
Recent Advances in Arboviral Vaccines: Emerging Platforms and Promising Innovations

Department of Pharmacology, Physiology and Neuroscience, School of Medicine, University of South Carolina, Columbia, SC 29209, USA
Biologics 2024, 4(1), 1-16; https://0-doi-org.brum.beds.ac.uk/10.3390/biologics4010001
Submission received: 22 November 2023
/
Revised: 19 December 2023
/
Accepted: 20 December 2023
/
Published: 22 December 2023
(This article belongs to the Special Issue Novel Vaccine Technologies and Platforms to Protect from Infectious Diseases)
Abstract
:Arboviruses are a group of viruses that are transmitted by arthropods, such as mosquitoes, and cause significant morbidity and mortality worldwide. Currently, there are only a few options, with restricted use, for effective vaccines against these viruses. However, recent advances in arboviral vaccine development have shown promising innovations that have potential in preclinical and clinical studies. Insect-specific viruses have been explored as a novel vaccine platform that can induce cross-protective immunity against related arboviruses. Nanoparticle-based vaccines have also been developed to enhance the immunogenicity and stability of viral antigens. Additionally, vaccines against mosquito salivary proteins that can modulate the host immune response and interfere with arboviral transmission are being explored. Synonymous recoding, such as random codon shuffling, codon deoptimization, and codon-pair deoptimization, is being investigated as a strategy to attenuate the replication of arboviruses in vertebrate cells, reducing the risk of reverting to wild-type virulence. Finally, mRNA vaccines have been developed to rapidly generate and express viral antigens in the host cells, eliciting robust and durable immune responses. The challenges and opportunities for arboviral vaccine development are outlined, and future directions for research and innovation are discussed.
1. Introduction
Vaccines have significantly reduced the burden of infectious and non-infectious diseases, improving public health and extending life expectancy. As emphasized by the World Health Organization (WHO), immunization played a pivotal role in eradicating smallpox, eliminating poliomyelitis from most regions, and reducing measles incidence by 83% in the past two decades [1,2]. Vaccines continue to combat diseases such as diphtheria, tetanus, pertussis, influenza, meningitis, and rotavirus gastroenteritis, and most recently, they were rapidly developed to combat the coronavirus disease 2019 (COVID-19) pandemic. As per the latest data, more than 12.7 billion doses of COVID-19 vaccines have been administered globally, protecting millions of lives and livelihoods [3,4,5].
Despite remarkable progress in vaccine technology, challenges persist, particularly in addressing vector-borne diseases that constitute nearly 20% of the global disease burden [6]. Mosquito-borne viral pathogens pose a major global health threat, with over 100 viral pathogens transmitted by mosquitoes affecting almost half of the world’s population and with an economic impact estimated at USD 12 billion annually [7]. These effects are particularly severe in low socioeconomic regions, where treatment and related costs can represent up to 25% of household income, exacerbating already dire financial circumstances [8]. Arthropod-borne viral (arboviral) infections commonly manifest as fevers, headaches, arthralgia, and rashes but can also lead to the development of more serious complications like Guillain–Barré syndrome [9,10,11,12]. Moreover, arboviral infections can also cause long-term neurological impairments and cognitive disabilities [13].
In 2018, despite the available vaccines, 109,000 cases of yellow fever and 51,000 fatalities were reported [14]. Similarly, in 2019, around 57,000 cases of Japanese encephalitis resulted in 21,000 deaths [15]. Dengue fever alone affects nearly 4 billion people in 129 countries, with close to 100 million symptomatic cases and 40,000 deaths every year [16]. Although a dengue fever vaccine recently became available, it is limited to individuals with confirmed prior infection due to the high risk of severe dengue in the unexposed population [17,18,19].
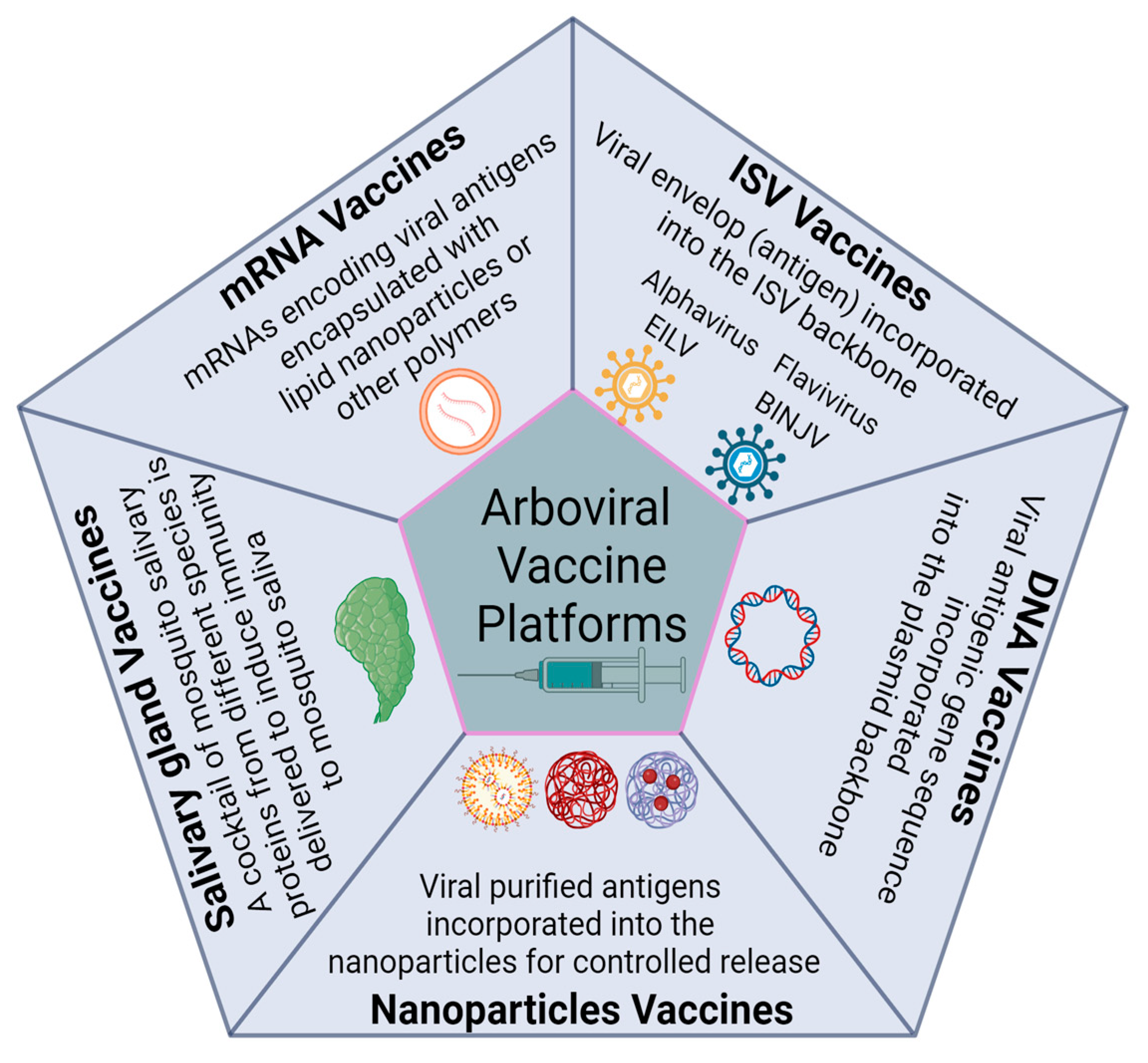
The persistent threat of arboviruses, most recently exemplified by the 2015–2016 zika outbreak, necessitates both innovative solutions and collaborative efforts to overcome these public health challenges [11,12,20]. By mimicking a natural infection, vaccines stimulate the immune system to generate antibodies and memory cells to combat viral, parasitic, and bacterial pathogens. Vaccine technologies span a broad range, from traditional whole-pathogen-inactivated and attenuated vaccines to novel designs based on nucleic acids, virus-like particles (VLPs), and conjugate vaccines. Each approach has advantages and disadvantages, reflecting the dynamic landscape of vaccine development. For instance, whole-pathogen vaccines that are either inactivated or attenuated can trigger strong immune responses but may pose safety issues. Conversely, engineered vaccines may have better safety profiles but might also induce less powerful immune reactions. In this article, we will explore the dynamic landscape of arboviral vaccines, focusing on the forefront of innovation. Notably, we will review the emerging platforms and promising advancements, including the utilization of insect-specific viruses as a vaccine platform, the development of nanoparticle-based vaccines, the exploration of mosquito salivary protein-based vaccines, and the groundbreaking field of mRNA vaccines (Figure 1).
2. Arboviral Vaccine Platforms
2.1. Insect-Specific Viruses Platform
Insect-specific viruses (ISVs) are a novel and promising platform for advancing recombinant vaccine development against various arboviral pathogens, such as dengue, zika, and chikungunya, owing to their distinctive attributes [21]. ISVs infect arthropod vectors such as mosquitoes and sandflies, but as these cannot replicate in vertebrate cells, their safety profile is superior to that of other vaccine platforms. Over 60 ISVs have been characterized so far [21]. Unlike attenuated vaccines that inherently carry the risk of virulence recurrence, ISVs are more suitable for people who have a compromised immune system, such as those who have HIV, cancer, or immunosuppressive therapy [22,23,24]. Moreover, the production of ISV-based vaccine candidates does not require chemical or physical inactivation processes, which can impair the immunogenicity and antigenicity of the vaccine candidate. By using recombinant DNA technology to generate chimeric constructs, ISV-based vaccines provide a promising opportunity for developing safer and more efficacious vaccine candidates against arboviruses and related pathogens. In the following sections, we discuss Eilat virus (EILV) and Binjari virus (BINJV), which are ISVs from alphavirus and flavivirus families, respectively, and how these have been used as platforms for designing vaccines for human alphaviruses and flaviviruses.
2.1.1. Eilat Virus (EILV)
The development of a vaccine against the chikungunya virus (CHIKV), a member of the Togaviridae family’s alphavirus genus, entailed inserting the genetic sequence encoding all three CHIKV structural proteins (E1, E2, and C) into the cDNA clone of the insect-specific alphavirus, EILV. Notably, both EILV and CHIKV belong to the alphavirus genus and share a similar genomic organization. In vitro, the replication-deficient ELIV/CHIKV chimeric vaccine candidates was capable of attachment, entry, and viral protein expression in vertebrate cells, similar to CHIKV.
