Brain Susceptibility to Methyl Donor Deficiency: From Fetal Programming to Aging Outcome in Rats


 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
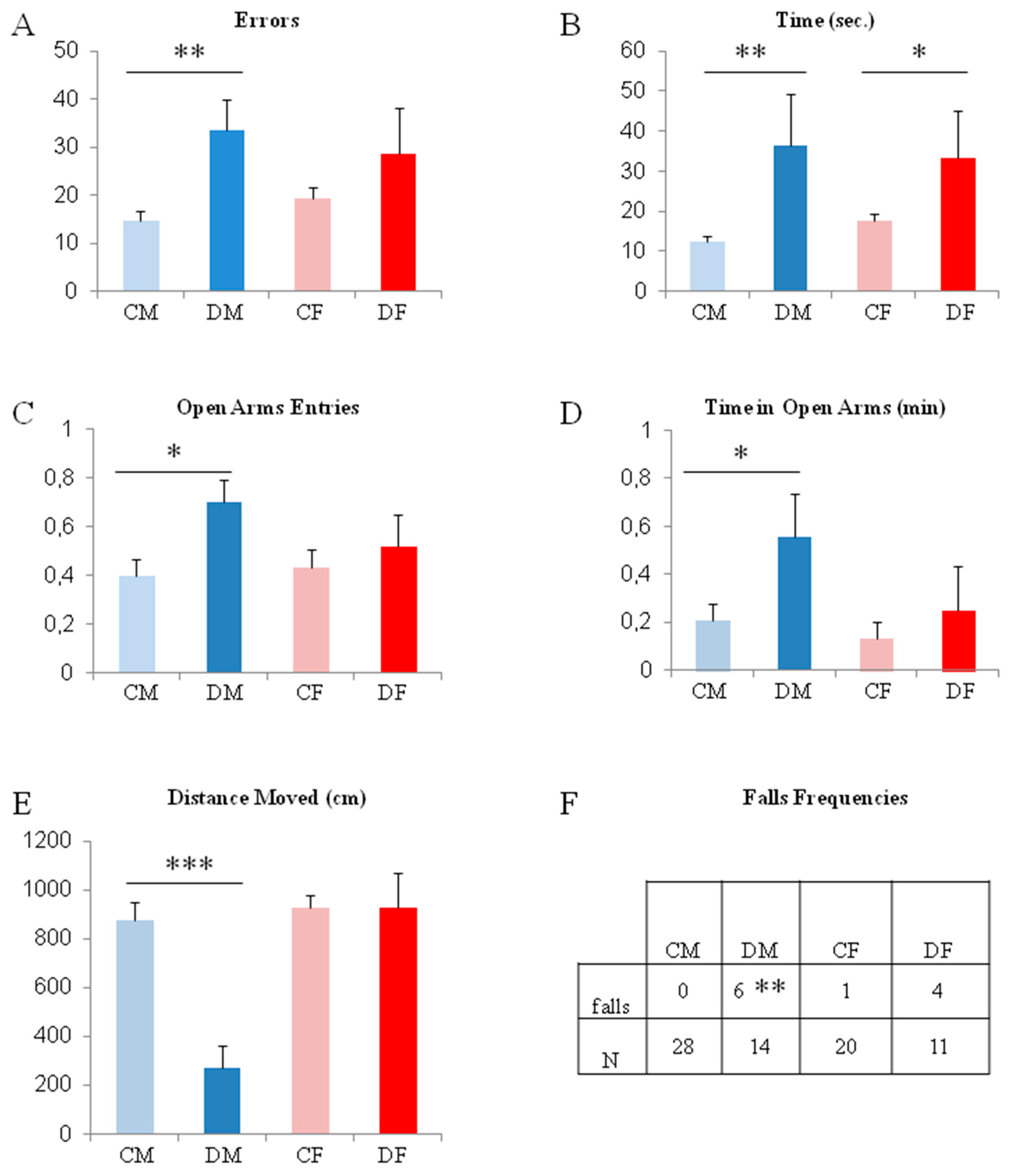
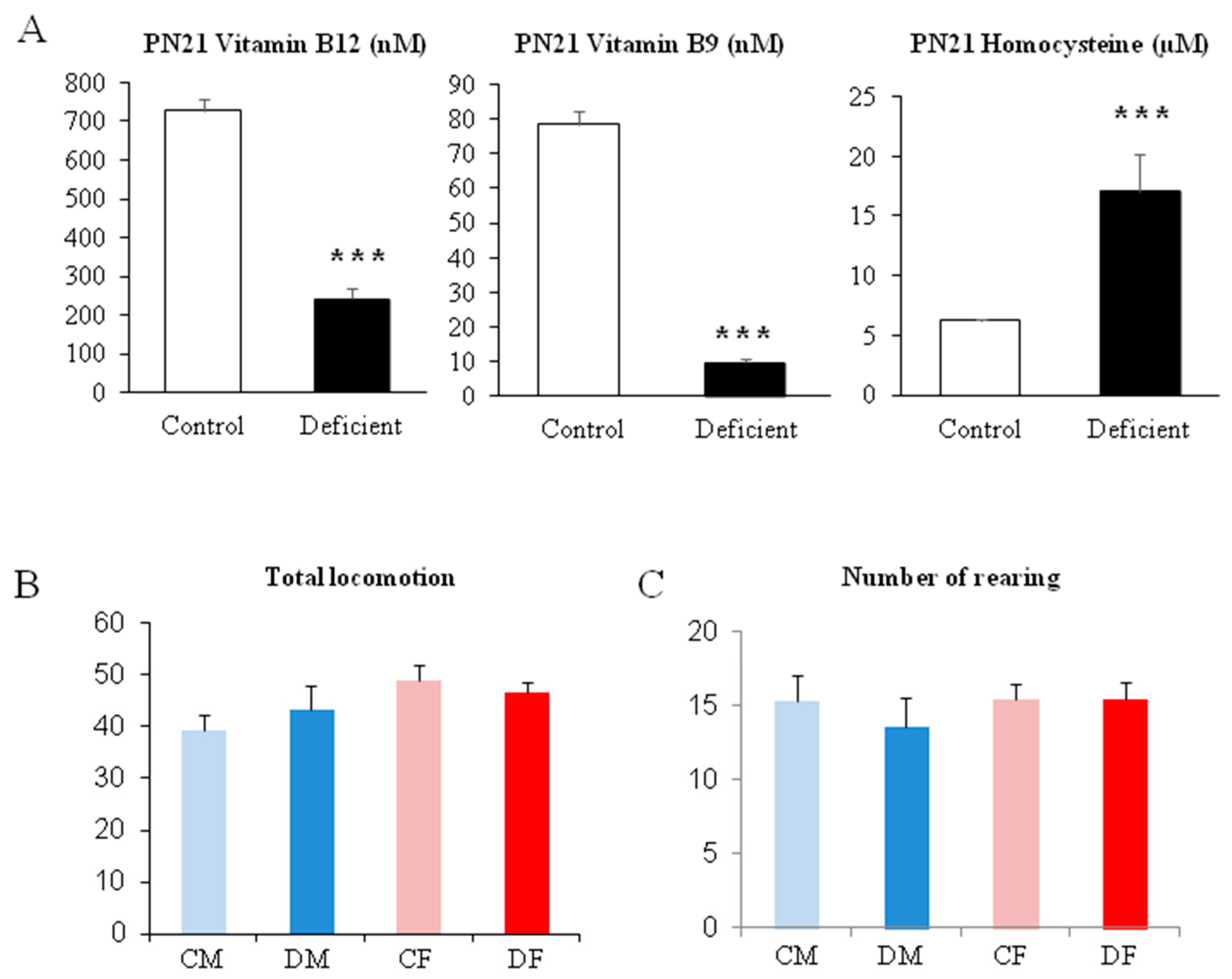
2.1. Gestational Deficiency Affects Young Rats During Post-Development
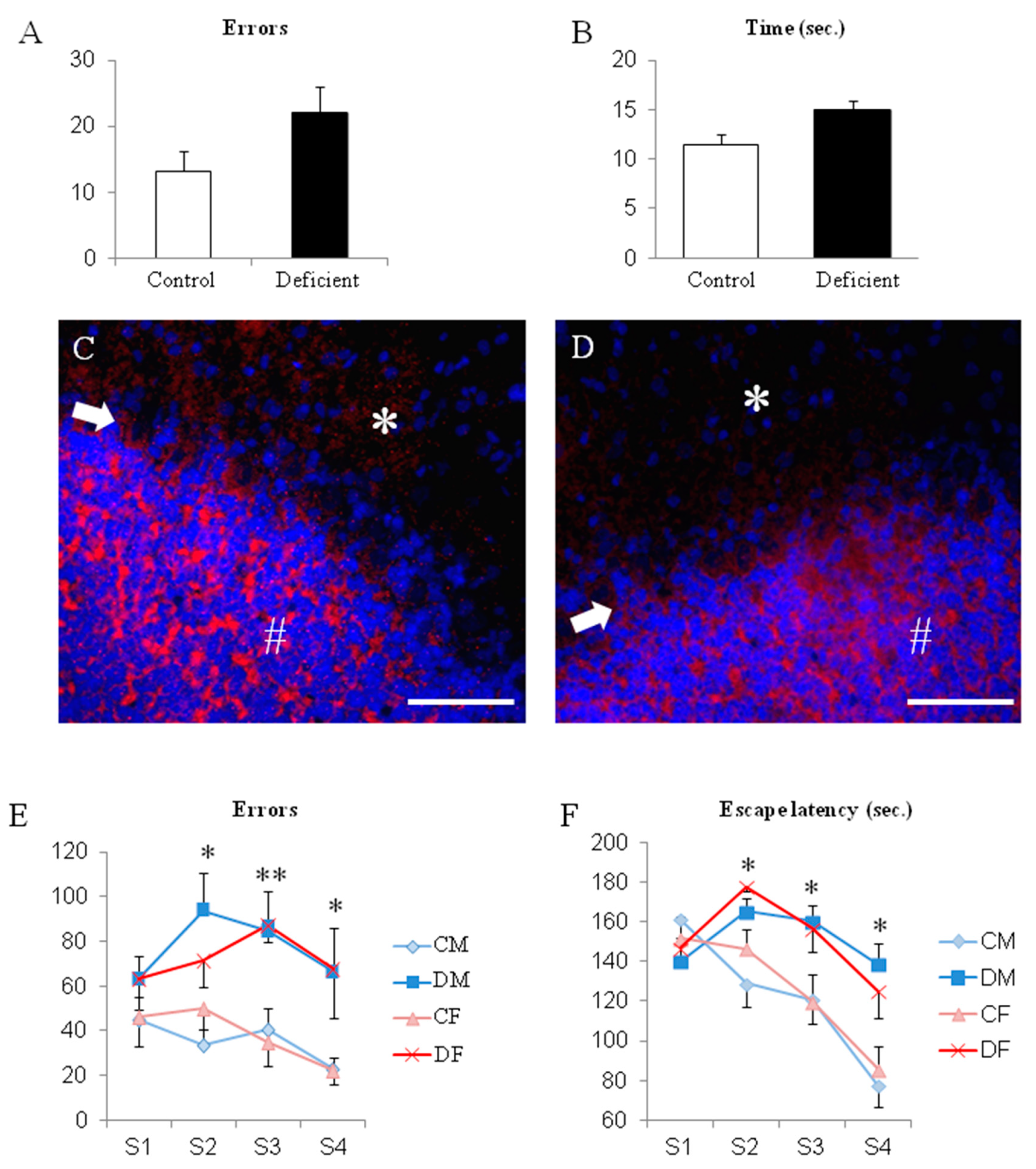
2.2. The Switch to a Normal Diet Restores Motor but not Cognitive Performances
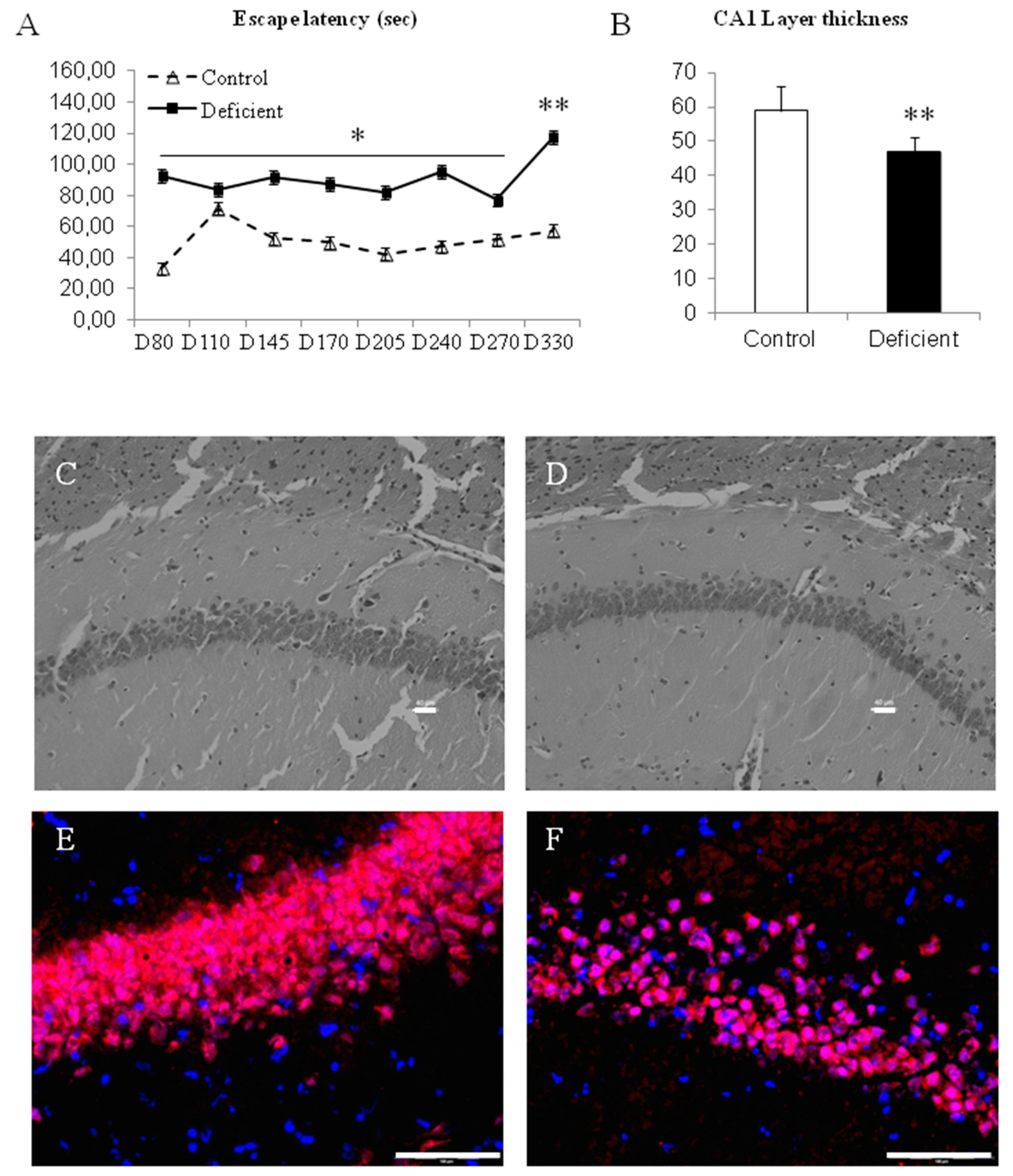
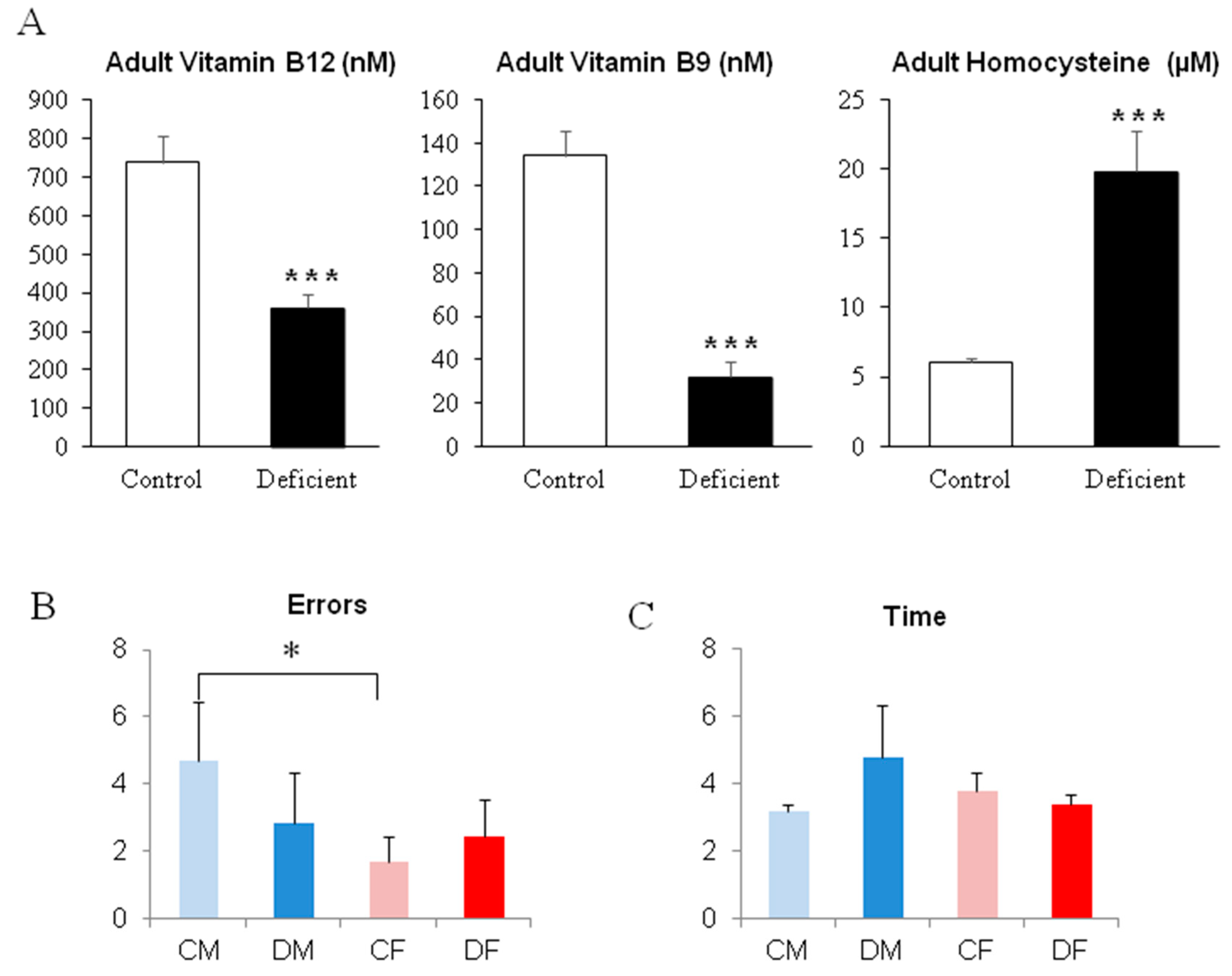
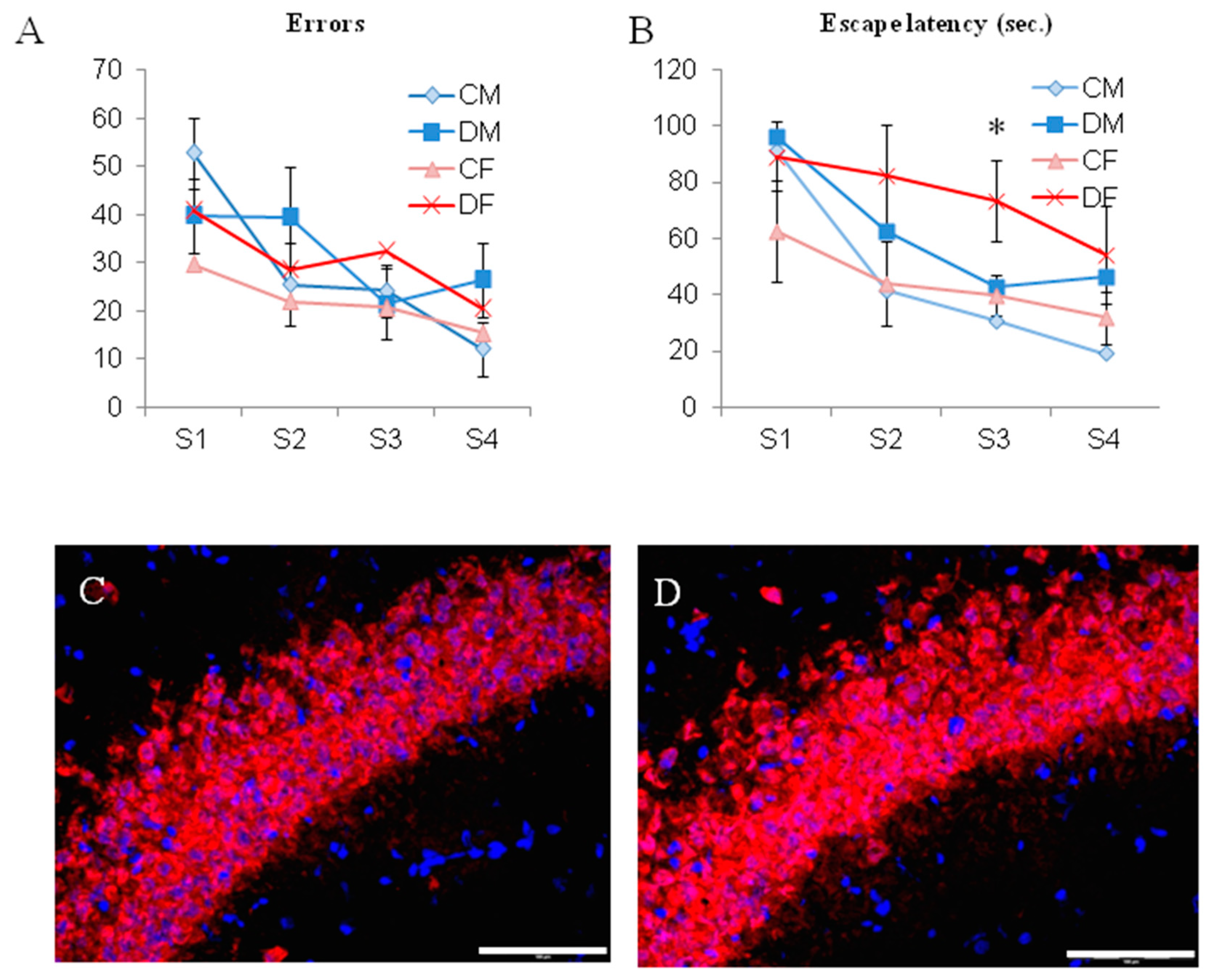
2.3. A Long Period of Deficiency During Adult Life Did Not Dramatically Affect Rat Behavior
3. Discussion
4. Methods
4.1. Animal Treatments
| Vitamins | Control Food (mg/kg) | Deficient Food (mg/kg) |
| B2 | 6.5 | 0.34 |
| B6 | 2.6 | 3.83 |
| B9 | 0.5 | 0.04 |
| B12 | 0.02 | 0.00 |
4.2. Biochemical Analyses
4.3. Behavioral Tests
4.3.1. Linear Walking Test
4.3.2. Elevated Plus Maze
4.3.3. Open-Field Test
4.4. Aquatic Maze
4.5. Histology
4.6. Statistics
Author Contributions
Funding
Conflicts of Interest
References
- Clarke, R. Vitamin B12, folic acid, and the prevention of dementia. New Engl. J. Med. 2006, 354, 2817–2819. [Google Scholar] [CrossRef] [PubMed]
- Miller, J.W. Assessing the association between vitamin B-12 status and cognitive function in older adults. Am. J. Clin. Nutr. 2006, 84, 1259–1260. [Google Scholar] [CrossRef] [PubMed]
