
Selection of Unique Molecules for Cancer Treatment by Distance-Based Method: Hypericin Effect on Respiratory Chain



Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Cultures
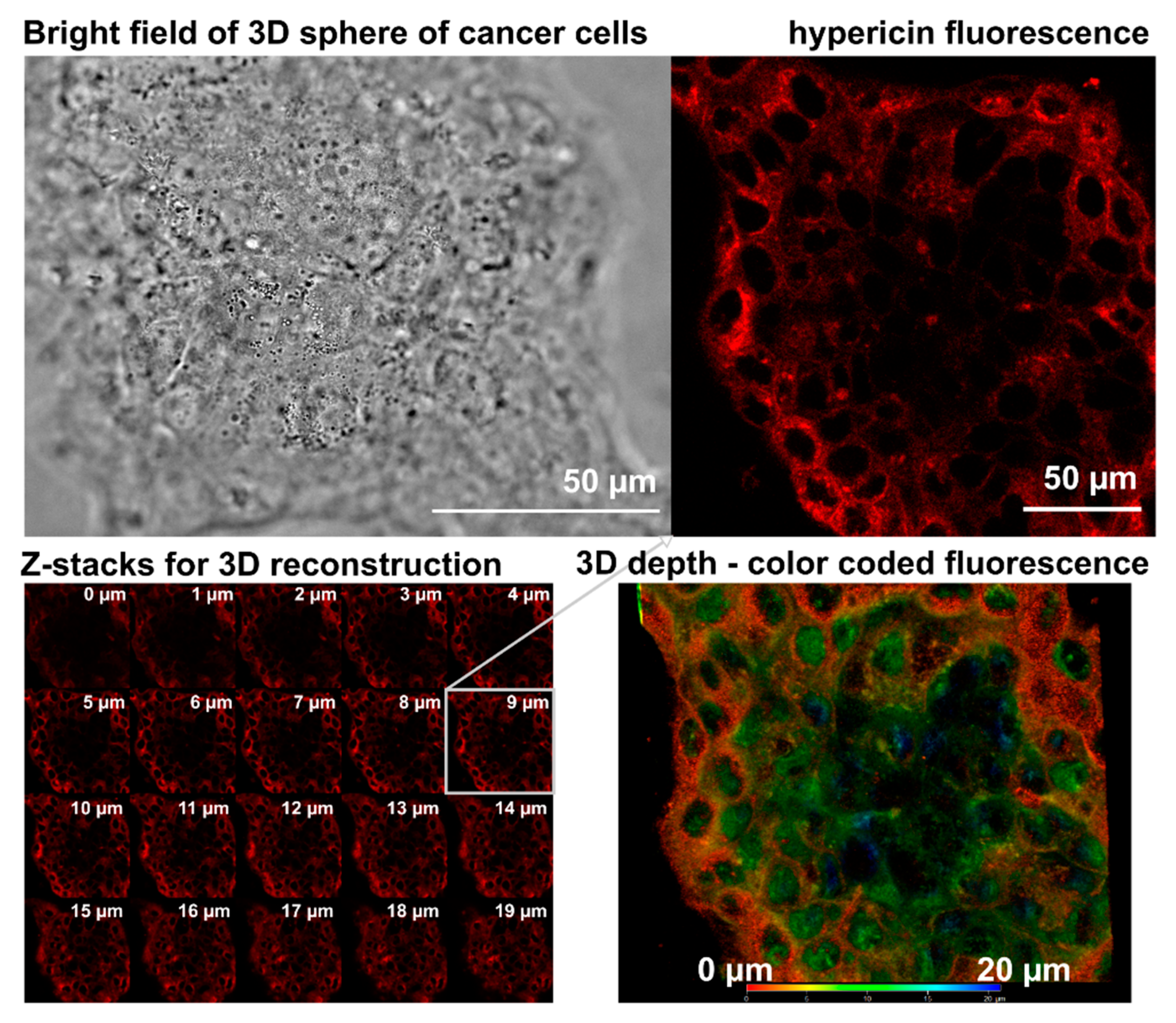
2.2. Confocal Fluorescence Microscopy
2.3. Western Blot
2.4. MTT Metabolic Assay
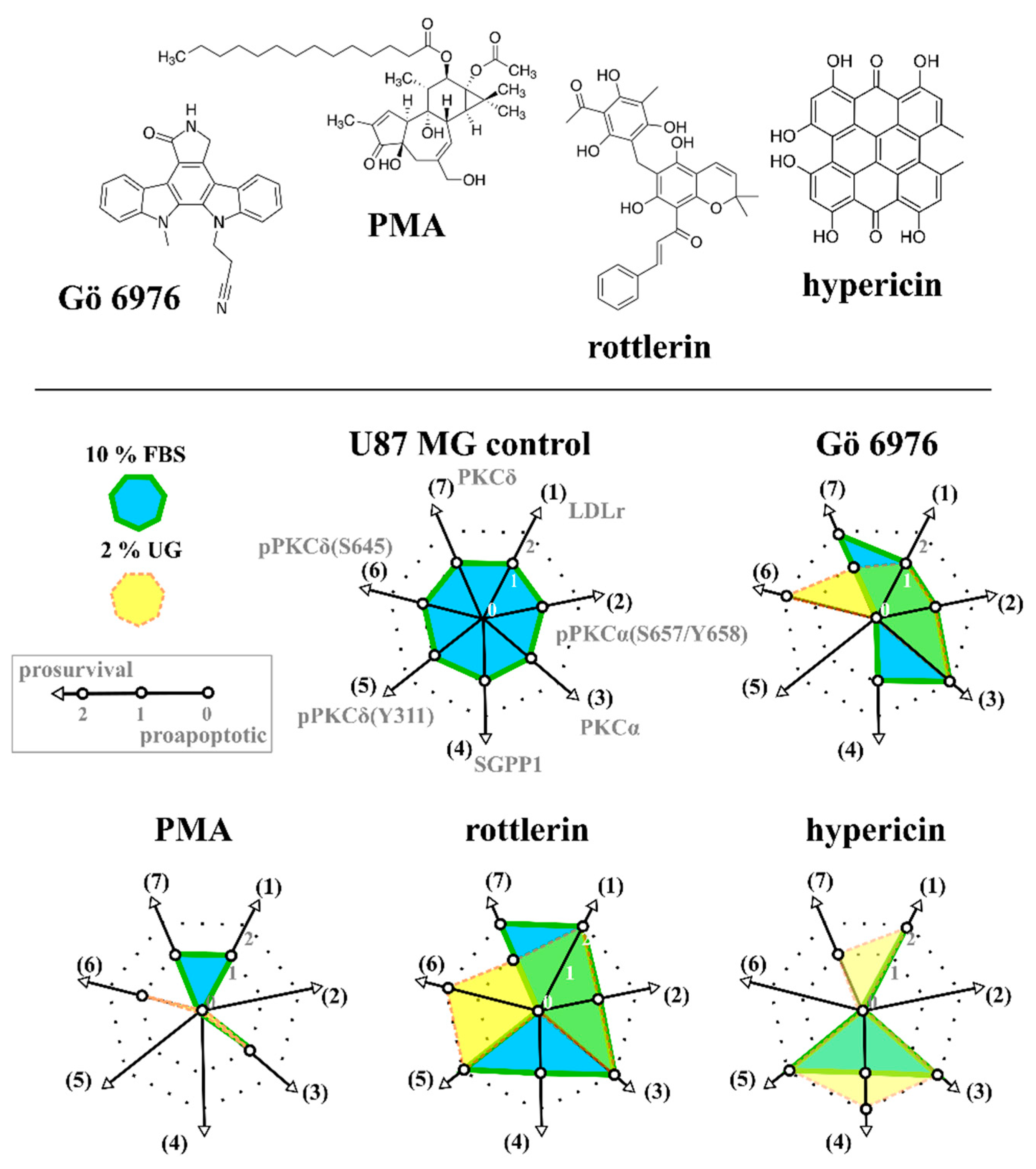
2.5. Evaluation Criteria to Create Spider Diagram Presentation
3. Results and Discussion
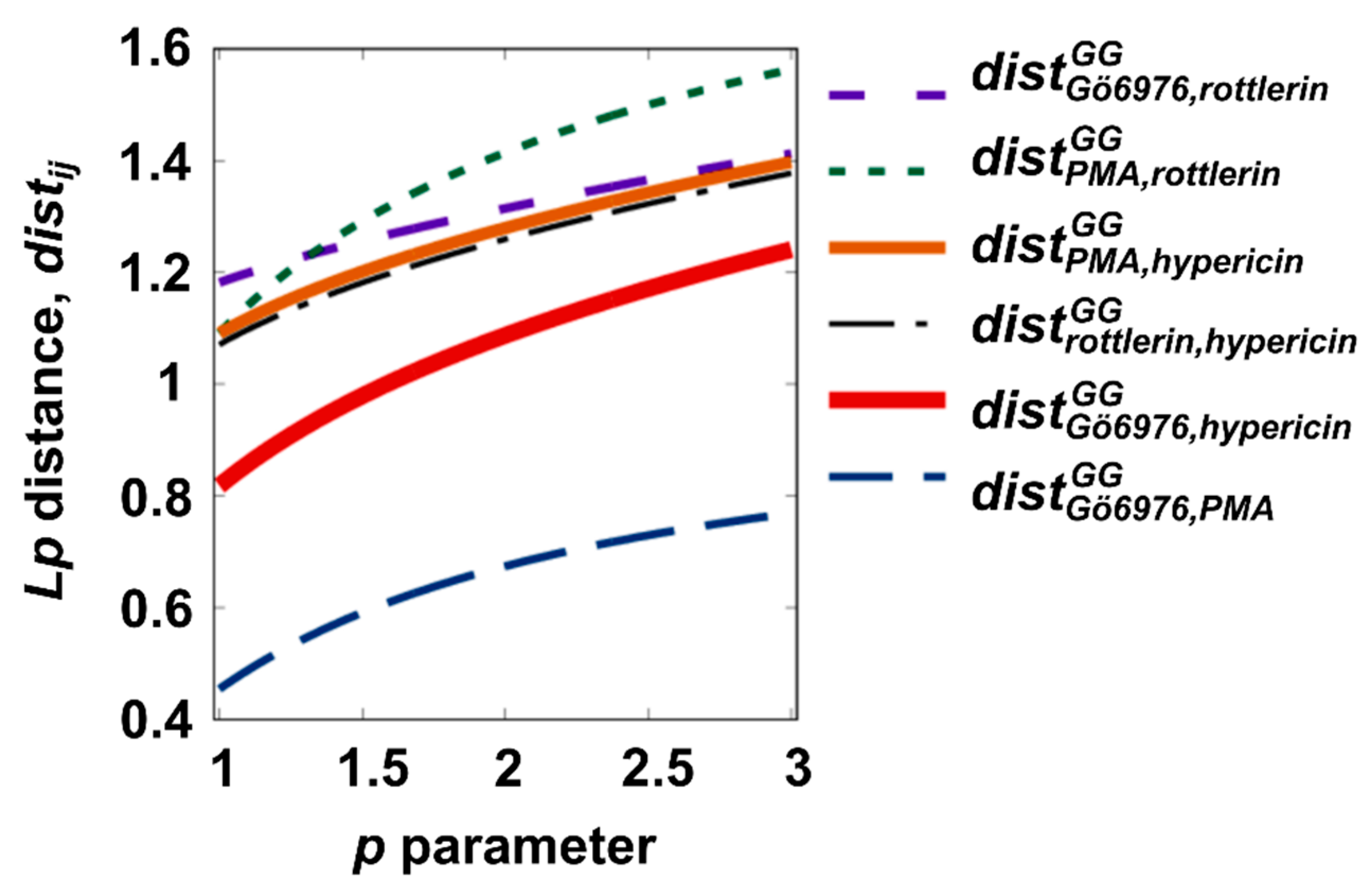
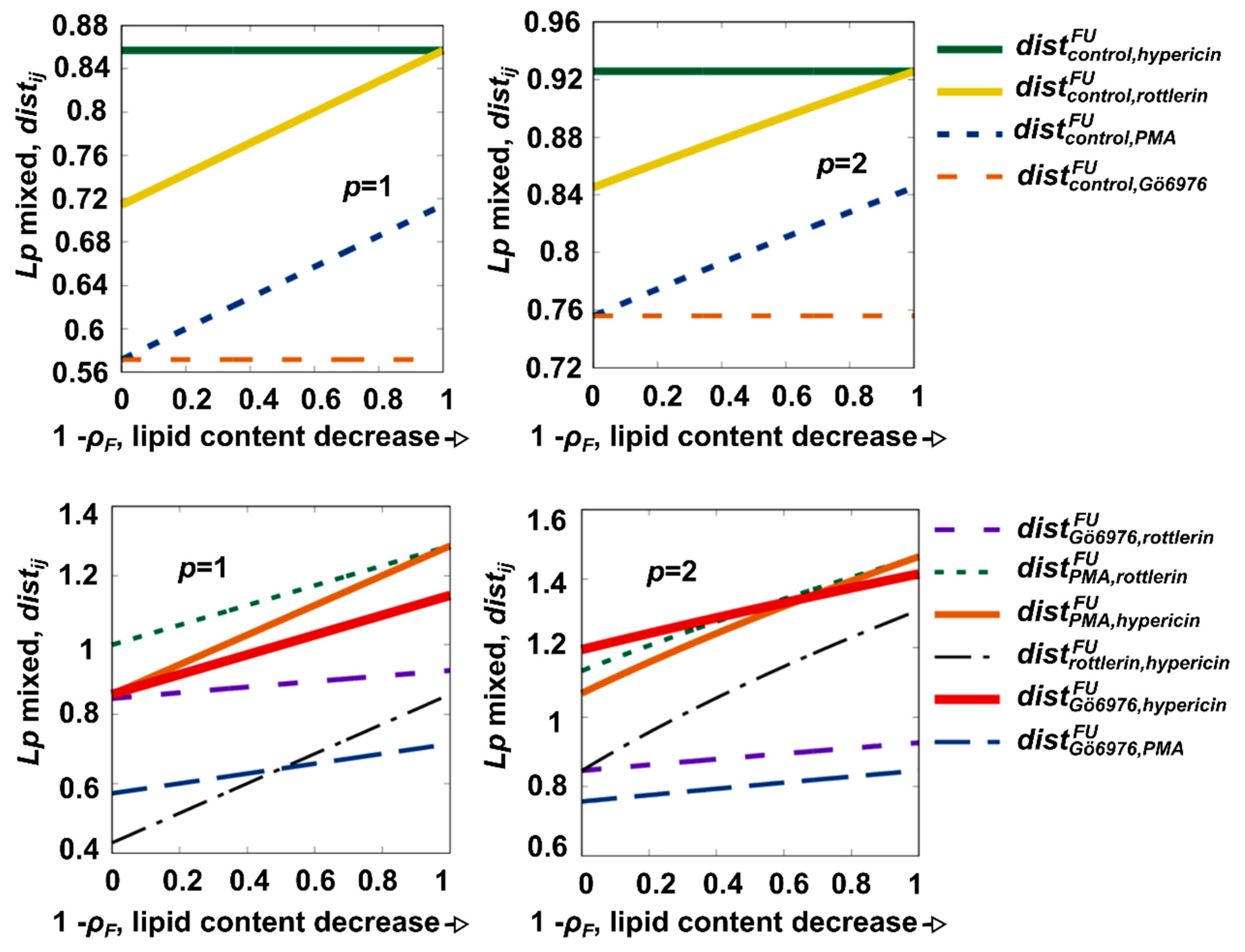
3.1. Selection of an Extraordinary Molecule between PKC Regulators by Distance-Based Analysis
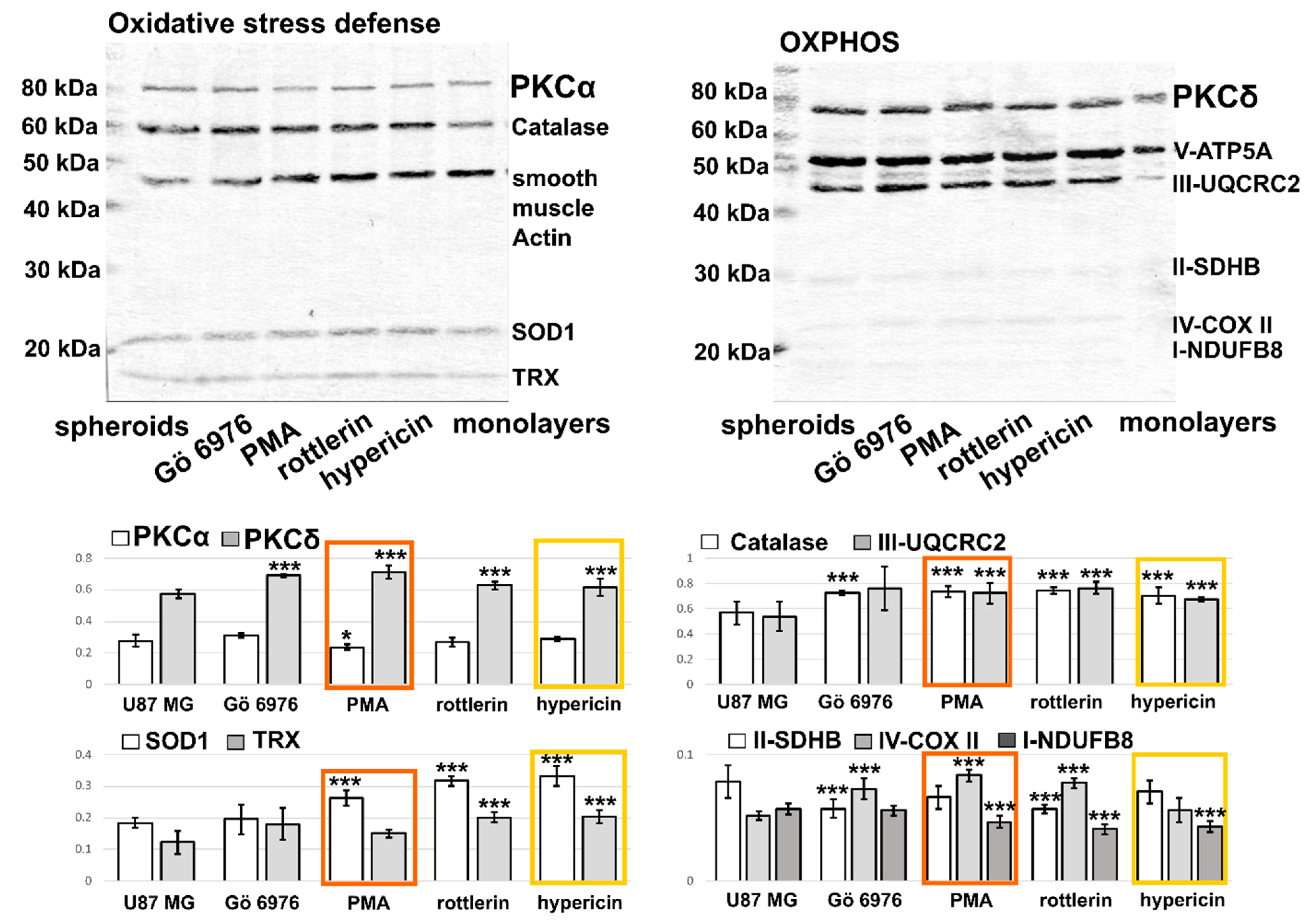
3.2. Validation of Selected Molecule in Monolayers and Spheroids of U87 MG Cells by Western Blot
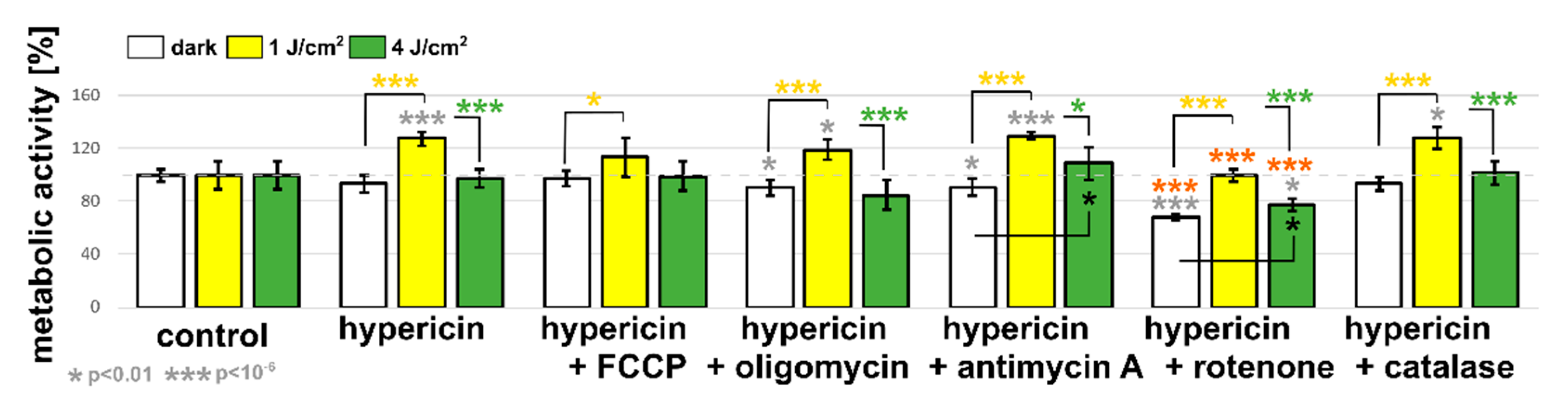
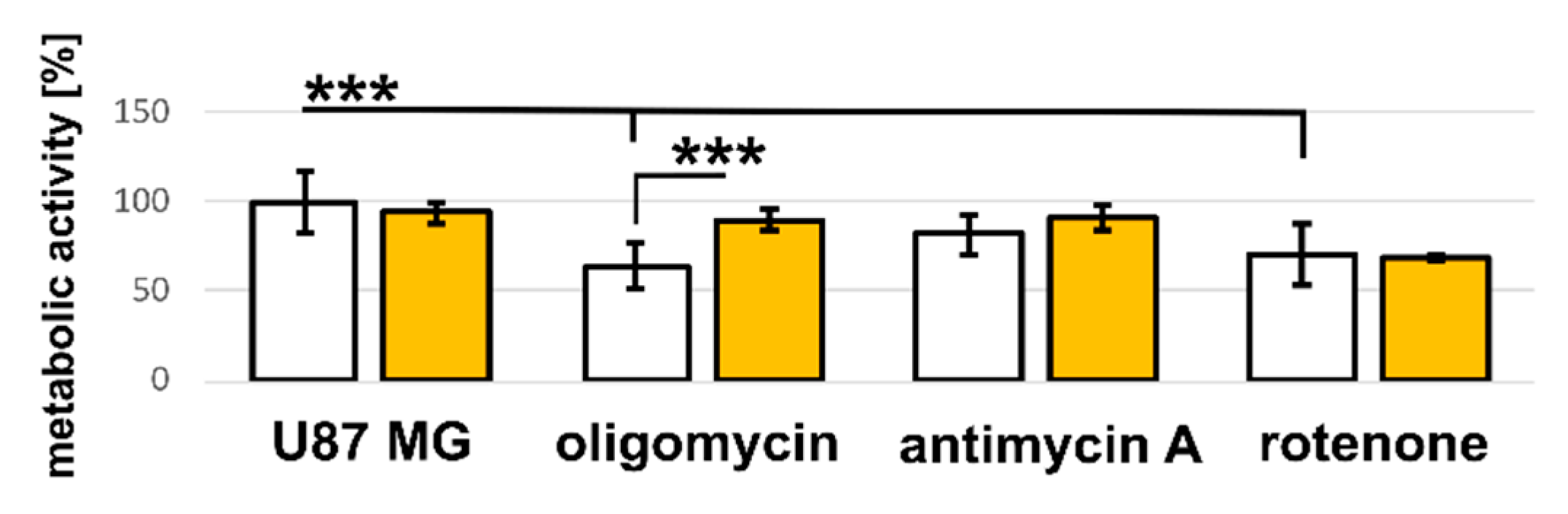
3.3. U87 MG Cells’ Response to Hypericin Treatment of the Monolayers
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yau, S.-T.; Nadis, S. The Shape of Inner Space; Basic Books: New York, NY, USA, 2010; ISBN 978-0-465-02023-2. [Google Scholar]
- Roweis, S.T.; Saul, L.K. Nonlinear dimensionality reduction by locally linear embedding. Science 2000, 290, 2323–2326. [Google Scholar] [CrossRef] [Green Version]
- Nies, H.W.; Zakaria, Z.; Mohamad, M.S.; Chan, W.H.; Zaki, N.; Sinnott, R.O.; Napis, S.; Chamoso, P.; Omatu, S.; Corchado, J.M. A review of computational methods for clustering genes with similar biological functions. Processes 2019, 7, 550. [Google Scholar] [CrossRef] [Green Version]
