
Targeting Hyaluronan Synthesis in Cancer: A Road Less Travelled

The Institute of Cancer Research, London SW3 6JB, UK
Biologics 2023, 3(4), 402-414; https://0-doi-org.brum.beds.ac.uk/10.3390/biologics3040022
Submission received: 10 October 2023
/
Revised: 10 November 2023
/
Accepted: 7 December 2023
/
Published: 12 December 2023
Abstract
:Hyaluronan is one of the major components of the extracellular matrix and is involved in the regulation of multiple processes in both human physiology and disease. In human cancers, hyaluronan metabolism displays remarkable alterations, leading to the accumulation of large amounts of hyaluronan matrices in the tumoural tissues. The altered levels of hyaluronan in the tumours stem from the enhanced expression and activity of hyaluronan synthases in both tumour and stromal cells. Moreover, hyaluronidase activity is also upregulated in cancer, leading to the generation of lower molecular weight hyaluronan fragments that in turn assist tumour growth, neo-angiogenesis and the metastatic cascade. Hyaluronan accumulation in malignant tissues not only assists tumour growth and metastases but is also associated with worse outcomes in cancer patients. Therefore, targeting hyaluronan synthesis emerges as an interesting strategy that might be employed for cancer treatment. This review article summarises current evidence and discusses ways to move forward in the field of targeting hyaluronan synthesis for cancer therapy.
1. Introduction
Hyaluronan is one of the most important components of the extracellular matrix surrounding the cells of several different tissues. Hyaluronan is synthesised and secreted to the extracellular matrix from cells like fibroblasts, endothelial and various types of immune cells. Hyaluronan production is indispensable for proper embryonic development, and knock-out of hyaluronan synthase 2 (HAS2) causes embryonic lethality in mice due to severe cardiac defects. Highlighting the importance of HAS2-synthesised hyaluronan, no activity-altering mutations have been found in humans. Apart from embryogenesis, hyaluronan is important for a multitude of normal tissue functions like skin hydration and joint lubrication. Hyaluronan can retain large amounts of water. Therefore, hyaluronan in the skin holds the appropriate amount of water and prevents wrinkle formation. In the joints, the water-retentive properties of hyaluronan allow for reduced friction and impact absorption during bone movements. The importance of hyaluronan for proper joint function is showcased by the fact that in diseases like osteoarthritis, intra-joint hyaluronan is reduced and injection of exogenous hyaluronan in the joint is used to alleviate pain.
Altered hyaluronan metabolism has been observed in many different human diseases. Specifically, hyaluronan synthesis and fragmentation has been observed in diabetes, chronic inflammation and viral infections [1,2,3,4]. Specifically in cancer, hyaluronan metabolism deregulation is a frequent event. In a multitude of cancer types, large increments of hyaluronan are detected in the tumour compared to the respective normal tissue. Furthermore, enhanced hyaluronan synthesis is accompanied by enhanced degradation owing to the higher expression and activity of hyaluronidases. The degradation of hyaluronan produces intra-tumoural hyaluronan fragments that promote inflammation, angiogenesis, tumour growth and metastasis. Therefore, targeting hyaluronan metabolism for the treatment of human pathologies, specifically cancer, emerges as a promising avenue. Nevertheless, the lack of small-molecule inhibitors of hyaluronan synthesis owing to the unknown mechanism of hyaluronan synthesis—until recently [5]—has hindered such approaches being introduced into the clinic and the data regarding the effectiveness of hyaluronan synthesis inhibition in vivo remain sparse.
In this review, I lay out the available evidence from pre-clinical models regarding the development of hyaluronan synthesis-interfering strategies and further discuss the potential of translating hyaluronan synthesis inhibition into the clinic. Moreover, I also propose some ideas (like the use of PROTACs) that can potentially help the field move forward.
2. Hyaluronan Metabolism
2.1. Hyaluronan Synthesis
Hyaluronan is a ubiquitously present polysaccharide in the human body and is synthesised by many different—if not all—cell types (e.g., fibroblasts, macrophages, endothelial and stem cells), albeit in a cell- and context-specific manner. Interestingly, hyaluronan is the only glycosaminoglycan that has evolved to be synthesised in the plasma membrane and not in the Golgi compartment. Moreover, hyaluronan is not covalently attached to proteins like the rest of the glycosaminoglycans. The transmembrane enzymes that synthesise hyaluronan are termed hyaluronan synthases (HASes) and there are three different isoenzymes in humans (HAS1-3), which derive from distinct genes. Of note, HASes display tissue-type specific expression and display different catalytic properties leading to the synthesis of different-molecular-weight hyaluronan [6]. It is worth mentioning that the hyaluronan synthase structure was only recently described for chlorella virus hyaluronan synthase, whereas the exact mechanism and structure of the human orthologue remains to be discovered.
Hyaluronan is comprised of glucuronic acid (GlcUA) and N-acetyl-glucosamine (GlcNAc), which are derived from their UDP-bound precursors (UDP–GlcUA and UDP–GlcNAc). In principle, both precursors can originate from glucose, albeit via different metabolic intermediates. UDP–GlcUA derives from 1-P-glucose, whereas UDP–GlcNAc derives from 6-P-fructose through a pathway known as hexosamine biosynthesis, which also requires glucosamine and acetyl-coA. These UDP-activated sugars are then used up by hyaluronan synthases to produce the hyaluronan moieties, which are comprised of repeating disaccharides of GlcUA bound to GlcNAc with alternate β-1,4 and β-1,3 glycosidic bonds [7,8]. Hyaluronan can reach up to millions of kDa in size and can be conformed into different 3D structures according to its size. Although hyaluronan can be found as a continuum of different sizes, usually it is roughly categorised as high-(HMW) or low-molecular-weight (LMW) hyaluronan. HMW hyaluronan can form globular or cylindrical structures among others, whereas LMW can form fibres or stacks [9,10].
Hyaluronan biosynthesis rates can be tightly regulated in both physiological and pathological conditions by a multitude of post-translational modifications. So far, HAS proteins have been shown to be decorated with post-translational modifications like ubiquitination and phosphorylation. Moreover, HAS dimerisation can also affect HAS hyaluronan-synthesising activity. Specifically, HAS2 activity is negatively regulated by AMPKα phosphorylation at Thr110 [11], while phosphorylation of Thr328 by casein kinase-1 (CK1) is essential for HAS2 activation [12]. HAS activity can be also induced through phosphorylation by PKC [13]. Moreover, ubiquitination of HAS2 in K190 residue potentiates its activity as the mutation of this site to K190R inhibits HAS2 [14]. Furthermore, the ubiquitin-specific protease USP17 removes poly-ubiquitination and stabilises HAS2, leading to enhanced hyaluronan production. On the other hand, USP4 preferentially removes mono-ubiquitination from HAS2 and displays the opposite effect on hyaluronan synthesis [15]. Interestingly, HAS2 can form homodimers or heterodimers with HAS3 [14]. Hyaluronan production is also regulated by growth factors like EGF, PDGF, HGF and TGF-β, which control HAS transcription and activity. Of note, growth factor treatment activates subcellular signalling that, in most cases, leads to the upregulation of hyaluronan synthesis, which makes hyaluronan an important intermediate in growth factor-induced cellular functions like the epithelial to mesenchymal transition (EMT) programme, differentiation, migration and proliferation. Several studies have also highlighted that HAS gene expression is under epigenetic control from HAS1-AS1 as well as chromatin modifications (reviewed in [4]). Therefore, hyaluronan synthesis seems to be a process that is under consistent tight control.
2.2. Hyaluronan Catabolism
Hyaluronan amounts in the extracellular and interstitial space are also under tight control through its enzymatic digestion [16]. Systemically, hyaluronan chains are initially degraded into smaller fragments interstitially, which are then transferred to the liver, kidneys and spleen for complete degradation [17,18,19,20]. Hyaluronan degradation is carried out by hyaluronidases and in humans, several hyaluronidase genes have been identified: HYAL-1, HYAL-2, PH-20 [21,22,23]. The interest in hyaluronan catabolism was also recently reignited with the discovery of two new proteins involved in the process, namely TMEM2 and HYBID [24,25]. Albeit TMEM2 was initially suggested to be a new hyaluronidase [20], a recent report suggested that TMEM2 is not a catalytic hyaluronidase, rather it is a regulator of hyaluronan metabolism via HYBID and HAS2 regulation [26]. Further research into this subject will hopefully help to identify the role of TMEM2 and HYBID on hyaluronan catabolism. In any case, both HYBID and TMEM2 appear to be important regulators in hyaluronan degradation.
Hyaluronan is a polysaccharide that displays high turnover rates (approximately 1–1.5 days) [20]. In light of the role of hyaluronan as a potent anti-oxidant [27], its fast recycling rate is not surprising since hyaluronan quenching of reactive oxygen species (ROS) leads to its non-enzymatic degradation [28,29] and hyaluronan of higher molecular sizes needs to be possibly replenished, while lower molecular size fragments occurring through reactions with ROS might be damaging to the tissue and need to be rapidly removed.