In vaccination and CHIKV challenge experiments in mouse and non-human primate (NHP) models, the EILV/CHIKV chimeric vaccine achieved fast, robust, and long-lasting immunogenicity and conferred protection with just a single dose. The production of neutralizing antibodies was fast and durable, completely protecting the mice and preventing the onset of fever and viremia in NHPs. In contrast, inactivated EILV/CHIKV chimeras achieved reduced efficacy but were still immunogenic, as evidenced by reduced animal mortality without entirely alleviating disease symptoms in specific mouse models. Moreover, the vaccination induced elevated levels of CHIKV-specific IgM and IgG antibodies, memory B cell responses, and CD8+ T cell responses, all of which contribute to strong innate and adaptive immunity. In addition, the vaccine stimulated the production of antiviral cytokines, including IFNα, IFNβ, IL-1β, IL-6, and IL-12; inflammatory cytokines, such as IL-1β and IL-17; and antigen-presenting cells (APCs) in vivo [25].
The EILV platform was further used to create chimeric vaccines targeting other pathogens, such as Eastern equine encephalitis virus (EEEV) and Venezuelan equine encephalitis virus (VEEV), demonstrating broad applicability. Animals (murine challenge model) that received single doses of monovalent or multivalent EILV/EEEV and EILV/VEEV live chimeras produced neutralizing antibodies and survived when exposed to wild-type viruses more than 8 weeks post-vaccination [26]. Notably, mice vaccinated with a single dose of the monovalent vaccine exhibited 50% protection within six days post-vaccination [24,26].
2.1.2. Binjari Virus (BINJV)
BINJV represents a recently identified insect-specific flavivirus that demonstrates the potential to serve as an expeditious vaccine platform, featuring a production timeline of two to three weeks [27]. This attribute positions BINJV as a highly promising tool for rapid responses to emerging pathogenic flaviviruses such as dengue virus (DENV), yellow fever virus (YFV), zika virus (ZIKV), West Nile virus (WNV), and Japanese encephalitis virus (JEV).
The prME gene-encoding structural proteins of BINJV are substituted with the corresponding genes from arboviruses using a refined circular polymerase extension reaction [18]. The resultant chimeric viruses exhibit preservation of both the structural and immunological characteristics inherent in the parental arboviruses. Furthermore, these chimeric viruses display efficient replication within mosquito cells while demonstrating an incapacity to replicate within vertebrate cells [18].
Among the notable chimeric vaccines is BINJV/ZIKV-prME, designed to counteract the zika virus. Experimental trials involving IFNAR-/- mice subjected to single doses of either 2 or 20 µg of the vial preparation yielded a substantial immune response [28]. This immune response was marked by the production of ZIKV-specific IgG1 and IgG2c and total IgG and the development of neutralizing antibodies. Although the 20 µg vaccine elicited higher responses, both dosage levels effectively reduced viremia to undetectable levels upon subsequent ZIKV challenge [23,24].
In summary, leveraging insect-specific viruses, particularly in developing chimeric vaccines, holds significant promise for addressing arboviruses and related pathogens. This approach presents a robust and effective vaccination strategy with enhanced reliability and potency.
2.2. DNA Vaccine Platform
DNA vaccine platforms represent a pioneering advancement in immunological prophylactics, utilizing recombinant plasmid DNA to elicit antigen-specific immune responses. These platforms circumvent traditional antigen purification processes by transfecting host cells with plasmids encoding pathogen-specific antigenic epitopes, facilitating endogenous antigen synthesis and presentation. This strategy offers expeditious adaptability and scalability, which are crucial for addressing emergent pathogenic challenges. Additionally, the ambient thermal stability of DNA vaccines obviates the need for stringent cold-chain logistics [29]. Despite their potential, DNA vaccines are subject to ongoing, rigorous research to ascertain their immunogenicity, safety profile, and long-term efficacy. Research has focused on enhancing the immunogenicity of genomic vaccines through various strategies, including adjuvant and delivery systems, promoter selection, administration methods, and antigen codon optimization [29].
The recent progress marks a significant step forward in the development of flavivirus DNA vaccines, with several candidates for DENV, ZIKV, and WNV currently undergoing clinical trials and more in preclinical stages. The tetravalent DENV vaccine TVDVVAX, which includes the adjuvant Vaxfectin, has been well received in Phase I trials, indicating its safety and ability to induce significant T-cell responses [30]. At the same time, VRC5283 for ZIKV has shown promising immunogenicity in a Phase II trial [31]. For WNV, the DNA vaccine candidates VRC302 and VRC303, differing in their promoter use, have undergone Phase I trials, with VRC303 demonstrating safety and immunogenicity, though further follow-up studies are pending [32,33].
Lately, DNA vaccine candidates have been developed and evaluated for the zika virus. The CHIKV-envelope construct (pMCE321) integrates E3, E2, and E1 genes linked via furin cleavage sites and optimized for envelope expression through codon and RNA optimization. This DNA vaccine shows efficacy in both mice and nonhuman primates, providing protection in mice and driving neutralizing titers in primates similar to human convalescent sera. A second vaccine construct incorporates CHIKV capsid and envelope-specific sequences, a Kozak sequence, and an immunoglobulin E leader sequence [34,35]. These vaccines, optimized for envelope expression and using innovative immunization techniques like adaptive constant-current electroporation, have elicited strong T-cell and humoral responses, underscoring the potential of DNA vaccines to provide comprehensive immune protection against challenging viral pathogens.
Additionally, it should be highlighted that viruses depend on the host’s translational machinery and tRNA pool for protein synthesis; viral genomes tend to reflect the codon usage patterns of their hosts. Thus, codon pair bias, reflected as a preferential or rare occurrence of certain codon pairs, can impact translation efficiency and kinetics, which may, in turn, influence protein folding and the co-expression of proteins with related functions.
Synonymous recoding, such as random codon shuffling, codon deoptimization, and codon-pair deoptimization, attenuates the replication of most viruses, including arboviral human pathogens in both vertebrate and invertebrate cells [36,37,38]. For instance, changing codons to one mutation away from a stop codon can decrease virus replication in vitro and in vivo in mice and mosquitoes [37]. Notably, CpG-rich sequences are naturally underrepresented in vertebrate RNA. However, when these sequences are present in viral RNA, they activate the zinc-finger antiviral protein (ZAP). This activation, in turn, initiates the interferon response, mediates RNA degradation, and inhibits translation [38,39]. As alphaviruses naturally have higher CpG dinucleotide frequencies, resembling unbiased mosquito RNA, avoiding CpG and UpA dinucleotides is a primary driver of viral genome codon distribution. Conversely, introducing CpG and/or UpA dinucleotides through synonymous recoding attenuates the replication of diverse vertebrate viruses, including ZIKV [40,41,42]. The deletion of the gene encoding ZAP recovers the replication of CpG-high virus mutants [38]. Notably, mosquito RNA lacks CpG dinucleotide bias, and CpG-high ZIKV mutants replicate more efficiently than wild-type ZIKV in mosquito cells and salivary glands, suggesting that deoptimizing arboviruses for vertebrate cell replication can paradoxically enhance mosquito cell replication [42].
The protein structure of attenuated viruses is identical to their wild-type counterparts, ensuring a specific and robust immune response. Attenuation through recoding introduces numerous nucleotide substitutions, significantly reducing the risk of reverting to wild-type virulence. Several arboviral vaccine candidates are currently under evaluation [37,38,43,44,45,46]. However, investigations to assess the attenuation extent, potential tissue infections, virus transmission potential, and persistent mosquito infections will be critical components of vaccine development.
2.3. Nanoparticle-Based Platform
Nanoparticles (NPs) are organic or inorganic versatile immunization platforms that potently stimulate the immune system and achieve durable antigen presentation. NPs effectively function as adjuvants by encapsulating therapeutic agents in a biocompatible matrix, preventing their degradation and enhancing both uptake and overall immunogenicity [47,48,49]. Furthermore, they facilitate regulated antigen release, forming reservoirs at the administration site. NPs, with dimensions ranging from 10 to 1000 nm, have gained significance in bioscience due to their distinctive properties and potential for drug and antigen delivery systems, and they are promising vaccine candidates because they can encapsulate, adsorb, or disperse antigens [49].
The choice of materials for nanoparticle matrices will determine the kinetics of antigen release at the injection site and upon distribution. Biodegradable nanoparticle matrices can be made of either natural (e.g., proteins, carbohydrates) or synthetic biopolymers. While natural biomaterials offer benefits such as biocompatibility, biodegradability, and low immunogenicity, synthetic versions are typically easier to optimize [50,51,52]. To enhance nanoparticle functionality, a range of physical–chemical parameters such as size, charge, and morphology need to be carefully adjusted. Particle size determines in vivo distribution, toxicity, and cellular targeting, with smaller nanoparticles (<25 nm) demonstrating rapid transport through the lymphatic system and potential accumulation in dendritic cells [53]. Lymphatic drainage is also a mechanism that influences lipid nanoparticles (LNPs)’ interaction with B cells to promote immune responses, with particles in the 20–200 nm range being especially effective at stimulating immune cells [54]. Understanding how particle shape and size influence immune responses will be important to advance the development of next-generation vaccines.
Experimental arbovirus vaccines often employ LNPs because of their high mono-dispersion, long-term stability, and relatively good thermal stability. LNPs typically comprise four components: an ionizable cationic lipid for self-assembly and antigen release, PEG–lipid conjugates for prolonged formulation half-life, cholesterol for stabilization, and natural phospholipids for lipid bilayer structure support [55,56]. LNPs enhance protein stability, protect against proteolytic degradation, and exhibit low toxicity by avoiding potentially harmful organic solvents [57]. While dengue vaccines have used different organic (e.g., bovine serum albumin, lipids) and inorganic materials (e.g., calcium chloride, carbon, calcium phosphate), zika virus vaccines have predominantly used lipid and poly(amidoamine) nanoparticles, and Japanese encephalitis virus vaccines have included protein-VLPs, chitosan, and poly(gamma-glutamic acid) nanoparticles [58,59,60]. Conversely, West Nile virus vaccines have applied gold, CpG oligodeoxynucleotide, polyethyleneimine, lipoprotein, and lipid-based nanoparticle, whereas zika virus vaccines have used lipid and amyloid-based nanoparticles [61,62,63]. In addition, Hepatitis B virus protein-derived VLPs have been assessed for dengue and Japanese Encephalitis vaccines [64,65,66].