- Scalabrino, G. The multi-faceted basis of vitamin B12 (cobalamin) neurotrophism in adult central nervous system: Lessons learned from its deficiency. Prog. Neurobiol. 2009, 88, 203–220. [Google Scholar] [CrossRef] [PubMed]
- Seshadri, S.; Beiser, A.; Selhub, J.; Jacques, P.F.; Rosenberg, I.H.; D’Agostino, R.B.; Wilson, P.W.; Wolf, P.A. Plasma homocysteine as a risk factor for dementia and Alzheimer′s disease. N Engl. J. Med. 2002, 346, 476–483. [Google Scholar] [CrossRef]
- Burdge, G.C.; Lillycrop, K.A. Nutrition, epigenetics, and developmental plasticity: Implications for understanding human disease. Annu. Rev. Nutr. 2010, 30, 315–339. [Google Scholar] [CrossRef]
- Daval, J.L.; Blaise, S.; Guéant, J.L. Vitamin B deficiency causes neural cell loss and cognitive impairment in the developing rat. Proc. Natl. Acad. Sci. USA 2009, 6, 106. [Google Scholar] [CrossRef]
- Troen, A.M.; Shea-Budgell, M.; Shukitt-Hale, B.; Smith, D.E.; Selhub, J.; Rosenberg, I.H. B-vitamin deficiency causes hyperhomocysteinemia and vascular cognitive impairment in mice. Proc. Natl. Acad. Sci. USA 2008, 105, 12474–12479. [Google Scholar] [CrossRef]
- Sechi, G.; Sechi, E.; Fois, C.; Kumar, N. Advances in clinical determinants and neurological manifestations of B vitamin deficiency in adults. Nutr. Rev. 2016, 74, 281–300. [Google Scholar] [CrossRef]
- Battaglia-Hsu, S.F.; Akchiche, N.; Noel, N.; Alberto, J.-M.; Jeannesson, E.; Orozco-Barrios, C.E.; Martinez-Fong, D.; Daval, J.-L.; Guéant, J.-L. Vitamin B12 deficiency reduces proliferation and promotes differentiation of neuroblastoma cells and up-regulates PP2A, proNGF, and TACE. Proc. Natl. Acad. Sci. USA 2009, 106, 21930–21935. [Google Scholar] [CrossRef]
