. and A.S.G.; writing—review and editing, M.O.S., A.G.-L. and P.C.S.; visualization, A.S.G. and M.O.S.; supervision, P.C.S. and A.S.G.; project administration, A.S.G., P.C.S. and M.O.S.; funding acquisition, A.S.G., A.G.-L. and P.C.S. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Australian Research Council (A00105175, A19937078, ARCFEL010G, DP0557058, DP120102415), Sea World Research and Rescue Foundation Australia (SWR/2/2012), and the US National Science Foundation (OCE-1536794, PC Sikkel, PI).
Acknowledgments
We thank the many volunteers and the Lizard Island Research Station (GBR) staff who helped maintain the gnathiid culture and provided equipment and facilities. We also thank Jessica Vorse, who conducted an earlier pilot study on the effects of temperature on gnathiid survival on the Great Barrier Reef; this was invaluable in the development of the final methodology implemented in the present study. We thank the municipality of Sibulan, and Dumaguete City, Negros Oriental, Philippines, for permission to conduct this study (0154-18 DA-BFAR). We also thank Hilconida P. Calumpong, Janet S. Estacion, Rene A. Abesamis, and the staff of the Silliman University Institute for Environmental and Marine Sciences for logistic support, equipment and use of facilities. We thank Jeremiah Gepaya and Lucille Jean Raterta for their field assistance and Dioscoro Inocencio for fish collections and field support. The Authors are also grateful to the three anonymous reviewers for their constructive comments.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bindoff, N.L.; Cheung, W.W.L.; Kairo, J.G.; Arístegui, J.; Guinder, V.A.; Hilmi, N.; Jiao, N.; Karim, M.S.; Levin, L.; O’Donoghue, S.; et al. Changing Ocean, Marine Ecosystems, and Dependent Communities; ETH Zurich: Zurich, Switzerland, 2019; pp. 477–587. [Google Scholar]
- Huey, R.B.; Stevenson, R.D. Integrating Thermal Physiology and Ecology of Ectotherms: A Discussion of Approaches. Am. Zool. 1979, 19, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Brett, J.R. 3. Temperature, 3.3. Animals, 3.32. Fishes-Functional Reponses. In Marine Ecology: A Comprehensive, Integrated Treatise on Life in Oceans and Coastal Waters; Kinne, O., Ed.; Wiley-Interscience: Hoboken, NJ, USA, 1984; Volume 1, pp. 515–560. [Google Scholar]
- Kinne, O. 3. Temperature, 3.3. Animals, 3.31. Invertebrates. In Marine Ecology: A Comprehensive, Integrated Treatise on Life in Oceans and Coastal Waters; Kinne, O., Ed.; Wiley-Interscience: Hoboken, NJ, USA, 1984; Volume 1, pp. 407–514. [Google Scholar]
- Newell, R.C.; Branch, G.M. The influence of temperature on the maintenance of metabolic energy balance in marine invertebrates. Adv. Mar. Biol. 1980, 17, 329–396. [Google Scholar] [CrossRef]
- Houde, E.D. Comparative growth, mortality, and energetics of marine fish larvae: Temperature and implied latitudinal effects. Fish. Bull. 1989, 87, 471–495. [Google Scholar]
- Schmidt-Nielsen, K. Animal Physiology: Adaptation and Environment; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Marsden, I.D. Effect of temperature on the microdistribution of the isopod Sphaeroma rugicauda from a saltmarsh habitat. Mar. Biol. 1976, 38, 117–128. [Google Scholar] [CrossRef]
- McNamara, J.C.; Moreira, P.S.; Moreira, G.S. The effect of salinity on the upper thermal limits of survival and metamorphosis during larval development in Macrobrachium amazonicum (Heller) (Decapoda, Palaemonidae). Crustaceana 1986, 50, 231–238. [Google Scholar] [CrossRef]
- Marcogliese, D.J. The impact of climate change on the parasites and infectious diseases of aquatic animals. Oie Rev. Sci. Tech. 2008, 27, 467–484. [Google Scholar] [CrossRef]
- Marcogliese, D.J. The Distribution and Abundance of Parasites in Aquatic Ecosystems in a Changing Climate: More than Just Temperature. Integr. Comp. Biol. 2016, 56, 611–619. [Google Scholar] [CrossRef] [Green Version]
- Munday, P.L.; Jones, G.P.; Pratchett, M.S.; Williams, A.J. Climate change and the future for coral reef fishes. Fish Fish. 2008, 9, 261–285. [Google Scholar] [CrossRef]
- Przeslawski, R.; Ahyong, S.; Byrne, M.; Wörheide, G.; Hutchings, P. Beyond corals and fish: The effects of climate change on noncoral benthic invertebrates of tropical reefs. Glob. Chang. Biol. 2008, 14, 2773–2795. [Google Scholar] [CrossRef]
- Donelson, J.M.; Munday, P.L.; McCormick, M.I.; Pankhurst, N.W.; Pankhurst, P.M. Effects of elevated water temperature and food availability on the reproductive performance of a coral reef fish. Mar. Ecol. Prog. Ser. 2010, 401, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Zarco-Perello, S.; Pratchett, M.; Liao, V. Temperature-growth performance curves for a coral reef fish, Acanthochromis polyacanthus. Galaxea J. Coral Reef Stud. 2012, 14, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.-C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef] [PubMed]
- Animals and Temperature: Phenotypic and Evolutionary Adaptation; Johnston, I.A.; Bennett, A.F. (Eds.) Cambridge University Press: Cambridge, UK, 2008. [Google Scholar]
- Hoey, A.S.; Howells, E.; Johansen, J.L.; Hobbs, J.-P.A.; Messmer, V.; McCowan, D.M.; Wilson, S.K.; Pratchett, M.S. Recent Advances in Understanding the Effects of Climate Change on Coral Reefs. Diversity 2016, 8, 12. [Google Scholar] [CrossRef] [Green Version]
- Fields, P.A.; Graham, J.B.; Rosenblatt, R.H.; Somero, G.N. Effects of expected global climate change on marine faunas. Trends Ecol. Evol. 1993, 8, 361–367. [Google Scholar] [CrossRef]
- Jennings, S.; Brander, K. Predicting the effects of climate change on marine communities and the consequences for fisheries. J. Mar. Syst. 2010, 79, 418–426. [Google Scholar] [CrossRef]
- Kordas, R.L.; Harley, C.D.G.; O’Connor, M.I. Community ecology in a warming world: The influence of temperature on interspecific interactions in marine systems. J. Exp. Mar. Biol. Ecol. 2011, 400, 218–226. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O. Climate change, coral bleaching and the future of the world’s coral reefs. Mar. Freshw. Res. 1999. [Google Scholar] [CrossRef] [Green Version]
- Bellwood, D.R.; Hughes, T.P. Regional-scale assembly rules and biodiversity of coral reefs. Science 2001, 292, 1532–1534. [Google Scholar] [CrossRef] [Green Version]
- Lough, J.M. Small change, big difference: Sea surface temperature distributions for tropical coral reef ecosystems, 1950-2011. J. Geophys. Res. Ocean. 2012, 117. [Google Scholar] [CrossRef] [Green Version]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Dietzel, A.; Eakin, C.M.; Heron, S.F.; Hoey, A.S.; Hoogenboom, M.O.; Liu, G.; et al. Global warming transforms coral reef assemblages. Nature 2018, 556, 492–496. [Google Scholar] [CrossRef]
- Lough, J.M.; Anderson, K.D.; Hughes, T.P. Increasing thermal stress for tropical coral reefs: 1871-2017. Sci. Rep. 2018, 8, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tewksbury, J.J.; Huey, R.B.; Deutsch, C.A. ECOLOGY: Putting the Heat on Tropical Animals. Science 2008, 320, 1296–1297. [Google Scholar] [CrossRef]