The use of nanoparticle-based vaccines to combat arbovirus infections is supported by strong reasoning; however, a notable weakness in this area lies in the lack of standardized methodologies across studies. Critical data related to nanoparticle formulation, physical–chemical characteristics (size, morphology, zeta potential, encapsulation or adsorption rates), and comprehensive in vitro and in vivo analyses were frequently omitted. Some studies overlooked investigations into the cytotoxicity of nanoparticle-based vaccines and their interactions with antigen-processing cells and other immune cells. While generally considered safe, it is crucial to explore the potential activation of key players in innate and adaptive immune responses, such as dendritic cells and T cells. Furthermore, a significant number of studies did not conduct challenge assays on nanoparticle-immunized mice, an essential step in assessing the protective efficacy of these vaccine formulations.
2.4. Mosquito Salivary Protein Vaccine
The mosquito salivary protein vaccine represents a novel approach, aiming to induce immunity to mosquito saliva rather than specific viral antigens. This innovative strategy targets the diverse molecules present in mosquito saliva that play a role in viral transmission and modulate the host’s immune response. The vaccine is rooted in the observation that mosquito saliva contains various molecules that facilitate viral transmission and infection by modulating the host’s immune response [67]. For instance, mosquito saliva can suppress the production of type I interferons, inhibit the activation of NK cells and macrophages, impair the function of dendritic cells and T cells, induce the production of anti-inflammatory cytokines, and increase vascular permeability, enhancing viral dissemination [68,69,70,71].
The pivotal role of CD4 and CD8 T cells, activated macrophages, and pDCs in the mosquito salivary protein vaccine is to mediate the adaptive immune response to mosquito saliva and enhance antiviral immunity [72]. CD4 T cells recognize mosquito salivary antigens presented by APCs via MHC class II molecules, assisting B cells in antibody production and CD8 T cells in cytotoxicity [73,74]. CD8 T cells, on the other hand, directly eliminate infected cells and release IL-10, IL-4, IL-13, and IFN-γ by recognizing mosquito salivary antigens presented by MHC class I molecules. pDCs produce type I interferons and other cytokines, activating NK cells and macrophages, inducing MHC class I expression on infected cells, and regulating the cytokine balance. Activated macrophages phagocytose and destroy pathogens while also presenting antigens to T cells and secreting inflammatory mediators [74,75,76].
Together, these immune cells collaborate to mount a robust and protective immune response against arboviruses upon a mosquito bite [73]. Immunizing with mosquito salivary proteins enables the host to generate antibodies that neutralize and prevent the immunosuppressive effects of these factors [77]. This approach could potentially result in a stronger and faster immune response upon a mosquito bite, ultimately reducing the viral load, disease severity, and transmission potential [67,76].
A single vaccine that is effective against multiple mosquito-borne diseases could have a profound impact on public health. The An. gambiae saliva (AGS-v) PLUS vaccine, a mosquito saliva vaccine, and an enhanced version of the first-generation AGS-v have demonstrated robust and specific immune responses across various formulations. AGS-v PLUS comprises five synthetic mosquito salivary peptides—four salivary peptides derived from An. gambiae salivary glands but common amongst Anopheles spp., Aedes spp., and Culex spp. ASG-v—and a fifth peptide. This fifth peptide is found in the saliva of many mosquito species, including An. gambiae, An. darling, Culex quinquefasciatus, A. aegypti, and A. albopictus, and aims to provoke an immune response that impedes the transmission of mosquito-borne pathogens [77]. Notably, the absence of adverse reactions such as redness or swelling post-mosquito bite indicates its favorable safety profile. The vaccine aims to provoke an immune response at mosquito bite sites to impede the transmission of mosquito-borne pathogens, such as the zika virus. Preclinical data in mice showed strong antibody responses induced by a fifth synthetic peptide, expanding the potential range of protection. More importantly, data from the phase-1 clinical trial are encouraging [77,78]. This vaccine candidate engages CD4 and CD8 T cells and activates macrophages and pDCs to counter the immunomodulatory effects of mosquito saliva, ultimately reducing the disease severity and transmission potential. These vaccines, administered with adjuvants (ISA-51), have shown safety, efficacy, and the potential to elicit a protective Th1 response at mosquito bite sites, which is crucial in preventing pathogen transmission [77].
The vaccine is being further developed for other mosquito-borne parasitic and viral pathogens and is undergoing Phase II clinical trials for zika by Imutex, a joint venture between PepTcell (trading as SEEK) and hVIVO. Moreover, while challenges persist, the AGS-v approach offers a promising avenue for addressing mosquito-borne diseases, providing a comprehensive and innovative solution to combat these pervasive health threats.
2.5. mRNA Vaccine
mRNA vaccines represent a powerful tool for developing diverse immunization methods. In this innovative approach, mRNA encoding a viral antigen is synthesized in vitro and administered into the host, leading to the expression of exogenous proteins and the induction of robust immune responses. The lab-made mRNA mimics the host mRNA, enhancing its stability and efficiency. Key features such as the 5′ cap-1, 5′, and 3′ untranslated elements and a poly(A)n tail protect the mRNA from exonucleases and facilitate antigen translation [79,80]. A short signal peptide at the N-terminus ensures the host cell secretes the proteins. Unmodified mRNA is susceptible to degradation by RNases, resulting in low expression. Adding nucleoside analogs (e.g., pseudouridine [Ψ]), purifying the lab-made mRNA, and encapsulating it in a lipid nanoparticle enhance cell uptake, leading to longer and higher expression [81,82]. These modifications increase mRNA stability and reduce immune detectability, preventing translation inhibition [83]. mRNA vaccine platforms enable the rapid development of vaccines, a critical capability in the face of emerging pandemics. Furthermore, the adaptable nature of the mRNA coding region allows for easy manipulation, facilitating the exploration of novel hypotheses in the pursuit of combating viral infections resistant to conventional vaccine strategies.
BioNTech, in collaboration with Pfizer and Fosun, developed and distributed the mRNA vaccine against COVID-19 [84]. The vaccine received regulatory approval from the UK in December 2020, followed by emergency use authorization from the FDA. Notably, the FDA granted full approval on 23 August 2021 [85].
The landscape of mRNA vaccine research is expanding, with several other candidates targeting diseases such as rabies, human cytomegalovirus, and influenza currently progressing through clinical trials [86,87,88,89]. Multiple mRNA vaccine candidates for arboviral pathogens transmitted by mosquitoes, leading to diseases like dengue, zika, chikungunya, and others, are at different stages of development and undergoing clinical trials, representing a spectrum of promising solutions [90,91,92,93,94,95,96]. These mRNA vaccines employ diverse strategies and various antigen targets, demonstrating the adaptability of this innovative approach. Table 1 provides an overview of the current progress in arboviral mRNA vaccine candidates and potential future breakthroughs in combating some of the important arboviral diseases.
3. Host Immune Response and Arboviral Vaccine Designing
The intricate immune mechanism in arboviral infections introduces an additional layer of complexity and challenges. Beyond directly neutralizing the virus, virus-specific antibodies play a protective role by engaging the Fc portion and binding to Fcγ receptors (FcγR) on immune cells [97]. This engagement triggers Antibody-Dependent Cellular Phagocytosis (ADCP), Antibody-Dependent Cellular Cytotoxicity (ADCC), and Complement-Dependent Cytotoxicity (CDC) [98]. While these processes are crucial for eliminating viruses and virus-infected cells, they also bring challenges, notably the potential for Antibody-Dependent Enhancement (ADE). ADE, observed in flaviviruses like dengue virus, involves non-neutralizing antibodies that could potentially facilitate the virus entry process, exacerbating the situation [99].
The four distinct serotypes of dengue virus (DENV1-4) give rise to serotype-specific neutralizing antibodies as well as cross-reacting non-neutralizing antibodies. This dual response facilitates ADCP, enables viruses to enter, and potentially exacerbates the disease pathology [99]. Both neutralizing and non-neutralizing antibodies exhibit cross-reactivity against a wide range of arboviral pathogens. For instance, dengue antibodies can cross-react with yellow fever and zika viruses. Also, antibodies from tick-borne encephalitis virus (TBEV) vaccine recipients may cross-react with the zika virus, posing challenges like difficulty engaging with virus-infected cells [100].
Moreover, in the context of ZIKV, the cellular expression of the envelope protein differs from that of the NS1 protein [101]. Unlike the NS1 protein, the ZIKV envelope protein, along with other flaviviruses, is not expressed at the cell surface [102]. Flaviviral particles emerge internally from the Golgi apparatus, and the structural proteins, including the envelope protein, are not readily accessible on the surfaces of infected cells [103]. Consequently, envelope-specific antibodies can bind to intact virions and trigger ADE, and these antibodies face limitations in binding to infected cells, preventing them from engaging protective Fc-mediated effector functions [104]. Though envelope-specific antibodies may provide sterilizing immunity against ZIKV, their protective efficacy may be limited due to the potential for ADE. In contrast, NS1-specific antibodies are emphasized for their efficacy in clearing virus-infected cells. This underscores the importance of identifying a suitable vaccine candidate.
Furthermore, recent research on potential monoclonal antibody treatment for the zika virus (CHIKV) emphasizes the crucial role of antibody-FcγR engagement in optimal infection clearance, mediating enhanced antibody effector mechanisms like ADCP [105]. The study, focused on CHIKV, underscores the significance of understanding antibody effector functions beyond direct virus neutralization. Virus-specific antibodies, particularly those engaging in Fc-mediated effector functions, are pivotal for protection. A comparative analysis of intact anti-CHIKV monoclonal antibodies with variants lacking significant binding to Fcγ receptors (FcγRs) and complement component 1q (C1q) reveals the importance of Fcγ receptor engagement in CHIKV clearance, highlighting enhanced antibody effector mechanisms such as ADCP. Antibodies from individuals with a natural CHIKV infection can inhibit viral egress and initiate ADCC through immune effector cells, emphasizing the necessity for protective FcγR-mediated antibody effector functions induced by CHIKV vaccination [106,107,108].
