Biodegradation of Bisphenol A by Sphingobium sp. YC-JY1 and the Essential Role of Cytochrome P450 Monooxygenase


Abstract
:
1. Introduction
2. Results and Discussion
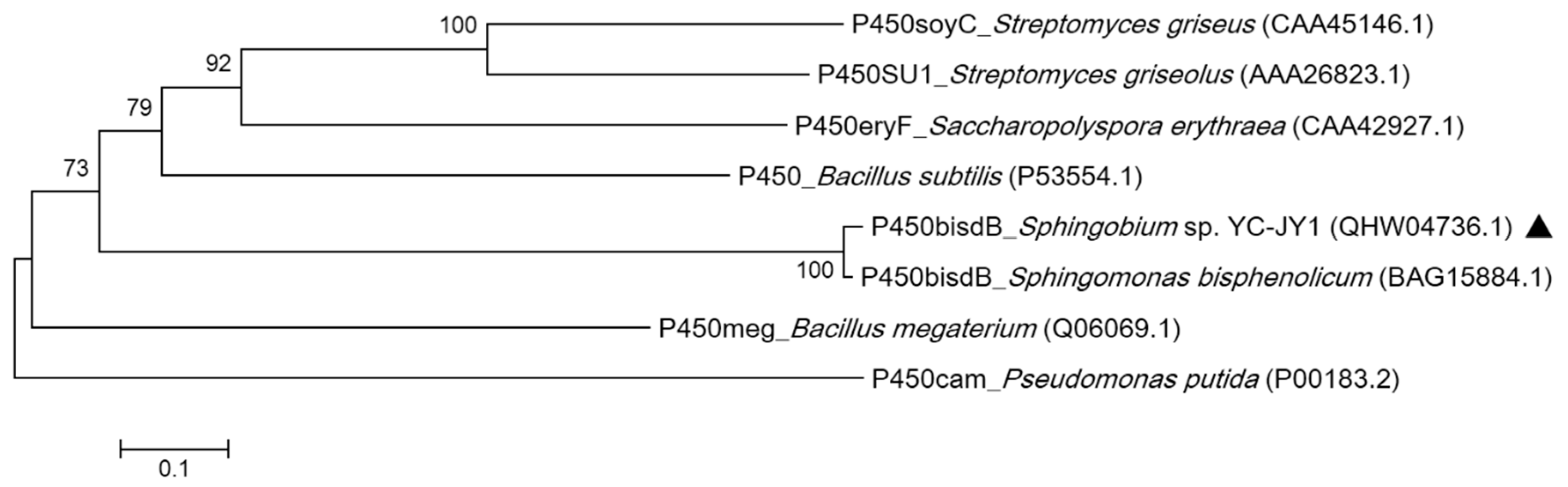
2.1. Strain Isolation and Identification
2.2. Effects of Environmental Factors on Biodegradation of BPA
2.3. Identification of Metabolites
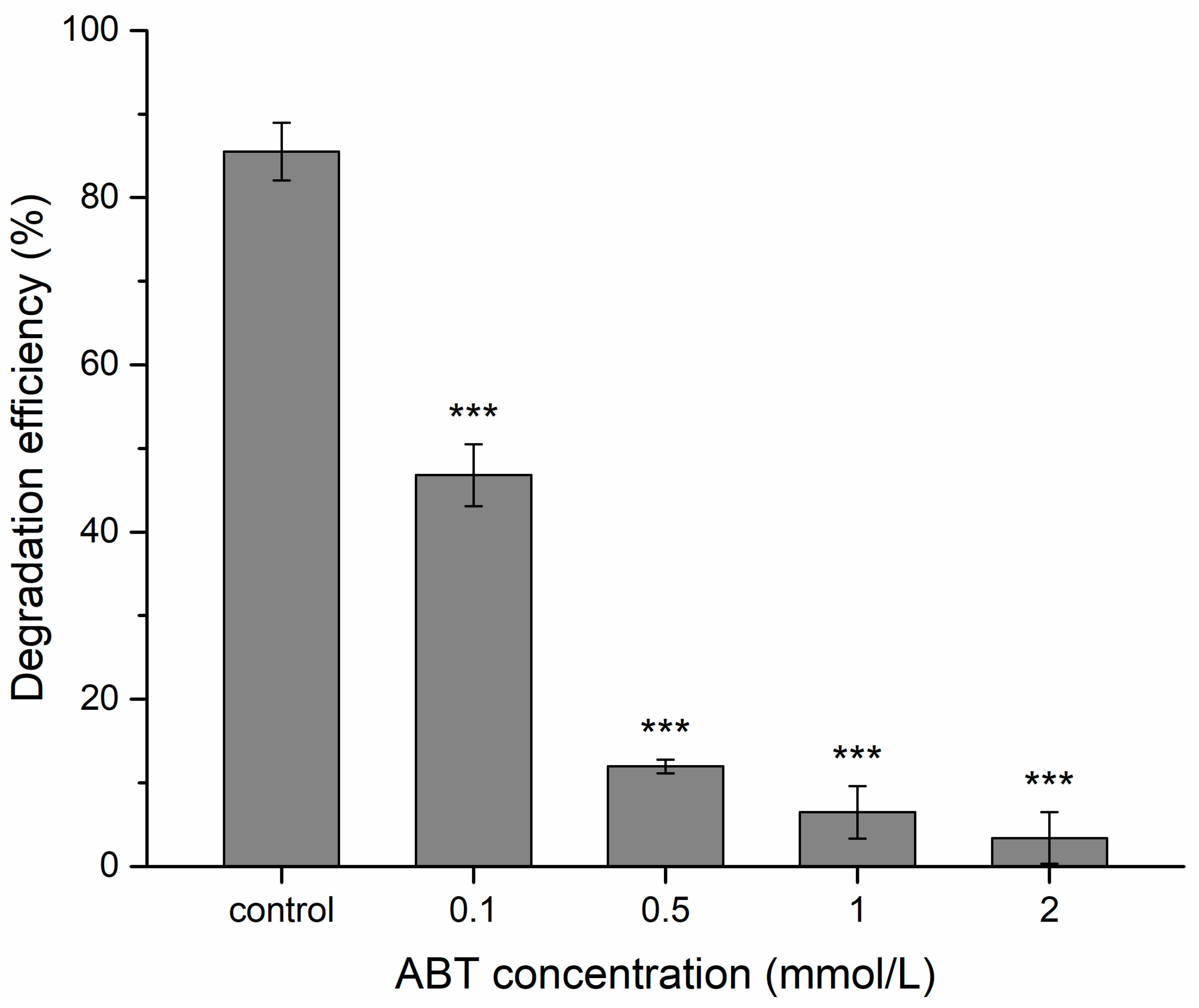
2.4. Effect of Cytochrome P450 Inhibitor and BPA Degradation Activity of Cytochrome P450
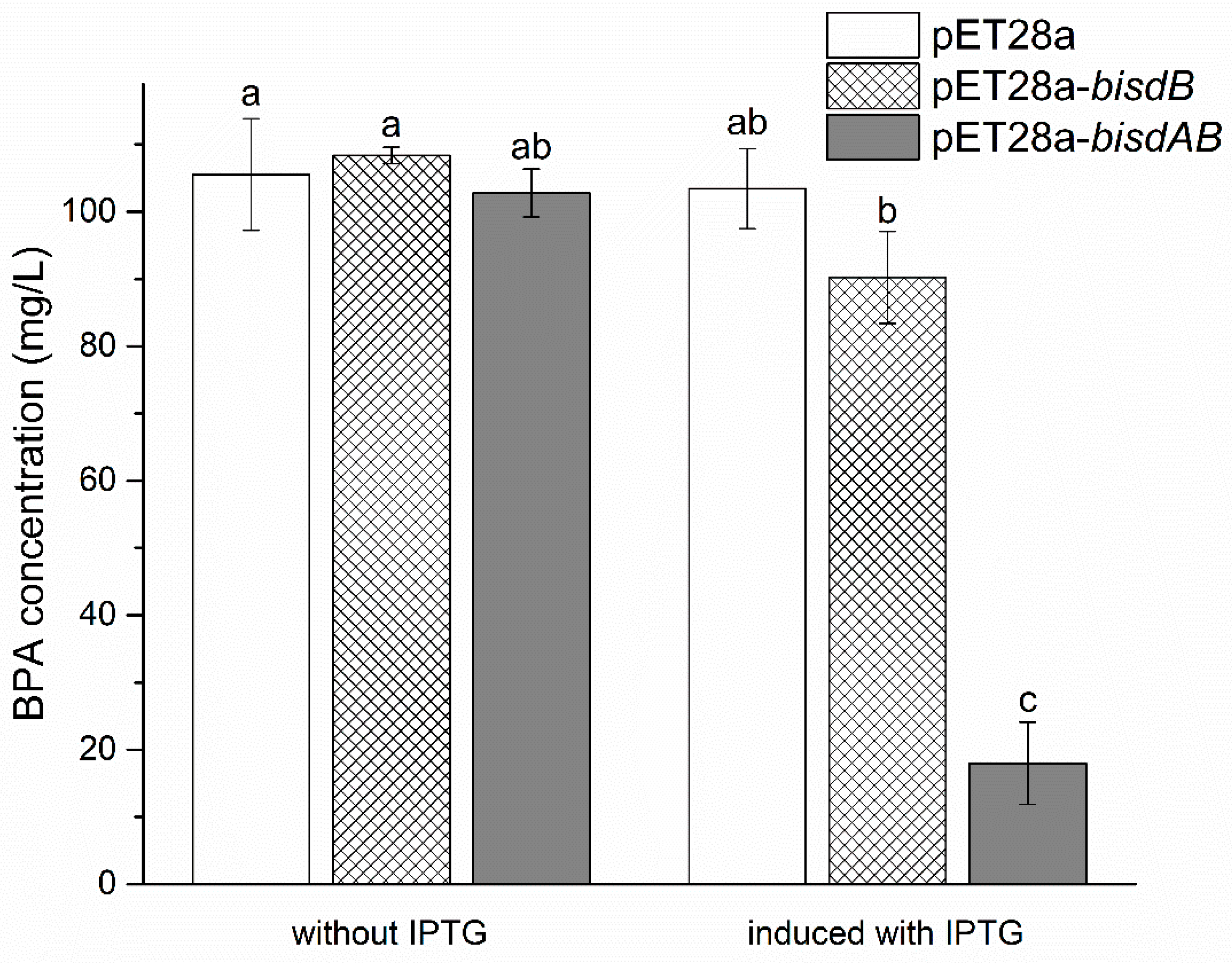
2.5. Essential Role of CYP Gene for BPA Degradation in Strain YC-JY1
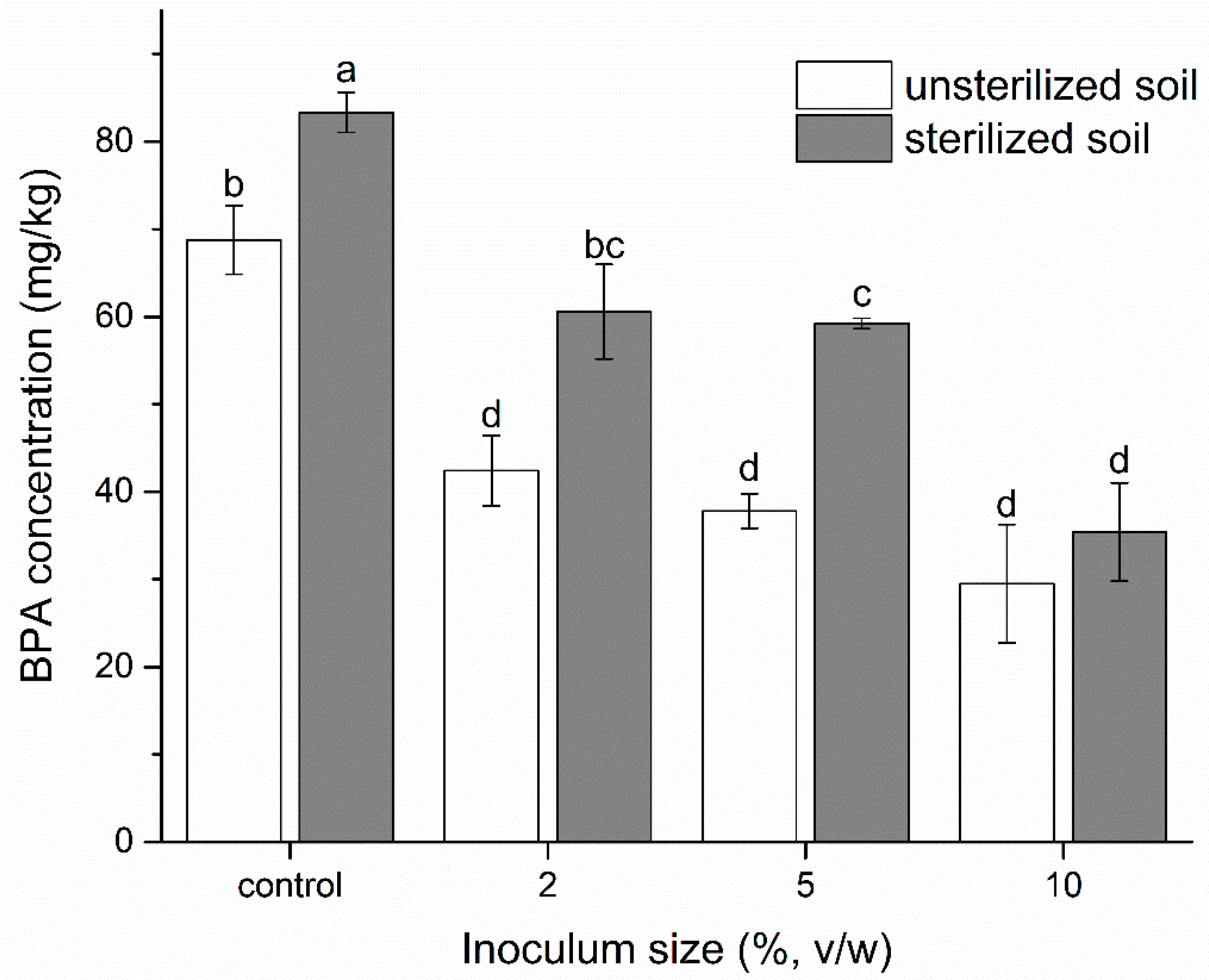
2.6. Effect of Inoculated YC-JY1 on Remediation of BPA-Contaminated Soil
3. Materials and Methods
3.1. Chemicals and Medium
3.2. Strain Isolation and Identification
3.3. Preparation of the Bacterial Suspension
3.4. Effects of Environmental Factors on BPA Biodegradation
3.5. Identification of Metabolites
3.6. Cytochrome P450 Inhibitor Experiment
3.7. Cloning and Expression of bisdB and bisdAB
3.8. Gene Knockout of bisdB
3.9. Remediation of BPA-Contaminated Soil
3.10. Analytical Methods and Statistical Analysis
3.11. Accession Numbers