- Yoshioka, P.M. Misidentification of the Bray-Curtis similarity index. Mar. Ecol. Prog. Ser. 2008, 368. [Google Scholar] [CrossRef] [Green Version]
- Jukes, T.H.; Cantor, C.R. Evolution of protein molecules BT—Mammalian protein metabolism. In Mammalian Protein Metabolism; Academic Press: New York, NY, USA, 1969; Volume III. [Google Scholar]
- Lenkavska, L.; Tomkova, S.; Horvath, D.; Huntosova, V. Searching for combination therapy by clustering methods: Stimulation of PKC in Golgi apparatus combined with hypericin induced PDT. Photodiagnosis Photodyn. Ther. 2020, 31. [Google Scholar] [CrossRef] [PubMed]
- Huntosova, V.; Horvath, D.; Seliga, R.; Wagnieres, G. Influence of Oxidative Stress on Time-Resolved Oxygen Detection by [Ru(Phen)3]2+ In Vivo and In Vitro. Molecules 2021, 26, 485. [Google Scholar] [CrossRef]
- Amos, S.; Mut, M.; DiPierro, C.G.; Carpenter, J.E.; Xiao, A.; Kohutek, Z.A.; Redpath, G.T.; Zhao, Y.; Wang, J.; Shaffrey, M.E.; et al. Protein kinase C-α-mediated regulation of low-density lipoprotein receptor-related protein and urokinase increases astrocytoma invasion. Cancer Res. 2007, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bornancin, F.; Parker, P.J. Phosphorylation of threonine 638 critically controls the dephosphorylation and inactivation of protein kinase Cα. Curr. Biol. 1996, 6. [Google Scholar] [CrossRef] [Green Version]
- Luo, B.; Prescott, S.M.; Topham, M.K. Protein kinase Cα phosphorylates and negatively regulates diacylglycerol kinase ζ. J. Biol. Chem. 2003, 278. [Google Scholar] [CrossRef] [Green Version]
- Reyland, M.E. Protein kinase Cδ and apoptosis. Biochem. Soc. Trans. 2007, 35, 1001–1004. [Google Scholar] [CrossRef]