The research also evaluates the impact of Fc effector functions in a mouse model of CHIKV-induced arthritis, emphasizing the indispensable role of intact antibodies for therapeutic efficacy. Fc effector functions contribute to alleviating foot swelling, expediting viral RNA clearance, and curtailing immune cell infiltration, with monocytes identified as a key player in clinical protection and viral RNA clearance [108].
Hence, the formulation of tactics that target a balanced immune response to all serotypes to reduce Antibody-Dependent Enhancement (ADE) is a crucial element. This underscores the need for continuous research in the pursuit of designing effective vaccines.
4. Non-Vaccine Methods to Control Arboviruses
Over the years, arbovirus vector control using insecticide-based methods has been the most common way of preventing arboviral diseases. However, the alarming rates of insecticide resistance among mosquito populations stimulated new approaches through increasing understanding of insect biology and insect-pathogen interactions [109,110]. Vector control strategies are now focusing on the interference of arbovirus development in vectors and impacting insect survival, such as enhancing vector immune system, manipulating vector microbiome, or editing vector genomes using clustered regularly interspaced short palindromic repeats-Cas systems. These approaches are primarily based on either population suppression or modification strategies [111,112,113]. Suppression strategies include the sterile insect technique, incompatible insect technique, and transgene-based technologies. In population modification strategies, pathogen-resistant mosquitoes are designed to be released into wild populations, where they can spread their heritable modifications to prevent pathogen transmission. Wolbachia, a heritable insect endosymbiont, when introduced into the mosquito, can crash a mosquito population by cytoplasmic incompatibility or reduced likelihood of pathogen transmission by infected mosquitoes through competitive interaction with viruses such as dengue, zika, and other arboviruses [114,115].
The World Mosquito Program has initiated trials involving the release of Wolbachia-infected mosquitoes in various countries [116]. Remarkably, city-wide deployment of these modified mosquitoes in Colombia resulted in a substantial reduction in dengue incidence by 94–97% in areas where the insects were well established [117]. Several other countries, including Australia, Malaysia, Indonesia, Vietnam, and Brazil, have also released Wolbachia-infected mosquitoes to curb the transmission of local mosquito-borne disease [116]. During early mosquito breeding season, California releases Wolbachia-infected male mosquitoes to suppress the Aedes mosquito population [113,118]. The UK-based company Oxitec (Oxford, UK) involves the release of antibiotic-dependent OX5034 male mosquitoes in the UK and Florida, USA [119]. These genetically modified mosquitoes have self-limiting genes preventing female mosquito offspring from reaching adulthood. These multifaceted interventions showcase the evolving landscape of vector control methodologies in the pursuit of mitigating arboviral diseases. Despite the success of traditional strategies and the promise of recent developments, the persistence of vector control failures in several countries emphasizes the continued significance of vaccination as the most effective means of preventing arboviral pathogens.
5. Conclusions and Future Perspectives
The current landscape of mosquito-borne arboviral vaccines emphasizes the need for the continued development of vaccines with broader protective capabilities. Advanced technologies, such as the ones mentioned earlier, provide potential solutions for creating next-generation vaccines with better vaccine efficacy and safety parameters. Table 2 in the article specifically highlights these considerations, detailing the potential limitations of current vaccine technologies that require attention.
Vaccines for four mosquito-borne arboviral infections are available: yellow fever, zika, dengue, and Japanese encephalitis, but their use is restricted. The yellow fever (YF) vaccine is a live attenuated vaccine that has been in use since the 1930s, especially in YF endemic areas [120]. YF-Vax is available in the USA and is recommended for international travelers traveling to YF-endemic areas, as well as for laboratory personnel at risk of exposure to the virulent yellow fever virus or concentrated vaccine strains. Japanese encephalitis (JE) vaccines, including inactivated options like MB-JEV and ENCEVAC, as well as live-attenuated vaccines such as SA-14-14-2 and the YFV-JEV chimeric vaccine, play a crucial role in safeguarding individuals in endemic regions [121,122,123]. IXIARO, an inactivated JE vaccine available in the United States, is recommended for international travelers [124]. The dengue vaccine (Dengvaxia) is a tetravalent live attenuated vaccine derived from a yellow fever 17D viral strain. It is approved for individuals under 45 years of age (under 16 in the USA) with a history of dengue exposure [125]. Recently, the chikungunya vaccine (Ixchiq) achieved FDA approval through the accelerated approval pathway, marking a significant advance in chikungunya prevention [126]. However, concerns persist. While the YF vaccine, though rare, is associated with neurotropic and viscerotropic diseases, live attenuated JEV vaccines show rare but severe vaccine-related adverse events [127,128]. Dengvaxia’s efficacy is limited, particularly against DENV2, and it increases the risk of severe infection in seronegatively vaccinated individuals [125]. Addressing these concerns is crucial for ensuring the safety and effectiveness of these vaccines in combating arboviral infections.
With the surge in cases and potential virus emergence in new areas, the imperative for new and enhanced arboviral vaccines is evident. Further studies on ISV-based vaccines, including the characterization of human immune responses, are essential for advancing these promising tools into clinical development. The emergence and re-emergence of mosquito-borne diseases necessitate rapid preventive strategies, focusing on understanding vector saliva components to inform the development of salivary-based vaccines. Despite challenges, ongoing experiments and clinical trials showcase the potential of these vaccines, especially when complemented with viral antigens. Additionally, nucleic acid vaccines for DENV and ZIKV exhibit promising results, showcasing the adaptability of this platform, particularly in the context of the COVID-19 pandemic. An ideal vaccine targeting multiple flaviviruses through humoral and cellular immunity is a challenging but crucial goal, with mRNA vaccines emerging as a leading candidate in this endeavor.
The future of arboviral vaccine development holds exciting prospects and challenges. Advances in ISV-,DNA, mRNA-, and nanoparticle-based platforms pave the way for more effective and versatile vaccines against mosquito-borne diseases. Further research on conserved epitopes and the development of pan-flaviviral immunity are critical. The proposed Solidarity Vaccine Trial by the WHO reflects global efforts to address these challenges collectively. However, responsible research practices must be upheld, avoiding extreme measures for determining vaccine efficacy [129]. As compared to vector control, vaccines and medications remain powerful tools for halting the spread of infectious diseases. Continued collaboration, innovation, and ethical considerations will shape the future landscape of arboviral vaccine development, contributing to the global fight against mosquito-borne illnesses.
Funding
NIH grant R21AI151475 provided funding for this work to Sujit Pujhari.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Minta, A.A.; Ferrari, M.; Antoni, S.; Portnoy, A.; Sbarra, A.; Lambert, B.; Hauryski, S.; Hatcher, C.; Nedelec, Y.; Datta, D.; et al. Progress Toward Regional Measles Elimination—Worldwide, 2000–2021. MMWR Morb. Mortal Wkly. Rep. 2022, 71, 1489–1495. [Google Scholar] [CrossRef] [PubMed]
- Badizadegan, K.; Kalkowska, D.A.; Thompson, K.M. Polio by the Numbers-A Global Perspective. J. Infect. Dis. 2022, 226, 1309–1318. [Google Scholar] [CrossRef] [PubMed]
- Kayser, V.; Ramzan, I. Vaccines and vaccination: History and emerging issues. Hum. Vaccin. Immunother. 2021, 17, 5255–5268. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, E. Coronavirus (COVID-19) Vaccinations. Available online: https://ourworldindata.org/covid-vaccinations (accessed on 19 November 2023).