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Kamaraj, M.; Sivaraj, R.; Venckatesh, R. Biodegradation of Bisphenol A by the tolerant bacterial species isolated from coastal regions of Chennai, Tamil Nadu, India. Int. Biodeterior. Biodegrad. 2014, 93, 216–222. [Google Scholar] [CrossRef]
- Yassine, O.; Mehdi, Z.; Patrick, D.; Brahima, S.; Gerardo, B.; Rino, D. Performance of a membrane bioreactor in extreme concentrations of bisphenol A. Water Sci. Technol. 2018, 77, 1505–1513. [Google Scholar]
- Fu, M.; Xing, J.; Ge, Z. Preparation of laccase-loaded magnetic nanoflowers and their recycling for efficient degradation of bisphenol A. Sci. Total Environ. 2019, 651, 2857–2865. [Google Scholar] [CrossRef] [PubMed]
- Olajuyigbe, F.M.; Adetuyi, O.Y.; Fatokun, C.O. Characterization of free and immobilized laccase from Cyberlindnera fabianii and application in degradation of bisphenol A. Int. J. Biol. Macromol. 2019, 125, 856–864. [Google Scholar] [CrossRef] [PubMed]
- Mohapatra, D.P.; Brar, S.K.; Tyagi, R.D.; Surampalli, R.Y. Physico-chemical pre-treatment and biotransformation of wastewater and wastewater sludge-fate of bisphenol A. Chemosphere 2010, 78, 923–941. [Google Scholar] [CrossRef]
