
Potential Beneficial Effects of Vitamin K in SARS-CoV-2 Induced Vascular Disease?

1
Biotheus (Hong Kong) Limited, Hong Kong Science and Technology Park, Hong Kong, China
2
Li Ka Shing Faculty of Medicine, School of Public Health, The University of Hong Kong, Hong Kong, China
3
School of Nursing, Tung Wah College, 31 Wylie Road, Homantin, Hong Kong, China
*
Author to whom correspondence should be addressed.
†
These authors contribute equally.
Immuno 2021, 1(1), 17-29; https://0-doi-org.brum.beds.ac.uk/10.3390/immuno1010003
Submission received: 21 January 2021
/
Revised: 11 February 2021
/
Accepted: 15 February 2021
/
Published: 17 February 2021
{kind=link}
Abstract
:Prevalent coagulopathy and thromboembolism are observed in severe COVID-19 patients with 40% of COVID-19 mortality being associated with cardiovascular complications. Abnormal coagulation parameters are related to poor prognosis in COVID-19 patients. Victims also displayed presence of extensive thrombosis in infected lungs. Vitamin K is well-known to play an essential role in the coagulation system. Latest study revealed an existing correlation between vitamin K deficiency and COVID-19 severity, highlighting a role of vitamin K, probably via coagulation modulation. In agreement, other recent studies also indicated that anti-coagulant treatments can reduce mortality in severe cases. Altogether, potential mechanisms linking COVID-19 with coagulopathy in which vitamin K may exert its modulating role in coagulation related with disease pathogenesis are established. In this review, we discuss the recent evidence supporting COVID-19 as a vascular disease and explore the potential benefits of using vitamin K against COVID-19 to improve disease outcomes.
1. Introduction
Since its emergence in December 2019 in Wuhan, China, the novel coronavirus known as the severe acute respiratory syndrome coronavirus-2 (SARS-CoV-2) and responsible for the disease Coronavirus disease 19 (COVID-19), has infected over 100,000,000 people and killed over 2,000,000 worldwide according to the World Health Organization [1]. Interestingly, despite intense research, the critical mechanisms underlying the patient morbidity and mortality remain largely obscure. One of the predominant theories favors the concept of a “cytokine storm” in which the immune response is exacerbated through the induction of an excessive pro-inflammatory cytokine response driving lung injury [2]. It was reported that presence of a high viral load causes massive destruction of lung tissues, in turn leading to hyperinflammation causing acute respiratory distress syndrome (ARDS) [3]. In addition to respiratory symptoms, a growing body of evidence also shows that the virus can specifically infects endothelial cells affecting thus the normal process of coagulation [4]. Severe COVID-19 patients were found to possess coagulopathy characterized by abnormal coagulation parameters [4,5] widespread presence of blood clots [6] as well as arterial and venous thromboembolism [7,8]. Furthermore, preliminary data from several studies seem to indicate that anticoagulant therapy is associated with lower mortality in COVID-19 patients [9]. Vitamin K is an essential component preventing blood clotting and a major player of the coagulation system of which a link between vitamin K deficiency and the worst COVID-19 outcomes was recently revealed [10]. In this review, we will discuss the mechanisms of COVID-19 as both respiratory and vascular disease prior to explore the potential beneficial role of vitamin K in COVID-19 pathogenesis.
2. Respiratory Illness Associated with COVID-19
It is well established that SARS-CoV-2 virus affects primarily the respiratory system with infection being both asymptomatic and symptomatic. Mechanistically, SARS-CoV-2 infection involves the binding to its functional receptor the Angiotensin converting enzyme 2 (ACE2) [11,12]. ACE2 is known to be highly expressed on lung epithelial cells as well as on endothelial cells [13]. As far as we know, clinical presentations of mild COVID-19 infection are wide-ranging and not much distinct from upper respiratory tract infections caused by various respiratory viruses such as influenza A virus (IAV) [14,15]. Fever, cough, myalgia and headache are commonly reported symptoms in COVID-19 patients.
COVID-19 respiratory symptoms are heterogeneous and may sometimes lead to serious complications. Similar to other severe respiratory diseases, severe forms of COVID-19 induce pneumonia, acute lung injury (ALI), ARDS and sepsis leading to multiple organ failure and death [16]. Studies have shown that the respiratory symptoms can worsen with development of ARDS occurring as fast as 9 days post onset [14]. Damage to the lungs characterized by a pulmonary ground glass opacification was observed by computed tomography (CT) scan in even asymptomatic cases indicating that the plethora of complications arising from COVID-19 is still far from being fully understood [17].
Cytokine storm is considered to be one of the major causes of ARDS and multiple-organ failure [18] and plays a crucial role in the process of disease aggravation [19]. The cytokine storm is the result of an exacerbated immune response resulting in the excessive production of pro-inflammatory cytokines. Whilst it is revealed that SARS-CoV-2 infection could alter both the innate and adaptive immunity [20,21], respiratory epithelial cells and myeloid cells are thought to play an important role in orchestrating innate immunity in the airway [22]. Infiltration of a large number of inflammatory immune cells is observed in the lungs from severe COVID-19 patients [23] with majority being macrophages and neutrophils [24]. Such increase in infiltration and accumulation of immune cells (macrophages, neutrophils) enhance the probability of rupture of atherosclerotic plaques potentially leading to cardiovascular complications.
Lung infiltration of macrophages has been reported in COVID-19 infection [25]. Pro-inflammatory cytokines such as IL-6 [26], IL-1 [27] and TNF [28] are thought to be produced by macrophages, reported to be hyper-induced during SARS-CoV-2 infection and are found to be positively correlated with disease severity relating to cytokine storms [29,30].
Neutrophils are the most abundant leukocytes in circulating blood which are responsible for the formation of the neutrophil extracellular trap (NET) [31]. Neutrophil elastase is a component of NET and is capable of the degradation of elastin within the pulmonary extracellular matrix which leads to the loss of elastic recoil of the lung and thus impairs normal lung function [32]. Interestingly, markers specific for NET formation are found to be elevated in COVID-19 patients and are up-regulated to a larger extent in patients who required mechanical ventilation when compared to patients with mild symptoms [33]. Moreover, elastin fragments are chemotactic to macrophages which are major drivers of the ongoing inflammation [32]. Furthermore, the macrophages secrete MMP proteins 8 and 9 which degrade COL1A1 resulting in formation of collagen-derived peptide Pro-Gly-Pro that can act as chemoattractant for neutrophils [32]. On the other hand, repairment of the ECM is driven by transforming growth factor (TGF)-β among other mediators [32]. Stored TGF-β in neutrophils could be activated by local elastase and contributes to the induction of pulmonary fibrosis through the differentiation of fibroblasts to myofibroblasts [34]. Therefore, by limiting viral infection through the generation of reactive oxygen species, by trapping the pathogen in the NET, and at the same time inducing pulmonary immunopathology and pulmonary fibrosis, neutrophils can act as double-edged sword in lung injuries.
3. Coagulopathy and COVID-19
A growing body of evidence suggests coagulopathy as a potential complication of COVID-19 resulting in higher risk of developing venous and arterial thromboembolism [7]. Indeed, patients with severe COVID-19 present with abnormal coagulation parameters which are associated with poor disease prognosis [35]. Likewise, COVID-19 patients present with higher than normal levels of fibrinogen [35], resulting from a high level of IL-6 in the serum. IL-6 is known to stimulate the production of fibrinogen by hepatocytes [36]. In addition, plasma levels of the procoagulation protein, von Willebrand factor is also increased in COVID-19 patients [5]. Levels of D-dimer and fibrin degradation product, which can reflect the occurrence of thrombosis and is associated with a diagnosis of disseminated intravascular coagulation (DIC), are found to be significantly enhanced in severe COVID-19 cases [35]. Although, the prevalence of DIC in COVID-19 is still in debate [4,35], pulmonary microthrombi formation is clearly observed in COVID-19 [37,38].
Pulmonary embolism, strokes and heart attacks can be a direct consequence of thrombosis. Indeed, pulmonary embolism is observed in 50% of COVID-19 patients admitted to ICU [39]. Adequate oxygenation and ventilation are recommended for COVID-19 patients with ARDS [40]; however, the development of pulmonary embolism may limit their usefulness by obstructing the circulation of oxygenated blood. Altogether, tackling thrombotic complications observed in COVID-19 patients need urgent investigation.
Autopsies of COVID-19 victims reveal a widespread presence of blood clots in infected lungs suggesting the involvement of pulmonary vascular endothelial cells in lung inflammation and coagulation [41]. Studies demonstrate that SARS-CoV-2 can infect endothelial cells, cells which represent one third of the total cells in lungs [42] and hence can contribute directly to thrombosis via endothelial cell lysis. Damage to the endothelial wall exposes the subendothelial collagen that is involved in platelet adhesion, activation and ultimately coagulation [43]. Secretion of factors involved in coagulation by the endothelial cells is also altered [44].