- Black, M.M. Effects of vitamin B12 and folate deficiency on brain development in children. Food Nutr. Bull. 2008, 29, S126–S131. [Google Scholar] [CrossRef]
- Huemer, M.; Bürer, C.; Jesina, P.; Kožich, V.; Landolt, M.A.; Suormala, T.; Fowler, B.; Savvopoulou, P.A.; Blair, E.; Brennerova, K.; et al. Clinical onset and course, response to treatment and outcome in 24 patients with the cblE or cblG remethylation defect complemented by genetic and in vitro enzyme study data. J. Inherit. Metab. Dis. 2014, 38, 957–967. [Google Scholar] [CrossRef] [PubMed]
- Durga, J.; Van Boxtel, M.P.; Schouten, E.G.; Kok, F.J.; Jolles, J.; Katan, M.B.; Verhoef, P. Effect of 3-year folic acid supplementation on cognitive function in older adults in the FACIT trial: A randomised, double blind, controlled trial. Lancet 2007, 369, 208–216. [Google Scholar] [CrossRef]
- Stanger, O.; Fowler, B.; Piertzik, K.; Huemer, M.; Haschke-Becher, E.; Semmler, A.; Lorenzl, S.; Linnebank, M. Homocysteine, folate and vitamin B12in neuropsychiatric diseases: Review and treatment recommendations. Expert Rev. Neurother. 2009, 9, 1393–1412. [Google Scholar] [CrossRef] [PubMed]
- Blaise, S.A.; Nédélec, E.; Schroeder, H.; Alberto, J.-M.; Bossenmeyer-Pourié, C.; Guéant, J.-L.; Daval, J.-L. Gestational Vitamin B Deficiency Leads to Homocysteine-Associated Brain Apoptosis and Alters Neurobehavioral Development in Rats. Am. J. Pathol. 2007, 170, 667–679. [Google Scholar] [CrossRef] [PubMed]