- Huang, Y.Q.; Wong, C.K.; Zheng, J.S.; Bouwman, H.; Barra, R.; Wahlstrom, B.; Neretin, L.; Wong, M.H. Bisphenol A (BPA) in China: A review of sources, environmental levels, and potential human health impacts. Environ. Int. 2012, 42, 91–99. [Google Scholar] [CrossRef]
- Eio, E.J.; Kawai, M.; Tsuchiya, K.; Yamamoto, S.; Toda, T. Biodegradation of bisphenol A by bacterial consortia. Int. Biodeterior. Biodegrad. 2014, 96, 166–173. [Google Scholar] [CrossRef]
- de Freitas, E.N.; Bubna, G.A.; Brugnari, T.; Kato, C.G.; Nolli, M.; Rauen, T.G.; de Fátima Peralta Muniz Moreira, R.; Peralta, R.A.; Bracht, A.; de Souza, C.G.M.; et al. Removal of bisphenol A by laccases from Pleurotus ostreatus and Pleurotus pulmonarius and evaluation of ecotoxicity of degradation products. Chem. Eng. J. 2017, 330, 1361–1369. [Google Scholar] [CrossRef]
- Li, G.; Zu, L.; Wong, P.K.; Hui, X.; Lu, Y.; Xiong, J.; An, T. Biodegradation and detoxification of bisphenol A with one newly-isolated strain Bacillus sp. GZB: Kinetics, mechanism and estrogenic transition. Bioresource Technol. 2012, 114, 224–230. [Google Scholar] [CrossRef]
- Suyamud, B.; Inthorn, D.; Panyapinyopol, B.; Thiravetyan, P. Biodegradation of bisphenol a by a newly isolated Bacillus megaterium strain ISO-2 from a polycarbonate industrial wastewater. Water Air Soil Poll. 2018, 229, 229–348. [Google Scholar] [CrossRef]