3. Hyaluronan in Normal Physiology
Hyaluronan displays important functions in normal physiology and development. The clearest demonstration of hyaluronan’s importance in normal development is that HAS2 knock-out is embryonically lethal in mice, since the lack of HAS2-synthesised hyaluronan leads to improper heart and vascular system development [30]. HAS3-synthesised hyaluronan also exerts important roles as it has been shown to be crucial for semicircular canal morphogenesis in the ear [31]. Hyaluronan, in a size-dependent manner, also contributes to the normal formation of breast tissue as its receptor RHAMM assists the EGF-regulated branching of the breast [32].
Skin is a tissue that displays a higher accumulation of hyaluronan. In the skin, hyaluronan retains water to regulate ion concentration, hydration and proper skin structure. During physiological ageing, hyaluronan size in the skin is reduced and LMW fragments promote inflammation. Interestingly, in photoaged skin, hyaluronan fragmentation by CEMIP and ROS is thought to promote the formation of wrinkles, by reducing tissue hydration and altering viscosity [33,34,35].
The immune system is another system that lies under the umbrella of hyaluronan-assisted regulation. Hyaluronan, by interacting with its principal receptor CD44, not only regulates immune cell function but also guides proper stem cell homing on bone marrow niches. Moreover, hyaluronan produced by endothelial cells allows for the attachment of immune cells to the endothelial glycocalyx and extravasation to the underlying tissue [35,36,37]. For a complete review of hyaluronan’s effects on the immune system, the reader is advised to look in [36].
Unambiguously, hyaluronan appears to be not only an important molecule for proper tissue and bodily functions but also indispensable for the development of life—a lesson learned from the HAS2-knock-out mice.
4. Hyaluronan Metabolism Deregulation in Tumours
Hyaluronan is highly upregulated in many tumour types including breast, prostate and lung cancer. While hyaluronan secretion by both cancer and stromal cells has been observed in tumours, most hyaluronan functions regarding tumour development, growth and progression can be understood through its interactions with its receptors CD44 and RHAMM, which are also highly expressed in human cancers.
In non-small cell lung cancer tumours, hyaluronan has been suggested as a novel biomarker recently [38], and in both prostate [39] and breast cancer [40], hyaluronan accumulation is associated with worse outcomes and increased incidence of metastases. Glioblastoma is a tumour type that is often found to produce high amounts of hyaluronan, which induces tumour cell migration, invasion, proliferation and therapy resistance, through its interactions with CD44 and RHAMM [41,42,43]. In gastric cancer cells, hyaluronan synthesis is also associated not only with local and nodal spread but also with reduced survival rates [44]. A marked increase of hyaluronan is also evident in malignant mesothelioma, where it promotes pro-tumourigenic activities [45,46]. Recently, it was also shown that hyaluronan remodels the microenvironment of mesenchymal colorectal tumours, promoting the appearance of a more heterogeneous epithelial environment that favours invasion through interaction with activated fibroblasts [47]. Hyaluronan is also involved in neo-angiogenesis—a crucial process of tumour development and progression [48,49]. Hyaluronan can induce VEGF-A and VEGFR1 expression in endothelial cells and subsequently promote angiogenesis [50]. Furthermore, CD44, HYAL-2 and hyaluronan are not only important for endothelial cell differentiation but also for cancer cell dissemination. Moreover, CD44-expressing cancer cells can adhere to endothelial cells—a critical initial step in tumour cell extravasation and subsequent metastasis [51]. Also, tumour cell-derived hyaluronidase can cleave hyaluronan in the tumour microenvironment to low-molecular-weight fragments that can promote tumour vascularisation [52].
Hyaluronan has also been established as a biomarker in several cancer types. In metastatic breast cancer, high plasma hyaluronan levels are associated with poor prognosis of patients [53]. In androgen receptor-negative breast cancer, high HAS2 expression is correlated with poor prognostic features like higher grade and lymphovascular invasion [54]. In an analysis of The Cancer Genome Atlas (TCGA), a study identified hyaluronan synthases and hyaluronidase-1 levels to be potent predictors of metastasis and poor patient survival [55]. In colorectal cancer, high tumour cytosolic hyaluronan levels are associated with unfavourable patient outcomes [56]. Hyaluronan’s presence in the urine has also been documented to be a biomarker to aid in diagnosing and grading transitional cell carcinoma of the urinary bladder [57,58]. Interestingly, work from the same group demonstrated that patients with a nephroblastoma of embryonic origin (Wilms’ tumour) display higher levels of urinary hyaluronan compared with patients after chemotherapy and surgery or patients suffering hypernephroma and normal individuals [59]. In these works, the authors also describe a method capable of detecting down to 0.2 µg/L of urinary hyaluronan, which makes the assay 50 times more sensitive than previously described assays. Future works will hopefully utilise this sensitive method to measure hyaluronan in other biological fluids (e.g., blood) and different tumour types to determine hyaluronan’s status as a cancer biomarker. Measuring biological fluid hyaluronan will hopefully offer an easy, rapid and accurate method to aid cancer diagnostics.
The upregulation of hyaluronan synthesis in tumours occurs usually through r=the upregulation of HAS expression. Multiple growth factors like TGF-β, HGF, PDGF, EGF and several cytokines upregulate hyaluronan production [4]. Specifically, in breast epithelial cells, TGF-β induces EMT that apparently depends on the upregulation of HAS2 via the activation of the p38 MAPK pathway. HAS2-synthesised hyaluronan then promotes a malignant phenotype via suppressing E-cadherin—a major anti-tumour protein [60,61,62]. Furthermore, HAS2-synthesised hyaluronan in breast cancer cells promotes local inflammation through enhancing the expression of cytokine-related genes and regulates CD206+ macrophage infiltration in vivo [63]. Interestingly, hyaluronan can regulate drug resistance through CD44. Specifically, hyaluronan/CD44 interactions activate the PI3K pathway, which promotes MDR protein expression, leading to chemotherapy resistance [64]. Hyaluronan also controls MDR expression through the Oct4–Sox2–NANOG signalling axis, a pathway involved in enhanced tumour therapy resistance [65]. Hyaluronan, through its interaction with CD44, can also regulate and promote cancer stem cell properties. For a comprehensive analysis of hyaluronan role on cancer stemness, refer to our previous review [66].
An intricacy observed in tumour-derived hyaluronan is that it is usually fragmented LMW hyaluronan. Indeed, the upregulation of hyaluronidases in tumours has been shown to promote hyaluronan cleavage and accumulation of a continuous supply of LMW hyaluronan that can drive local inflammation and tumour growth and invasion [67,68]. Notably, hyaluronidase upregulation was noted in breast cancer and typically, ER-negative cells display higher hyaluronidase expression than ER-positive cells [69]. A significant elevation of hyaluronidases has been also documented in prostate cancer, where they are associated with advanced tumour progression [70]. It is also important to note that combined upregulation of HAS and hyaluronidases drives tumour growth, as shown in several cancer models [68]. On the other hand, high-molecular-weight hyaluronan displays significant anti-cancer properties. Specifically, in naked mole rats, hyaluronan with a very high molecular weight promotes cancer resistance in this rodent [71]. These cytoprotective properties of very-high-molecular-weight hyaluronan are associated with the p53 pathway [72]. Interestingly, these cytoprotective properties of high-molecular-weight hyaluronan, which also relies on quenching of oxidative stress, can be imported to other species, as mice that expressed naked-mole-rat hyaluronan synthase displayed increased longevity [73]. Whether this very-high-molecular-weight hyaluronan can be utilised for cancer treatment is an exciting possibility that remains to be investigated.
5. Targeting Hyaluronan Synthesis
Arguably, according to the available documented evidence, hyaluronan metabolism deregulation seems to be a frequent event in human cancer. Albeit many efforts have been focused on inhibiting hyaluronan/CD44 interactions using a multitude of approaches like antibodies and nanomedicine, this aspect of intervention has not flourished into clinical accomplishments so far. Interestingly, a significant number of efforts have focused on utilising hyaluronan as a drug carrier, owing to its biocompatibility. Moreover, since many different types of cancer cells overexpress the hyaluronan receptors CD44 and RHAMM, hyaluronan-based nanocarriers can transfer chemotherapeutics effectively in CD44–or RHAMM–expressing tumour cells [74,75]. Although hyaluronan metabolism per se has been shown to be deregulated in cancer, the development of specific drugs targeting hyaluronan metabolism and specifically HASes has been hindered so far by the unknown mechanism and structure of HASes. Bellow, I review the available evidence regarding targeting hyaluronan synthesis with small molecules and genetic manipulation and discuss possible avenues of moving forward in the future.