The idea of using anticoagulant therapy in COVID-19 patients to lower the mortality is well established [9]. In fact, the coagulation process is a balance between procoagulation and anticoagulation factors that require a strict control. Dysregulation towards either ends could lead to thrombophilia or coagulopathy. Protein C and protein S are among the key players in this process [45]. Interestingly, a low protein C activity is found in severe and aged COVID-19 patients favoring a hypercoagulability state [46].
Taken together, SARS-CoV-2 should not be regarded as an ordinary respiratory virus solely, but a virus which may possess a much broader tropism and could induce systemic symptoms and complications. Understanding the different disease mechanisms caused by infection will be vital in drug discovery for COVID-19 treatment.
4. Vitamin K
Vitamin K was first discovered by Henrik Dam in the early 1930s. This lipid soluble factor was first isolated for its “antihemorrhagic” properties [47,48,49]. Because of its requirement for hemostasis, Dam designated this factor as “Koagulations vitamin”, hence vitamin K. A second isoform, named K2, was isolated few years later by Edward Doisy from putrefied fish meals [50]. The importance of the discovery of vitamin K was highlighted when the prophylactic treatment of newborns who presented with vitamin K deficiency was shown to decrease significantly the neonatal mortality and thus was awarded a Nobel prize in Physiology or Medicine in 1943 [51]. The existence of vitamin K has been known for over 80 years mainly due to its involvement in coagulation. Subsequent discovery of different isoforms has suggested other potential functions of vitamin K beyond coagulation. Nowadays, vitamin K remains a fundamental bioactive compound used as supplement in optimizing body function. The following sections will first give an overview of vitamin K and its functions prior to addressing its potential benefits in prophylaxis and treatment of COVID-19.
5. Structure, Uptake and Distribution of Vitamin K
Naturally, vitamin K exists as two vitamers: K1 and K2. Structurally K1 belongs to the phylloquinone family whereas K2 chemical structure, which was elucidated in 1960, is related to the menaquinones [52]. Vitamin K1 represents the predominant form that can be found in daily diet [53,54] and is mainly present in green vegetables and fruits [55,56]. In mammalian cells in absence of bacteria, vitamin K1 was shown to be able to convert into vitamin K2 MK-4 isoform [57]. Normally K2 form is primarily bacterial in origin, and is produced in the human intestines. It can also be found in fermented food, meat and cheese [58]. Interestingly, the highest content of K2 is found in a Japanese dish named Natto. In terms of vitamin K intake, isoform K1 is known to be poorly absorbed when compared to larger side chained menaquinones [59]. Thus, it is predicted that as high as 70% of MK7,8,9 can be absorbed and distributed to extrahepatic tissue [60]. Overall, as much as 95% of extrahepatic vitamin K comes from dietary menaquinones, not phylloquinones. A healthy adult consumption of vitamin K should be around 1 µg/day/kg [61] and specifically 50 to 600 µg/day for vitamin K1 and 5 to 600 µg/day for vitamin K2 [58]. Although, low amounts in µg are sufficient already to maintain the daily body requirements due to an efficient vitamin K recycling system developed in mammals, studies have shown that majority of healthy adults are sub-clinically deficient for vitamin K in their circulation [62]. Contrarily to vitamin K1, which is rapidly removed from the circulation [63] and mainly remains in the liver [64], K2 form is known to be equally distributed between the circulation and the extra-hepatic tissues [65]. Thus, K2 is thought to provide a rapid and localized protective response whilst action of K1 is found to be more widespread. Commercially, there are two vitamin K2 forms available, named MK-4 and MK-7. MK-4 has a relatively short half-life of up to three hours, whilst MK-7 can remain stable for up to 3 days [60]. Vitamin K1 and MK-4 present similar properties whereas vitamin K2 larger isoforms (for example MK-7,8,9,10) are proposed to also possess function beyond coagulation [66]. Indeed, the presence of large side chains confer potential hydrophilic properties that are different from the K1 and MK-4 forms. Since vitamin K, even given at high doses has no reported side effects [67], its potential prophylactic benefits supplementation may be advisable.
6. Vitamin K: The Coagulation Switch and Beyond
The circulatory system is a complex network containing 60,000 miles of blood vessels through the body. Its success is intrinsically linked to two intertwine properties: circulation and coagulation of the blood. Circulation refers to the ability of blood to flow freely, and deliver the needed nutrients and oxygen to cells, whilst coagulation refers to the capacity in injury situation to stop the leak and repair, thus maintaining hemostasis. Under normal conditions, coagulation system is balanced towards the anticoagulation state [68]. Vitamin K is an essential “switch” in balancing coagulation and anticoagulation process [69]. Indeed, vitamin K acts as a cofactor in the activation of extra-hepatic and hepatic vitamin K-dependent proteins (VKDPs) including pro-thrombin and clotting factors VII, IX, X, major factors involved in blood coagulation. On the other hand, vitamin K can also trigger key anticoagulants via VKDPs for producing proteins C, S and Z [70,71,72]. In the presence of vitamin K, the glutamate (Glu) residues present on these proteins are carboxylated into gamma-carboxyglutamic acid (Gla) by γ-glutamyl carboxylase (GGCX) enzyme, enzyme that uses vitamin K as a cofactor for its activity [73]. Glu is modified into Gla on the coagulation factors of which these proteins display a higher affinity for calcium enabling them to form calcium bridges and bind to the surface membrane phospholipids prior to clot assembling [74,75,76]. It is important to note that vitamin K does not start the clotting process, it only enhances the coagulation system to work effectively. While vitamin K involvement in coagulation is well established, it is also a key component of the anticoagulation response. This response is facilitated through the activation of protein C, S and Z. Vitamin K-dependent protein C activation can inhibit clotting factors V and VIII which are responsible for clot generation [45,69].
Beyond its essential role in coagulation, vitamin K is suggested to possess immunomodulatory functions as well as preventing vascular calcification. Studies have shown that K2 form has more potent anti-inflammatory effect when compared to K1 [77]. K2 acts as an immunosuppressive compound to modulate expression of a multitude of pro-inflammatory cytokines such as TNF, IL-1α, IL-1β and suppresses IL-6 release [77,78,79]. It can also impair T cell activation and proliferation [80].
Besides, vitamin K has been shown to activate extra-hepatic VKDPs such as the Matrix Gla-protein (MGP), Osteocalcin and Gla-rich protein (GRP) [81,82,83]. MGP is mainly expressed in cartilage and vasculature and involved in ECM remodeling responsible for preventing vascular calcification [84] and thus plays a fundamental role in vascular health [85,86]. It has been suggested that vitamin K dependent MGP plays an important role in elastin degradation in the lungs phenomenon that is accelerated in pulmonary disease [87,88]. Furthermore, vascular calcification is often observed in chronic kidney disease patients [89], patients who have been reported to be more prone to develop severe form of COVID-19 highlighting the importance of vitamin K and MGP [90].
7. Using Vitamin K to Improve COVID-19 Outcomes
Very little is known concerning the potential benefits of using vitamin K to improve COVID-19 outcomes, however it is clearly established that patients with severe COVID-19, present with prevalent signs of coagulopathy and thromboembolism [4,7,39]. Impaired coagulation function has been demonstrated in COVID-19 patients [91]. Findings from several recent studies have further suggested that anticoagulant therapy is beneficial and can lower the mortality in COVID-19 patients [9,92]. Furthermore, patients with pre-conditions such as diabetes, hypertension and cardiovascular disease which are known to be associated with vitamin K deficiency [93,94,95] are prompt to develop a more severe COVID-19 disease [96]. This is particularly evident in patients suffering from chronic kidney disease (CKD), a population characterized by enhanced number of severe COVID-19 cases [90]. These patients suffer from subclinical vitamin K deficiency resulting from its high demand for the activation of VKDPs to inhibit calcification [97,98]. As a result, CKD patients are shown to present with high levels of non-phosphorylated non-carboxylated MGP, increasing the risk of vascular calcification and development of cardiovascular disease. Vitamin K supplementation of CKD patients was shown to reach target tissues including the vessel wall as well as improve the consequences resulting from vitamin K deficiency [97]. Furthermore, the progression of cardiovascular calcification in healthy adults was significantly reduced when supplemented with daily phylloquinone (0.5 mg) [99]. The CKD population serves as a valuable indicator when addressing potential consequences of poor vitamin K status, a status that represents an aggravating risk factor in COVID-19. Indeed, recently a direct association between low levels of vitamin K and severe cases of COVID-19 was reported [10]. Altogether, this evidence points to the existence of a possible link between vitamin K and COVID-19 as well as highlight the potential benefits of using vitamin K as a supplement.