- Petcu, E.B.; Sfredel, V.; Platt, D.; Herndon, J.G.; Kessler, C.; Popa-Wagner, A. Cellular and Molecular Events Underlying the Dysregulated Response of the Aged Brain to Stroke: A Mini-Review. Gerontology 2008, 54, 6–17. [Google Scholar] [CrossRef]
- McGee, M.; Bainbridge, S.; Fontaine-Bisson, B. A crucial role for maternal dietary methyl donor intake in epigenetic programming and fetal growth outcomes. Nutr. Rev. 2018, 76, 469–478. [Google Scholar] [CrossRef]
- Reynolds, E. Vitamin B12, folic acid, and the nervous system. Lancet. Neurol. 2006, 5, 949–960. [Google Scholar] [CrossRef]
- McKee, S.E.; Reyes, T.M. Effect of supplementation with methyl-donor nutrients on neurodevelopment and cognition: Considerations for future research. Nutr. Rev. 2018, 76, 497–511. [Google Scholar] [CrossRef]
- Zeisel, S.H.; Craciunescu, C.N. Dietary Choline Reverses Some, but Not All, Effects of Folate Deficiency on Neurogenesis and Apoptosis in Fetal Mouse Brain. SciVee 2010, 140, 1162–1166. [Google Scholar]
- Laanpere, M.; Altmäe, S.; Nilsson, T.K.; Yngve, A.; Salumets, A.; Stavreus-Evers, A.; Stavreus-Evers, A. Folate-mediated one-carbon metabolism and its effect on female fertility and pregnancy viability. Nutr. Rev. 2010, 68, 99–113. [Google Scholar] [CrossRef]
- Geoffroy, A.; Kerek, R.; Pourié, G.; Helle, D.; Guéant, J.L.; Daval, J.L. Bossenmeyer-Pourié. Late Maternal Folate Supplementation Rescues from Methyl Donor Deficiency-Associated Brain Defects by Restoring Let-7 and miR-34 Pathways. Mol. Neurobiol. 2017, 54, 5017–5033. [Google Scholar] [CrossRef] [PubMed]