- Sunday, J.M.; Bates, A.E.; Dulvy, N.K. Global analysis of thermal tolerance and latitude in ectotherms. Proc. R. Soc. B: Biol. Sci. 2011, 278, 1823–1830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habary, A.; Johansen, J.L.; Nay, T.J.; Steffensen, J.F.; Rummer, J.L. Adapt, move or die - how will tropical coral reef fishes cope with ocean warming? Glob. Chang. Biol. 2017, 23, 566–577. [Google Scholar] [CrossRef]
- Lough, J.M. 10th Anniversary Review: A changing climate for coral reefs. J. Environ. Monit. 2008, 10, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Hughes, T.P.; Kerry, J.T.; Álvarez-Noriega, M.; Álvarez-Romero, J.G.; Anderson, K.D.; Baird, A.H.; Babcock, R.C.; Beger, M.; Bellwood, D.R.; Berkelmans, R.; et al. Global warming and recurrent mass bleaching of corals. Nature 2017, 543, 373–377. [Google Scholar] [CrossRef]
- Hughes, T.P.; Anderson, K.D.; Connolly, S.R.; Heron, S.F.; Kerry, J.T.; Lough, J.M.; Baird, A.H.; Baum, J.K.; Berumen, M.L.; Bridge, T.C.; et al. Spatial and temporal patterns of mass bleaching of corals in the Anthropocene. Science 2018, 359, 80–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliver, J.K.; Berkelmans, R.; Eakin, C.M. Coral Bleaching in Space and Time. In Coral Bleaching: Patterns, Processes, Causes and Consequences; van Oppen, M.J.H., Lough, J.M., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 27–49. [Google Scholar] [CrossRef]
- Skirving, W.J.; Heron, S.F.; Marsh, B.L.; Liu, G.; De La Cour, J.L.; Geiger, E.F.; Eakin, C.M. The relentless march of mass coral bleaching: A global perspective of changing heat stress. Coral Reefs 2019, 38, 547–557. [Google Scholar] [CrossRef] [Green Version]
- Hughes, T.P.; Kerry, J.T.; Baird, A.H.; Connolly, S.R.; Chase, T.J.; Dietzel, A.; Hill, T.; Hoey, A.S.; Hoogenboom, M.O.; Jacobson, M.; et al. Global warming impairs stock–recruitment dynamics of corals. Nature 2019, 568, 387–390. [Google Scholar] [CrossRef]
- Raymundo, L.J.; Burdick, D.; Hoot, W.C.; Miller, R.M.; Brown, V.; Reynolds, T.; Gault, J.; Idechong, J.; Fifer, J.; Williams, A. Successive bleaching events cause mass coral mortality in Guam, Micronesia. Coral Reefs 2019, 38, 677–700. [Google Scholar] [CrossRef]
- Mora, C.; Ospína, A.F. Tolerance to high temperatures and potential impact of sea warming on reef fishes of Gorgona Island (tropical eastern Pacific). Mar. Biol. 2001, 139, 765–769. [Google Scholar] [CrossRef]
- Booth, D.; Beretta, G. Changes in a fish assemblage after a coral bleaching event. Mar. Ecol. Prog. Ser. 2002, 245, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Spalding, M.D.; Jarvis, G.E. The impact of the 1998 coral mortality on reef fish communities in the Seychelles. Mar. Pollut. Bull. 2002, 44, 309–321. [Google Scholar] [CrossRef]
- Jones, G.P.; McCormick, M.I.; Srinivasan, M.; Eagle, J.V. Coral decline threatens fish biodiversity in marine reserves. Proc. Natl. Acad. Sci. USA 2004, 101, 8251–8253. [Google Scholar] [CrossRef] [Green Version]
- Ospina, A.F.; Mora, C. Effect of body size on reef fish tolerance to extreme low and high temperatures. Environ. Biol. Fishes 2004, 70, 339–343. [Google Scholar] [CrossRef]
- Roessig, J.M.; Woodley, C.M.; Cech, J.J.; Hansen, L.J. Effects of global climate change on marine and estuarine fishes and fisheries. Rev. Fish Biol. Fish. 2004, 14, 251–275. [Google Scholar] [CrossRef]
- Graham, N.A.J.; Wilson, S.K.; Jennings, S.; Polunin, N.V.C.; Bijoux, J.P.; Robinson, J. Dynamic fragility of oceanic coral reef ecosystems. Proc. Natl. Acad. Sci. USA 2006, 103, 8425–8429. [Google Scholar] [CrossRef] [Green Version]
- Garpe, K.C.; Yahya, S.A.S.; Lindahl, U.; Öhman, M.C. Long-term effects of the 1998 coral bleaching event on reef fish assemblages. Mar. Ecol. Prog. Ser. 2006, 315, 237–247. [Google Scholar] [CrossRef]
- Wilson, S.K.; Graham, N.A.J.; Pratchett, M.S.; Jones, G.P.; Polunin, N.V.C. Multiple disturbances and the global degradation of coral reefs: Are reef fishes at risk or resilient? Glob. Chang. Biol. 2006, 12, 2220–2234. [Google Scholar] [CrossRef]
- Pratchett, M.S.; Munday, P.; Wilson, S.K.; Graham, N.A.J.; Cinner, J.; Bellwood, D.R.; Jones, G.P.; Polunin, N.V.C.; McClanahan, T.R. Effects Of Climate-Induced Coral Bleaching On Coral-Reef Fishes‚ Ecological And Economic Consequences. In Oceanography and Marine Biology: An Annual Reviewiology; Gibson, R.N., Atkinson, R.J.A., Gordon, J.D.M., Eds.; Taylor & Francis: Milton Park, UK, 2008; pp. 257–302. [Google Scholar]
- Clark, T.D.; Roche, D.G.; Binning, S.A.; Speers-Roesch, B.; Sundin, J. Maximum thermal limits of coral reef damselfishes are size dependent and resilient to near-future ocean acidification. J. Exp. Biol. 2017, 220, 3519–3526. [Google Scholar] [CrossRef] [Green Version]
- Triki, Z.; Wismer, S.; Levorato, E.; Bshary, R. A decrease in the abundance and strategic sophistication of cleaner fish after environmental perturbations. Glob. Chang. Biol. 2018, 24, 481–489. [Google Scholar] [CrossRef] [PubMed]
- Plaisance, L.; Knowlton, N.; Paulay, G.; Meyer, C. Reef-associated crustacean fauna: Biodiversity estimates using semi-quantitative sampling and DNA barcoding. Coral Reefs 2009, 28, 977–986. [Google Scholar] [CrossRef] [Green Version]
- Plaisance, L.; Caley, M.J.; Brainard, R.E.; Knowlton, N. The diversity of coral reefs: What are we missing? PLoS ONE 2011, 6, e25026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brandl, S.J.; Tornabene, L.; Goatley, C.H.R.; Casey, J.M.; Morais, R.A.; Côté, I.M.; Baldwin, C.C.; Parravicini, V.; Schiettekatte, N.M.D.; Bellwood, D.R. Demographic dynamics of the smallest marine vertebrates fuel coral reef ecosystem functioning. Science 2019, 364, 1189–1192. [Google Scholar] [CrossRef] [PubMed]
- De Meeûs, T.; Renaud, F. Parasites within the new phylogeny of eukaryotes. Trends Parasitol. 2002, 18, 247–250. [Google Scholar] [CrossRef]
- Rohde, K. Ecology of Marine Parasites; University of Queensland Press: St Lucia, Australia, 1982; p. 245. [Google Scholar]
- Dobson, A.; Lafferty, K.D.; Kuris, A.M.; Hechinger, R.F.; Jetz, W. Homage to linnaeus: How many parasites? How many hosts? Light Evol. 2009, 2, 63–82. [Google Scholar] [CrossRef] [Green Version]
- Hatcher, M.J.; Dunn, A.M. Parasites in Ecological Communities: From Interactions to Ecosystems; Cambridge University Press: Cambridge, UK, 2011. [Google Scholar] [CrossRef] [Green Version]
- Okamura, B.; Hartigan, A.; Naldoni, J. Extensive uncharted biodiversity: The parasite dimension. Integr. Comp. Biol. 2018, 56, 1132–1145. [Google Scholar] [CrossRef]
- Hudson, P.J.; Dobson, A.P.; Lafferty, K.D. Is a healthy ecosystem one that is rich in parasites? Trends Ecol. Evol. 2006, 21, 381–385. [Google Scholar] [CrossRef]
- Kuris, A.M.; Hechinger, R.F.; Shaw, J.C.; Whitney, K.L.; Aguirre-Macedo, L.; Boch, C.A.; Dobson, A.P.; Dunham, E.J.; Fredensborg, B.L.; Huspeni, T.C.; et al. Ecosystem energetic implications of parasite and free-living biomass in three estuaries. Nature 2008, 454, 515–518. [Google Scholar] [CrossRef]
- Hatcher, M.J.; Dick, J.T.A.; Dunn, A.M. Diverse effects of parasites in ecosystems: Linking interdependent processes. Front. Ecol. Environ. 2012, 10, 186–194. [Google Scholar] [CrossRef]
- Hatcher, M.J.; Dick, J.T.A.; Dunn, A.M. Parasites that change predator or prey behaviour can have keystone effects on community composition. Biol. Lett. 2014, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohde, K. Ecology and biogeography of marine parasites. Adv. Mar. Biol. 2002, 43, 1–86. [Google Scholar] [PubMed]