8. Vitamin K: An Anticoagulation Option for COVID-19?
A serious hypercoagulable state has been observed in many severe COVID-19 cases and associated with poor prognostic outcome [17,35]. Contrarily to severe IAV cases, multiple blood clots are observed in the lungs at the site of SARS-CoV-2 infection [100]. As mentioned earlier, SARS-CoV-2 can directly infect endothelial cells which are known to express significant amount of receptor ACE2 [13]. Endothelial cells play a direct role in coagulation. Indeed, they secrete coagulation inhibitors like protein S as well as provide receptors for anticoagulant proteins present in the blood that interfere with clot formation (like protein C) [45]. Thus, the imbalance of coagulation system by altering/lysing of endothelial cells after infection can significantly contribute to thrombosis. On 25th March 2020, the International Society of Thrombosis and Hemostasis (ISTH) introduced provisional guidelines for the management of coagulopathy in COVID-19 patients. There are numerous anticoagulants with various mode of actions that are in use clinically.
A low prophylactic dose of low molecular heparin (LMWH), an anticoagulant, was suggested to be given to all COVID-19 patients requiring hospitalization as long as no contraindications such as active bleeding was recorded [101]. While studies have shown beneficial effect of LMWH on COVID-19 patients in terms of reduce mortality [9], in clinical practice severely infected patients still continue to clot and fail to response adequately to both prophylactic and therapeutic doses [102,103]. This might be resulting from the fact that COVID-19 patients present with low levels of anti-thrombin and higher levels of fibrinogen, which contribute to heparin resistance [102]. Indeed, hyperfibrinogenemia was clearly demonstrated in patients with severe COVID-19 and was shown to reduce significantly LMWH efficacy to reduce clot formation [104]. Furthermore, due to the risk of venous thromboembolism, pulmonary embolism and renal insufficiency resulting from SARS-CoV-2 [102], the use of unfractioned heparin (UFH) might be a better choice of anticoagulant [105]. Indeed, patients who present with pulmonary embolism and receive LMWH are at an increased risk of bleeding that cannot be stopped further supporting the use of UFH.
Direct oral anticoagulant (DOAC) drugs are currently broadly administered as anticoagulant treatments. This novel class of anticoagulant act directly on selective blood clotting factors to prevent formation of blood clots. However, their use in COVID-19 patients remain controversial. Indeed, up to now there is very limited clinical data on safety or efficacy of DOAC in COVID-19 patients [106]. Meanwhile, ample evidence suggests a direct impact on the cytochrome P450 pathway which is observed in both antiviral treatment (remdesivir, dexamethasone), as well as COVID-19 disease. DOACs are also known to alter the same P450 pathway [107]. Thus, combined antiviral and anticoagulant treatment using DOAC might cause drug-drug interactions resulting in potential decrease or increase in anticoagulation activity. A recent study in Italy on COVID-19 patients where DOAC treatment was simultaneously administered with antiviral drugs showed that all patients presented with alarming increase of DOAC at plasma levels [108]. Altogether, it seems prudent not to start DOAC treatment in COVID-19 treated patients until more evidence is established.
Although progressively substituted by DOAC, Vitamin K antagonists (VKAs), such as Warfarin for example, still remain important anticoagulant drugs. VKAs interrupt the vitamin K cycle through inhibition of vitamin K 2,3-epoxide reductase leading to deficiency in vitamin K. As previously mentioned, vitamin K has also anti-coagulant properties through activation of protein C, S and Z. In contrast to proteins C and Z which are mainly localized in the liver, half of protein S is synthesized in endothelial cells playing thus a fundamental role in local prevention of thrombosis [109,110,111]. Interestingly it has been shown that uptake of supplements of vitamin K1 does not alter VKA anticoagulant efficacy [112]. However, potential interference might be observed with high amounts of vitamin K2 (MK-7, MK-8, MK-9 and larger isoforms) indicating that people undertaking VKA should avoid consuming food or supplements with high amounts of K2 analogues. Furthermore, a beneficial decrease in both level of inactivated factor II and osteocalcin was observed in patients who increase their dietary intake of vitamin K when undergoing VKA treatment [112]. Altogether, combining VKAs with an increase of vitamin K1 uptake should be considered in COVID-19 patients.
Up to date WHO has approved three vaccines: Pfizer/BioNTech, Moderna and Astra-Zeneca. Previous studies to determine whether influenza vaccination interferes with anticoagulant therapy are controversial and still under debate [113,114]. However, most recent studies seem to point to a lack of significant effect of vaccine on anticoagulant therapy [115]. Thus, current global recommendations encourage patients on anticoagulant therapy to receive a vaccine to protect against COVID-19 and monitor their blood level thinning following vaccine when taking Warfarin for example.
Altogether a more aggressive anticoagulation approach should be undertaken when treating patients with severe form of COVID-19. As mentioned earlier, given that vitamin K is a key component in blood clotting, a combined administration of UFH, anti-thrombin supplement and vitamin K should be explored as such remedy can both promote anticoagulation (vitamin K) and block formation of blood clots (UFH & anti-thrombin).
9. Vitamin K: An Immunomodulatory Option for COVID-19?
Beyond its potential beneficial effect to prevent coagulopathy, vitamin K is also known to play an important role in immunomodulation. Indeed, in vitro studies have shown that vitamin K is associated with an impaired production of proinflammatory cytokines [77,116]. Inhibition of TNF, IL-1 and IL-6 by vitamin K has been showed [77,78,79]. Interestingly, these are among the most important cytokines activated during SARS-CoV-2 infection [29,117], which contribute to cytokine storm leading to ARDS in severe COVID-19 patients [118]. Furthermore, the loss of the alveolar-capillary membrane integrity is a hallmark of ARDS [23]. Proteins C and S, which are activated by vitamin K, are known to play protective role and maintain the integrity of this membrane [119,120]. Thus, vitamin K administration may help in attenuating cytokines levels as well as protect the integrity of alveolar-capillary membrane, thus reducing the risk for ARDS development in COVID-19 patients.
10. Vitamin K: Vascular Health Promoter to Protect against Cardiovascular Complications and Lung Fibrosis?
Finally, 40% of deaths from SARS-CoV-2 infection are related to cardiovascular complications [121]. Interestingly, through activation of MGP, vitamin K can prevent development of arterial calcification [122,123,124], a process known to cause cardiovascular disease [65] as well as maintain arterial elasticity [125]. Vitamin K dependent MGP protects elastic fibers against mineralization, fibers which are fundamental parts of the extracellular matrix (ECM) and play a crucial role in lung fibrosis [126]. Low vitamin K status is found to be associated with increased elastin degradation in pulmonary disease [127]. Furthermore, MGP is known to be highly expressed in the lung, and modulation to MGP and inability to activate (carboxylate resulting from insufficient levels of vitamin K) is suggested to be a contributor to lung fibrosis. Thus, vitamin K is considered to promote vascular health and reduce the risk for development of lung fibrosis in COVID-19.
11. Concluding Remarks
SARS-CoV-2 emerges as an uncommon disease with a plethora of signs and symptoms previously unseen in respiratory infections. It is initially considered as a respiratory illness prior to also becoming a vascular condition.
A possible link to low levels of vitamin K and severe cases of COVID-19 was lately reported [6]. The diverse and distinct roles of vitamin K in modulating blood clotting, elastin degradation, immunomodulation, and managing vascular health, together with the low toxicity of vitamin K in humans makes vitamin K an attractive remedy using prophylactically as supplement or therapeutically to improve COVID-19 disease outcomes. A summary describing the potential involvement of coagulopathy in the pathogenesis of SARS-CoV-2 viral infection is illustrated in Figure 1. More research is needed immediately to further investigate its potentials.
Author Contributions
S.M.Y.L. conceptualized the work. M.K., T.F.Y., G.C.H.L. and S.M.Y.L. wrote the manuscript, and approved the manuscript for its intellectual content. All authors have read and agreed to the published version of the manuscript.
Funding
S.M.Y.L. is supported by Research Grants Council (RGC) of Hong Kong, General Research Fund (GRF) [grant number 17116018].