- Mathieu, E.; Ritchie, H.; Ortiz-Ospina, E.; Roser, M.; Hasell, J.; Appel, C.; Giattino, C.; Rodes-Guirao, L. A global database of COVID-19 vaccinations. Nat. Hum. Behav. 2021, 5, 947–953. [Google Scholar] [CrossRef] [PubMed]
- Torto, B.; Tchouassi, D.P. Grand Challenges in Vector-Borne Disease Control Targeting Vectors. Front. Trop. Dis. 2021, 1, 635356. [Google Scholar] [CrossRef]
- Aryaprema, V.S.; Steck, M.R.; Peper, S.T.; Xue, R.D.; Qualls, W.A. A systematic review of published literature on mosquito control action thresholds across the world. PLoS Negl. Trop. Dis. 2023, 17, e0011173. [Google Scholar] [CrossRef]
- Greenwood, B. Treatment of malaria--a continuing challenge. N. Engl. J. Med. 2014, 371, 474–475. [Google Scholar] [CrossRef]
- Ladzinski, A.T.; Tai, A.; Rumschlag, M.T.; Smith, C.S.; Mehta, A.; Boapimp, P.; Edewaard, E.J.; Douce, R.W.; Morgan, L.F.; Wang, M.S.; et al. Clinical Characteristics of the 2019 Eastern Equine Encephalitis Outbreak in Michigan. Open Forum Infect. Dis. 2023, 10, ofad206. [Google Scholar] [CrossRef]
- Pujhari, S.K.; Prabhakar, S.; Ratho, R.K.; Modi, M.; Sharma, M.; Mishra, B. A novel mutation (S227T) in domain II of the envelope gene of Japanese encephalitis virus circulating in North India. Epidemiol. Infect. 2011, 139, 849–856. [Google Scholar] [CrossRef]
- Pujhari, S.; Brustolin, M.; Macias, V.M.; Nissly, R.H.; Nomura, M.; Kuchipudi, S.V.; Rasgon, J.L. Heat shock protein 70 (Hsp70) mediates Zika virus entry, replication, and egress from host cells. Emerg. Microbes Infect. 2019, 8, 8–16. [Google Scholar] [CrossRef]
- Dodson, B.L.; Pujhari, S.; Rasgon, J.L. Vector competence of selected North American Anopheles and Culex mosquitoes for Zika virus. PeerJ 2018, 6, e4324. [Google Scholar] [CrossRef]
- Pujhari, S.K.; Prabhakar, S.; Ratho, R.; Mishra, B.; Modi, M.; Sharma, S.; Singh, P. Th1 immune response takeover among patients with severe Japanese encephalitis infection. J. Neuroimmunol. 2013, 263, 133–138. [Google Scholar] [CrossRef]
- Gaythorpe, K.A.; Hamlet, A.; Jean, K.; Garkauskas Ramos, D.; Cibrelus, L.; Garske, T.; Ferguson, N. The global burden of yellow fever. Elife 2021, 10, 64670. [Google Scholar] [CrossRef]
- Moore, S.M. The current burden of Japanese encephalitis and the estimated impacts of vaccination: Combining estimates of the spatial distribution and transmission intensity of a zoonotic pathogen. PLoS Negl. Trop. Dis. 2021, 15, e0009385. [Google Scholar] [CrossRef]
- Roy, S.K.; Bhattacharjee, S. Dengue virus: Epidemiology, biology, and disease aetiology. Can. J. Microbiol. 2021, 67, 687–702. [Google Scholar] [CrossRef]
- Torresi, J.; Ebert, G.; Pellegrini, M. Vaccines licensed and in clinical trials for the prevention of dengue. Hum. Vaccin. Immunother. 2017, 13, 1059–1072. [Google Scholar] [CrossRef]
- Osorio, J.E.; Wallace, D.; Stinchcomb, D.T. A recombinant, chimeric tetravalent dengue vaccine candidate based on a dengue virus serotype 2 backbone. Expert. Rev. Vaccines 2016, 15, 497–508. [Google Scholar] [CrossRef]
- Freedman, D.O. A new dengue vaccine (TAK-003) now WHO recommended in endemic areas; what about travelers? J. Travel. Med. 2023, 30, taad132. [Google Scholar] [CrossRef]
- Clancy, I.L.; Jones, R.T.; Power, G.M.; Logan, J.G.; Iriart, J.A.B.; Massad, E.; Kinsman, J. Public health messages on arboviruses transmitted by Aedes aegypti in Brazil. BMC Public Health 2021, 21, 1362. [Google Scholar] [CrossRef]
- Gomez, M.; Martinez, D.; Munoz, M.; Ramirez, J.D. Aedes aegypti and Ae. albopictus microbiome/virome: New strategies for controlling arboviral transmission? Parasit. Vectors 2022, 15, 287. [Google Scholar] [CrossRef]
- Tanelus, M.; Lopez, K.; Smith, S.; Muller, J.A.; Porier, D.L.; Auguste, D.I.; Stone, W.B.; Paulson, S.L.; Auguste, A.J. Exploring the immunogenicity of an insect-specific virus vectored Zika vaccine candidate. Sci. Rep. 2023, 13, 19948. [Google Scholar] [CrossRef]
- Hobson-Peters, J.; Harrison, J.J.; Watterson, D.; Hazlewood, J.E.; Vet, L.J.; Newton, N.D.; Warrilow, D.; Colmant, A.M.G.; Taylor, C.; Huang, B.; et al. A recombinant platform for flavivirus vaccines and diagnostics using chimeras of a new insect-specific virus. Sci. Transl. Med. 2019, 11, eaax7888. [Google Scholar] [CrossRef]
- Carvalho, V.L.; Long, M.T. Perspectives on New Vaccines against Arboviruses Using Insect-Specific Viruses as Platforms. Vaccines 2021, 9, 263. [Google Scholar] [CrossRef]
- Adam, A.; Luo, H.; Osman, S.R.; Wang, B.; Roundy, C.M.; Auguste, A.J.; Plante, K.S.; Peng, B.H.; Thangamani, S.; Frolova, E.I.; et al. Optimized production and immunogenicity of an insect virus-based chikungunya virus candidate vaccine in cell culture and animal models. Emerg. Microbes Infect. 2021, 10, 305–316. [Google Scholar] [CrossRef]
- Erasmus, J.H.; Seymour, R.L.; Kaelber, J.T.; Kim, D.Y.; Leal, G.; Sherman, M.B.; Frolov, I.; Chiu, W.; Weaver, S.C.; Nasar, F. Novel Insect-Specific Eilat Virus-Based Chimeric Vaccine Candidates Provide Durable, Mono- and Multivalent, Single-Dose Protection against Lethal Alphavirus Challenge. J. Virol. 2018, 92, e01274-17. [Google Scholar] [CrossRef]
- Habarugira, G.; Harrison, J.J.; Moran, J.; Suen, W.W.; Colmant, A.M.G.; Hobson-Peters, J.; Isberg, S.R.; Bielefeldt-Ohmann, H.; Hall, R.A. A chimeric vaccine protects farmed saltwater crocodiles from West Nile virus-induced skin lesions. npj Vaccines 2023, 8, 93. [Google Scholar] [CrossRef]
- Hazlewood, J.E.; Tang, B.; Yan, K.; Rawle, D.J.; Harrison, J.J.; Hall, R.A.; Hobson-Peters, J.; Suhrbier, A. The Chimeric Binjari-Zika Vaccine Provides Long-Term Protection against ZIKA Virus Challenge. Vaccines 2022, 10, 85. [Google Scholar] [CrossRef]
- Pagliari, S.; Dema, B.; Sanchez-Martinez, A.; Montalvo Zurbia-Flores, G.; Rollier, C.S. DNA Vaccines: History, Molecular Mechanisms and Future Perspectives. J. Mol. Biol. 2023, 435, 168297. [Google Scholar] [CrossRef]
- Danko, J.R.; Kochel, T.; Teneza-Mora, N.; Luke, T.C.; Raviprakash, K.; Sun, P.; Simmons, M.; Moon, J.E.; De La Barrera, R.; Martinez, L.J.; et al. Safety and Immunogenicity of a Tetravalent Dengue DNA Vaccine Administered with a Cationic Lipid-Based Adjuvant in a Phase 1 Clinical Trial. Am. J. Trop. Med. Hyg. 2018, 98, 849–856. [Google Scholar] [CrossRef]
- Dowd, K.A.; Ko, S.Y.; Morabito, K.M.; Yang, E.S.; Pelc, R.S.; DeMaso, C.R.; Castilho, L.R.; Abbink, P.; Boyd, M.; Nityanandam, R.; et al. Rapid development of a DNA vaccine for Zika virus. Science 2016, 354, 237–240. [Google Scholar] [CrossRef]
- Ledgerwood, J.E.; Pierson, T.C.; Hubka, S.A.; Desai, N.; Rucker, S.; Gordon, I.J.; Enama, M.E.; Nelson, S.; Nason, M.; Gu, W.; et al. A West Nile virus DNA vaccine utilizing a modified promoter induces neutralizing antibody in younger and older healthy adults in a phase I clinical trial. J. Infect. Dis. 2011, 203, 1396–1404. [Google Scholar] [CrossRef]
- Dutta, S.K.; Langenburg, T. A Perspective on Current Flavivirus Vaccine Development: A Brief Review. Viruses 2023, 15, 860. [Google Scholar] [CrossRef]
- Muthumani, K.; Lankaraman, K.M.; Laddy, D.J.; Sundaram, S.G.; Chung, C.W.; Sako, E.; Wu, L.; Khan, A.; Sardesai, N.; Kim, J.J.; et al. Immunogenicity of novel consensus-based DNA vaccines against Chikungunya virus. Vaccine 2008, 26, 5128–5134. [Google Scholar] [CrossRef]
- Mallilankaraman, K.; Shedlock, D.J.; Bao, H.; Kawalekar, O.U.; Fagone, P.; Ramanathan, A.A.; Ferraro, B.; Stabenow, J.; Vijayachari, P.; Sundaram, S.G.; et al. A DNA vaccine against chikungunya virus is protective in mice and induces neutralizing antibodies in mice and nonhuman primates. PLoS Negl. Trop. Dis. 2011, 5, e928. [Google Scholar] [CrossRef]
- Brule, C.E.; Grayhack, E.J. Synonymous Codons: Choose Wisely for Expression. Trends Genet. 2017, 33, 283–297. [Google Scholar] [CrossRef]