- Larroque-Cardoso, P.; Swiader, A.; Ingueneau, C.; Nègre-Salvayre, A.; Elbaz, M.; Reyland, M.E.; Salvayre, R.; Vindis, C. Role of protein kinase C δ in ER stress and apoptosis induced by oxidized LDL in human vascular smooth muscle cells. Cell Death Dis. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Basu, A.; Pal, D. Two faces of protein kinase Cδ: The contrasting roles of PKCδ in cell survival and cell death. Sci. World J. 2010, 10, 673080. [Google Scholar] [CrossRef]
- Le, T.L.; Joseph, S.R.; Yap, A.S.; Stow, J.L. Protein kinase C regulates endocytosis and recycling of E-cadherin. Am. J. Physiol. Cell Physiol. 2002, 283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, K.R.; Becker, K.P.; Facchinetti, M.M.; Hannun, Y.A.; Obeid, L.M. PKC-dependent Activation of Sphingosine Kinase 1 and Translocation to the Plasma Membrane. J. Biol. Chem. 2002, 277. [Google Scholar] [CrossRef] [Green Version]
- Conway, A.M.; Pyne, N.J.; Pyne, S. Ceramide-dependent regulation of p42/p44 mitogen-activated protein kinase and c-Jun N-terminal-directed protein kinase in cultured airway smooth muscle cells. Cell. Signal. 2000, 12. [Google Scholar] [CrossRef]
- Newton, A.C. Protein kinase C: Poised to signal. Am. J. Physiol. Endocrinol. Metab. 2010, 298. [Google Scholar] [CrossRef] [Green Version]