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
- World Health Organization. Weekly Operational Update on COVID-19; World Health Organization: Geneva, Switzerland, 2021. [Google Scholar]
- Coperchini, F.; Chiovato, L.; Croce, L.; Magri, F.; Rotondi, M. The cytokine storm in COVID-19: An overview of the involvement of the chemokine/chemokine-receptor system. Cytokine Growth Factor Rev. 2020, 53, 25–32. [Google Scholar] [CrossRef]
- Song, P.; Li, W.; Xie, J.; Hou, Y.; You, C. Cytokine storm induced by SARS-CoV-2. Clin. Chim. Acta 2020, 509, 280–287. [Google Scholar] [CrossRef] [PubMed]
- Connors, J.M.; Levy, J.H. COVID-19 and its implications for thrombosis and anticoagulation. Blood 2020, 135, 2033–2040. [Google Scholar] [CrossRef]
- Panigada, M.; Bottino, N.; Tagliabue, P.; Grasselli, G.; Novembrino, C.; Chantarangkul, V.; Pesenti, A.; Peyvandi, F.; Tripodi, A. Hypercoagulability of COVID-19 patients in intensive care unit: A report of thromboelastography findings and other parameters of hemostasis. J. Thromb. Haemost. 2020, 18, 1738–1742. [Google Scholar] [CrossRef]
- Klok, F.A.; Kruip, M.; van der Meer, N.J.M.; Arbous, M.S.; Gommers, D.; Kant, K.M.; Kaptein, F.H.J.; van Paassen, J.; Stals, M.A.M.; Huisman, M.V.; et al. Confirmation of the high cumulative incidence of thrombotic complications in critically ill ICU patients with COVID-19: An updated analysis. Thromb. Res. 2020, 191, 148–150. [Google Scholar] [CrossRef] [PubMed]
- Lodigiani, C.; Iapichino, G.; Carenzo, L.; Cecconi, M.; Ferrazzi, P.; Sebastian, T.; Kucher, N.; Studt, J.D.; Sacco, C.; Alexia, B.; et al. Venous and arterial thromboembolic complications in COVID-19 patients admitted to an academic hospital in Milan, Italy. Thromb. Res. 2020, 191, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Middeldorp, S.; Coppens, M.; van Haaps, T.F.; Foppen, M.; Vlaar, A.P.; Müller, M.C.A.; Bouman, C.C.S.; Beenen, L.F.M.; Kootte, R.S.; Heijmans, J.; et al. Incidence of venous thromboembolism in hospitalized patients with COVID-19. J. Thromb. Haemost. 2020. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; Bai, H.; Chen, X.; Gong, J.; Li, D.; Sun, Z. Anticoagulant treatment is associated with decreased mortality in severe coronavirus disease 2019 patients with coagulopathy. J. Thromb. Haemost. 2020, 18, 1094–1099. [Google Scholar] [CrossRef]
- Dofferhoff, A.S.M.; Piscaer, I.; Schurgers, L.J.; Visser, M.P.J.; van den Ouweland, J.M.W.; de Jong, P.A.; Gosens, R.; Hackeng, T.M.; van Daal, H.; Lux, P.; et al. Reduced vitamin K status as a potentially modifiable risk factor of severe COVID-19. Clin. Infect. Dis. 2020. [Google Scholar] [CrossRef]
- Letko, M.; Marzi, A.; Munster, V. Functional assessment of cell entry and receptor usage for SARS-CoV-2 and other lineage B betacoronaviruses. Nat. Microbiol. 2020, 5, 562–569. [Google Scholar] [CrossRef] [Green Version]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell 2020, 181, 281–292. [Google Scholar] [CrossRef]
- Varga, Z.; Flammer, A.J.; Steiger, P.; Haberecker, M.; Andermatt, R.; Zinkernagel, A.S.; Mehra, M.R.; Schuepbach, R.A.; Ruschitzka, F.; Moch, H. Endothelial cell infection and endotheliitis in COVID-19. Lancet 2020, 395, 1417–1418. [Google Scholar] [CrossRef]
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical features of patients infected with 2019 novel coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- She, J.; Jiang, J.; Ye, L.; Hu, L.; Bai, C.; Song, Y. 2019 novel coronavirus of pneumonia in Wuhan, China: Emerging attack and management strategies. Clin. Transl. Med. 2020, 9, 19. [Google Scholar] [CrossRef] [PubMed]
- Zaim, S.; Chong, J.H.; Sankaranarayanan, V.; Harky, A. COVID-19 and Multiorgan Response. Curr. Probl. Cardiol. 2020, 45, 100618. [Google Scholar] [CrossRef] [PubMed]
- Guan, W.J.; Ni, Z.Y.; Hu, Y.; Liang, W.H.; Ou, C.Q.; He, J.X.; Liu, L.; Shan, H.; Lei, C.L.; Hui, D.S.C.; et al. Clinical Characteristics of Coronavirus Disease 2019 in China. N. Engl. J. Med. 2020, 382, 1708–1720. [Google Scholar] [CrossRef] [PubMed]
- Chousterman, B.G.; Swirski, F.K.; Weber, G.F. Cytokine storm and sepsis disease pathogenesis. Semin. Immunopathol. 2017, 39, 517–528. [Google Scholar] [CrossRef] [PubMed]
- Shimabukuro-Vornhagen, A.; Gödel, P.; Subklewe, M.; Stemmler, H.J.; Schlößer, H.A.; Schlaak, M.; Kochanek, M.; Böll, B.; von Bergwelt-Baildon, M.S. Cytokine release syndrome. J. Immunother. Cancer 2018, 6, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giamarellos-Bourboulis, E.J.; Netea, M.G.; Rovina, N.; Akinosoglou, K.; Antoniadou, A.; Antonakos, N.; Damoraki, G.; Gkavogianni, T.; Adami, M.E.; Katsaounou, P.; et al. Complex Immune Dysregulation in COVID-19 Patients with Severe Respiratory Failure. Cell Host Microbe 2020, 27, 992–1000. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Nie, J.; Wang, H.; Zhao, Q.; Xiong, Y.; Deng, L.; Song, S.; Ma, Z.; Mo, P.; Zhang, Y. Characteristics of peripheral lymphocyte subset alteration in COVID-19 pneumonia. J. Infect. Dis. 2020, 221, 1762–1769. [Google Scholar] [CrossRef] [Green Version]
- Yoshikawa, T.; Hill, T.; Li, K.; Peters, C.J.; Tseng, C.T. Severe acute respiratory syndrome (SARS) coronavirus-induced lung epithelial cytokines exacerbate SARS pathogenesis by modulating intrinsic functions of monocyte-derived macrophages and dendritic cells. J. Virol. 2009, 83, 3039–3048. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Shi, L.; Wang, Y.; Zhang, J.; Huang, L.; Zhang, C.; Liu, S.; Zhao, P.; Liu, H.; Zhu, L.; et al. Pathological findings of COVID-19 associated with acute respiratory distress syndrome. Lancet Respir. Med. 2020, 8, 420–422. [Google Scholar] [CrossRef]
- Barnes, B.J.; Adrover, J.M.; Baxter-Stoltzfus, A.; Borczuk, A.; Cools-Lartigue, J.; Crawford, J.M.; Dassler-Plenker, J.; Guerci, P.; Huynh, C.; Knight, J.S.; et al. Targeting potential drivers of COVID-19: Neutrophil extracellular traps. J. Exp. Med. 2020, 217, e20200652. [Google Scholar] [CrossRef]
- Tian, S.; Hu, W.; Niu, L.; Liu, H.; Xu, H.; Xiao, S.Y. Pulmonary Pathology of Early-Phase 2019 Novel Coronavirus (COVID-19) Pneumonia in Two Patients with Lung Cancer. J. Thorac. Oncol. 2020, 15, 700–704. [Google Scholar] [CrossRef]
- Yan, Y.; Yang, Y.; Wang, F.; Ren, H.; Zhang, S.; Shi, X.; Yu, X.; Dong, K. Clinical characteristics and outcomes of patients with severe covid-19 with diabetes. BMJ Open Diabetes Res. Care 2020, 8, e001343. [Google Scholar] [CrossRef] [PubMed]
- Conti, P.; Ronconi, G.; Caraffa, A.; Gallenga, C.E.; Ross, R.; Frydas, I.; Kritas, S.K. Induction of pro-inflammatory cytokines (IL-1 and IL-6) and lung inflammation by Coronavirus-19 (COVI-19 or SARS-CoV-2): Anti-inflammatory strategies. J. Biol. Regul. Homeost. Agents 2020, 34, 1. [Google Scholar] [CrossRef]