5.1. Small Molecules
4-methyl-umbelliferone (4-MU) is unequivocally the most studied small molecule inhibitor of hyaluronan synthesis. 4-MU is a plant-derived natural extract from chamomile and is categorised as a coumarin analogue. It inhibits hyaluronan synthesis by two distinct and sometimes complementary mechanisms. Firstly, 4-MU is glucuronidated by UDP–glucuronosyltransferase (UGT)—the enzyme that normally transfers UDP to GlcUA to produce UDP–GlcUA)—into 4-MU-glucuronide. Thus, the availability of UDP–GlcUA is lowered and subsequently, hyaluronan synthesis is reduced. Secondly, 4-MU reduces HAS expression through an unknown mechanism, although this observation is cell-type-dependent [76].
4-MU has shown a promising capacity to inhibit several aspects of tumour growth and development. In colon cancer, in vitro 4-MU inhibits proliferation and migration, whereas in vivo synergises with other drugs to enhance anti-tumour immunity [77,78,79]. In mice models of pancreatic cancer, 4-MU treatment improves survival rates and limits the formation of metastases in the liver [80,81,82,83,84]. Prostate cancer is another tumour type that 4-MU administration reduces the growth of tumours in rodents [85,86]. In ovarian and breast cancer, which frequently display remarkable increments of hyaluronan, 4-MU effectively inhibits hyaluronan production and attenuates cancer cell proliferation, migration and invasion [87,88,89,90,91,92]. Of note, hyaluronan has been shown to be effective against many other types of tumours like glioblastoma, hepatocellular carcinoma, melanoma, chronic myeloid leukaemia, osteosarcoma and oesophageal squamous cell carcinoma [76,93]. On top of the remarkable success of 4-MU in pre-clinical models, it is also worth noting that 4-MU is an already approved drug for use in humans for the treatment of biliary spasm; it decreases hyaluronan in humans and displays a great safety profile [94,95]. Therefore, it appears that 4-MU fulfils many criteria as a very promising candidate drug that can possibly enter clinical trials as an adjuvant for already approved cancer-targeting schemes.
Apart from 4-MU, in previous work, we also identified salicylate—the precursor of the widely used acetyl-salicylate (aspirin)—as a potent inhibitor of hyaluronan synthesis in breast cancer cells. Mechanistically, salicylate activates AMPK, which in turn inhibits hyaluronan production, possibly through the phosphorylation of HAS2—the main HAS isoform expressed in the context of breast cancer. Furthermore, salicylate has also potently inhibited the migration and proliferation of triple-negative breast cancer cells [96]. In our efforts to identify a more potent small molecule hyaluronan synthesis inhibitor, we discovered 5′-Deoxy-5′-(1,3-Diphenyl-2-Imidazolidinyl)-Thymidine (DDIT) as a potent inhibitor of hyaluronan synthesis. Of note, DDIT appears to be approximately 12 times more potent than 4-MU as a hyaluronan synthesis inhibitor. Although DDIT inhibits both HAS2 and HAS3 in different types of cancer cells (breast and lung cancer, glioblastoma cells), in that study, we were not able to pinpoint the exact mechanism of DDIT-mediated HAS inhibition. Future studies will hopefully address DDIT’s mode of action. DDIT treatment also caused marked reductions in breast cancer cell proliferation, migration, invasion and cancer stem cell self-renewal [97].
A collective presentation of the available hyaluronan synthesis inhibitors and their effects on tumours is summarised in Table 1. Certainly, in the future, more studies will hopefully discover potent hyaluronan synthesis inhibitors that can also exert significant anti-cancer properties.
5.2. Genetic Manipulation
Since HASes display elevated expression in human cancer, inhibiting their expression in tumour cells via genetic manipulation seems to be a promising idea. The feasibility of this approach has been demonstrated so far in vitro. Owing to the recognition of HAS2 as the most highly expressed HAS isoform in human tumours, most studies are focused on the genetic inhibition of this specific isoform rather than HAS1 or HAS3. In breast cancer, silencing HAS2 utilising specific siRNAs suppressed the malignant phenotype of tumour cells [104]. The validity of targeting HAS2 at the gene level utilising CRISPR/Cas9 on tumour cells has not been assessed. Although germline loss of HAS2 via knock-out leads to embryonic lethality, somatic loss of HAS2 might be tolerated as it was successfully performed in rat chondrosarcoma chondrocytes [105]. Furthermore, the genetic loss of HAS2 in somatic cell lineages might be tolerated owing to the upregulation of HAS1 and HAS3 observed after silencing HAS2 with siRNAs [104]—probably to compensate for the loss of extracellular hyaluronan synthesised from HAS2. Whether this mechanism is activated in the case of CRISPR/Cas9-mediated HAS2 knock-out is not known and hopefully future studies will enhance our understanding regarding this aspect.
Utilising HAS2-AS1 is also an appealing strategy to target cancer cells, since HAS2 expression and therefore hyaluronan synthesis lie under the control of this long non-coding RNA. However, there are conflicting reports regarding the role of HAS2-AS1 since it has been shown that HAS2-AS1 can promote TGF-β-induced EMT in mouse mammary epithelial cells [106], while in triple-negative human cancer cells, the introduction of HAS2-AS1 reduces aggressiveness in a hyaluronan-independent manner [107]. Interestingly, hyaluronan production from human aortic smooth muscle cells appears to lie under the control of HAS2-AS1 [108]. These results point to a cell type- and species-specific regulation of hyaluronan synthesis by HAS2-AS1. In any case, HAS2-AS1 seems to be an appealing way to possibly intervene with hyaluronan metabolism and further studies might clarify this issue in the future. Moreover, in vivo experimentation might provide useful insight into the further development of such therapeutic approaches.
5.3. Moving Forward
Clearly, to this day, there is only a handful of studies investigating the targeting of hyaluronan synthesis in human cancer, probably owing to the lack of HAS mechanism, structure and appropriate antibodies. Studies that will focus on resolving these issues might lead to significant blooming of hyaluronan synthesis-targeting strategies. Furthermore, novel approaches that might be employed for hyaluronan synthase targeting include proteolysis targeting chimeric (PROTAC) technology, which has shown remarkable results in specific protein targeting. PROTAC is a chemical knock-down approach that degrades a selected target protein through the ubiquitin-proteasome system [109]. In cancer, the use of PROTACs has shown promising initial results [110,111,112,113]. For example, hypoxia-activated PROTACs have shown a potent ability to degrade EGFR in hypoxic tumour cells specifically [114]. In another study, protein degraders were used to downregulate tumour-related proteins like ALK and EGFR in non-small lung and breast cancer cells, leading to a reduction in their aggressive properties. Of note, these PROTACs are well tolerated in vivo [115]. Since enzymes involved in the ubiquitination and stabilisation of HAS2 have already been identified, namely USP4 and USP17 [15], one could envision utilising this approach to specifically target HAS2-expressing tumour cells. For example, designing PROTACs that will lead to enhanced degradation of USP17, which stabilises HAS2, might lead to reduced HAS2 protein levels and subsequently reduce tumour hyaluronan. Subsequently, downregulation of HAS2-synthesised hyaluronan will hopefully lead to the inhibition of tumour development, progression and metastases according to the tumour aspects that hyaluronan is shown to regulate. Furthermore, PROTACs directly targeting the HAS2 protein could be designed, although such an approach depends on the successful discovery of human HAS2 structure. The feasibility of such a scheme will hopefully be addressed in future studies both in vitro and in vivo.
6. Conclusions
So far, the available literature suggests that tackling hyaluronan synthesis can be of therapeutic value in cancer treatment (Figure 1). Undoubtedly, a lot remains to be learned in the field of hyaluronan synthesis in general and in cancer specifically. The main challenge that the field is currently facing is the discovery of the human HAS structure and identification of its differential regulation between normal and cancerous tissues. Identifying differential modes of HAS activation in cancer compared to normal physiology will hopefully lead to successful targeting of tumour hyaluronan metabolism and spare any side effects.
It is clear that targeting hyaluronan synthesis in cancer is currently only in its infancy but hopefully will blossom into a field that will yield useful therapeutic strategies that will assist the current clinical advancements in the treatment of human cancers.
Funding
This research received no external funding.
Conflicts of Interest
The author declares no conflict of interest.