- Moretti, R.; Caruso, P. The Controversial Role of Homocysteine in Neurology: From Labs to Clinical Practice. Int. J. Mol. Sci. 2019, 20, 231. [Google Scholar] [CrossRef] [PubMed]
- Blusztajn, J.K.; Slack, B.E.; Mellott, T.J. Neuroprotective Actions of Dietary Choline. Nutrrients 2017, 9, 815. [Google Scholar] [CrossRef] [PubMed]
- Kerek, R.; Geoffroy, A.; Bison, A.; Martin, N.; Akchiche, N.; Pourié, G.; Helle, D.; Guéant, J.-L.; Bossenmeyer-Pourié, C.; Daval, J.-L. Early methyl donor deficiency may induce persistent brain defects by reducing Stat3 signaling targeted by miR-124. Cell Death Dis. 2013, 4, e755. [Google Scholar] [CrossRef] [PubMed]
- Geoffroy, A.; Saber-Cherif, L.; Pourié, G.; Helle, D.; Umoret, R.; Guéant, J.-L.; Bossenmeyer-Pourié, C.; Daval, J.-L. Developmental Impairments in a Rat Model of Methyl Donor Deficiency: Effects of a Late Maternal Supplementation with Folic Acid. Int. J. Mol. Sci. 2019, 20, 973. [Google Scholar] [CrossRef]
- McGarel, C.; Pentieva, K.; Strain, J.J.; McNulty, H. Emerging roles for folate and related B-vitamins in brain health across the lifecycle. Proc. Nutr. Soc. 2015, 74, 46–55. [Google Scholar] [CrossRef]
- Grojean, S.; Schroeder, H.; Pourié, G.; Charriaut-Marlangue, C.; Koziel, V.; Desor, D.; Vert, P.; Daval, J.-L. Histopathological alterations and functional brain deficits after transient hypoxia in the newborn rat pup: A long term follow-up. Neurobiol. Dis. 2003, 14, 265–278. [Google Scholar] [CrossRef]