- Haighton, L.A.; Hlywka, J.J.; Doull, J.; Kroes, R.; Lynch, B.S.; Munro, I.C. An evaluation of the possible carcinogenicity of bisphenol A to humans. Regul. Toxicol. Pharm. 2002, 35, 238–254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nowak, K.; Jablonska, E.; Ratajczak-Wrona, W. Immunomodulatory effects of synthetic endocrine disrupting chemicals on the development and functions of human immune cells. Environ. Int. 2019, 125, 350–364. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Cheng, W.; Feng, Y.; Wei, H.; Liang, F.; Wang, Y. Interactions between three typical endocrine-disrupting chemicals (EDCs) in binary mixtures exposure on myocardial differentiation of mouse embryonic stem cell. Chemosphere 2017, 178, 378–383. [Google Scholar] [CrossRef] [PubMed]
- Berger, A.; Ziv-Gal, A.; Cudiamat, J.; Wang, W.; Zhou, C.; Flaws, J.A. The effects of in utero bisphenol A exposure on the ovaries in multiple generations of mice. Reprod. Toxicol. 2016, 60, 39–52. [Google Scholar] [CrossRef] [Green Version]
- Eltoukhy, A.; Jia, Y.; Nahurira, R.; Abo-Kadoum, M.A.; Khokhar, I.; Wang, J.; Yan, Y. Biodegradation of endocrine disruptor Bisphenol A by Pseudomonas putida strain YC-AE1 isolated from polluted soil, Guangdong, China. BMC Microbiol. 2020, 20, 11. [Google Scholar] [CrossRef] [Green Version]