References
- Garantziotis, S.; Savani, R.C. Hyaluronan biology: A complex balancing act of structure, function, location and context. Matrix Biol. 2019, 78–79, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Chanmee, T.; Itano, N. Hyaluronan: Metabolism and Function. Biomolecules 2020, 10, 1525. [Google Scholar] [CrossRef] [PubMed]
- Heldin, P.; Kolliopoulos, C.; Lin, C.Y.; Heldin, C.H. Involvement of hyaluronan and CD44 in cancer and viral infections. Cell Signal. 2020, 65, 109427. [Google Scholar] [CrossRef] [PubMed]
- Heldin, P.; Lin, C.Y.; Kolliopoulos, C.; Chen, Y.H.; Skandalis, S.S. Regulation of hyaluronan biosynthesis and clinical impact of excessive hyaluronan production. Matrix Biol. 2019, 78–79, 100–117. [Google Scholar] [CrossRef] [PubMed]
- Maloney, F.P.; Kuklewicz, J.; Corey, R.A.; Bi, Y.; Ho, R.; Mateusiak, L.; Pardon, E.; Steyaert, J.; Stansfeld, P.J.; Zimmer, J. Structure, substrate recognition and initiation of hyaluronan synthase. Nature 2022, 604, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Itano, N.; Sawai, T.; Yoshida, M.; Lenas, P.; Yamada, Y.; Imagawa, M.; Shinomura, T.; Hamaguchi, M.; Yoshida, Y.; Ohnuki, Y.; et al. Three isoforms of mammalian hyaluronan synthases have distinct enzymatic properties. J. Biol. Chem. 1999, 274, 25085–25092. [Google Scholar] [CrossRef] [PubMed]
- Vigetti, D.; Karousou, E.; Viola, M.; Deleonibus, S.; De Luca, G.; Passi, A. Hyaluronan: Biosynthesis and signaling. Biochim. Biophys. Acta 2014, 1840, 2452–2459. [Google Scholar] [CrossRef]
- Caon, I.; Parnigoni, A.; Viola, M.; Karousou, E.; Passi, A.; Vigetti, D. Cell Energy Metabolism and Hyaluronan Synthesis. J. Histochem. Cytochem. 2021, 69, 35–47. [Google Scholar] [CrossRef]
- Spagnoli, C.; Korniakov, A.; Ulman, A.; Balazs, E.A.; Lyubchenko, Y.L.; Cowman, M.K. Hyaluronan conformations on surfaces: Effect of surface charge and hydrophobicity. Carbohydr. Res. 2005, 340, 929–941. [Google Scholar] [CrossRef]
- Cowman, M.K.; Matsuoka, S. Experimental approaches to hyaluronan structure. Carbohydr. Res. 2005, 340, 791–809. [Google Scholar] [CrossRef]
- Vigetti, D.; Clerici, M.; Deleonibus, S.; Karousou, E.; Viola, M.; Moretto, P.; Heldin, P.; Hascall, V.C.; De Luca, G.; Passi, A. Hyaluronan synthesis is inhibited by adenosine monophosphate-activated protein kinase through the regulation of HAS2 activity in human aortic smooth muscle cells. J. Biol. Chem. 2011, 286, 7917–7924. [Google Scholar] [CrossRef] [PubMed]
- Kasai, K.; Kuroda, Y.; Takabuchi, Y.; Nitta, A.; Kobayashi, T.; Nozaka, H.; Miura, T.; Nakamura, T. Phosphorylation of Thr(328) in hyaluronan synthase 2 is essential for hyaluronan synthesis. Biochem. Biophys. Res. Commun. 2020, 533, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, Y.; Kasai, K.; Nanashima, N.; Nozaka, H.; Nakano, M.; Chiba, M.; Yoneda, M.; Nakamura, T. 4-Methylumbelliferone inhibits the phosphorylation of hyaluronan synthase 2 induced by 12-O-tetradecanoyl-phorbol-13-acetate. Biomed. Res. 2013, 34, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Karousou, E.; Kamiryo, M.; Skandalis, S.S.; Ruusala, A.; Asteriou, T.; Passi, A.; Yamashita, H.; Hellman, U.; Heldin, C.H.; Heldin, P. The activity of hyaluronan synthase 2 is regulated by dimerization and ubiquitination. J. Biol. Chem. 2010, 285, 23647–23654. [Google Scholar] [CrossRef] [PubMed]
- Mehic, M.; de Sa, V.K.; Hebestreit, S.; Heldin, C.H.; Heldin, P. The deubiquitinating enzymes USP4 and USP17 target hyaluronan synthase 2 and differentially affect its function. Oncogenesis 2017, 6, e348. [Google Scholar] [CrossRef] [PubMed]
- Stern, R.; Kogan, G.; Jedrzejas, M.J.; Soltes, L. The many ways to cleave hyaluronan. Biotechnol. Adv. 2007, 25, 537–557. [Google Scholar] [CrossRef] [PubMed]
- Stern, R. Hyaluronan catabolism: A new metabolic pathway. Eur. J. Cell Biol. 2004, 83, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Tobisawa, Y.; Fujita, N.; Yamamoto, H.; Ohyama, C.; Irie, F.; Yamaguchi, Y. The cell surface hyaluronidase TMEM2 is essential for systemic hyaluronan catabolism and turnover. J. Biol. Chem. 2021, 297, 101281. [Google Scholar] [CrossRef]
- Jadin, L.; Bookbinder, L.H.; Frost, G.I. A comprehensive model of hyaluronan turnover in the mouse. Matrix Biol. 2012, 31, 81–89. [Google Scholar] [CrossRef]
- Pandey, M.S.; Harris, E.N.; Weigel, J.A.; Weigel, P.H. The cytoplasmic domain of the hyaluronan receptor for endocytosis (HARE) contains multiple endocytic motifs targeting coated pit-mediated internalization. J. Biol. Chem. 2008, 283, 21453–21461. [Google Scholar] [CrossRef]
- Shuttleworth, T.L.; Wilson, M.D.; Wicklow, B.A.; Wilkins, J.A.; Triggs-Raine, B.L. Characterization of the murine hyaluronidase gene region reveals complex organization and cotranscription of Hyal1 with downstream genes, Fus2 and Hyal3. J. Biol. Chem. 2002, 277, 23008–23018. [Google Scholar] [CrossRef] [PubMed]
- Flannery, C.R.; Little, C.B.; Hughes, C.E.; Caterson, B. Expression and activity of articular cartilage hyaluronidases. Biochem. Biophys. Res. Commun. 1998, 251, 824–829. [Google Scholar] [CrossRef] [PubMed]
- Cherr, G.N.; Yudin, A.I.; Overstreet, J.W. The dual functions of GPI-anchored PH-20: Hyaluronidase and intracellular signaling. Matrix Biol. 2001, 20, 515–525. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Nagaoka, A.; Nakamura, S.; Sugiyama, Y.; Okada, Y.; Inoue, S. Murine homologue of the human KIAA1199 is implicated in hyaluronan binding and depolymerization. FEBS Open Bio 2013, 3, 352–356. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Tobisawa, Y.; Inubushi, T.; Irie, F.; Ohyama, C.; Yamaguchi, Y. A mammalian homolog of the zebrafish transmembrane protein 2 (TMEM2) is the long-sought-after cell-surface hyaluronidase. J. Biol. Chem. 2017, 292, 7304–7313. [Google Scholar] [CrossRef]
- Sato, S.; Miyazaki, M.; Fukuda, S.; Mizutani, Y.; Mizukami, Y.; Higashiyama, S.; Inoue, S. Human TMEM2 is not a catalytic hyaluronidase, but a regulator of hyaluronan metabolism via HYBID (KIAA1199/CEMIP) and HAS2 expression. J. Biol. Chem. 2023, 299, 104826. [Google Scholar] [CrossRef] [PubMed]
- Sudha, P.N.; Rose, M.H. Beneficial effects of hyaluronic acid. Adv. Food Nutr. Res. 2014, 72, 137–176. [Google Scholar]