- Carrau, L.; Rezelj, V.V.; Noval, M.G.; Levi, L.I.; Megrian, D.; Blanc, H.; Weger-Lucarelli, J.; Moratorio, G.; Stapleford, K.A.; Vignuzzi, M. Chikungunya Virus Vaccine Candidates with Decreased Mutational Robustness Are Attenuated In Vivo and Have Compromised Transmissibility. J. Virol. 2019, 93, e00775-19. [Google Scholar] [CrossRef]
- van Bree, J.W.M.; Visser, I.; Duyvestyn, J.M.; Aguilar-Bretones, M.; Marshall, E.M.; van Hemert, M.J.; Pijlman, G.P.; van Nierop, G.P.; Kikkert, M.; Rockx, B.H.G.; et al. Novel approaches for the rapid development of rationally designed arbovirus vaccines. One Health 2023, 16, 100565. [Google Scholar] [CrossRef]
- Shaw, A.E.; Rihn, S.J.; Mollentze, N.; Wickenhagen, A.; Stewart, D.G.; Orton, R.J.; Kuchi, S.; Bakshi, S.; Collados, M.R.; Turnbull, M.L.; et al. The antiviral state has shaped the CpG composition of the vertebrate interferome to avoid self-targeting. PLoS Biol. 2021, 19, e3001352. [Google Scholar] [CrossRef]
- Fros, J.J.; Dietrich, I.; Alshaikhahmed, K.; Passchier, T.C.; Evans, D.J.; Simmonds, P. CpG and UpA dinucleotides in both coding and non-coding regions of echovirus 7 inhibit replication initiation post-entry. Elife 2017, 6, e29112. [Google Scholar] [CrossRef]
- Sharp, C.P.; Thompson, B.H.; Nash, T.J.; Diebold, O.; Pinto, R.M.; Thorley, L.; Lin, Y.T.; Sives, S.; Wise, H.; Clohisey Hendry, S.; et al. CpG dinucleotide enrichment in the influenza A virus genome as a live attenuated vaccine development strategy. PLoS Pathog. 2023, 19, e1011357. [Google Scholar] [CrossRef]
- Trus, I.; Udenze, D.; Karniychuk, U. Generation of CpG-Recoded Zika Virus Vaccine Candidates. Methods Mol. Biol. 2022, 2410, 289–302. [Google Scholar]
- Manokaran, G.; Sujatmoko; McPherson, K.G.; Simmons, C.P. Attenuation of a dengue virus replicon by codon deoptimization of nonstructural genes. Vaccine 2019, 37, 2857–2863. [Google Scholar] [CrossRef]
- Li, P.; Ke, X.; Wang, T.; Tan, Z.; Luo, D.; Miao, Y.; Sun, J.; Zhang, Y.; Liu, Y.; Hu, Q.; et al. Zika Virus Attenuation by Codon Pair Deoptimization Induces Sterilizing Immunity in Mouse Models. J. Virol. 2018, 92, e00701-18. [Google Scholar] [CrossRef]
- Nougairede, A.; De Fabritus, L.; Aubry, F.; Gould, E.A.; Holmes, E.C.; de Lamballerie, X. Random codon re-encoding induces stable reduction of replicative fitness of Chikungunya virus in primate and mosquito cells. PLoS Pathog. 2013, 9, e1003172. [Google Scholar] [CrossRef]
- de Fabritus, L.; Nougairede, A.; Aubry, F.; Gould, E.A.; de Lamballerie, X. Attenuation of tick-borne encephalitis virus using large-scale random codon re-encoding. PLoS Pathog. 2015, 11, e1004738. [Google Scholar] [CrossRef]
- Jia, F.; Liu, X.; Li, L.; Mallapragada, S.; Narasimhan, B.; Wang, Q. Multifunctional nanoparticles for targeted delivery of immune activating and cancer therapeutic agents. J. Control Release 2013, 172, 1020–1034. [Google Scholar] [CrossRef]
- Priyanka; Abusalah, M.A.H.; Chopra, H.; Sharma, A.; Mustafa, S.A.; Choudhary, O.P.; Sharma, M.; Dhawan, M.; Khosla, R.; Loshali, A.; et al. Nanovaccines: A game changing approach in the fight against infectious diseases. Biomed. Pharmacother. 2023, 167, 115597. [Google Scholar]
- Lv, T.; Meng, Y.; Liu, Y.; Han, Y.; Xin, H.; Peng, X.; Huang, J. RNA nanotechnology: A new chapter in targeted therapy. Colloids Surf. B Biointerfaces 2023, 230, 113533. [Google Scholar] [CrossRef]
- Kucuk, N.; Primozic, M.; Knez, Z.; Leitgeb, M. Sustainable Biodegradable Biopolymer-Based Nanoparticles for Healthcare Applications. Int. J. Mol. Sci. 2023, 24, 3188. [Google Scholar] [CrossRef]
- Idrees, H.; Zaidi, S.Z.J.; Sabir, A.; Khan, R.U.; Zhang, X.; Hassan, S.U. A Review of Biodegradable Natural Polymer-Based Nanoparticles for Drug Delivery Applications. Nanomaterials 2020, 10, 1970. [Google Scholar] [CrossRef]
- Gamage, A.; Thiviya, P.; Mani, S.; Ponnusamy, P.G.; Manamperi, A.; Evon, P.; Merah, O.; Madhujith, T. Environmental Properties and Applications of Biodegradable Starch-Based Nanocomposites. Polymers 2022, 14, 4578. [Google Scholar] [CrossRef]
- Kulkarni, S.A.; Feng, S.S. Effects of particle size and surface modification on cellular uptake and biodistribution of polymeric nanoparticles for drug delivery. Pharm. Res. 2013, 30, 2512–2522. [Google Scholar] [CrossRef]
- Goode, E.F.; Roussos Torres, E.T.; Irshad, S. Lymph Node Immune Profiles as Predictive Biomarkers for Immune Checkpoint Inhibitor Response. Front. Mol. Biosci. 2021, 8, 674558. [Google Scholar] [CrossRef]
- Eygeris, Y.; Gupta, M.; Kim, J.; Sahay, G. Chemistry of Lipid Nanoparticles for RNA Delivery. Acc. Chem. Res. 2022, 55, 2–12. [Google Scholar] [CrossRef]
- Fan, Y.; Marioli, M.; Zhang, K. Analytical characterization of liposomes and other lipid nanoparticles for drug delivery. J. Pharm. Biomed. Anal. 2021, 192, 113642. [Google Scholar] [CrossRef]
- Lin, P.C.; Lin, S.; Wang, P.C.; Sridhar, R. Techniques for physicochemical characterization of nanomaterials. Biotechnol. Adv. 2014, 32, 711–726. [Google Scholar] [CrossRef]
- Okamoto, S.; Yoshii, H.; Matsuura, M.; Kojima, A.; Ishikawa, T.; Akagi, T.; Akashi, M.; Takahashi, M.; Yamanishi, K.; Mori, Y. Poly-gamma-glutamic acid nanoparticles and aluminum adjuvant used as an adjuvant with a single dose of Japanese encephalitis virus-like particles provide effective protection from Japanese encephalitis virus. Clin. Vaccine Immunol. 2012, 19, 17–22. [Google Scholar] [CrossRef]
- de Souza, G.A.P.; Rocha, R.P.; Goncalves, R.L.; Ferreira, C.S.; de Mello Silva, B.; de Castro, R.F.G.; Rodrigues, J.F.V.; Junior, J.; Malaquias, L.C.C.; Abrahao, J.S.; et al. Nanoparticles as Vaccines to Prevent Arbovirus Infection: A Long Road Ahead. Pathogens 2021, 10, 36. [Google Scholar] [CrossRef]
- Nevagi, R.J.; Khalil, Z.G.; Hussein, W.M.; Powell, J.; Batzloff, M.R.; Capon, R.J.; Good, M.F.; Skwarczynski, M.; Toth, I. Polyglutamic acid-trimethyl chitosan-based intranasal peptide nano-vaccine induces potent immune responses against group A streptococcus. Acta Biomater. 2018, 80, 278–287. [Google Scholar] [CrossRef]
- Hossain, M.U.; Keya, C.A.; Das, K.C.; Hashem, A.; Omar, T.M.; Khan, M.A.; Rakib-Uz-Zaman, S.M.; Salimullah, M. An Immunopharmacoinformatics Approach in Development of Vaccine and Drug Candidates for West Nile Virus. Front. Chem. 2018, 6, 246. [Google Scholar] [CrossRef]
- Kose, N.; Fox, J.M.; Sapparapu, G.; Bombardi, R.; Tennekoon, R.N.; de Silva, A.D.; Elbashir, S.M.; Theisen, M.A.; Humphris-Narayanan, E.; Ciaramella, G.; et al. A lipid-encapsulated mRNA encoding a potently neutralizing human monoclonal antibody protects against chikungunya infection. Sci. Immunol. 2019, 4, aaw6647. [Google Scholar] [CrossRef]
- De Filette, M.; Soehle, S.; Ulbert, S.; Richner, J.; Diamond, M.S.; Sinigaglia, A.; Barzon, L.; Roels, S.; Lisziewicz, J.; Lorincz, O.; et al. Vaccination of mice using the West Nile virus E-protein in a DNA prime-protein boost strategy stimulates cell-mediated immunity and protects mice against a lethal challenge. PLoS ONE 2014, 9, e87837. [Google Scholar] [CrossRef]
- Nooraei, S.; Bahrulolum, H.; Hoseini, Z.S.; Katalani, C.; Hajizade, A.; Easton, A.J.; Ahmadian, G. Virus-like particles: Preparation, immunogenicity and their roles as nanovaccines and drug nanocarriers. J. Nanobiotechnol. 2021, 19, 59. [Google Scholar] [CrossRef]
- Krol, E.; Brzuska, G.; Szewczyk, B. Production and Biomedical Application of Flavivirus-like Particles. Trends Biotechnol. 2019, 37, 1202–1216. [Google Scholar] [CrossRef]
- Urakami, A.; Ngwe Tun, M.M.; Moi, M.L.; Sakurai, A.; Ishikawa, M.; Kuno, S.; Ueno, R.; Morita, K.; Akahata, W. An Envelope-Modified Tetravalent Dengue Virus-Like-Particle Vaccine Has Implications for Flavivirus Vaccine Design. J. Virol. 2017, 91, 10–1128. [Google Scholar] [CrossRef]