- Newton, A.C. Lipid activation of protein kinases. J. Lipid. Res. 2009, 50, S266–S271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steinberg, S.F. Structural basis of protein kinase C isoform function. Physiol. Rev. 2008, 88, 1341–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misuth, M.; Joniova, J.; Horvath, D.; Dzurova, L.; Nichtova, Z.; Novotova, M.; Miskovsky, P.; Stroffekova, K.; Huntosova, V. The flashlights on a distinct role of protein kinase C δ: Phosphorylation of regulatory and catalytic domain upon oxidative stress in glioma cells. Cell. Signal. 2017, 34. [Google Scholar] [CrossRef] [PubMed]
- Dzurová, L.; Petrovajova, D.; Nadova, Z.; Huntosova, V.; Miskovsky, P.; Stroffekova, K. The role of anti-apoptotic protein kinase Cα in response to hypericin photodynamic therapy in U-87 MG cells. Photodiagnosis Photodyn. Ther. 2014, 11. [Google Scholar] [CrossRef] [PubMed]
- Huntosova, V.; Stroffekova, K. Hypericin in the dark: Foe or ally in photodynamic therapy? Cancers 2016, 8, 93. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, S. Protein Kinase C{alpha} (PKC{alpha}): Regulation and Biological Function. J. Biochem. 2002, 132, 669–675. [Google Scholar] [CrossRef] [PubMed]
- Bornancin, F.; Parker, P.J. Phosphorylation of protein kinase C-α on serine 657 controls the accumulation of active enzyme and contributes to its phosphatase-resistant state. J. Biol. Chem. 1997, 272. [Google Scholar] [CrossRef] [Green Version]
- Rezk, C. A model for the homotopy theory of homotopy theory. Trans. Am. Math. Soc. 2000, 353. [Google Scholar] [CrossRef] [Green Version]
- Bergner, J.E. Three models for the homotopy theory of homotopy theories. Topology 2007, 46. [Google Scholar] [CrossRef] [Green Version]
- Awodey, S. Natural models of homotopy type theory. Math. Struct. Comput. Sci. 2018, 28. [Google Scholar] [CrossRef] [Green Version]