- Rohde, K. Species diversity of parasites on the Great Barrier Reef. Z. Für Parasitenkd. 1976, 50, 93–94. [Google Scholar] [CrossRef]
- Poulin, R.; Blasco-Costa, I.; Randhawa, H.S. Integrating parasitology and marine ecology: Seven challenges towards greater synergy. J. Sea Res. 2016, 113, 3–10. [Google Scholar] [CrossRef]
- Marcogliese, D.J. Implications of climate change for parasitism of animals in the aquatic environment. Can. J. Zool. 2001, 79, 1331–1352. [Google Scholar] [CrossRef]
- Sikkel, P.C.; Richardson, M.A.; Sun, D.; Narvaez, P.; Feeney, W.E.; Grutter, A.S. Changes in abundance of fish-parasitic gnathiid isopods associated with warm-water bleaching events on the northern Great Barrier Reef. Coral Reefs 2019, 38, 721–730. [Google Scholar] [CrossRef]
- Sikkel, P.C.; Welicky, R.L. The Ecological Significance of Parasitic Crustaceans. In Parasitic Crustacea; Springer: London, UK, 2019; pp. 421–477. [Google Scholar]
- Claar, D.C.; Wood, C.L. Pulse Heat Stress and Parasitism in a Warming World. Trends Ecol. Evol. 2020, 35, 704–715. [Google Scholar] [CrossRef]
- Gehman, A.-L.M.; Hall, R.J.; Byers, J.E. Host and parasite thermal ecology jointly determine the effect of climate warming on epidemic dynamics. Proc. Natl. Acad. Sci. USA 2018, 115, 744–749. [Google Scholar] [CrossRef] [Green Version]
- Brazenor, A.K.; Hutson, K.S. Effects of temperature and salinity on the life cycle of Neobenedenia sp. (Monogenea: Capsalidae) infecting farmed barramundi (Lates calcarifer). Parasitol. Res. 2015, 114, 1875–1886. [Google Scholar] [CrossRef]
- Grutter, A.S. Spatial and temporal variations of the ectoparasites of seven reef fish species from Lizard Island and Heron Island, Australia. Mar. Ecol. Prog. Ser. 1994, 115, 21–30. [Google Scholar] [CrossRef]
- Grutter, A.S. Parasite removal rates by the cleaner wrasse Labroides dimidiatus. Mar. Ecol. Prog. Ser. 1996, 130, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Grutter, A.S.; Poulin, R. Cleaning of Coral Reef Fishes by the Wrasse Labroides dimidiatus: Influence of Client Body Size and Phylogeny. Copeia 1998, 1998, 1447707. [Google Scholar] [CrossRef]
- Smit, N.J.; Davies, A.J. The curious life-style of the parasitic stages of Gnathiid isopods. Adv. Parasitol. 2004, 58, 289–391. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K. Life history of gnathiid isopods—Current knowledge and and future directions. Plankton Benthos Res. 2007, 2, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Lafferty, K.D.; Kuris, A.M. Trophic strategies, animal diversity and body size. Trends Ecol. Evol. 2002, 17, 507–513. [Google Scholar] [CrossRef]
- Raffel, T.R.; Martin, L.B.; Rohr, J.R. Parasites as predators: Unifying natural enemy ecology. Trends Ecol. Evol. 2008, 23, 610–618. [Google Scholar] [CrossRef]
- Grutter, A.S. Parasite infection rather than tactile stimulation is the proximate cause of cleaning behaviour in reef fish. Proc. R. Soc. B Biol. Sci. 2001, 268, 1361–1365. [Google Scholar] [CrossRef] [Green Version]
- Grutter, A.S. Feeding ecology of the fish ectoparasite Gnathia sp. (Crustacea: Isopoda) from the Great Barrier Reef, and its implications for fish cleaning behaviour. Mar. Ecol. Prog. Ser. 2003, 259, 295–302. [Google Scholar] [CrossRef]
- Sikkel, P.C.; Cheney, K.L.; Côté, I.M. In situ evidence for ectoparasites as a proximate cause of cleaning interactions in reef fish. Anim. Behav. 2004, 68, 241–247. [Google Scholar] [CrossRef]
- Sikkel, P.C.; Herzlieb, S.E.; Kramer, D.L. Compensatory cleaner-seeking behavior following spawning in female yellowtail damselfish. Mar. Ecol. Prog. Ser. 2005, 296, 1–11. [Google Scholar] [CrossRef]
- Cheney, K.L.; Côté, I.M. Mutualism or parasitism? The variable outcome of cleaning symbioses. Biol. Lett. 2005, 1, 162–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Binning, S.A.; Roche, D.G.; Grutter, A.S.; Colosio, S.; Sun, D.; Miest, J.; Bshary, R. Cleaner wrasse indirectly affect the cognitive performance of a damselfish through ectoparasite removal. Proc. R. Soc. B Biol. Sci. 2018, 285. [Google Scholar] [CrossRef] [PubMed]
- Sellers, J.C.; Holstein, D.M.; Botha, T.L.; Sikkel, P.C. Lethal and sublethal impacts of a micropredator on post-settlement Caribbean reef fishes. Oecologia 2019, 189, 293–305. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.M.; Grutter, A.S. Parasitic isopods (Gnathia sp.) reduce haematocrit in captive blackeye thicklip (Labridae) on the Great Barrier Reef. J. Fish Biol. 2005, 66, 860–864. [Google Scholar] [CrossRef]
- Hayes, P.M.; Smit, N.J.; Grutter, A.S.; Davies, A.J. Unexpected response of a captive blackeye thicklip, Hemigymnus melapterus (Bloch), from Lizard Island, Australia, exposed to juvenile isopods Gnathia aureamaculosa Ferreira & Smit. J. Fish Dis. 2011, 34, 563–566. [Google Scholar] [CrossRef]
- Grutter, A.S.; Pickering, J.L.; McCallum, H.; McCormick, M.I. Impact of micropredatory gnathiid isopods on young coral reef fishes. Coral Reefs 2008, 27, 655–661. [Google Scholar] [CrossRef]
- Jones, C.M.; Grutter, A.S. Reef-based micropredators reduce the growth of post-settlement damselfish in captivity. Coral Reefs 2008, 27, 677–684. [Google Scholar] [CrossRef]
- Penfold, R.; Grutter, A.S.; Kuris, A.M.; McCormick, M.I.; Jones, C.M. Interactions between juvenile marine fish and gnathiid isopods: Predation versus micropredation. Mar. Ecol. Prog. Ser. 2008, 357, 111–119. [Google Scholar] [CrossRef]
- Artim, J.M.; Sellers, J.C.; Sikkel, P.C. Micropredation by gnathiid isopods on settlement stage reef fish in the eastern Caribbean Sea. Bull. Mar. Sci. 2015, 91, 479–487. [Google Scholar] [CrossRef]
- Grutter, A.S.; Blomberg, S.P.; Fargher, B.; Kuris, A.M.; McCormick, M.I.; Warner, R.R. Size-related mortality due to gnathiid isopod micropredation correlates with settlement size in coral reef fishes. Coral Reefs 2017, 36, 549–559. [Google Scholar] [CrossRef]
- Curtis, L.M.; Grutter, A.S.; Smit, N.J.; Davies, A.J. Gnathia aureamaculosa, a likely definitive host of Haemogregarina balistapi and potential vector for Haemogregarina bigemina between fishes of the Great Barrier Reef, Australia. Int. J. Parasitol. 2013, 43, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Honma, Y.; Chiba, A. Pathological changes in the branchial chamber wall of stingrays, Dasyatis spp., associated with the presence of juvenile gnathiids (Isopoda, Crustacea). Fish Pathol. 1991, 26, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Grutter, A.S. Spatiotemporal Variation and Feeding Selectivity in the Diet of the Cleaner Fish Labroides dimidiatus. Copeia 1997, 1997, 1447754. [Google Scholar] [CrossRef]
- Grutter, A.S. Cleaning symbioses from the parasites’ perspective. Parasitology 2002, 124, 65–81. [Google Scholar] [CrossRef]
- Grutter, A.S.; Blomberg, S.P.; Box, S.; Bshary, R.; Ho, O.; Madin, E.M.P.; McClure, E.C.; Meekan, M.G.; Murphy, J.M.; Richardson, M.A.; et al. Changes in local free-living parasite populations in response to cleaner manipulation over 12 years. Oecologia 2019, 190, 783–797. [Google Scholar] [CrossRef]