- Guerrero, D.; Cantaert, T.; Misse, D. Aedes Mosquito Salivary Components and Their Effect on the Immune Response to Arboviruses. Front. Cell Infect. Microbiol. 2020, 10, 407. [Google Scholar] [CrossRef]
- Wichit, S.; Diop, F.; Hamel, R.; Talignani, L.; Ferraris, P.; Cornelie, S.; Liegeois, F.; Thomas, F.; Yssel, H.; Misse, D. Aedes Aegypti saliva enhances chikungunya virus replication in human skin fibroblasts via inhibition of the type I interferon signaling pathway. Infect. Genet. Evol. 2017, 55, 68–70. [Google Scholar] [CrossRef]
- Schneider, B.S.; Soong, L.; Coffey, L.L.; Stevenson, H.L.; McGee, C.E.; Higgs, S. Aedes aegypti saliva alters leukocyte recruitment and cytokine signaling by antigen-presenting cells during West Nile virus infection. PLoS ONE 2010, 5, e11704. [Google Scholar] [CrossRef]
- Schneider, B.S.; Soong, L.; Zeidner, N.S.; Higgs, S. Aedes aegypti salivary gland extracts modulate anti-viral and TH1/TH2 cytokine responses to sindbis virus infection. Viral Immunol. 2004, 17, 565–573. [Google Scholar] [CrossRef]
- Lefteri, D.A.; Bryden, S.R.; Pingen, M.; Terry, S.; McCafferty, A.; Beswick, E.F.; Georgiev, G.; Van der Laan, M.; Mastrullo, V.; Campagnolo, P.; et al. Mosquito saliva enhances virus infection through sialokinin-dependent vascular leakage. Proc. Natl. Acad. Sci. USA 2022, 119, e2114309119. [Google Scholar] [CrossRef]
- Fong, S.W.; Kini, R.M.; Ng, L.F.P. Mosquito Saliva Reshapes Alphavirus Infection and Immunopathogenesis. J. Virol. 2018, 92, e01004-17. [Google Scholar] [CrossRef]
- Vogt, M.B.; Lahon, A.; Arya, R.P.; Kneubehl, A.R.; Spencer Clinton, J.L.; Paust, S.; Rico-Hesse, R. Mosquito saliva alone has profound effects on the human immune system. PLoS Negl. Trop. Dis. 2018, 12, e0006439. [Google Scholar] [CrossRef]
- Guerrero, D.; Vo, H.T.M.; Lon, C.; Bohl, J.A.; Nhik, S.; Chea, S.; Man, S.; Sreng, S.; Pacheco, A.R.; Ly, S.; et al. Evaluation of cutaneous immune response in a controlled human in vivo model of mosquito bites. Nat. Commun. 2022, 13, 7036. [Google Scholar] [CrossRef]
- Demeure, C.E.; Brahimi, K.; Hacini, F.; Marchand, F.; Peronet, R.; Huerre, M.; St-Mezard, P.; Nicolas, J.F.; Brey, P.; Delespesse, G.; et al. Anopheles mosquito bites activate cutaneous mast cells leading to a local inflammatory response and lymph node hyperplasia. J. Immunol. 2005, 174, 3932–3940. [Google Scholar] [CrossRef]
- Depinay, N.; Hacini, F.; Beghdadi, W.; Peronet, R.; Mecheri, S. Mast cell-dependent down-regulation of antigen-specific immune responses by mosquito bites. J. Immunol. 2006, 176, 4141–4146. [Google Scholar] [CrossRef]
- Manning, J.E.; Oliveira, F.; Coutinho-Abreu, I.V.; Herbert, S.; Meneses, C.; Kamhawi, S.; Baus, H.A.; Han, A.; Czajkowski, L.; Rosas, L.A.; et al. Safety and immunogenicity of a mosquito saliva peptide-based vaccine: A randomised, placebo-controlled, double-blind, phase 1 trial. Lancet 2020, 395, 1998–2007. [Google Scholar] [CrossRef]
- Friedman-Klabanoff, D.J.; Birkhold, M.; Short, M.T.; Wilson, T.R.; Meneses, C.R.; Lacsina, J.R.; Oliveira, F.; Kamhawi, S.; Valenzuela, J.G.; Hunsberger, S.; et al. Safety and immunogenicity of AGS-v PLUS, a mosquito saliva peptide vaccine against arboviral diseases: A randomized, double-blind, placebo-controlled Phase 1 trial. EBioMedicine 2022, 86, 104375. [Google Scholar] [CrossRef]
- Ramanathan, A.; Robb, G.B.; Chan, S.H. mRNA capping: Biological functions and applications. Nucleic Acids Res. 2016, 44, 7511–7526. [Google Scholar] [CrossRef]
- Thillier, Y.; Decroly, E.; Morvan, F.; Canard, B.; Vasseur, J.J.; Debart, F. Synthesis of 5’ cap-0 and cap-1 RNAs using solid-phase chemistry coupled with enzymatic methylation by human (guanine-N(7))-methyl transferase. RNA 2012, 18, 856–868. [Google Scholar] [CrossRef]
- Kariko, K.; Muramatsu, H.; Welsh, F.A.; Ludwig, J.; Kato, H.; Akira, S.; Weissman, D. Incorporation of pseudouridine into mRNA yields superior nonimmunogenic vector with increased translational capacity and biological stability. Mol. Ther. 2008, 16, 1833–1840. [Google Scholar] [CrossRef]
- Reichmuth, A.M.; Oberli, M.A.; Jaklenec, A.; Langer, R.; Blankschtein, D. mRNA vaccine delivery using lipid nanoparticles. Ther. Deliv. 2016, 7, 319–334. [Google Scholar] [CrossRef]
- Maruggi, G.; Zhang, C.; Li, J.; Ulmer, J.B.; Yu, D. mRNA as a Transformative Technology for Vaccine Development to Control Infectious Diseases. Mol. Ther. 2019, 27, 757–772. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Perez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Lamb, Y.N. BNT162b2 mRNA COVID-19 Vaccine: First Approval. Drugs 2021, 81, 495–501. [Google Scholar] [CrossRef]
- Schnee, M.; Vogel, A.B.; Voss, D.; Petsch, B.; Baumhof, P.; Kramps, T.; Stitz, L. An mRNA Vaccine Encoding Rabies Virus Glycoprotein Induces Protection against Lethal Infection in Mice and Correlates of Protection in Adult and Newborn Pigs. PLoS Negl. Trop. Dis. 2016, 10, e0004746. [Google Scholar] [CrossRef]
- Alberer, M.; Gnad-Vogt, U.; Hong, H.S.; Mehr, K.T.; Backert, L.; Finak, G.; Gottardo, R.; Bica, M.A.; Garofano, A.; Koch, S.D.; et al. Safety and immunogenicity of a mRNA rabies vaccine in healthy adults: An open-label, non-randomised, prospective, first-in-human phase 1 clinical trial. Lancet 2017, 390, 1511–1520. [Google Scholar] [CrossRef]
- Nelson, C.S.; Jenks, J.A.; Pardi, N.; Goodwin, M.; Roark, H.; Edwards, W.; McLellan, J.S.; Pollara, J.; Weissman, D.; Permar, S.R. Human Cytomegalovirus Glycoprotein B Nucleoside-Modified mRNA Vaccine Elicits Antibody Responses with Greater Durability and Breadth than MF59-Adjuvanted gB Protein Immunization. J. Virol. 2020, 94, 10–1128. [Google Scholar] [CrossRef]
- Villanueva, M.T. An mRNA universal vaccine for influenza. Nat. Rev. Drug Discov. 2023, 22, 98. [Google Scholar] [CrossRef]
- Roth, C.; Cantaert, T.; Colas, C.; Prot, M.; Casademont, I.; Levillayer, L.; Thalmensi, J.; Langlade-Demoyen, P.; Gerke, C.; Bahl, K.; et al. A Modified mRNA Vaccine Targeting Immunodominant NS Epitopes Protects Against Dengue Virus Infection in HLA Class I Transgenic Mice. Front. Immunol. 2019, 10, 1424. [Google Scholar] [CrossRef]
- Shaw, C.A.; August, A.; Bart, S.; Booth, P.J.; Knightly, C.; Brasel, T.; Weaver, S.C.; Zhou, H.; Panther, L. A phase 1, randomized, placebo-controlled, dose-ranging study to evaluate the safety and immunogenicity of an mRNA-based chikungunya virus vaccine in healthy adults. Vaccine 2023, 41, 3898–3906. [Google Scholar] [CrossRef]
- Medina-Magues, L.G.; Gergen, J.; Jasny, E.; Petsch, B.; Lopera-Madrid, J.; Medina-Magues, E.S.; Salas-Quinchucua, C.; Osorio, J.E. mRNA Vaccine Protects against Zika Virus. Vaccines 2021, 9, 1464. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Pelc, R.S.; Muramatsu, H.; Andersen, H.; DeMaso, C.R.; Dowd, K.A.; Sutherland, L.L.; Scearce, R.M.; Parks, R.; et al. Zika virus protection by a single low-dose nucleoside-modified mRNA vaccination. Nature 2017, 543, 248–251. [Google Scholar] [CrossRef]
- Essink, B.; Chu, L.; Seger, W.; Barranco, E.; Le Cam, N.; Bennett, H.; Faughnan, V.; Pajon, R.; Paila, Y.D.; Bollman, B.; et al. The safety and immunogenicity of two Zika virus mRNA vaccine candidates in healthy flavivirus baseline seropositive and seronegative adults: The results of two randomised, placebo-controlled, dose-ranging, phase 1 clinical trials. Lancet Infect. Dis. 2023, 23, 621–633. [Google Scholar] [CrossRef]
- Wollner, C.J.; Richner, M.; Hassert, M.A.; Pinto, A.K.; Brien, J.D.; Richner, J.M. A Dengue Virus Serotype 1 mRNA-LNP Vaccine Elicits Protective Immune Responses. J. Virol. 2021, 95, 10–1128. [Google Scholar] [CrossRef]
- Taylor, N.P. Moderna Scraps Lead mRNA Chikungunya Candidate after Phase 1, Slowing Push Beyond Prophylactic Vaccines. Available online: https://www.fiercebiotech.com/biotech/moderna-scraps-lead-mrna-antibody-candidate-after-phase-1-slowing-push-beyond-prophylactic (accessed on 21 November 2023).