- Pourié, G.; Martin, N.; Bossenmeyer-Pourié, C.; Akchiche, N.; Gueant-Rodriguez, R.M.; Geoffroy, A.; Jeannesson, E.; Chehadeh, S.E.H.; Mimoun, K.; Brachet, P.; et al. Folate- and vitamin B12–deficient diet during gestation and lactation alters cerebellar synapsin expression via impaired influence of estrogen nuclear receptor α. Faseb J. 2015, 29, 3713–3725. [Google Scholar] [CrossRef]
- Etchell, A.; Adhikari, A.; Weinberg, L.S.; Choo, A.L.; Garnett, E.O.; Chow, H.M.; Chang, S.-E. A systematic literature review of sex differences in childhood language and brain development. Neuropsychology 2018, 114, 19–31. [Google Scholar] [CrossRef]
- Sallaberry, C.; Ardais, A.P.; Rocha, A.; Borges, M.F.; Fioreze, G.T.; Mioranzza, S.; Nunes, F.; Pagnussat, N.; Botton, P.H.S.; Porciúncula, L.O.; et al. Sex differences in the effects of pre- and postnatal caffeine exposure on behavior and synaptic proteins in pubescent rats. Prog. Neuro-Psychopharmacol. Boil. Psychiatry 2018, 81, 416–425. [Google Scholar] [CrossRef]
- Yang, F.; Tao, J.; Xu, L.; Zhao, N.; Chen, J.; Chen, W.; Zhu, Y.; Qiu, J. Estradiol decreases rat depressive behavior by estrogen receptor beta but not alpha: No correlation with plasma corticosterone. Neuroreport 2014, 25, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Patki, G.; Salvi, A.; Kelly, M.; Salim, S. Behavioral effects of early life maternal trauma witness in rats. Prog. Neuro-Psychopharmacol. Boil. Psychiatry 