- Demairé, C.; Triki, Z.; Binning, S.A.; Glauser, G.; Roche, D.G.; Bshary, R. Reduced access to cleaner fish negatively impacts the physiological state of two resident reef fishes. Mar. Biol. 2020, 167, 48. [Google Scholar] [CrossRef]
- Grutter, A.; Bejarano, S.; Cheney, K.; Goldizen, A.; Sinclair-Taylor, T.; Waldie, P. Effects of the cleaner fish Labroides dimidiatus on grazing fishes and coral reef benthos. Mar. Ecol. Prog. Ser. 2020, 643, 99–114. [Google Scholar] [CrossRef]
- Hughes, T.P.; Kerry, J.T.; Simpson, T. Large-scale bleaching of corals on the Great Barrier Reef. Ecology 2018, 99, 501. [Google Scholar] [CrossRef] [Green Version]
- Arceo, H.O.; Quibilan, M.C.; Aliño, P.M.; Lim, G.; Licuanan, W.Y. Coral bleaching in Philippine reefs: Coincident evidences with mesoscale thermal anomalies. Bull. Mar. Sci. 2001, 69, 579–593. [Google Scholar]
- Raymundo, L.; Maypa, A. Recovery of the Apo Island Marine Reserve, Philippines, 2 years after the El Niño bleaching event. Coral Reefs 2002, 21, 260–261. [Google Scholar] [CrossRef]
- Magdaong, E.T.; Fujii, M.; Yamano, H.; Licuanan, W.Y.; Maypa, A.; Campos, W.L.; Alcala, A.C.; White, A.T.; Apistar, D.; Martinez, R. Long-term change in coral cover and the effectiveness of marine protected areas in the Philippines: A meta-analysis. Hydrobiologia 2014, 733, 5–17. [Google Scholar] [CrossRef] [Green Version]
- Grutter, A.S.; Feeney, W.E.; McClure, E.C.; Narvaez, P.; Smit, N.J.; Sun, D.; Sikkel, P.C. Practical methods for culturing parasitic gnathiid isopods. Int. J. Parasitol. 2020, 50, 825–837. [Google Scholar] [CrossRef] [PubMed]
- Sikkel, P.C.; Tuttle, L.J.; Cure, K.; Coile, A.M.; Hixon, M.A. Low Susceptibility of Invasive Red Lionfish (Pterois volitans) to a Generalist Ectoparasite in Both Its Introduced and Native Ranges. PLoS ONE 2014, 9, e95854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santos, T.R.N.; Sikkel, P.C. Habitat associations of fish-parasitic gnathiid isopods in a shallow reef system in the central Philippines. Mar. Biodivers. 2017, 49, 83–96. [Google Scholar] [CrossRef]
- Artim, J.M.; Sikkel, P.C. Comparison of sampling methodologies and estimation of population parameters for a temporary fish ectoparasite. Int. J. Parasitol. Parasites Wildl. 2016, 5, 145–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graba-Landry, A.C.; Loffler, Z.; McClure, E.C.; Pratchett, M.S.; Hoey, A.S. Impaired growth and survival of tropical macroalgae (Sargassum spp.) at elevated temperatures. Coral Reefs 2020, 39, 475–486. [Google Scholar] [CrossRef]
- Pacific Ocean: Great Barrier Reef: Sea Water Temperature Compared to Long Term Averages. Available online: http://data.aims.gov.au/aimsrtds/yearlytrends.xhtml (accessed on 13 November 2018).
- Water Temperature at Lizard Island Graph. Australian Institute of Marine Science AIMS. Available online: http://data.aims.gov.au/aimsrtds/datatool.xhtml?from=2018-10-15&thru=2018-10-20&period=MONTH&aggregations=AVG&channels=3523 (accessed on 13 November 2018).
- NOAA Coral Reef Watch Program. Available online: https://coralreefwatch.noaa.gov/satellite/vs/philippines.php (accessed on 5 January 2018).
- Therneau, T.M.; Grambsch, P.M. Modeling Survival Data: Extending the Cox Model; Springer: New York, NY, USA, 2000. [Google Scholar]
- Therneau, T. Coxme: Mixed Effects Cox Models. R Package Version 2.2-3. 2015. Available online: http://CRAN.R-project.org/package=coxme (accessed on 17 July 2020).
- Fox, J.; Weisberg, S. Multivariate Linear Models in R. In An R Companion to Applied Regression, 2nd ed.; Thousand Oaks: Los Angeles, CA, USA, 2011; pp. 1–31. [Google Scholar]
- Ganachaud, A.; Sen Gupta, A.S.; Orr, J.C.; Wijffels, S.E.; Ridgway, K.R.; Hemer, M.A.; Maes, C.; Steinberg, C.R.; Tribollet, A.D.; Qiu, B.; et al. Observed and expected changes to the tropical Pacific Ocean. In Vulnerability of Tropical Pacific Fisheries and Aquaculture to Climate Change; Bell, J.D., Johnson, J.E., Hobday, A.J., Eds.; Secretariat of the Pacific Community: Noumea, New Caledonia, 2011; pp. 101–187. [Google Scholar]
- Huey, R.B.; Kingsolver, J.G. Climate Warming, Resource Availability, and the Metabolic Meltdown of Ectotherms. Am. Nat. 2019, 194, E140–E150. [Google Scholar] [CrossRef]
- Smale, D.A.; Wernberg, T.; Oliver, E.C.J.; Thomsen, M.; Harvey, B.P.; Straub, S.C.; Burrows, M.T.; Alexander, L.V.; Benthuysen, J.A.; Donat, M.G.; et al. Marine heatwaves threaten global biodiversity and the provision of ecosystem services. Nat. Clim. Chang. 2019, 9, 306–312. [Google Scholar] [CrossRef] [Green Version]
- Frölicher, T.L.; Fischer, E.M.; Gruber, N. Marine heatwaves under global warming. Nature 2018, 560, 360–364. [Google Scholar] [CrossRef]
- Oliver, E.C.J.; Donat, M.G.; Burrows, M.T.; Moore, P.J.; Smale, D.A.; Alexander, L.V.; Benthuysen, J.A.; Feng, M.; Gupta, A.S.; Hobday, A.J.; et al. Longer and more frequent marine heatwaves over the past century. Nat. Commun. 2018, 9, 1324. [Google Scholar] [CrossRef]
- Compton, T.J.; Rijkenberg, M.J.A.; Drent, J.; Piersma, T. Thermal tolerance ranges and climate variability: A comparison between bivalves from differing climates. J. Exp. Mar. Biol. Ecol. 2007, 352, 200–211. [Google Scholar] [CrossRef] [Green Version]
- Stillman, J.H.; Somero, G.N. A comparative analysis of the upper thermal tolerance limits of eastern pacific porcelain crabs, genus Petrolisthes: Influences of latitude, vertical zonation, acclimation, and phylogeny. Physiol. Biochem. Zool. 2000, 73, 200–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attrill, M.J.; Kelmo, F.; Jones, M.B. Impact of the 1997-98 El Niño event on the coral reef-associated echinoderm assemblage from northern Bahia, northeastern Brazil. Clim. Res. 2004, 26, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Chan, B.K.K.; Morritt, D.; De Pirro, M.; Leung, K.M.Y.; Williams, G.A. Summer mortality: Effects on the distribution and abundance of the acorn barnacle Tetraclita japonica on tropical shores. Mar. Ecol. Prog. Ser. 2006, 328, 195–204. [Google Scholar] [CrossRef]
- Jokiel, P.L.; Coles, S.L. Coral Reefs to elevated temperature. Environ. Conserv. 1990, 8, 155–162. [Google Scholar]
- Mouritsen, K.N.; Poulin, R. Parasitism, climate oscillations and the structure of natural communities. Oikos 2002, 97, 462–468. [Google Scholar] [CrossRef] [Green Version]
- Poulin, R. Global warming and temperature-mediated increases in cercarial emergence in trematode parasites. Parasitology 2006, 132, 143–151. [Google Scholar] [CrossRef]
- Thieltges, D.W.; Fredensborg, B.L.; Studer, A.; Poulin, R. Large-scale patterns in trematode richness and infection levels in marine crustacean hosts. Mar. Ecol. Prog. Ser. 2009, 389, 139–147. [Google Scholar] [CrossRef]
- Pechenik, J.A.; Fried, B. Effect of temperature on survival and infectivity of Echinostoma trivolvis cercariae: A test of the energy limitation hypothesis. Parasitology 1995, 111, 373–378. [Google Scholar] [CrossRef]
- Lyholt, H.C.K.; Buchmann, K. Diplostomum spathaceum:effects of temperature and light on cercarial shedding and infection of rainbow trout. Dis. Aquat. Org. 1996, 25, 169–173. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, A.M. The influence of temperature on the survival and infectivity of the cercariae of Echinoparyphium recurvatum (Digenea: Echinostomatidae). Parasitology 1999, 118, 383–388. [Google Scholar] [CrossRef] [PubMed]