- Lobos, J.H.; Leib, T.K.; Su, T.M. Biodegradation of bisphenol A and other bisphenols by a gram-negative aerobic bacterium. Appl. Environ. Microbiol. 1992, 58, 1823–1831. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, M.; Akahira, A.; Oshiman, K.; Tsuchido, T.; Matsumura, Y. Purification of cytochrome P450 and ferredoxin, involved in bisphenol A degradation, from Sphingomonas sp. strain AO1. Appl. Environ. Microbiol. 2005, 71, 8024–8030. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, M.; Maki, J.; Oshiman, K.; Matsumura, Y.; Tsuchido, T. Biodegradation of bisphenol A by cells and cell lysate from Sphingomonas sp. strain AO1. Biodegradation 2005, 16, 449–459. [Google Scholar] [CrossRef]
- Sasaki, M.; Tsuchido, T.; Matsumura, Y. Molecular cloning and characterization of cytochrome P450 and ferredoxin genes involved in bisphenol A degradation in Sphingomonas bisphenolicum strain AO1. J. Appl. Microbiol. 2008, 105, 1158–1169. [Google Scholar] [CrossRef]
- Zhou, N.A.; Kjeldal, H.; Gough, H.L.; Nielsen, J.L. Identification of Putative Genes Involved in Bisphenol A Degradation Using Differential Protein Abundance Analysis of Sphingobium sp. BiD32. Environ. Sci. Technol. 2015, 49, 12232–12241. [Google Scholar] [CrossRef]