- Li, X.; Xu, S.; Yu, M.; Wang, K.; Tao, Y.; Zhou, Y.; Shi, J.; Zhou, M.; Wu, B.; Yang, Z.; et al. Risk factors for severity and mortality in adult COVID-19 inpatients in Wuhan. J. Allergy Clin. Immunol. 2020, 146, 110–118. [Google Scholar] [CrossRef]
- Wang, J.; Jiang, M.; Chen, X.; Montaner, L.J. Cytokine storm and leukocyte changes in mild versus severe SARS-CoV-2 infection: Review of 3939 COVID-19 patients in China and emerging pathogenesis and therapy concepts. J. Leukoc. Biol. 2020, 108, 17–41. [Google Scholar] [CrossRef]
- Chen, X.; Zhao, B.; Qu, Y.; Chen, Y.; Xiong, J.; Feng, Y.; Men, D.; Huang, Q.; Liu, Y.; Yang, B. Detectable serum SARS-CoV-2 viral load (RNAaemia) is closely correlated with drastically elevated interleukin 6 (IL-6) level in critically ill COVID-19 patients. Clin. Infect. Dis. 2020, 71, 1937–1942. [Google Scholar] [CrossRef]
- Kolaczkowska, E.; Kubes, P. Neutrophil recruitment and function in health and inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, T.; O’Reilly, P.; Antony, V.B.; Gaggar, A.; Thannickal, V.J. Matrix Remodeling in Pulmonary Fibrosis and Emphysema. Am. J. Respir. Cell Mol. Biol. 2016, 54, 751–760. [Google Scholar] [CrossRef] [Green Version]
- Zuo, Y.; Yalavarthi, S.; Shi, H.; Gockman, K.; Zuo, M.; Madison, J.A.; Blair, C.N.; Weber, A.; Barnes, B.J.; Egeblad, M.; et al. Neutrophil extracellular traps in COVID-19. JCI Insight 2020, 5, e138999. [Google Scholar] [CrossRef] [Green Version]
- Chen, W. A potential treatment of COVID-19 with TGF-β blockade. Int. J. Biol. Sci. 2020, 16, 1954–1955. [Google Scholar] [CrossRef] [PubMed]
- Tang, N.; Li, D.; Wang, X.; Sun, Z. Abnormal coagulation parameters are associated with poor prognosis in patients with novel coronavirus pneumonia. J. Thromb. Haemost. 2020, 18, 844–847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt-Arras, D.; Rose-John, S. IL-6 pathway in the liver: From physiopathology to therapy. J. Hepatol. 2016, 64, 1403–1415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atallah, B.; Mallah, S.I.; AlMahmeed, W. Anticoagulation in COVID-19. Eur. Heart J. Cardiovasc. Pharmacother. 2020, 6, 260–261. [Google Scholar] [CrossRef]
- McGonagle, D.; O’Donnell, J.S.; Sharif, K.; Emery, P.; Bridgewood, C. Immune mechanisms of pulmonary intravascular coagulopathy in COVID-19 pneumonia. Lancet Rheumatol. 2020, 2, e437–e445. [Google Scholar] [CrossRef]
- Bompard, F.; Monnier, H.; Saab, I.; Tordjman, M.; Abdoul, H.; Fournier, L.; Sanchez, O.; Lorut, C.; Chassagnon, G.; Revel, M.P. Pulmonary embolism in patients with Covid-19 pneumonia. Eur. Respir. J. 2020. [Google Scholar] [CrossRef]
- Liu, X.; Liu, X.; Xu, Y.; Xu, Z.; Huang, Y.; Chen, S.; Li, S.; Liu, D.; Lin, Z.; Li, Y. Ventilatory Ratio in Hypercapnic Mechanically Ventilated Patients with COVID-19–associated Acute Respiratory Distress Syndrome. Am. J. Respir. Crit. Care Med. 2020, 201, 1297–1299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wichmann, D.; Sperhake, J.P.; Lütgehetmann, M.; Steurer, S.; Edler, C.; Heinemann, A.; Heinrich, F.; Mushumba, H.; Kniep, I.; Schröder, A.S.; et al. Autopsy Findings and Venous Thromboembolism in Patients with COVID-19. Ann. Intern. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Zeng, H.; Pappas, C.; Belser, J.A.; Houser, K.V.; Zhong, W.; Wadford, D.A.; Stevens, T.; Balczon, R.; Katz, J.M.; Tumpey, T.M. Human pulmonary microvascular endothelial cells support productive replication of highly pathogenic avian influenza viruses: Possible involvement in the pathogenesis of human H5N1 virus infection. J. Virol. 2012, 86, 667–678. [Google Scholar] [CrossRef] [Green Version]
- Farndale, R.W.; Sixma, J.J.; Barnes, M.J.; de Groot, P.G. The role of collagen in thrombosis and hemostasis. J. Thromb. Haemost. 2004, 2, 561–573. [Google Scholar] [CrossRef]
- Frantzeskaki, F.; Armaganidis, A.; Orfanos, S.E. Immunothrombosis in Acute Respiratory Distress Syndrome: Cross Talks between Inflammation and Coagulation. Respiration 2017, 93, 212–225. [Google Scholar] [CrossRef]
- Esmon, C.T.; Vigano-D’Angelo, S.; D’Angelo, A.; Comp, P.C. Anticoagulation proteins C and S. Adv. Exp. Med. Biol. 1987, 214, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Tabatabai, A.; Rabin, J.; Menaker, J.; Madathil, R.; Galvagno, S.; Menne, A.; Chow, J.H.; Grazioli, A.; Herr, D.; Tanaka, K.; et al. Factor VIII and Functional Protein C Activity in Critically Ill Patients with Coronavirus Disease 2019: A Case Series. A A Pract. 2020, 14, e01236. [Google Scholar] [CrossRef] [PubMed]
- Dam, H. The antihaemorrhagic vitamin of the chick. Biochem. J. 1935, 29, 1273–1285. [Google Scholar] [CrossRef] [Green Version]
- Dam, H.; Schönheyder, F. A deficiency disease in chicks resembling scurvy. Biochem. J. 1934, 28, 1355–1359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McFarlane, W.D.; Graham, W.R., Jr.; Richardson, F. The fat-soluble vitamin requirements of the chick: The vitamin A and vitamin D content of fish meal and meat meal. Biochem. J. 1931, 25, 358–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKee, R.W.; Binkley, S.B.; MacCorquodale, D.W.; Thayer, S.A.; Doisy, E.A. The Isolation of Vitamins K1 And K2. J. Am. Chem. Soc. 1939, 61, 1295. [Google Scholar] [CrossRef]
- Zetterström, R.H.C.P. Dam (1895–1976) and E. A. Doisy (1893–1986): The discovery of antihaemorrhagic vitamin and its impact on neonatal health. Acta Paediatr. 2006, 95, 642–644. [Google Scholar] [CrossRef]
- Jacobsen, B.K.; Dam, H. Vitamin K in bacteria. Biochim. Biophys. Acta 1960, 40, 211–216. [Google Scholar] [CrossRef]
- Gijsbers, B.L.; Jie, K.-S.G.; Vermeer, C. Effect of food composition on vitamin K absorption in human volunteers. Br. J. Nutr. 1996, 76, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Schurgers, L.J.; Vermeer, C. Determination of phylloquinone and menaquinones in food. Effect of food matrix on circulating vitamin K concentrations. Haemostasis 2000, 30, 298–307. [Google Scholar] [CrossRef]
- Dismore, M.L.; Haytowitz, D.B.; Gebhardt, S.E.; Peterson, J.W.; Booth, S.L. Vitamin K content of nuts and fruits in the US diet. J. Am. Diet. Assoc. 2003, 103, 1650–1652. [Google Scholar] [CrossRef]
- Tarento, T.D.; McClure, D.D.; Talbot, A.M.; Regtop, H.L.; Biffin, J.R.; Valtchev, P.; Dehghani, F.; Kavanagh, J.M. A potential biotechnological process for the sustainable production of vitamin K1. Crit. Rev. Biotechnol. 2019, 39, 1–19. [Google Scholar] [CrossRef]