- Shen, H.C.; Bie, C.Y.C.; Chiu, D.K.Y. A texture-based distance measure for classification. Pattern Recognit. 1993, 26. [Google Scholar] [CrossRef]
- Horvath, D.; Ulicny, J.; Brutovsky, B. Self-organised manifold learning and heuristic charting via adaptive metrics. Conn. Sci. 2016, 28. [Google Scholar] [CrossRef]
- Angerer, T.B.; Magnusson, Y.; Landberg, G.; Fletcher, J.S. Lipid heterogeneity resulting from fatty acid processing in the human breast cancer microenvironment identified by GCIB-ToFSIMS imaging. Anal. Chem. 2016, 88. [Google Scholar] [CrossRef] [PubMed]
- Long, J.; Zhang, C.-J.; Zhu, N.; Du, K.; Yin, Y.-F.; Tan, X.; Liao, D.-F.; Qin, L. Lipid metabolism and carcinogenesis, cancer development. Am. J. Cancer Res. 2018, 8, 778. [Google Scholar]
- Deisboeck, T.S.; Wang, Z.; MacKlin, P.; Cristini, V. Multiscale cancer modeling. Annu. Rev. Biomed. Eng. 2011, 13. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Bordas, V.; Deisboeck, T.S. Discovering molecular targets in cancer with multiscale modeling. Drug Dev. Res. 2011, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, D.; Brutovsky, B. Toward understanding of the role of reversibility of phenotypic switching in the evolution of resistance to therapy. Phys. Lett. A 2018, 382, 1586–1600. [Google Scholar] [CrossRef] [Green Version]
- Huntosova, V.; Wagnieres, G. CHAPTER 15: pO2 Measurements in Biological Tissues by Luminescence Lifetime Spectroscopy: Strategies to Exploit or Minimize Phototoxic Effects in Tumors. In Quenched-Phosphorescence Detection of Molecular Oxygen; Royal Society of Chemistry: London, UK, 2018. [Google Scholar]