- Toledo, R.; Muñoz-Antoli, C.; Pérez, M.; Esteban, J.G. Survival and infectivity of Hypoderaeum conoideum and Euparyphium albuferensis cercariae under laboratory conditions. J. Helminthol. 1999, 73, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Mouritsen, K.N. The Hydrobia ulvae—Maritrema subdolum association: Influence of temperature, salinity, light, water-pressure and secondary host exudates on cercarial emergence and longevity. J. Helminthol. 2002, 76, 341–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thieltges, D.W.; Rick, J. Effect of temperature on emergence, survival and infectivity of cercariae of the marine trematode Renicola roscovita (Digenea: Renicolidae). Dis. Aquat. Org. 2006, 73, 63–68. [Google Scholar] [CrossRef]
- Leiva, N.V.; Manríquez, P.H.; Aguilera, V.M.; González, M.T. Temperature and pCO2 jointly affect the emergence and survival of cercariae from a snail host: Implications for future parasitic infections in the Humboldt Current system. Int. J. Parasitol. 2019, 49, 49–61. [Google Scholar] [CrossRef]
- Franzova, V.A.; MacLeod, C.D.; Wang, T.; Harley, C.D.G. Complex and interactive effects of ocean acidification and warming on the life span of a marine trematode parasite. Int. J. Parasitol. 2019, 49, 1015–1021. [Google Scholar] [CrossRef]
- Conley, D.C.; Curtis, M.A. Effects of temperature and photoperiod on the duration of hatching, swimming, and copepodid survival of the parasitic copepod Salmincola edwardsii. Can. J. Zool. 1993, 71, 972–976. [Google Scholar] [CrossRef]
- Hirazawa, N.; Takano, R.; Hagiwara, H.; Noguchi, M.; Narita, M. The influence of different water temperatures on Neobenedenia girellae (Monogenea) infection, parasite growth, egg production and emerging second generation on amberjack Seriola dumerili (Carangidae) and the histopathological effect of this parasite on fish skin. Aquaculture 2010, 299, 2–7. [Google Scholar] [CrossRef]
- Mahmoud, N.E.; Fahmy, M.M.; Abuowarda, M.M.; Zaki, M.; Ismail, E.M.; Ismael, E.S. Mediterranean sea fry; a source of isopod infestation problem in Egypt with reference to the effect of salinity and temperature on the survival of Livoneca redmanii (Isopoda: Cymothoidae) juvenile stages. J. Egypt. Soc. Parasitol. 2019, 49, 235–242. [Google Scholar]
- Evans, N.A. The influence of environmental temperature upon transmission of the cercariae of Echinostoma liei (Digenea: Echinostomatidae). Parasitology 1985, 90, 269–275. [Google Scholar] [CrossRef]
- Lo, C.T.; Lee, K.M. Pattern of Emergence and the Effects of Temperature and Light on the Emergence and Survival of Heterophyid Cercariae (Centrocestus formosanus and Haplorchis pumilio). J. Parasitol. 1996, 82, 347. [Google Scholar] [CrossRef] [PubMed]
- Fried, B.; Ponder, E.L. Effects of temperature on survival, infectivity and in vitro encystment of the cercariae of Echinostoma caproni. J. Helminthol. 2003, 77, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Tucker, C.S.; Sommerville, C.; Wootten, R. The effect of temperature and salinity on the settlement and survival of copepodids of Lepeophtheirus salmonis (Krøyer, 1837) on Atlantic salmon, Salmo salar L. J. Fish Dis. 2000, 23, 309–320. [Google Scholar] [CrossRef]
- Peck, L.S.; Webb, K.E.; Bailey, D.M. Extreme sensitivity of biological function to temperature in Antarctic marine species. Funct. Ecol. 2004, 18, 625–630. [Google Scholar] [CrossRef] [Green Version]
- Pörtner, H.O.; Knust, R. Climate change affects marine fishes through the oxygen limitation of thermal tolerance. Science 2007, 315, 95–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meyrowitsch, D.; Christensen, N.; Hindsbo, O. Effects of temperature and host density on the snail-finding capacity of cercariae of Echinostoma caproni (Digenea: Echinostomatidae). Parasitology 1991, 102, 391–395. [Google Scholar] [CrossRef]
- Cech, J.J.; Moyle, P.B. Fishes: An Introduction to Ichthyology, 5th ed.; Prentice Hall: Upper Saddle River, NJ, USA, 2004. [Google Scholar]
- Gagliano, M.; McCormick, M.I.; Meekan, M.G. Temperature-induced shifts in selective pressure at a critical developmental transition. Oecologia 2007, 152, 219–225. [Google Scholar] [CrossRef]
- Nilsson, G.E.; Crawley, N.; Lunde, I.G.; Munday, P.L. Elevated temperature reduces the respiratory scope of coral reef fishes. Glob. Chang. Biol. 2009, 15, 1405–1412. [Google Scholar] [CrossRef]
- Nay, T.J.; Johansen, J.L.; Habary, A.; Steffensen, J.F.; Rummer, J.L. Behavioural thermoregulation in a temperature-sensitive coral reef fish, the five-lined cardinalfish (Cheilodipterus quinquelineatus). Coral Reefs 2015, 34, 1261–1265. [Google Scholar] [CrossRef]
- Jones, C.M.; Miller, T.L.; Grutter, A.S.; Cribb, T.H. Natatory-stage cymothoid isopods: Description, molecular identification and evolution of attachment. Int. J. Parasitol. 2008, 38, 477–491. [Google Scholar] [CrossRef]
- Shodipo, M.O.; Gomez, R.D.C.; Welicky, R.L.; Sikkel, P.C. Apparent kleptoparasitism in fish—parasitic gnathiid isopods. Parasitol. Res. 2018, 118, 653–655. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, M.D.; Artim, J.D.; Hendrick, G.C.; Packard, A.J.; Sikkel, P.C. Fish-Parasitic Gnathiid Isopods Metamorphose Following Invertebrate-Derived Meal. J. Parasitol. 2019, 105, 793. [Google Scholar] [CrossRef] [PubMed]
- Ota, Y.; Hoshino, O.; Hirose, M.; Tanaka, K.; Hirose, E. Third-stage larva shifts host fish from teleost to elasmobranch in the temporary parasitic isopod, Gnathia trimaculata (Crustacea; Gnathiidae). Mar. Biol. 2012, 159, 2333–2347. [Google Scholar] [CrossRef]
- Sikkel, P.C.; Welicky, R.L.; Artim, J.M.; McCammon, A.M.; Sellers, J.C.; Coile, A.M.; Jenkins, W.G. Nocturnal migration reduces exposure to micropredation in a coral reef fish. Bull. Mar. Sci. 2017, 93, 475–489. [Google Scholar] [CrossRef]
- Artim, J.M.; Sikkel, P.C. Live coral repels a common reef fish ectoparasite. Coral Reefs 2013, 32, 487–494. [Google Scholar] [CrossRef]
- Artim, J.M.; Nicholson, M.D.; Hendrick, G.C.; Brandt, M.; Smith, T.; Sikkel, P.C. Abundance of a Cryptic Generalist Parasite Reflects Degradation of an Ecosystem. Ecosphere 2020, in press. [Google Scholar]
- Dupont, S.; Pörtner, H. Get ready for ocean acidification. Nature 2013, 498, 429. [Google Scholar] [CrossRef]
- Paula, J.R.; Otjacques, E.; Hildebrandt, C.; Grutter, A.S.; Rosa, R. Ocean Acidification Does Not Affect Fish Ectoparasite Survival. Oceans 2020, 1, 3. [Google Scholar] [CrossRef] [Green Version]
- Carlson, C.J.; Burgio, K.R.; Dougherty, E.R.; Phillips, A.J.; Bueno, V.M.; Clements, C.F.; Castaldo, G.; Dallas, T.A.; Cizauskas, C.A.; Cumming, G.S.; et al. Parasite biodiversity faces extinction and redistribution in a changing climate. Sci. Adv. 2017, 3. [Google Scholar] [CrossRef] [Green Version]
- Carlson, C.J.; Cizauskas, C.A.; Burgio, K.R.; Clements, C.F.; Harris, N.C. The More Parasites, the Better? Science 2013, 342, 1041. [Google Scholar] [CrossRef] [Green Version]
- Cizauskas, C.A.; Carlson, C.J.; Burgio, K.R.; Clements, C.F.; Dougherty, E.R.; Harris, N.C.; Phillips, A.J. Parasite vulnerability to climate change: An evidence-based functional trait approach. R. Soc. Open Sci. 2017, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
Figure 1.