- Davidson, R.T.; Foley, A.L.; Engelke, J.A.; Suttie, J.W. Conversion of dietary phylloquinone to tissue menaquinone-4 in rats is not dependent on gut bacteria. J. Nutr. 1998, 128, 220–223. [Google Scholar] [CrossRef] [Green Version]
- Marles, R.J.; Roe, A.L.; Oketch-Rabah, H.A. US Pharmacopeial Convention safety evaluation of menaquinone-7, a form of vitamin K. Nutr. Rev. 2017, 75, 553–578. [Google Scholar] [CrossRef] [PubMed]
- Akbulut, A.C.; Pavlic, A.; Petsophonsakul, P.; Halder, M.; Maresz, K.; Kramann, R.; Schurgers, L. Vitamin K2 needs an RDI separate from vitamin K1. Nutrients 2020, 12, 1852. [Google Scholar] [CrossRef] [PubMed]
- Schurgers, L.J.; Teunissen, K.J.; Hamulyák, K.; Knapen, M.H.; Vik, H.; Vermeer, C. Vitamin K-containing dietary supplements: Comparison of synthetic vitamin K1 and natto-derived menaquinone-7. Blood 2007, 109, 3279–3283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Vitamin and Mineral Requirements in Human Nutrition; World Health Organization: Geneva, Switzerland, 2004. [Google Scholar]
- Binkley, N.C.; Krueger, D.C.; Engelke, J.A.; Foley, A.L.; Suttie, J.W. Vitamin K supplementation reduces serum concentrations of under-gamma-carboxylated osteocalcin in healthy young and elderly adults. Am. J. Clin. Nutr. 2000, 72, 1523–1528. [Google Scholar] [CrossRef] [Green Version]
- Shearer, M.J.; Mallinson, C.N.; Webster, G.R.; Barkhan, P. Clearance from plasma and excretion in urine, faeces and bile of an intravenous dose of tritiated vitamin K 1 in man. Br. J. Haematol. 1972, 22, 579–588. [Google Scholar] [CrossRef]
- Schurgers, L.J.; Vermeer, C. Differential lipoprotein transport pathways of K-vitamins in healthy subjects. Biochim. Biophys. Acta 2002, 1570, 27–32. [Google Scholar] [CrossRef]
- Spronk, H.M.; Soute, B.A.; Schurgers, L.J.; Thijssen, H.H.; De Mey, J.G.; Vermeer, C. Tissue-specific utilization of menaquinone-4 results in the prevention of arterial calcification in warfarin-treated rats. J. Vasc. Res. 2003, 40, 531–537. [Google Scholar] [CrossRef]
- Halder, M.; Petsophonsakul, P.; Akbulut, A.C.; Pavlic, A.; Bohan, F.; Anderson, E.; Maresz, K.; Kramann, R.; Schurgers, L. Vitamin K: Double bonds beyond coagulation insights into differences between vitamin K1 and K2 in health and disease. Int. J. Mol. Sci. 2019, 20, 896. [Google Scholar] [CrossRef] [Green Version]
- Kaneki, M.; Hedges, S.J.; Hosoi, T.; Fujiwara, S.; Lyons, A.; Ishida, N.; Nakagawa, M.; Takechi, M.; Sano, Y.; Mizuno, Y. Japanese fermented soybean food as the major determinant of the large geographic difference in circulating levels of vitamin K2: Possible implications for hip-fracture risk. Nutrition 2001, 17, 315–321. [Google Scholar] [CrossRef]
- Dahlbäck, B. Blood coagulation. Lancet 2000, 355, 1627–1632. [Google Scholar] [CrossRef]
- Espana, F.; Medina, P.; Navarro, S.; Zorio, E.; Estellés, A.; Aznar, J. The multifunctional protein C system. Curr. Med. Chem. Cardiovasc. Hematol. Agents 2005, 3, 119–131. [Google Scholar] [CrossRef]
- Olson, R.E. The function and metabolism of vitamin K. Annu. Rev. Nutr. 1984, 4, 281–337. [Google Scholar] [CrossRef] [PubMed]
- Suttie, J.W. Vitamin K-dependent carboxylase. Annu. Rev. Biochem. 1985, 54, 459–477. [Google Scholar] [CrossRef] [PubMed]
- Danziger, J. Vitamin K-dependent proteins, warfarin, and vascular calcification. Clin. J. Am. Soc. Nephrol. 2008, 3, 1504–1510. [Google Scholar] [CrossRef]
- Nelsestuen, G.L.; Zytkovicz, T.H.; Howard, J.B. The mode of action of vitamin K. Identification of gamma-carboxyglutamic acid as a component of prothrombin. J. Biol. Chem. 1974, 249, 6347–6350. [Google Scholar] [CrossRef]
- Schurgers, L.J.; Spronk, H.M. Differential cellular effects of old and new oral anticoagulants: Consequences to the genesis and progression of atherosclerosis. Thromb. Haemost. 2014, 112, 909–917. [Google Scholar] [CrossRef] [PubMed]
- Nelsestuen, G.L.; Suttie, J.W. Mode of action of vitamin K. Calcium binding properties of bovine prothrombin. Biochemistry 1972, 11, 4961–4964. [Google Scholar] [CrossRef]
- Ellison, E.H.; Castellino, F.J. Adsorption of vitamin K-dependent blood coagulation proteins to spread phospholipid monolayers as determined from combined measurements of the surface pressure and surface protein concentration. Biochemistry 1998, 37, 7997–8003. [Google Scholar] [CrossRef]
- Reddi, K.; Henderson, B.; Meghji, S.; Wilson, M.; Poole, S.; Hopper, C.; Harris, M.; Hodges, S.J. Interleukin 6 production by lipopolysaccharide-stimulated human fibroblasts is potently inhibited by naphthoquinone (vitamin K) compounds. Cytokine 1995, 7, 287–290. [Google Scholar] [CrossRef]
- Pan, M.H.; Maresz, K.; Lee, P.S.; Wu, J.C.; Ho, C.T.; Popko, J.; Mehta, D.S.; Stohs, S.J.; Badmaev, V. Inhibition of TNF-α, IL-1α, and IL-1β by Pretreatment of Human Monocyte-Derived Macrophages with Menaquinone-7 and Cell Activation with TLR Agonists In Vitro. J. Med. Food 2016, 19, 663–669. [Google Scholar] [CrossRef]
- Ohsaki, Y.; Shirakawa, H.; Miura, A.; Giriwono, P.E.; Sato, S.; Ohashi, A.; Iribe, M.; Goto, T.; Komai, M. Vitamin K suppresses the lipopolysaccharide-induced expression of inflammatory cytokines in cultured macrophage-like cells via the inhibition of the activation of nuclear factor κB through the repression of IKKα/β phosphorylation. J. Nutr. Biochem. 2010, 21, 1120–1126. [Google Scholar] [CrossRef]
- Myneni, V.D.; Mezey, E. Immunomodulatory effect of vitamin K2: Implications for bone health. Oral Dis. 2018, 24, 67–71. [Google Scholar] [CrossRef] [PubMed]
- Price, P.A.; Urist, M.R.; Otawara, Y. Matrix Gla protein, a new gamma-carboxyglutamic acid-containing protein which is associated with the organic matrix of bone. Biochem. Biophys. Res. Commun. 1983, 117, 765–771. [Google Scholar] [CrossRef]
- Hauschka, P.V.; Lian, J.B.; Gallop, P.M. Direct identification of the calcium-binding amino acid, gamma-carboxyglutamate, in mineralized tissue. Proc. Natl. Acad. Sci. USA 1975, 72, 3925–3929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willems, B.A.; Vermeer, C.; Reutelingsperger, C.P.; Schurgers, L.J. The realm of vitamin K dependent proteins: Shifting from coagulation toward calcification. Mol. Nutr. Food Res. 2014, 58, 1620–1635. [Google Scholar] [CrossRef]
- Schurgers, L.J.; Uitto, J.; Reutelingsperger, C.P. Vitamin K-dependent carboxylation of matrix Gla-protein: A crucial switch to control ectopic mineralization. Trends Mol. Med. 2013, 19, 217–226. [Google Scholar] [CrossRef]
- Luo, G.; Ducy, P.; McKee, M.D.; Pinero, G.J.; Loyer, E.; Behringer, R.R.; Karsenty, G. Spontaneous calcification of arteries and cartilage in mice lacking matrix GLA protein. Nature 1997, 386, 78–81. [Google Scholar] [CrossRef] [PubMed]