- Soltes, L.; Mendichi, R.; Kogan, G.; Schiller, J.; Stankovska, M.; Arnhold, J. Degradative action of reactive oxygen species on hyaluronan. Biomacromolecules 2006, 7, 659–668. [Google Scholar] [CrossRef]
- Agren, U.M.; Tammi, R.H.; Tammi, M.I. Reactive oxygen species contribute to epidermal hyaluronan catabolism in human skin organ culture. Free Radic. Biol. Med. 1997, 23, 996–1001. [Google Scholar] [CrossRef]
- Camenisch, T.D.; Spicer, A.P.; Brehm-Gibson, T.; Biesterfeldt, J.; Augustine, M.L.; Calabro, A.; Jr Kubalak, S.; Klewer, S.E.; McDonald, J.A. Disruption of hyaluronan synthase-2 abrogates normal cardiac morphogenesis and hyaluronan-mediated transformation of epithelium to mesenchyme. J. Clin. Investig. 2000, 106, 349–360. [Google Scholar] [CrossRef]
- Munjal, A.; Hannezo, E.; Tsai, T.Y.; Mitchison, T.J.; Megason, S.G. Extracellular hyaluronate pressure shaped by cellular tethers drives tissue morphogenesis. Cell 2021, 184, 6313–6325.e18. [Google Scholar] [CrossRef] [PubMed]
- Tolg, C.; Yuan, H.; Flynn, S.M.; Basu, K.; Ma, J.; Tse, K.C.K.; Kowalska, B.; Vulkanesku, D.; Cowman, M.K.; McCarthy, J.B.; et al. Hyaluronan modulates growth factor induced mammary gland branching in a size dependent manner. Matrix Biol. 2017, 63, 117–132. [Google Scholar] [CrossRef] [PubMed]
- Yoshida, H.; Nagaoka, A.; Komiya, A.; Aoki, M.; Nakamura, S.; Morikawa, T.; Ohtsuki, R.; Sayo, T.; Okada, Y.; Takahashi, Y. Reduction of hyaluronan and increased expression of HYBID (alias CEMIP and KIAA1199) correlate with clinical symptoms in photoaged skin. Br. J. Dermatol. 2018, 179, 136–144. [Google Scholar] [CrossRef] [PubMed]
- Papakonstantinou, E.; Roth, M.; Karakiulakis, G. Hyaluronic acid: A key molecule in skin aging. Dermato-Endocrinol. 2012, 4, 253–258. [Google Scholar] [CrossRef] [PubMed]
- Zoller, M. CD44, Hyaluronan, the Hematopoietic Stem Cell, and Leukemia-Initiating Cells. Front. Immunol. 2015, 6, 235. [Google Scholar]
- Jiang, D.; Liang, J.; Noble, P.W. Hyaluronan as an immune regulator in human diseases. Physiol. Rev. 2011, 91, 221–264. [Google Scholar] [CrossRef]
- Lee-Sayer, S.S.; Dong, Y.; Arif, A.A.; Olsson, M.; Brown, K.L.; Johnson, P. The where, when, how, and why of hyaluronan binding by immune cells. Front. Immunol. 2015, 6, 150. [Google Scholar] [CrossRef]
- Gong, J.; Guan, M.; Kim, H.; Moshayedi, N.; Mehta, S.; Cook-Wiens, G.; Larson, B.K.; Zhou, J.; Patel, R.; Lapite, I.; et al. Tumor hyaluronan as a novel biomarker in non-small cell lung cancer: A retrospective study. Oncotarget 2022, 13, 1202–1214. [Google Scholar] [CrossRef]
- Josefsson, A.; Adamo, H.; Hammarsten, P.; Granfors, T.; Stattin, P.; Egevad, L.; Laurent, A.E.; Wikstrom, P.; Bergh, A. Prostate cancer increases hyaluronan in surrounding nonmalignant stroma, and this response is associated with tumor growth and an unfavorable outcome. Am. J. Pathol. 2011, 179, 1961–1968. [Google Scholar] [CrossRef]
- Schwertfeger, K.L.; Cowman, M.K.; Telmer, P.G.; Turley, E.A.; McCarthy, J.B. Hyaluronan, Inflammation, and Breast Cancer Progression. Front. Immunol. 2015, 6, 236. [Google Scholar] [CrossRef]
- Yan, T.; Chen, X.; Zhan, H.; Yao, P.; Wang, N.; Yang, H.; Zhang, C.; Wang, K.; Hu, H.; Li, J.; et al. Interfering with hyaluronic acid metabolism suppresses glioma cell proliferation by regulating autophagy. Cell Death Dis. 2021, 12, 486. [Google Scholar] [CrossRef] [PubMed]
- Park, J.B.; Kwak, H.J.; Lee, S.H. Role of hyaluronan in glioma invasion. Cell Adh. Migr. 2008, 2, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Pibuel, M.A.; Poodts, D.; Diaz, M.; Hajos, S.E.; Lompardia, S.L. The scrambled story between hyaluronan and glioblastoma. J. Biol. Chem. 2021, 296, 100549. [Google Scholar] [CrossRef] [PubMed]
- Setala, L.P.; Tammi, M.I.; Tammi, R.H.; Eskelinen, M.J.; Lipponen, P.K.; Agren, U.M.; Parkkinen, J.; Alhava, E.M.; Kosma, V.M. Hyaluronan expression in gastric cancer cells is associated with local and nodal spread and reduced survival rate. Br. J. Cancer 1999, 79, 1133–1138. [Google Scholar] [CrossRef] [PubMed]
- Vidergar, R.; Balduit, A.; Zacchi, P.; Agostinis, C.; Mangogna, A.; Belmonte, B.; Grandolfo, M.; Salton, F.; Biolo, M.; Zanconati, F.; et al. C1q-HA Matrix Regulates the Local Synthesis of Hyaluronan in Malignant Pleural Mesothelioma by Modulating HAS3 Expression. Cancers 2021, 13, 416. [Google Scholar] [CrossRef] [PubMed]
- Asplund, T.; Versnel, M.A.; Laurent, T.C.; Heldin, P. Human mesothelioma cells produce factors that stimulate the production of hyaluronan by mesothelial cells and fibroblasts. Cancer Res. 1993, 53, 388–392. [Google Scholar] [PubMed]
- Martinez-Ordonez, A.; Duran, A.; Ruiz-Martinez, M.; Cid-Diaz, T.; Zhang, X.; Han, Q.; Kinoshita, H.; Muta, Y.; Linares, J.F.; Kasashima, H.; et al. Hyaluronan driven by epithelial aPKC deficiency remodels the microenvironment and creates a vulnerability in mesenchymal colorectal cancer. Cancer Cell 2023, 41, 252–271.e9. [Google Scholar] [CrossRef] [PubMed]
- Spinelli, F.M.; Vitale, D.L.; Demarchi, G.; Cristina, C.; Alaniz, L. The immunological effect of hyaluronan in tumor angiogenesis. Clin. Transl. Immunol. 2015, 4, e52. [Google Scholar] [CrossRef]
- Pardue, E.L.; Ibrahim, S.; Ramamurthi, A. Role of hyaluronan in angiogenesis and its utility to angiogenic tissue engineering. Organogenesis 2008, 4, 203–214. [Google Scholar] [CrossRef]
- Ghose, S.; Biswas, S.; Datta, K.; Tyagi, R.K. Dynamic Hyaluronan drives liver endothelial cells towards angiogenesis. BMC Cancer 2018, 18, 648. [Google Scholar] [CrossRef]
- Olofsson, B.; Porsch, H.; Heldin, P. Knock-down of CD44 regulates endothelial cell differentiation via NFkappaB-mediated chemokine production. PLoS ONE 2014, 9, e90921. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Pearlman, E.; Diaconu, E.; Guo, K.; Mori, H.; Haqqi, T.; Markowitz, S.; Willson, J.; Sy, M.S. Expression of hyaluronidase by tumor cells induces angiogenesis in vivo. Proc. Natl. Acad. Sci. USA 1996, 93, 7832–7837. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Wallwiener, M.; Rudolph, A.; Cuk, K.; Eilber, U.; Celik, M.; Modugno, C.; Trumpp, A.; Heil, J.; Marme, F.; et al. Plasma hyaluronic acid level as a prognostic and monitoring marker of metastatic breast cancer. Int. J. Cancer 2016, 138, 2499–2509. [Google Scholar] [CrossRef]