Lizard Island Research Station; Kaplan-Meier survival curves for unfed gnathiids per temperature treatment. Shaded areas are 95% confidence intervals.
Figure 1.
Lizard Island Research Station; Kaplan-Meier survival curves for unfed gnathiids per temperature treatment. Shaded areas are 95% confidence intervals.
Figure 2.
Great Barrier Reef; Kaplan-Meier survival curves for fed gnathiids per temperature treatment for (a) stage one, (b) stage two, and (c) stage three juveniles. For ease of interpretation, 95% confidence intervals are not included.
Figure 2.
Great Barrier Reef; Kaplan-Meier survival curves for fed gnathiids per temperature treatment for (a) stage one, (b) stage two, and (c) stage three juveniles. For ease of interpretation, 95% confidence intervals are not included.


Figure 3.
Philippines; Trial 1, Kaplan-Meier survival curves for gnathiids per temperature treatment. Shaded areas are 95% confidence intervals.
Figure 3.
Philippines; Trial 1, Kaplan-Meier survival curves for gnathiids per temperature treatment. Shaded areas are 95% confidence intervals.

Figure 4.
Philippines; Trial 2, Kaplan-Meier survival curves for gnathiids per temperature treatment. Shaded areas are 95% confidence intervals.
Figure 4.
Philippines; Trial 2, Kaplan-Meier survival curves for gnathiids per temperature treatment. Shaded areas are 95% confidence intervals.


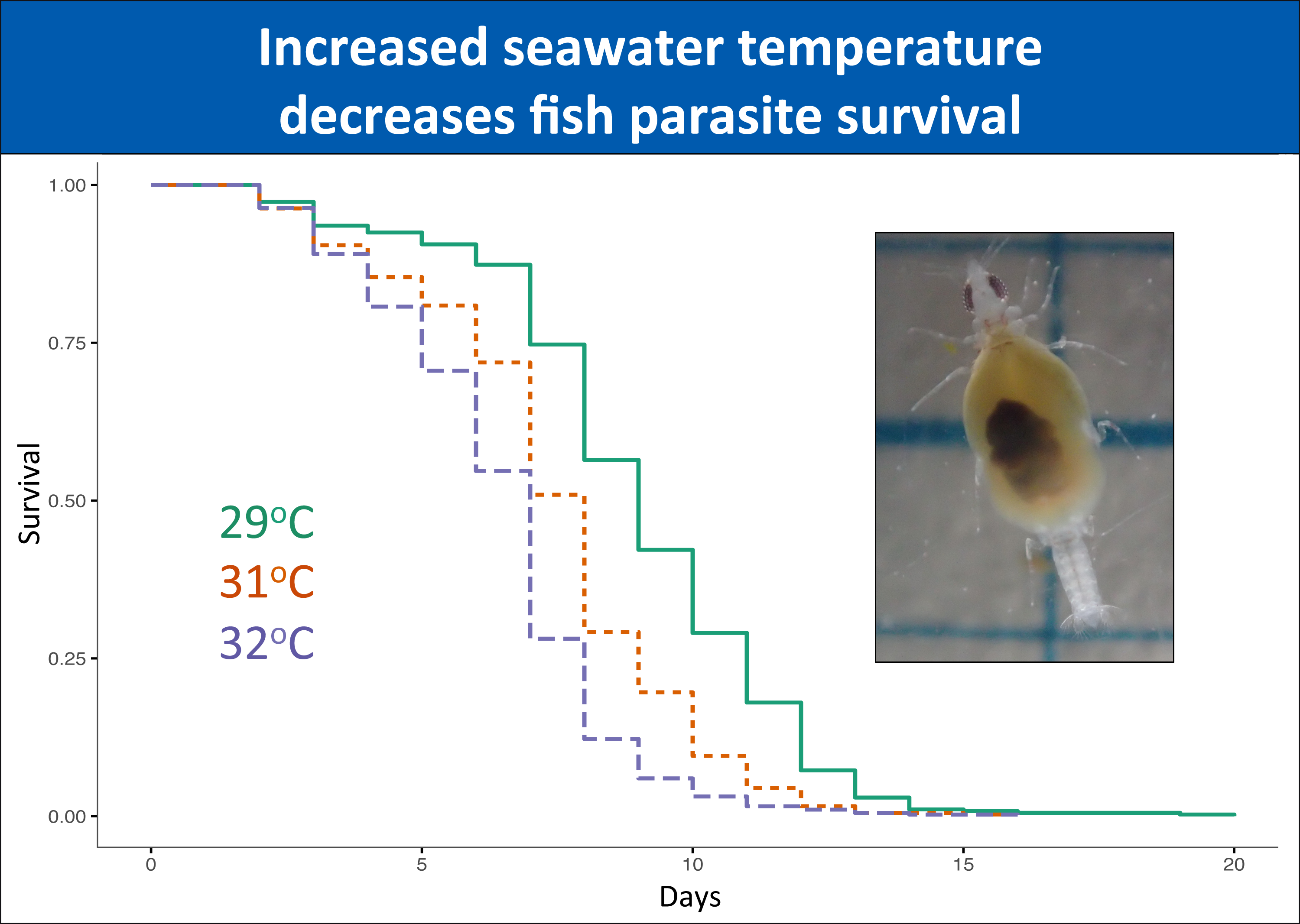{kind=link}
{kind=link}
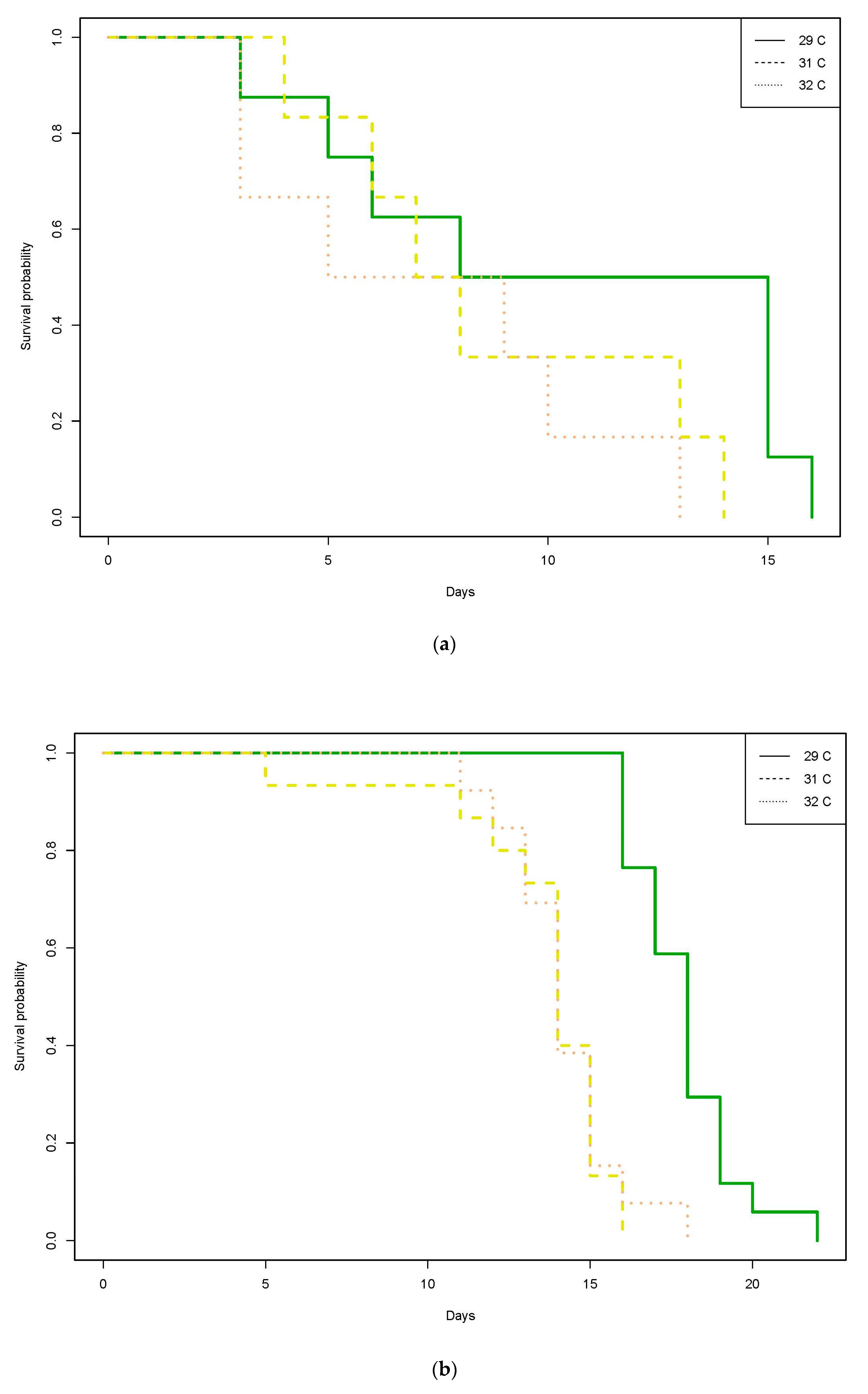{kind=link}
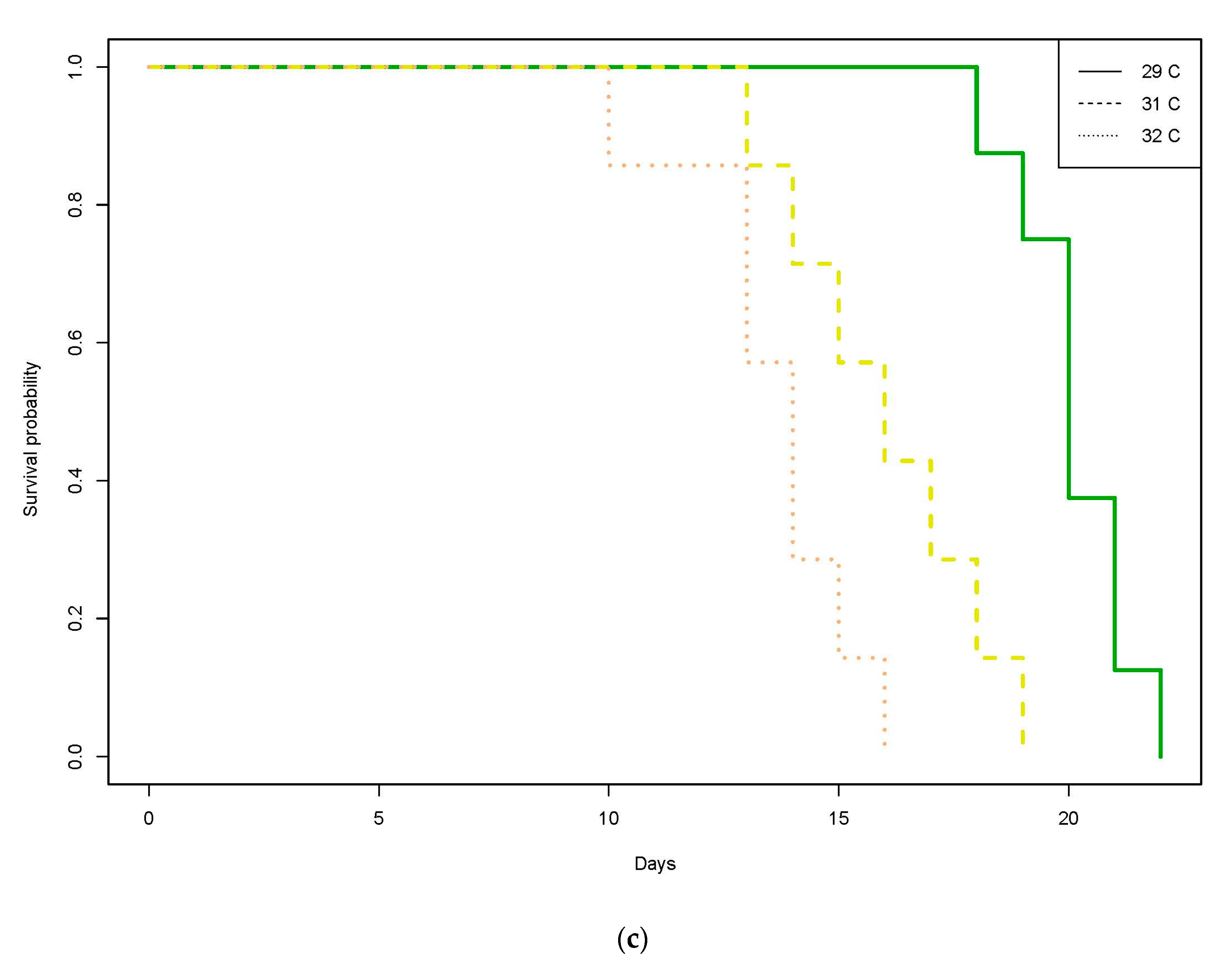{kind=link}