- Munroe, P.B.; Olgunturk, R.O.; Fryns, J.-P.; Van Maldergem, L.; Ziereisen, F.; Yuksel, B.; Gardiner, R.M.; Chung, E. Mutations in the gene encoding the human matrix Gla protein cause Keutel syndrome. Nat. Genet. 1999, 21, 142–144. [Google Scholar] [CrossRef] [PubMed]
- Piscaer, I.; Wouters, E.F.M.; Vermeer, C.; Janssens, W.; Franssen, F.M.E.; Janssen, R. Vitamin K deficiency: The linking pin between COPD and cardiovascular diseases? Respir. Res. 2017, 18, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssen, R.; Vermeer, C. Vitamin K deficit and elastolysis theory in pulmonary elasto-degenerative diseases. Med Hypotheses 2017, 108, 38–41. [Google Scholar] [CrossRef]
- Jono, S.; Shioi, A.; Ikari, Y.; Nishizawa, Y. Vascular calcification in chronic kidney disease. J. Bone Miner. Metab. 2006, 24, 176–181. [Google Scholar] [CrossRef]
- ERA-EDTA Council, ERACODA Working Group. Chronic kidney disease is a key risk factor for severe COVID-19: A call to action by the ERA-EDTA. Nephrol. Dial. Transplant. 2021, 36, 87–94. [Google Scholar] [CrossRef]
- Han, H.; Yang, L.; Liu, R.; Liu, F.; Wu, K.L.; Li, J.; Liu, X.H.; Zhu, C.L. Prominent changes in blood coagulation of patients with SARS-CoV-2 infection. Clin. Chem. Lab. Med. 2020, 58, 1116–1120. [Google Scholar] [CrossRef] [Green Version]
- Kollias, A.; Kyriakoulis, K.G.; Dimakakos, E.; Poulakou, G.; Stergiou, G.S.; Syrigos, K. Thromboembolic risk and anticoagulant therapy in COVID-19 patients: Emerging evidence and call for action. Br. J. Haematol. 2020, 189, 846–847. [Google Scholar] [CrossRef]
- Manna, P.; Kalita, J. Beneficial role of vitamin K supplementation on insulin sensitivity, glucose metabolism, and the reduced risk of type 2 diabetes: A review. Nutrition 2016, 32, 732–739. [Google Scholar] [CrossRef]
- Campbell, A.W. Vitamin K2 in the Prevention of Cardiovascular Diseases and Diabetes. Altern. Ther. Health Med. 2017, 23, 8–10. [Google Scholar]
- Van Ballegooijen, A.J.; Cepelis, A.; Visser, M.; Brouwer, I.A.; van Schoor, N.M.; Beulens, J.W. Joint Association of Low Vitamin D and Vitamin K Status with Blood Pressure and Hypertension. Hypertension 2017, 69, 1165–1172. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X.; et al. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]
- Cozzolino, M.; Mangano, M.; Galassi, A.; Ciceri, P.; Messa, P.; Nigwekar, S. Vitamin K in chronic kidney disease. Nutrients 2019, 11, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cranenburg, E.C.; Schurgers, L.J.; Uiterwijk, H.H.; Beulens, J.W.; Dalmeijer, G.W.; Westerhuis, R.; Magdeleyns, E.J.; Herfs, M.; Vermeer, C.; Laverman, G.D. Vitamin K intake and status are low in hemodialysis patients. Kidney Int. 2012, 82, 605–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shea, M.K.; O’Donnell, C.J.; Hoffmann, U.; Dallal, G.E.; Dawson-Hughes, B.; Ordovas, J.M.; Price, P.A.; Williamson, M.K.; Booth, S.L. Vitamin K supplementation and progression of coronary artery calcium in older men and women. Am. J. Clin. Nutr. 2009, 89, 1799–1807. [Google Scholar] [CrossRef]
- Ackermann, M.; Verleden, S.E.; Kuehnel, M.; Haverich, A.; Welte, T.; Laenger, F.; Vanstapel, A.; Werlein, C.; Stark, H.; Tzankov, A.; et al. Pulmonary Vascular Endothelialitis, Thrombosis, and Angiogenesis in Covid-19. N. Engl. J. Med. 2020, 383, 120–128. [Google Scholar] [CrossRef]
- Thachil, J.; Tang, N.; Gando, S.; Falanga, A.; Cattaneo, M.; Levi, M.; Clark, C.; Iba, T. ISTH interim guidance on recognition and management of coagulopathy in COVID-19. J. Thromb. Haemost. 2020, 18, 1023–1026. [Google Scholar] [CrossRef] [PubMed]
- Barrett, C.D.; Moore, H.B.; Yaffe, M.B.; Moore, E.E. ISTH interim guidance on recognition and management of coagulopathy in COVID-19: A comment. J. Thromb. Haemost. 2020. [Google Scholar] [CrossRef] [PubMed]
- Thachil, J. The versatile heparin in COVID-19. J. Thromb. Haemost. 2020, 18, 1020–1022. [Google Scholar] [CrossRef] [Green Version]
- Harr, J.N.; Moore, E.E.; Chin, T.L.; Ghasabyan, A.; Gonzalez, E.; Wohlauer, M.V.; Sauaia, A.; Banerjee, A.; Silliman, C.C. Postinjury hyperfibrinogenemia compromises efficacy of heparin-based venous thromboembolism prophylaxis. Shock 2014, 41, 33–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turshudzhyan, A. Anticoagulation Options for Coronavirus Disease 2019 (COVID-19)-Induced Coagulopathy. Cureus 2020, 12, e8150. [Google Scholar] [CrossRef] [PubMed]
- Schutgens, R.E. DOAC in COVID-19: Yes or No? HemaSphere 2021, 5, e526. [Google Scholar] [CrossRef] [PubMed]
- Driggin, E.; Madhavan, M.V.; Bikdeli, B.; Chuich, T.; Laracy, J.; Biondi-Zoccai, G.; Brown, T.S.; Der Nigoghossian, C.; Zidar, D.A.; Haythe, J. Cardiovascular considerations for patients, health care workers, and health systems during the COVID-19 pandemic. J. Am. Coll. Cardiol. 2020, 75, 2352–2371. [Google Scholar] [CrossRef]
- Testa, S.; Prandoni, P.; Paoletti, O.; Morandini, R.; Tala, M.; Dellanoce, C.; Giorgi-Pierfranceschi, M.; Betti, M.; Battista Danzi, G.; Pan, A. Direct oral anticoagulant plasma levels’ striking increase in severe COVID-19 respiratory syndrome patients treated with antiviral agents: The Cremona experience. J. Thromb. Haemost. 2020, 18, 1320–1323. [Google Scholar] [CrossRef]
- Burstyn-Cohen, T.; Heeb, M.J.; Lemke, G. Lack of protein S in mice causes embryonic lethal coagulopathy and vascular dysgenesis. J. Clin. Investig. 2009, 119, 2942–2953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fair, D.S.; Marlar, R.A.; Levin, E.G. Human endothelial cells synthesize protein S. Blood 1986, 67, 1168–1171. [Google Scholar] [CrossRef] [Green Version]
- Stern, D.; Brett, J.; Harris, K.; Nawroth, P. Participation of endothelial cells in the protein C-protein S anticoagulant pathway: The synthesis and release of protein S. J. Cell Biol. 1986, 102, 1971–1978. [Google Scholar] [CrossRef]
- Schurgers, L.J.; Shearer, M.J.; Hamulyák, K.; Stöcklin, E.; Vermeer, C. Effect of vitamin K intake on the stability of oral anticoagulant treatment: Dose-response relationships in healthy subjects. Blood 2004, 104, 2682–2689. [Google Scholar] [CrossRef]
- Paliani, U.; Filippucci, E.; Gresele, P. Significant potentiation of anticoagulation by flu-vaccine during the season 2001–2002. Haematologica 2003, 88, 599–600. [Google Scholar] [PubMed]
- Kuo, A.; Brown, J.; Clinard, V. Effect of influenza vaccination on international normalized ratio during chronic warfarin therapy. J. Clin. Pharm. Ther. 2012, 37, 505–509. [Google Scholar] [CrossRef] [PubMed]