- Zhang, H.; Tsang, J.Y.; Ni, Y.B.; Chan, S.K.; Chan, K.F.; Cheung, S.Y.; Tse, G.M. Hyaluronan synthase 2 is an adverse prognostic marker in androgen receptor-negative breast cancer. J. Clin. Pathol. 2016, 69, 1055–1062. [Google Scholar] [CrossRef] [PubMed]
- Morera, D.S.; Hennig, M.S.; Talukder, A.; Lokeshwar, S.D.; Wang, J.; Garcia-Roig, M.; Ortiz, N.; Yates, T.J.; Lopez, L.E.; Kallifatidis, G.; et al. Hyaluronic acid family in bladder cancer: Potential prognostic biomarkers and therapeutic targets. Br. J. Cancer 2017, 117, 1507–1517. [Google Scholar] [CrossRef] [PubMed]
- Llaneza, A.; Vizoso, F.; Rodriguez, J.C.; Raigoso, P.; Garcia-Muniz, J.L.; Allende, M.T.; Garcia-Moran, M. Hyaluronic acid as prognostic marker in resectable colorectal cancer. Br. J. Surg. 2000, 87, 1690–1696. [Google Scholar] [CrossRef] [PubMed]
- Passerotti, C.C.; Bonfim, A.; Martins, J.R.; Dall’Oglio, M.F.; Sampaio, L.O.; Mendes, A.; Ortiz, V.; Srougi, M.; Dietrich, C.P.; Nader, H.B. Urinary hyaluronan as a marker for the presence of residual transitional cell carcinoma of the urinary bladder. Eur. Urol. 2006, 49, 71–75. [Google Scholar] [CrossRef]
- Passerotti, C.C.; Srougi, M.; Bomfim, A.C.; Martins, J.R.; Leite, K.R.; Dos Reis, S.T.; Sampaio, L.O.; Ortiz, V.; Dietrich, C.P.; Nader, H.B. Testing for urinary hyaluronate improves detection and grading of transitional cell carcinoma. Urol. Oncol. 2011, 29, 710–715. [Google Scholar] [CrossRef]
- Jeronimo, S.M.; Sales, A.O.; Fernandes, M.Z.; Melo, F.P.; Sampaio, L.O.; Dietrich, C.P.; Nader, H.B. Glycosaminoglycan structure and content differ according to the origins of human tumors. Braz. J. Med. Biol. Res. 1994, 27, 2253–2258. [Google Scholar]
- Porsch, H.; Bernert, B.; Mehic, M.; Theocharis, A.D.; Heldin, C.H.; Heldin, P. Efficient TGFbeta-induced epithelial-mesenchymal transition depends on hyaluronan synthase HAS2. Oncogene 2013, 32, 4355–4365. [Google Scholar] [CrossRef]
- Zoltan-Jones, A.; Huang, L.; Ghatak, S.; Toole, B.P. Elevated hyaluronan production induces mesenchymal and transformed properties in epithelial cells. J. Biol. Chem. 2003, 278, 45801–45810. [Google Scholar] [CrossRef]
- Koyama, H.; Hibi, T.; Isogai, Z.; Yoneda, M.; Fujimori, M.; Amano, J.; Kawakubo, M.; Kannagi, R.; Kimata, K.; Taniguchi, S.; et al. Hyperproduction of hyaluronan in neu-induced mammary tumor accelerates angiogenesis through stromal cell recruitment: Possible involvement of versican/PG-M. Am. J. Pathol. 2007, 170, 1086–1099. [Google Scholar] [CrossRef] [PubMed]
- Witschen, P.M.; Chaffee, T.S.; Brady, N.J.; Huggins, D.N.; Knutson, T.P.; LaRue, R.S.; Munro, S.A.; Tiegs, L.; McCarthy, J.B.; Nelson, A.C.; et al. Tumor Cell Associated Hyaluronan-CD44 Signaling Promotes Pro-Tumor Inflammation in Breast Cancer. Cancers 2020, 12, 1325. [Google Scholar] [CrossRef] [PubMed]
- Misra, S.; Ghatak, S.; Toole, B. Regulation of MDR1 expression and drug resistance by a positive feedback loop involving hyaluronan, phosphoinositide 3-kinase, and ErbB2. J. Biol. Chem. 2005, 280, 20310–20315. [Google Scholar] [CrossRef]
- Bourguignon, L.Y.; Wong, G.; Earle, C.; Chen, L. Hyaluronan-CD44v3 interaction with Oct4-Sox2-Nanog promotes miR-302 expression leading to self-renewal, clonal formation, and cisplatin resistance in cancer stem cells from head and neck squamous cell carcinoma. J. Biol. Chem. 2012, 287, 32800–32824. [Google Scholar] [CrossRef] [PubMed]
- Skandalis, S.S.; Karalis, T.T.; Chatzopoulos, A.; Karamanos, N.K. Hyaluronan-CD44 axis orchestrates cancer stem cell functions. Cell Signal. 2019, 63, 109377. [Google Scholar] [CrossRef]
- Stern, R. Hyaluronidases in cancer biology. Semin. Cancer Biol. 2008, 18, 275–280. [Google Scholar] [CrossRef]
- McAtee, C.O.; Barycki, J.J.; Simpson, M.A. Emerging roles for hyaluronidase in cancer metastasis and therapy. Adv. Cancer Res. 2014, 123, 1–34. [Google Scholar]
- Wang, X.Y.; Tan, J.X.; Vasse, M.; Delpech, B.; Ren, G.S. Comparison of hyaluronidase expression, invasiveness and tubule formation promotion in ER (−) and ER (+) breast cancer cell lines in vitro. Chin. Med. J. 2009, 122, 1300–1304. [Google Scholar]
- Kovar, J.L.; Johnson, M.A.; Volcheck, W.M.; Chen, J.; Simpson, M.A. Hyaluronidase expression induces prostate tumor metastasis in an orthotopic mouse model. Am. J. Pathol. 2006, 169, 1415–1426. [Google Scholar] [CrossRef]
- Tian, X.; Azpurua, J.; Hine, C.; Vaidya, A.; Myakishev-Rempel, M.; Ablaeva, J.; Mao, Z.; Nevo, E.; Gorbunova, V.; Seluanov, A. High-molecular-mass hyaluronan mediates the cancer resistance of the naked mole rat. Nature 2013, 499, 346–349. [Google Scholar] [CrossRef] [PubMed]
- Takasugi, M.; Firsanov, D.; Tombline, G.; Ning, H.; Ablaeva, J.; Seluanov, A.; Gorbunova, V. Naked mole-rat very-high-molecular-mass hyaluronan exhibits superior cytoprotective properties. Nat. Commun. 2020, 11, 2376. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Tian, X.; Lu, J.Y.; Boit, K.; Ablaeva, J.; Zakusilo, F.T.; Emmrich, S.; Firsanov, D.; Rydkina, E.; Biashad, S.A.; et al. Increased hyaluronan by naked mole-rat Has2 improves healthspan in mice. Nature 2023, 621, 196–205. [Google Scholar] [CrossRef] [PubMed]
- Nikitovic, D.; Kouvidi, K.; Kavasi, R.M.; Berdiaki, A.; Tzanakakis, G.N. Hyaluronan/Hyaladherins—A Promising Axis for Targeted Drug Delivery in Cancer. Curr. Drug Deliv. 2016, 13, 500–511. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Qi, H.; Xu, J.; Guo, P.; Chen, F.; Li, F.; Yang, X.; Sheng, N.; Wu, Y.; Pan, W. Hyaluronan-based nanocarriers with CD44-overexpressed cancer cell targeting. Pharm. Res. 2014, 31, 2988–3005. [Google Scholar] [CrossRef] [PubMed]
- Vitale, D.L.; Icardi, A.; Rosales, P.; Spinelli, F.M.; Sevic, I.; Alaniz, L.D. Targeting the Tumor Extracellular Matrix by the Natural Molecule 4-Methylumbelliferone: A Complementary and Alternative Cancer Therapeutic Strategy. Front. Oncol. 2021, 11, 710061. [Google Scholar] [CrossRef] [PubMed]
- Heffler, M.; Golubovskaya, V.M.; Conroy, J.; Liu, S.; Wang, D.; Cance, W.G.; Dunn, K.B. FAK and HAS inhibition synergistically decrease colon cancer cell viability and affect expression of critical genes. Anticancer Agents Med. Chem. 2013, 13, 584–594. [Google Scholar] [CrossRef]
- Wang, T.P.; Pan, Y.R.; Fu, C.Y.; Chang, H.Y. Down-regulation of UDP-glucose dehydrogenase affects glycosaminoglycans synthesis and motility in HCT-8 colorectal carcinoma cells. Exp. Cell Res. 2010, 316, 2893–2902. [Google Scholar] [CrossRef]