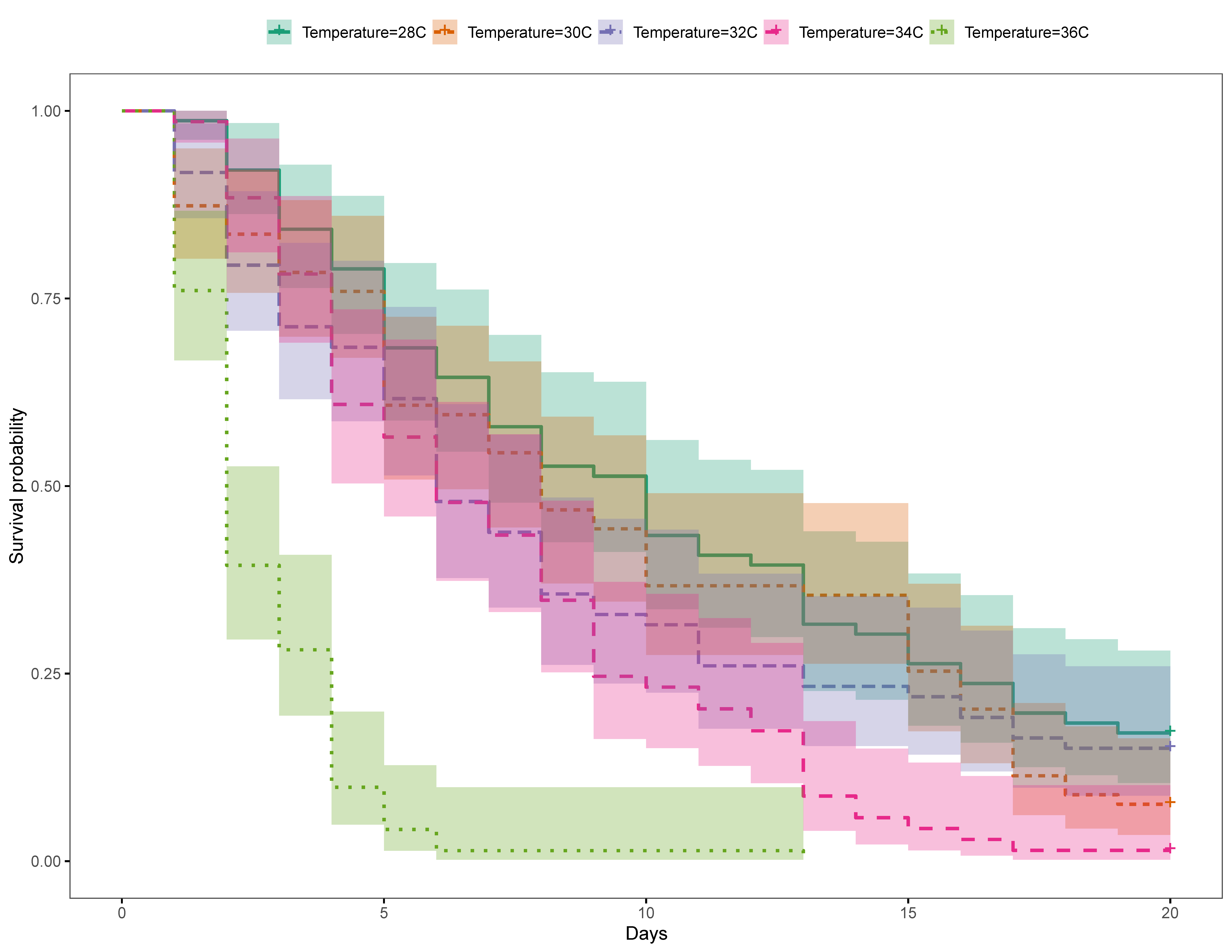{kind=link}
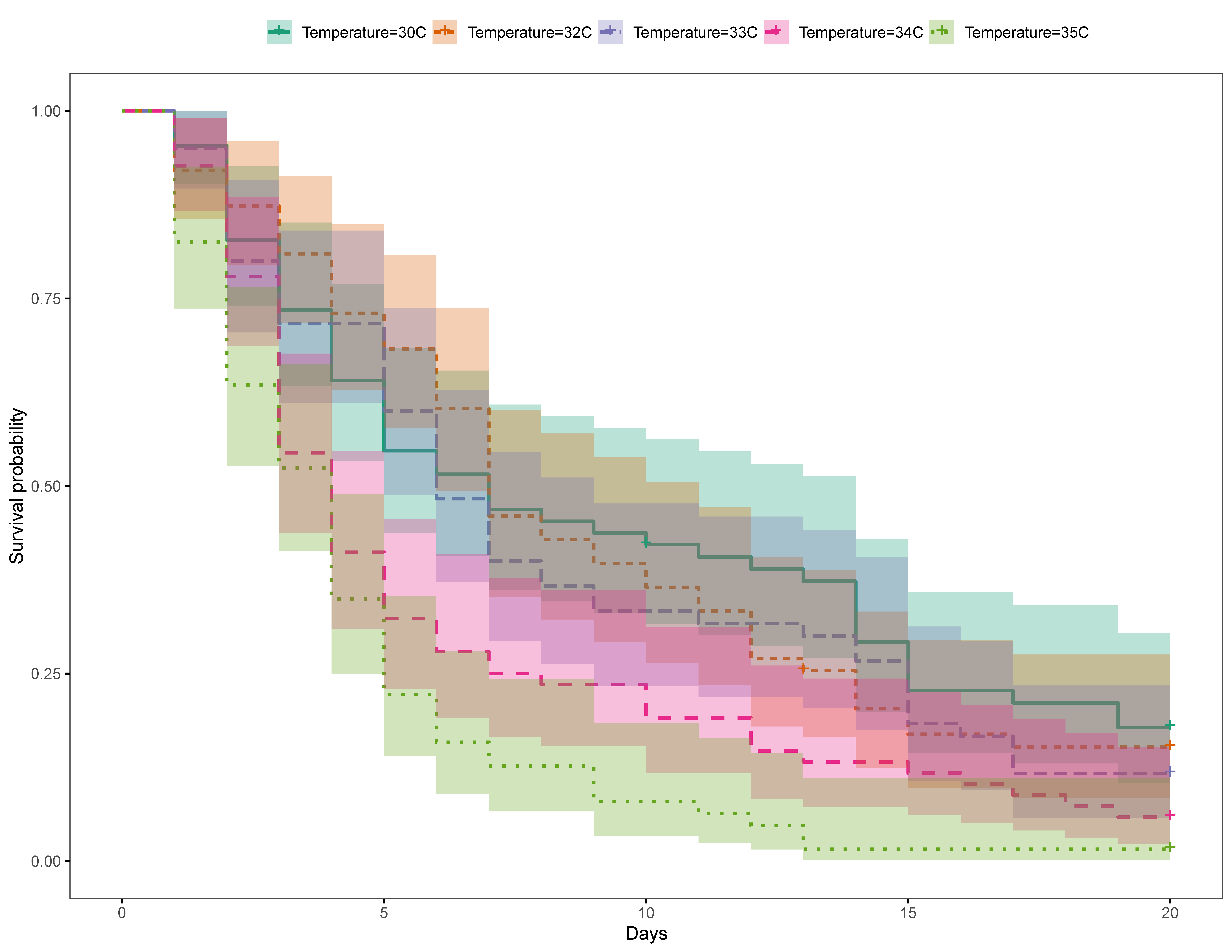{kind=link}
Table 1.
Great Barrier Reef; Analysis of deviance table (Type II tests) for unfed gnathiid survival among temperature treatments and juvenile stages for Cox model. Bolded values are ones mentioned in main text. *** p < 0.001.
Table 1.
Great Barrier Reef; Analysis of deviance table (Type II tests) for unfed gnathiid survival among temperature treatments and juvenile stages for Cox model. Bolded values are ones mentioned in main text. *** p < 0.001.
| Df | Chisq | Pr(>Chisq) | ||
|---|---|---|---|---|
| Temperature | 2 | 183.76 | <0.0001 | *** |
| Headwidth | 1 | 1.87 | 0.1719 | |
| Stage | 2 | 26.93 | <0.0001 | *** |
| Temperature × Headwidth | 2 | 0.07 | 0.9639 | |
| Temperature × Stage | 4 | 3.06 | 0.5471 | |
| Headwidth × Stage | 2 | 23.85 | <0.0001 | *** |
| Temperature × Headwidth × Stage | 4 | 6.51 | 0.1642 |
Table 2.
Great Barrier Reef; Analysis of deviance table (Type II tests) for fed gnathiid survival among temperature treatments and juvenile stages for Cox model. Bolded values are ones mentioned in main text. ** p < 0.01, *** p < 0.001.
Table 2.
Great Barrier Reef; Analysis of deviance table (Type II tests) for fed gnathiid survival among temperature treatments and juvenile stages for Cox model. Bolded values are ones mentioned in main text. ** p < 0.01, *** p < 0.001.
| Df | Chisq | Pr(>Chisq) | ||
|---|---|---|---|---|
| Temperature | 2 | 28.2063 | 7.50 × 10−7 | *** |
| Headwidth | 1 | 1.9549 | 0.16206 | |