2018, 81, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.; Ryan, C.; Curley, A.; Mulcaire, J.; Kelly, J.P. Sex differences in baseline and drug-induced behavioural responses in classical behavioural tests. Prog. Neuro-Psychopharmacol. Boil. Psychiatry 2012, 37, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Fu, H.; Han, X.; Hu, X.; Gu, H.; Chen, Y.; Wei, Q.; Hu, Q. Role of Synaptic Structural Plasticity in Impairments of Spatial Learning and Memory Induced by Developmental Lead Exposure in Wistar Rats. PLoS ONE 2014, 9, e115556. [Google Scholar] [CrossRef] [Green Version]
- Mattson, M.P.; Kruman, I.; Duan, W.I.; Kruman, I. Folic acid and homocysteine in age-related disease. Ageing Res. Rev. 2002, 1, 95–111. [Google Scholar] [CrossRef]
- Chan, A.; Tchantchou, F.; Graves, V.; Rozen, R.; Shea, T.B. Dietary and genetic compromise in folate availability reduces acetylcholine, cognitive performance and increases aggression: Critical role of S-adenosyl methionine. J. Nutr. Heal. Aging 2008, 12, 252–261. [Google Scholar] [CrossRef]
- Khan, M.M.; Xiao, J.; Patel, D.; LeDoux, M.S. DNA damage and neurodegenerative phenotypes in aged Ciz1 null mice. Neurobiol. Aging 2018, 62, 180–190. [Google Scholar] [CrossRef]
- Spriggs, M.; Cadwallader, C.; Hamm, J.; Tippett, L.; Kirk, I. Age-related alterations in human neocortical plasticity. Brain Res. Bull. 2017, 130, 53–59. [Google Scholar] [CrossRef]
- McKay, B.E.; Turner, R.W. Physiological and morphological development of the rat cerebellar Purkinje cell. J. Physiol. 2005, 567, 829–850. [Google Scholar] [CrossRef]