- Malvicini, M.; Fiore, E.; Ghiaccio, V.; Piccioni, F.; Rizzo, M.; Olmedo Bonadeo, L.; Garcia, M.; Rodriguez, M.; Bayo, J.; Peixoto, E.; et al. Tumor Microenvironment Remodeling by 4-Methylumbelliferone Boosts the Antitumor Effect of Combined Immunotherapy in Murine Colorectal Carcinoma. Mol. Ther. 2015, 23, 1444–1455. [Google Scholar] [CrossRef]
- Cheng, X.B.; Sato, N.; Kohi, S.; Koga, A.; Hirata, K. 4-Methylumbelliferone inhibits enhanced hyaluronan synthesis and cell migration in pancreatic cancer cells in response to tumor-stromal interactions. Oncol. Lett. 2018, 15, 6297–6301. [Google Scholar] [CrossRef]
- Sato, N.; Cheng, X.B.; Kohi, S.; Koga, A.; Hirata, K. Targeting hyaluronan for the treatment of pancreatic ductal adenocarcinoma. Acta Pharm. Sin. B 2016, 6, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Nagase, H.; Kudo, D.; Suto, A.; Yoshida, E.; Suto, S.; Negishi, M.; Kakizaki, I.; Hakamada, K. 4-Methylumbelliferone Suppresses Hyaluronan Synthesis and Tumor Progression in SCID Mice Intra-abdominally Inoculated With Pancreatic Cancer Cells. Pancreas 2017, 46, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, H.; Yoshihara, S.; Kudo, D.; Morohashi, H.; Kakizaki, I.; Kon, A.; Takagaki, K.; Sasaki, M. 4-methylumbelliferone, a hyaluronan synthase suppressor, enhances the anticancer activity of gemcitabine in human pancreatic cancer cells. Cancer Chemother. Pharmacol. 2006, 57, 165–170. [Google Scholar] [CrossRef]
- Yoshida, E.; Kudo, D.; Nagase, H.; Shimoda, H.; Suto, S.; Negishi, M.; Kakizaki, I.; Endo, M.; Hakamada, K. Antitumor effects of the hyaluronan inhibitor 4-methylumbelliferone on pancreatic cancer. Oncol. Lett. 2016, 12, 2337–2344. [Google Scholar] [CrossRef] [PubMed]
- Lokeshwar, V.B.; Lopez, L.E.; Munoz, D.; Chi, A.; Shirodkar, S.P.; Lokeshwar, S.D.; Escudero, D.O.; Dhir, N.; Altman, N. Antitumor activity of hyaluronic acid synthesis inhibitor 4-methylumbelliferone in prostate cancer cells. Cancer Res. 2010, 70, 2613–2623. [Google Scholar] [CrossRef] [PubMed]
- Yates, T.J.; Lopez, L.E.; Lokeshwar, S.D.; Ortiz, N.; Kallifatidis, G.; Jordan, A.; Hoye, K.; Altman, N.; Lokeshwar, V.B. Dietary supplement 4-methylumbelliferone: An effective chemopreventive and therapeutic agent for prostate cancer. J. Natl. Cancer Inst. 2015, 107, djv085. [Google Scholar] [CrossRef]
- An, G.; Park, S.; Lee, M.; Lim, W.; Song, G. Antiproliferative Effect of 4-Methylumbelliferone in Epithelial Ovarian Cancer Cells Is Mediated by Disruption of Intracellular Homeostasis and Regulation of PI3K/AKT and MAPK Signaling. Pharmaceutics 2020, 12, 640. [Google Scholar] [CrossRef]
- Tamura, R.; Yokoyama, Y.; Yoshida, H.; Imaizumi, T.; Mizunuma, H. 4-Methylumbelliferone inhibits ovarian cancer growth by suppressing thymidine phosphorylase expression. J. Ovarian Res. 2014, 7, 94. [Google Scholar] [CrossRef]
- Urakawa, H.; Nishida, Y.; Wasa, J.; Arai, E.; Zhuo, L.; Kimata, K.; Kozawa, E.; Futamura, N.; Ishiguro, N. Inhibition of hyaluronan synthesis in breast cancer cells by 4-methylumbelliferone suppresses tumorigenicity in vitro and metastatic lesions of bone in vivo. Int. J. Cancer 2012, 130, 454–466. [Google Scholar] [CrossRef]
- Brett, M.E.; Bomberger, H.E.; Doak, G.R.; Price, M.A.; McCarthy, J.B.; Wood, D.K. In vitro elucidation of the role of pericellular matrix in metastatic extravasation and invasion of breast carcinoma cells. Integr. Biol. 2018, 10, 242–252. [Google Scholar] [CrossRef]
- Okuda, H.; Kobayashi, A.; Xia, B.; Watabe, M.; Pai, S.K.; Hirota, S.; Xing, F.; Liu, W.; Pandey, P.R.; Fukuda, K.; et al. Hyaluronan synthase HAS2 promotes tumor progression in bone by stimulating the interaction of breast cancer stem-like cells with macrophages and stromal cells. Cancer Res. 2012, 72, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Karalis, T.T.; Heldin, P.; Vynios, D.H.; Neill, T.; Buraschi, S.; Iozzo, R.V.; Karamanos, N.K.; Skandalis, S.S. Tumor-suppressive functions of 4-MU on breast cancer cells of different ER status: Regulation of hyaluronan/HAS2/CD44 and specific matrix effectors. Matrix Biol. 2019, 78–79, 118–138. [Google Scholar] [CrossRef] [PubMed]
- Twarock, S.; Freudenberger, T.; Poscher, E.; Dai, G.; Jannasch, K.; Dullin, C.; Alves, F.; Prenzel, K.; Knoefel, W.T.; Stoecklein, N.H.; et al. Inhibition of oesophageal squamous cell carcinoma progression by in vivo targeting of hyaluronan synthesis. Mol. Cancer 2011, 10, 30. [Google Scholar] [CrossRef] [PubMed]
- Rosser, J.I.; Nagy, N.; Goel, R.; Kaber, G.; Demirdjian, S.; Saxena, J.; Bollyky, J.B.; Frymoyer, A.R.; Pacheco-Navarro, A.E.; Burgener, E.B.; et al. Oral hymecromone decreases hyaluronan in human study participants. J. Clin. Investig. 2022, 132, e157983. [Google Scholar] [CrossRef] [PubMed]
- Abate, A.; Dimartino, V.; Spina, P.; Costa, P.L.; Lombardo, C.; Santini, A.; Del Piano, M.; Alimonti, P. Hymecromone in the treatment of motor disorders of the bile ducts: A multicenter, double-blind, placebo-controlled clinical study. Drugs Exp. Clin. Res. 2001, 27, 223–231. [Google Scholar] [PubMed]
- Karalis, T.T.; Chatzopoulos, A.; Kondyli, A.; Aletras, A.J.; Karamanos, N.K.; Heldin, P.; Skandalis, S.S. Salicylate suppresses the oncogenic hyaluronan network in metastatic breast cancer cells. Matrix Biol. Plus 2020, 6–7, 100031. [Google Scholar] [CrossRef] [PubMed]
- Karalis, T.; Shiau, A.K.; Gahman, T.C.; Skandalis, S.S.; Heldin, C.H.; Heldin, P. Identification of a Small Molecule Inhibitor of Hyaluronan Synthesis, DDIT, Targeting Breast Cancer Cells. Cancers 2022, 14, 5800. [Google Scholar] [CrossRef]
- Pibuel, M.A.; Poodts, D.; Diaz, M.; Molinari, Y.A.; Franco, P.G.; Hajos, S.E.; Lompardia, S.L. Antitumor effect of 4MU on glioblastoma cells is mediated by senescence induction and CD44, RHAMM and p-ERK modulation. Cell Death Discov. 2021, 7, 280. [Google Scholar] [CrossRef]
- Piccioni, F.; Fiore, E.; Bayo, J.; Atorrasagasti, C.; Peixoto, E.; Rizzo, M.; Malvicini, M.; Tirado-Gonzalez, I.; Garcia, M.G.; Alaniz, L.; et al. 4-methylumbelliferone inhibits hepatocellular carcinoma growth by decreasing IL-6 production and angiogenesis. Glycobiology 2015, 25, 825–835. [Google Scholar] [CrossRef]