- Boneh, A.; Mandla, S.; Tenenhouse, H.S. Phorbol myristate acetate activates protein kinase C, stimulates the phosphorylation of endogenous proteins and inhibits phosphate transport in mouse renal tubules. Biochim. Biophys. Acta Mol. Cell Res. 1989, 1012. [Google Scholar] [CrossRef]
- Misuth, M.; Joniova, J.; Belej, D.; Hrivnak, S.; Horvath, D.; Huntosova, V. Estimation of PKCδ autophosphorylation in U87 MG glioma cells: Combination of experimental, conceptual and numerical approaches. J. Biophotonics 2017, 10. [Google Scholar] [CrossRef] [PubMed]
- Tomkova, S.; Misuth, M.; Lenkavska, L.; Miskovsky, P.; Huntosova, V. In vitro identification of mitochondrial oxidative stress production by time-resolved fluorescence imaging of glioma cells. Biochim. Biophys. Acta Mol. Cell Res. 2018, 1865. [Google Scholar] [CrossRef]
- Grimm, S. Respiratory chain complex II as general sensor for apoptosis. Biochim. Biophys. Acta Bioenerg. 2013, 1827, 565–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, R.Z.; Jiang, S.; Zhang, L.; Yu, Z. Bin Mitochondrial electron transport chain, ROS generation and uncoupling (Review). Int. J. Mol. Med. 2019, 44, 3–15. [Google Scholar]
- Ighodaro, O.M.; Akinloye, O.A. First line defence antioxidants-superoxide dismutase (SOD), catalase (CAT) and glutathione peroxidase (GPX): Their fundamental role in the entire antioxidant defence grid. Alex. J. Med. 2018, 54. [Google Scholar] [CrossRef] [Green Version]