- Rahimi, O.; Claiborne, B.J. Morphological development and maturation of granule neuron dendrites in the rat dentate gyrus. Prog. Brain Res. 2007, 163, 167–181. [Google Scholar]
- Shea, T.B.; Rogers, E. Lifetime requirement of the methionine cycle for neuronal development and maintenance. Curr. Opin. Psychiatry 2014, 27, 138–142. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, Z.; Coelho, D.; Kokten, T.; Alberto, J.-M.; Umoret, R.; Daval, J.-L.; Guéant, J.-L.; Bossenmeyer-Pourié, C.; Pourié, G. Brain Susceptibility to Methyl Donor Deficiency: From Fetal Programming to Aging Outcome in Rats. Int. J. Mol. Sci. 2019, 20, 5692. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225692
Hassan Z, Coelho D, Kokten T, Alberto J-M, Umoret R, Daval J-L, Guéant J-L, Bossenmeyer-Pourié C, Pourié G. Brain Susceptibility to Methyl Donor Deficiency: From Fetal Programming to Aging Outcome in Rats. International Journal of Molecular Sciences. 2019; 20(22):5692. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225692
Chicago/Turabian StyleHassan, Ziad, David Coelho, Tunay Kokten, Jean-Marc Alberto, Rémy Umoret, Jean-Luc Daval, Jean-Louis Guéant, Carine Bossenmeyer-Pourié, and Grégory Pourié. 2019. "Brain Susceptibility to Methyl Donor Deficiency: From Fetal Programming to Aging Outcome in Rats" International Journal of Molecular Sciences 20, no. 22: 5692. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms20225692