- Iorio, A.; Basileo, M.; Marcucci, M.; Guercini, F.; Camilloni, B.; Paccamiccio, E.; Vecchioli, M.; Iorio, A.M. Influenza vaccination and vitamin K antagonist treatment: A placebo-controlled, randomized, double-blind crossover study. Arch. Intern. Med. 2010, 170, 609–616. [Google Scholar] [CrossRef] [Green Version]
- Koshihara, Y.; Hoshi, K.; Shiraki, M. Vitamin K2 (menatetrenone) inhibits prostaglandin synthesis in cultured human osteoblast-like periosteal cells by inhibiting prostaglandin H synthase activity. Biochem. Pharmacol. 1993, 46, 1355–1362. [Google Scholar] [CrossRef]
- Chen, G.; Wu, D.; Guo, W.; Cao, Y.; Huang, D.; Wang, H.; Wang, T.; Zhang, X.; Chen, H.; Yu, H. Clinical and immunological features of severe and moderate coronavirus disease 2019. J. Clin. Investig. 2020, 130, 2620–2629. [Google Scholar] [CrossRef] [Green Version]
- Ragab, D.; Salah Eldin, H.; Taeimah, M.; Khattab, R.; Salem, R. The COVID-19 Cytokine Storm; What We Know So Far. Front. Immunol. 2020, 11, 1446. [Google Scholar] [CrossRef]
- Puig, F.; Fuster, G.; Adda, M.; Blanch, L.; Farre, R.; Navajas, D.; Artigas, A. Barrier-protective effects of activated protein C in human alveolar epithelial cells. PLoS ONE 2013, 8, e56965. [Google Scholar] [CrossRef] [Green Version]
- Urawa, M.; Kobayashi, T.; D’Alessandro-Gabazza, C.N.; Fujimoto, H.; Toda, M.; Roeen, Z.; Hinneh, J.A.; Yasuma, T.; Takei, Y.; Taguchi, O.; et al. Protein S is protective in pulmonary fibrosis. J. Thromb. Haemost. 2016, 14, 1588–1599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akhmerov, A.; Marbán, E. COVID-19 and the Heart. Circ. Res. 2020, 126, 1443–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dalmeijer, G.; van der Schouw, Y.T.; Magdeleyns, E.; Ahmed, N.; Vermeer, C.; Beulens, J. The effect of menaquinone-7 supplementation on circulating species of matrix Gla protein. Atherosclerosis 2012, 225, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Dalmeijer, G.W.; van der Schouw, Y.T.; Vermeer, C.; Magdeleyns, E.J.; Schurgers, L.J.; Beulens, J.W. Circulating matrix Gla protein is associated with coronary artery calcification and vitamin K status in healthy women. J. Nutr. Biochem. 2013, 24, 624–628. [Google Scholar] [CrossRef] [PubMed]
- Rennenberg, R.J.; De Leeuw, P.W.; Kessels, A.G.; Schurgers, L.J.; Vermeer, C.; Van Engelshoven, J.M.; Kemerink, G.J.; Kroon, A.A. Calcium scores and matrix Gla protein levels: Association with vitamin K status. Eur. J. Clin. Investig. 2010, 40, 344–349. [Google Scholar] [CrossRef] [PubMed]
- Braam, L.A.; Hoeks, A.P.; Brouns, F.; Hamulyák, K.; Gerichhausen, M.J.; Vermeer, C. Beneficial effects of vitamins D and K on the elastic properties of the vessel wall in postmenopausal women: A follow-up study. Thromb. Haemost. 2004, 91, 373–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enomoto, N.; Suda, T.; Kono, M.; Kaida, Y.; Hashimoto, D.; Fujisawa, T.; Inui, N.; Nakamura, Y.; Imokawa, S.; Funai, K.; et al. Amount of elastic fibers predicts prognosis of idiopathic pulmonary fibrosis. Respir. Med. 2013, 107, 1608–1616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piscaer, I.; van den Ouweland, J.M.W.; Vermeersch, K.; Reynaert, N.L.; Franssen, F.M.E.; Keene, S.; Wouters, E.F.M.; Janssens, W.; Vermeer, C.; Janssen, R. Low Vitamin K Status Is Associated with Increased Elastin Degradation in Chronic Obstructive Pulmonary Disease. J. Clin. Med. 2019, 8, 1116. [Google Scholar] [CrossRef] [Green Version]
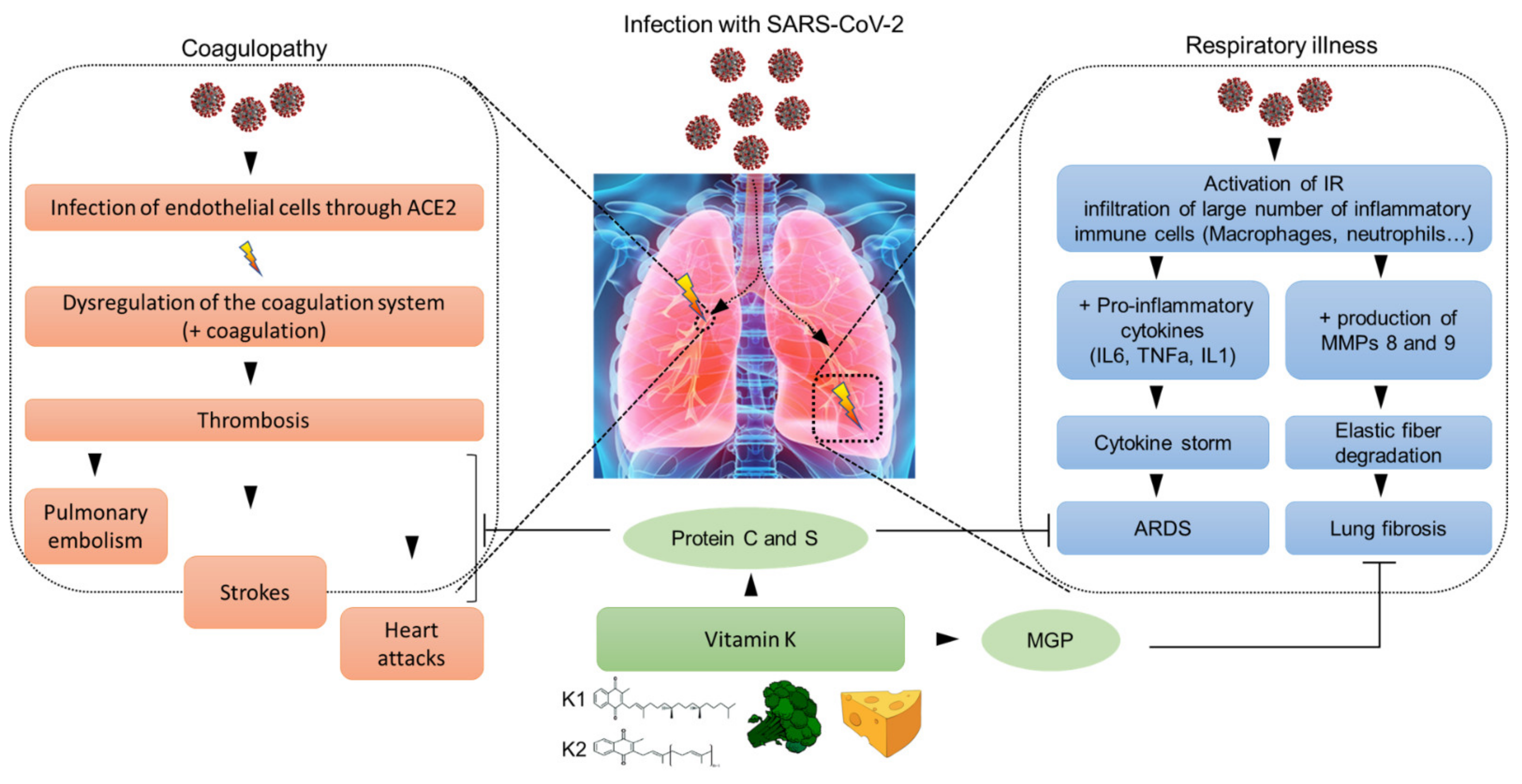
Figure 1.
The potential involvement of coagulopathy in the pathogenesis of SARS-CoV-2 viral infection and potential beneficial effects of vitamin K. Following SARS-CoV-2 infection, the virus binds to lung epithelial cells and endothelial cells that highly express its receptor Angiotensin converting enzyme 2 (ACE2) leading to both respiratory (respiratory illness) and vascular (coagulopathy) complications. The infection of lung epithelial cells triggers the activation of an immune response (IR) leading to secretion of pro-inflammatory cytokines (Interleukin 6 (IL6), Tumor Necrosis Factor alpha (TNFa) and Interleukin 1 (IL1)) and metalloproteinases (MMPs) by infiltrated macrophages. The presence of a high viral load causes massive destruction of lung tissues resulting in acute respiratory distress syndrome (ARDS) as well as lung fibrosis (right panel). The infection of endothelial cells affects the normal process of coagulation leading to venous and arterial thromboembolism that can result in strokes, heart attacks and pulmonary embolism (left panel). The diverse and distinct roles of vitamin K in modulating blood clotting, elastin degradation, immunomodulation and managing vascular health are summarized (lower panel).
Figure 1.
The potential involvement of coagulopathy in the pathogenesis of SARS-CoV-2 viral infection and potential beneficial effects of vitamin K. Following SARS-CoV-2 infection, the virus binds to lung epithelial cells and endothelial cells that highly express its receptor Angiotensin converting enzyme 2 (ACE2) leading to both respiratory (respiratory illness) and vascular (coagulopathy) complications. The infection of lung epithelial cells triggers the activation of an immune response (IR) leading to secretion of pro-inflammatory cytokines (Interleukin 6 (IL6), Tumor Necrosis Factor alpha (TNFa) and Interleukin 1 (IL1)) and metalloproteinases (MMPs) by infiltrated macrophages. The presence of a high viral load causes massive destruction of lung tissues resulting in acute respiratory distress syndrome (ARDS) as well as lung fibrosis (right panel). The infection of endothelial cells affects the normal process of coagulation leading to venous and arterial thromboembolism that can result in strokes, heart attacks and pulmonary embolism (left panel). The diverse and distinct roles of vitamin K in modulating blood clotting, elastin degradation, immunomodulation and managing vascular health are summarized (lower panel).

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kudelko, M.; Yip, T.F.; Hei Law, G.C.; Lee, S.M.Y. Potential Beneficial Effects of Vitamin K in SARS-CoV-2 Induced Vascular Disease? Immuno 2021, 1, 17-29. https://0-doi-org.brum.beds.ac.uk/10.3390/immuno1010003
AMA Style
Kudelko M, Yip TF, Hei Law GC, Lee SMY. Potential Beneficial Effects of Vitamin K in SARS-CoV-2 Induced Vascular Disease? Immuno. 2021; 1(1):17-29. https://0-doi-org.brum.beds.ac.uk/10.3390/immuno1010003
Chicago/Turabian StyleKudelko, Mateusz, Tsz Fung Yip, Grace Chun Hei Law, and Suki Man Yan Lee. 2021. "Potential Beneficial Effects of Vitamin K in SARS-CoV-2 Induced Vascular Disease?" Immuno 1, no. 1: 17-29. https://0-doi-org.brum.beds.ac.uk/10.3390/immuno1010003