- Ferrall-Fairbanks, M.C.; Ball, M.; Padron, E.; Altrock, P.M. Leveraging Single-Cell RNA Sequencing Experiments to Model Intratumor Heterogeneity. JCO Clin. Cancer Inform. 2019. [Google Scholar] [CrossRef]
- Papandreou, I.; Cairns, R.A.; Fontana, L.; Lim, A.L.; Denko, N.C. HIF-1 mediates adaptation to hypoxia by actively downregulating mitochondrial oxygen consumption. Cell Metab. 2006, 3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semenza, G.L.; Jiang, B.H.; Leung, S.W.; Passantino, R.; Concordat, J.P.; Maire, P.; Giallongo, A. Hypoxia response elements in the aldolase A, enolase 1, and lactate dehydrogenase a gene promoters contain essential binding sites for hypoxia-inducible factor 1. J. Biol. Chem. 1996, 271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huntosova, V.; Novotova, M.; Nichtova, Z.; Balogova, L.; Maslanakova, M.; Petrovajova, D.; Stroffekova, K. Assessing light-independent effects of hypericin on cell viability, ultrastructure and metabolism in human glioma and endothelial cells. Toxicol. Vitr. 2017, 40. [Google Scholar] [CrossRef] [PubMed]
- Berridge, M.V.; Tan, A.S. Characterization of the Cellular Reduction of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT): Subcellular Localization, Substrate Dependence, and Involvement of Mitochondrial Electron Transport in MTT Reduction. Arch. Biochem. Biophys. 1993, 303. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecule | Metabolism | Endocytosis | Respiration |
|---|---|---|---|
| (1)(2)(3)(4) | (5)(6)(7) | (8)(9)(10)(11) | |
| - | 1, 1, 1, 1 | 1, 1, 1, | 1, 1, 1, 1 |
| Gö6976 | 0, 2, 2, 1 | 1, 1, 1 | 1, 1, 1, 1 |