- Kudo, D.; Kon, A.; Yoshihara, S.; Kakizaki, I.; Sasaki, M.; Endo, M.; Takagaki, K. Effect of a hyaluronan synthase suppressor, 4-methylumbelliferone, on B16F-10 melanoma cell adhesion and locomotion. Biochem. Biophys. Res. Commun. 2004, 321, 783–787. [Google Scholar] [CrossRef]
- Lompardia, S.L.; Diaz, M.; Papademetrio, D.L.; Pibuel, M.; Alvarez, E.; Hajos, S.E. 4-methylumbelliferone and imatinib combination enhances senescence induction in chronic myeloid leukemia cell lines. Investig. New Drugs 2017, 35, 1–10. [Google Scholar] [CrossRef]
- Arai, E.; Nishida, Y.; Wasa, J.; Urakawa, H.; Zhuo, L.; Kimata, K.; Kozawa, E.; Futamura, N.; Ishiguro, N. Inhibition of hyaluronan retention by 4-methylumbelliferone suppresses osteosarcoma cells in vitro and lung metastasis in vivo. Br. J. Cancer 2011, 105, 1839–1849. [Google Scholar] [CrossRef] [PubMed]
- Kretschmer, I.; Freudenberger, T.; Twarock, S.; Yamaguchi, Y.; Grandoch, M.; Fischer, J.W. Esophageal Squamous Cell Carcinoma Cells Modulate Chemokine Expression and Hyaluronan Synthesis in Fibroblasts. J. Biol. Chem. 2016, 291, 4091–4106. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, L.; Brown, T.J.; Heldin, P. Silencing of hyaluronan synthase 2 suppresses the malignant phenotype of invasive breast cancer cells. Int. J. Cancer 2007, 120, 2557–2567. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Askew, E.B.; Knudson, C.B.; Knudson, W. CRISPR/Cas9 knockout of HAS2 in rat chondrosarcoma chondrocytes demonstrates the requirement of hyaluronan for aggrecan retention. Matrix Biol. 2016, 56, 74–94. [Google Scholar] [CrossRef] [PubMed]
- Kolliopoulos, C.; Lin, C.Y.; Heldin, C.H.; Moustakas, A.; Heldin, P. Has2 natural antisense RNA and Hmga2 promote Has2 expression during TGFbeta-induced EMT in breast cancer. Matrix Biol. 2019, 80, 29–45. [Google Scholar] [CrossRef]
- Parnigoni, A.; Caon, I.; Teo, W.X.; Hua, S.H.; Moretto, P.; Bartolini, B.; Viola, M.; Karousou, E.; Yip, G.W.; Gotte, M.; et al. The natural antisense transcript HAS2-AS1 regulates breast cancer cells aggressiveness independently from hyaluronan metabolism. Matrix Biol. 2022, 109, 140–161. [Google Scholar] [CrossRef]
- Caon, I.; Bartolini, B.; Moretto, P.; Parnigoni, A.; Carava, E.; Vitale, D.L.; Alaniz, L.; Viola, M.; Karousou, E.; De Luca, G.; et al. Sirtuin 1 reduces hyaluronan synthase 2 expression by inhibiting nuclear translocation of NF-kappaB and expression of the long-noncoding RNA HAS2-AS1. J. Biol. Chem. 2020, 295, 3485–3496. [Google Scholar] [CrossRef]
- Gao, H.; Sun, X.; Rao, Y. PROTAC Technology: Opportunities and Challenges. ACS Med. Chem. Lett. 2020, 11, 237–240. [Google Scholar] [CrossRef]
- Burke, M.R.; Smith, A.R.; Zheng, G. Overcoming Cancer Drug Resistance Utilizing PROTAC Technology. Front. Cell Dev. Biol. 2022, 10, 872729. [Google Scholar] [CrossRef]
- Bekes, M.; Langley, D.R.; Crews, C.M. PROTAC targeted protein degraders: The past is prologue. Nat. Rev. Drug Discov. 2022, 21, 181–200. [Google Scholar] [CrossRef] [PubMed]
- Li, J.W.; Zheng, G.; Kaye, F.J.; Wu, L. PROTAC therapy as a new targeted therapy for lung cancer. Mol. Ther. 2023, 31, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Pu, W.; Zheng, Q.; Ai, M.; Chen, S.; Peng, Y. Proteolysis-targeting chimeras (PROTACs) in cancer therapy. Mol. Cancer 2022, 21, 99. [Google Scholar] [CrossRef] [PubMed]
- Cheng, W.; Li, S.; Wen, X.; Han, S.; Wang, S.; Wei, H.; Song, Z.; Wang, Y.; Tian, X.; Zhang, X. Development of hypoxia-activated PROTAC exerting a more potent effect in tumor hypoxia than in normoxia. Chem. Commun. 2021, 57, 12852–12855. [Google Scholar] [CrossRef]
- Ren, C.; Sun, N.; Liu, H.; Kong, Y.; Sun, R.; Qiu, X.; Chen, J.; Li, Y.; Zhang, J.; Zhou, Y.; et al. Discovery of a Brigatinib Degrader SIAIS164018 with Destroying Metastasis-Related Oncoproteins and a Reshuffling Kinome Profile. J. Med. Chem. 2021, 64, 9152–9165. [Google Scholar] [CrossRef]
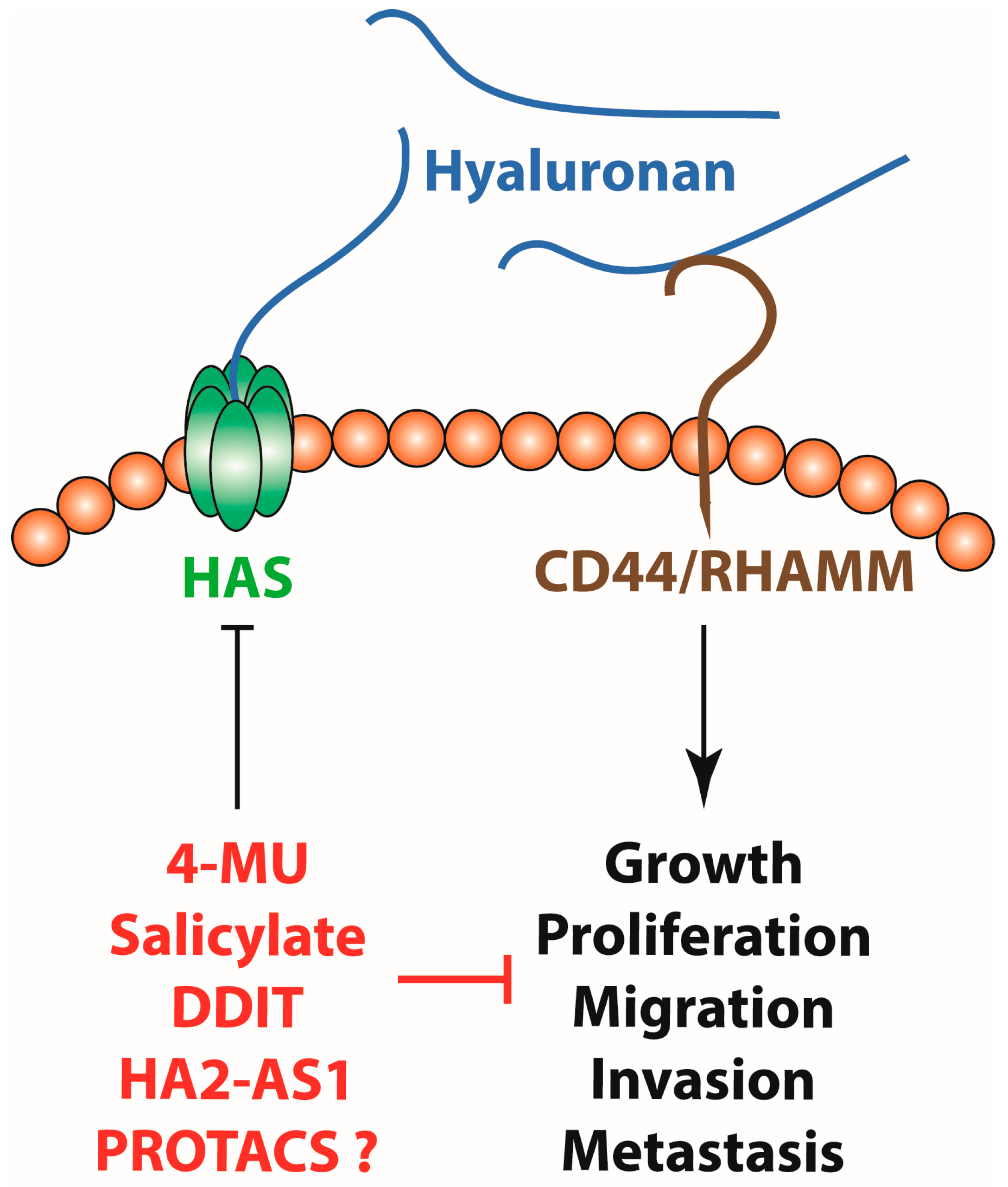
Figure 1.
Hyaluronan synthesis inhibition with small molecules like 4-MU, salicylate and DDIT, genetic manipulation through HAS2-AS1, as well as PROTACs might be of therapeutic value for the treatment of cancer by inhibiting tumour growth, proliferation, invasion and metastasis.
Figure 1.
Hyaluronan synthesis inhibition with small molecules like 4-MU, salicylate and DDIT, genetic manipulation through HAS2-AS1, as well as PROTACs might be of therapeutic value for the treatment of cancer by inhibiting tumour growth, proliferation, invasion and metastasis.


{kind=link}
Table 1.
Hyaluronan synthesis inhibitors and their effects in different tumour types.
| Inhibitor | Tumour Type | Effect | Reference |
|---|---|---|---|
| 4-methyl-umbelliferone (4-MU) | Colon | Inhibits proliferation and migration | [77,78,79] |
| Pancreatic | Improves survival rates and limits formation of metastases in the liver | [80,81,82,83,84] | |
| Prostate | Reduces tumour growth | [85,86] | |
| Gynaecological (ovarian and breast) | Attenuates cancer cell proliferation, migration and invasion | [87,88,89,90,91,92] | |
| Glioblastoma | Decreases cell migration and induces senescence | [98] | |
| Hepatocellular carcinoma | Inhibits angiogenesis and IL-6 production | [99] | |
| Melanoma | Reduces adhesion and locomotion | [100] | |
| Chronic myeloid leukaemia | Inhibits proliferation and induces senescence | [101] | |
| Osteosarcoma | Inhibits proliferation, migration, invasion and lung metastasis | [102] | |
| Oesophageal squamous cell carcinoma | Inhibits generation of cancer-associated fibroblasts | [103] | |
| Salicylate | Breast | Inhibits proliferation and migration | [96] |
| 5′-Deoxy-5′-(1,3-Diphenyl-2-Imidazolidinyl)-Thymidine (DDIT) | Breast | Reduces proliferation, migration, invasion and cancer stem cell self-renewal | [97] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Karalis, T. Targeting Hyaluronan Synthesis in Cancer: A Road Less Travelled. Biologics 2023, 3, 402-414. https://0-doi-org.brum.beds.ac.uk/10.3390/biologics3040022
AMA Style
Karalis T. Targeting Hyaluronan Synthesis in Cancer: A Road Less Travelled. Biologics. 2023; 3(4):402-414. https://0-doi-org.brum.beds.ac.uk/10.3390/biologics3040022
Chicago/Turabian StyleKaralis, Theodoros. 2023. "Targeting Hyaluronan Synthesis in Cancer: A Road Less Travelled" Biologics 3, no. 4: 402-414. https://0-doi-org.brum.beds.ac.uk/10.3390/biologics3040022