- Forthal, D.N.; Moog, C. Fc receptor-mediated antiviral antibodies. Curr. Opin. HIV AIDS 2009, 4, 388–393. [Google Scholar] [CrossRef]
- Sawant, J.; Patil, A.; Kurle, S. A Review: Understanding Molecular Mechanisms of Antibody-Dependent Enhancement in Viral Infections. Vaccines 2023, 11, 1240. [Google Scholar] [CrossRef]
- Katzelnick, L.C.; Gresh, L.; Halloran, M.E.; Mercado, J.C.; Kuan, G.; Gordon, A.; Balmaseda, A.; Harris, E. Antibody-dependent enhancement of severe dengue disease in humans. Science 2017, 358, 929–932. [Google Scholar] [CrossRef]
- Endale, A.; Medhin, G.; Darfiro, K.; Kebede, N.; Legesse, M. Magnitude of Antibody Cross-Reactivity in Medically Important Mosquito-Borne Flaviviruses: A Systematic Review. Infect. Drug Resist. 2021, 14, 4291–4299. [Google Scholar] [CrossRef]
- Brown, W.C.; Akey, D.L.; Konwerski, J.R.; Tarrasch, J.T.; Skiniotis, G.; Kuhn, R.J.; Smith, J.L. Extended surface for membrane association in Zika virus NS1 structure. Nat. Struct. Mol. Biol. 2016, 23, 865–867. [Google Scholar] [CrossRef]
- Akey, D.L.; Brown, W.C.; Dutta, S.; Konwerski, J.; Jose, J.; Jurkiw, T.J.; DelProposto, J.; Ogata, C.M.; Skiniotis, G.; Kuhn, R.J.; et al. Flavivirus NS1 structures reveal surfaces for associations with membranes and the immune system. Science 2014, 343, 881–885. [Google Scholar] [CrossRef] [PubMed]
- Barnard, T.R.; Abram, Q.H.; Lin, Q.F.; Wang, A.B.; Sagan, S.M. Molecular Determinants of Flavivirus Virion Assembly. Trends Biochem. Sci. 2021, 46, 378–390. [Google Scholar] [CrossRef] [PubMed]
- Bailey, M.J.; Broecker, F.; Duehr, J.; Arumemi, F.; Krammer, F.; Palese, P.; Tan, G.S. Antibodies Elicited by an NS1-Based Vaccine Protect Mice against Zika Virus. mBio 2019, 10, e02861-18. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.M.; Roy, V.; Gunn, B.M.; Huang, L.; Edeling, M.A.; Mack, M.; Fremont, D.H.; Doranz, B.J.; Johnson, S.; Alter, G.; et al. Optimal therapeutic activity of monoclonal antibodies against chikungunya virus requires Fc-FcgammaR interaction on monocytes. Sci. Immunol. 2019, 4, eaav5062. [Google Scholar] [CrossRef] [PubMed]
- Fric, J.; Bertin-Maghit, S.; Wang, C.I.; Nardin, A.; Warter, L. Use of human monoclonal antibodies to treat Chikungunya virus infection. J. Infect. Dis. 2013, 207, 319–322. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Liss, N.M.; Chen, D.H.; Liao, M.; Fox, J.M.; Shimak, R.M.; Fong, R.H.; Chafets, D.; Bakkour, S.; Keating, S.; et al. Neutralizing Monoclonal Antibodies Block Chikungunya Virus Entry and Release by Targeting an Epitope Critical to Viral Pathogenesis. Cell Rep. 2015, 13, 2553–2564. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.M.; Roy, V.; Gunn, B.M.; Bolton, G.R.; Fremont, D.H.; Alter, G.; Diamond, M.S.; Boesch, A.W. Enhancing the therapeutic activity of hyperimmune IgG against chikungunya virus using FcgammaRIIIa affinity chromatography. Front. Immunol. 2023, 14, 1153108. [Google Scholar] [CrossRef] [PubMed]
- Achee, N.L.; Grieco, J.P.; Vatandoost, H.; Seixas, G.; Pinto, J.; Ching-Ng, L.; Martins, A.J.; Juntarajumnong, W.; Corbel, V.; Gouagna, C.; et al. Alternative strategies for mosquito-borne arbovirus control. PLoS Negl. Trop. Dis. 2019, 13, e0006822. [Google Scholar]
- Dusfour, I.; Vontas, J.; David, J.P.; Weetman, D.; Fonseca, D.M.; Corbel, V.; Raghavendra, K.; Coulibaly, M.B.; Martins, A.J.; Kasai, S.; et al. Management of insecticide resistance in the major Aedes vectors of arboviruses: Advances and challenges. PLoS Negl. Trop. Dis. 2019, 13, e0007615. [Google Scholar] [CrossRef]
- Rasgon, J. Population replacement strategies for controlling vector populations and the use of Wolbachia pipientis for genetic drive. J. Vis. Exp. 2007, 5, 225. [Google Scholar]
- Rasgon, J.L. Dengue fever: Mosquitoes attacked from within. Nature 2011, 476, 407–408. [Google Scholar] [CrossRef]
- Liu, Y.; Yu, J.; Li, J. A Mosquito Population Suppression Model by Releasing Wolbachia-Infected Males. Bull. Math. Biol. 2022, 84, 121. [Google Scholar] [CrossRef]
- Weng, S.C.; Masri, R.A.; Akbari, O.S. Advances and challenges in synthetic biology for mosquito control. Trends Parasitol. 2023. [Google Scholar] [CrossRef]
- Li, M.; Yang, T.; Bui, M.; Gamez, S.; Wise, T.; Kandul, N.P.; Liu, J.; Alcantara, L.; Lee, H.; Edula, J.R.; et al. Suppressing mosquito populations with precision guided sterile males. Nat. Commun. 2021, 12, 5374. [Google Scholar] [CrossRef]
- Lenharo, M. Dengue is spreading. Can new vaccines and antivirals halt its rise? Nature 2023, 623, 470. [Google Scholar] [CrossRef]
- Velez, I.D.; Tanamas, S.K.; Arbelaez, M.P.; Kutcher, S.C.; Duque, S.L.; Uribe, A.; Zuluaga, L.; Martinez, L.; Patino, A.C.; Barajas, J.; et al. Reduced dengue incidence following city-wide wMel Wolbachia mosquito releases throughout three Colombian cities: Interrupted time series analysis and a prospective case-control study. PLoS Negl. Trop. Dis. 2023, 17, e0011713. [Google Scholar] [CrossRef]
- CDC Mosquitoes with Wolbachia for Reducing Numbers of Aedes aegypti Mosquitoes. Available online: https://www.cdc.gov/mosquitoes/mosquito-control/community/emerging-methods/wolbachia.html (accessed on 17 December 2023).
- Meghani, Z. Regulation of genetically engineered (GE) mosquitoes as a public health tool: A public health ethics analysis. Global Health 2022, 18, 21. [Google Scholar] [CrossRef]
- Barrett, A.D.T. Yellow fever live attenuated vaccine: A very successful live attenuated vaccine but still we have problems controlling the disease. Vaccine 2017, 35, 5951–5955. [Google Scholar] [CrossRef]
- Hoke, C.H.; Nisalak, A.; Sangawhipa, N.; Jatanasen, S.; Laorakapongse, T.; Innis, B.L.; Kotchasenee, S.; Gingrich, J.B.; Latendresse, J.; Fukai, K.; et al. Protection against Japanese encephalitis by inactivated vaccines. N. Engl. J. Med. 1988, 319, 608–614. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.I.; Lee, Y.M. Japanese encephalitis: The virus and vaccines. Hum. Vaccin. Immunother. 2014, 10, 263–279. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Wang, Z.; Yu, Y. Protection of SA14-14-2 live attenuated Japanese encephalitis vaccine against the wild-type JE viruses. Chin. Med. J. 2003, 116, 941–943. [Google Scholar] [PubMed]
- Erra, E.O.; Kantele, A. The Vero cell-derived, inactivated, SA14-14-2 strain-based vaccine (Ixiaro) for prevention of Japanese encephalitis. Expert. Rev. Vaccines 2015, 14, 1167–1179. [Google Scholar] [CrossRef] [PubMed]
- Aguiar, M.; Stollenwerk, N.; Halstead, S.B. The risks behind Dengvaxia recommendation. Lancet Infect. Dis. 2016, 16, 882–883. [Google Scholar] [CrossRef] [PubMed]
- FDA FDA Approves First Vaccine to Prevent Disease Caused by Chikungunya Virus. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-first-vaccine-prevent-disease-caused-chikungunya-virus (accessed on 21 November 2023).
- Ma, H.Y.; Lai, C.C.; Chiu, N.C.; Lee, P.I. Adverse events following immunization with the live-attenuated recombinant Japanese encephalitis vaccine (IMOJEV(R)) in Taiwan, 2017–2018. Vaccine 2020, 38, 5219–5222. [Google Scholar] [CrossRef] [PubMed]
- Kitchener, S. Viscerotropic and neurotropic disease following vaccination with the 17D yellow fever vaccine, ARILVAX. Vaccine 2004, 22, 2103–2105. [Google Scholar] [CrossRef]
- Callaway, E. Scores of coronavirus vaccines are in competition—How will scientists choose the best? Nature 2020. [Google Scholar] [CrossRef]
Figure 1.
Overview of the vaccine platforms targeting arboviruses, as explored in the article.


{kind=link}
Table 1.
mRNA vaccines for some of the important arboviral pathogens in different stages of development.
Table 1.
mRNA vaccines for some of the important arboviral pathogens in different stages of development.
| Vaccine Name | Target Virus | Encoded Proteins | Nucleoside Modification | Delivery Method | Animal Model | Outcome | Reference |
|---|---|---|---|---|---|---|---|
| mRNA-1325 | ZIKV | prM/E | No | LNP | Mice | Complete protection, reduction in viral titers and fetal resorption | [94] |
| Nucleoside-modified RNA vaccine | ZIKV | prM/ENV | Yes | LNP | Macaques | Potent neutralizing antibodies, protection, prevention of congenital transmission and fetal abnormalities | [93] |
| mRNA-LNP | ZIKV | prM/E | No | LNP | Mice, Macaques | Host-protective antibodies, sterilizing immunity, prevention of vertical transmission | [92] |
| DENV1-NS | DENV | NS3, NS4B, NS5 | No | LNP | Mice | Potent T cell response, reduction in DENV1 infection | [90] |
| prM/E mRNA-LNP | DENV | prM/E | Yes | LNP | Mice | Neutralizing antibodies, CD4+ and CD8+ T cells, increased ADE levels | [95] |
| mRNA-1944 | CHIKV | CHKV-24 monoclonal antibody | Yes | LNP | Mice | Potent neutralizing antibody, prevention of viremia | [96] |
| mRNA-1388 | CHIKV | prM/E | Yes | LNP | Mice, Macaques | Potent neutralizing antibody, protection against challenge | [91] |
Table 2.
Possible constraints of the vaccine technologies.
| Vaccine Platform | Limitations |
|---|---|
| Insect-specific viruses (ISVs) | Safety and efficacy require further research. |
| DNA vaccine | Limited immune response in humans. Delivery issues, difficulties in effective DNA delivery to host cells. Potential stability issues and genomic integration risks. Risk of triggering anti-DNA antibodies, causing autoimmune reactions. Challenging regulatory approval. |
| Nanoparticle-based vaccines | Low stability, structural heterogeneity, potential immunogenicity, high toxicity, and off-target activity. Challenges in efficient delivery to target sites. |
| Salivary protein vaccines | Ongoing research to understand biology. Vaccines derived from one mosquito species’ saliva might not be effective against different mosquito species. The approach will be ineffective for arboviral infections acquired via sexual or blood transfusion. Potential issues with allergic responses or negative effects from exposure to saliva proteins. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Pujhari, S. Recent Advances in Arboviral Vaccines: Emerging Platforms and Promising Innovations. Biologics 2024, 4, 1-16. https://0-doi-org.brum.beds.ac.uk/10.3390/biologics4010001
AMA Style
Pujhari S. Recent Advances in Arboviral Vaccines: Emerging Platforms and Promising Innovations. Biologics. 2024; 4(1):1-16. https://0-doi-org.brum.beds.ac.uk/10.3390/biologics4010001
Chicago/Turabian StylePujhari, Sujit. 2024. "Recent Advances in Arboviral Vaccines: Emerging Platforms and Promising Innovations" Biologics 4, no. 1: 1-16. https://0-doi-org.brum.beds.ac.uk/10.3390/biologics4010001