| PMA | 0, 2, 2, 1 | 0, 0, 2 | 1, 1, 2, 2 |
| rottlerin | 2, 0, 0, 1 | 2, 2, 2 | 2, 2, 2, 2 |
| hypericin | 1, 0, 0, 0 | 1, 0, 0 | 0, 1, 1, 1 |
| Molecule | in 10% FBS | in 2% UG |
|---|---|---|
| (1)(2)(3)(4)(5)(6)(7) | (1)(2)(3)(4)(5)(6)(7) | |
| - | 1, 1, 1, 1, 1, 1, 1 | 1, 1, 1, 1, 1, 1, 1 |
| Gö6976 | 1, 1, 2, 1, 0, 2, 1 | 1, 1, 2, 0, 0, 2, 1 |
| PMA | 1, 0, 1, 0, 0, 0, 1 | 0, 0, 1, 0, 0, 1, 0 |
| rottlerin | 2, 1, 2, 1, 2, 0, 2 | 2, 1, 2, 0, 2, 2, 1 |
| hypericin | 2, 0, 2, 1, 2, 0, 0 | 2, 0, 2, 2, 2, 0, 1 |
| Parameter | Pro-Apoptotic | Prosurvival | ||
|---|---|---|---|---|
| 10% FBS | 2% UG | 10% FBS | 2% UG | |
| Gö 6976 | 28 | 28 | 28 | 28 |
| PMA | 57 | 71 | 0 | 0 |
| rottlerin | 14 | 14 | 57 | 71 |
| hypericin | 28 | 28 | 42 | 57 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horvath, D.; Tomkova, S.; Huntosova, V. Selection of Unique Molecules for Cancer Treatment by Distance-Based Method: Hypericin Effect on Respiratory Chain. Biophysica 2021, 1, 222-237. https://0-doi-org.brum.beds.ac.uk/10.3390/biophysica1020017
Horvath D, Tomkova S, Huntosova V. Selection of Unique Molecules for Cancer Treatment by Distance-Based Method: Hypericin Effect on Respiratory Chain. Biophysica. 2021; 1(2):222-237. https://0-doi-org.brum.beds.ac.uk/10.3390/biophysica1020017
Chicago/Turabian StyleHorvath, Denis, Silvia Tomkova, and Veronika Huntosova. 2021. "Selection of Unique Molecules for Cancer Treatment by Distance-Based Method: Hypericin Effect on Respiratory Chain" Biophysica 1, no. 2: 222-237. https://0-doi-org.brum.beds.ac.uk/10.3390/biophysica1020017