| Stage | 2 | 21.3179 | 2.35 × 10−5 | *** |
| Temperature × Headwidth | 2 | 6.0753 | 0.04795 | * |
| Temperature × Stage | 4 | 13.6392 | 0.00854 | ** |
| Headwidth × Stage | 2 | 3.0686 | 0.21561 | |
| Temperature × Headwidth × Stage | 4 | 3.3548 | 0.5003 |
Table 3.
Philippines; Analysis of deviance table (Type II tests) for Trial 1 and 2 gnathiid survival for Cox model. Bolded values are ones mentioned in main text.
Table 3.
Philippines; Analysis of deviance table (Type II tests) for Trial 1 and 2 gnathiid survival for Cox model. Bolded values are ones mentioned in main text.
| Trial | Df | Chisq | Pr(>Chisq) | |
|---|---|---|---|---|
| 1 | Temperature | 4 | 24.927 | <0.0001 |
| 2 | Temperature | 4 | 8.4374 | 0.07681 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Shodipo, M.O.; Duong, B.; Graba-Landry, A.; Grutter, A.S.; Sikkel, P.C. Effect of Acute Seawater Temperature Increase on the Survival of a Fish Ectoparasite. Oceans 2020, 1, 215-236. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans1040016
AMA Style
Shodipo MO, Duong B, Graba-Landry A, Grutter AS, Sikkel PC. Effect of Acute Seawater Temperature Increase on the Survival of a Fish Ectoparasite. Oceans. 2020; 1(4):215-236. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans1040016
Chicago/Turabian StyleShodipo, Mary O., Berilin Duong, Alexia Graba-Landry, Alexandra S. Grutter, and Paul C. Sikkel. 2020. "Effect of Acute Seawater Temperature Increase on the Survival of a Fish Ectoparasite" Oceans 1, no. 4: 215-236. https://0-doi-org.brum.beds.ac.uk/10.3390/oceans1040016