- Zhang, C.; Zeng, G.; Yuan, L.; Yu, J.; Li, J.; Huang, G.; Xi, B.; Liu, H. Aerobic degradation of bisphenol A by Achromobacter xylosoxidans strain B-16 isolated from compost leachate of municipal solid waste. Chemosphere 2007, 68, 181–190. [Google Scholar] [CrossRef]
- Toyama, T.; Sato, Y.; Inoue, D.; Sei, K.; Chang, Y.C.; Kikuchi, S.; Ike, M. Biodegradation of bisphenol A and bisphenol F in the rhizosphere sediment of Phragmites australis. J. Biosci. Bioeng. 2009, 108, 147–150. [Google Scholar] [CrossRef] [PubMed]
- Fischer, J.; Kappelmeyer, U.; Kastner, M.; Schauer, F.; Heipieper, H.J. The degradation of bisphenol A by the newly isolated bacterium Cupriavidus basilensis JF1 can be enhanced by biostimulation with phenol. Int. Biodeterior. Biodegrad. 2010, 64, 324–330. [Google Scholar] [CrossRef]
- Zuhlke, M.K.; Schluter, R.; Mikolasch, A.; Zuhlke, D.; Giersberg, M.; Schindler, H.; Henning, A.K.; Frenzel, H.; Hammer, E.; Lalk, M.; et al. Biotransformation and reduction of estrogenicity of bisphenol A by the biphenyl-degrading Cupriavidus basilensis. Appl. Microbiol. Biotechnol. 2017, 101, 3743–3758. [Google Scholar] [CrossRef] [PubMed]
- Tsutsumi, Y.; Haneda, T.; Nishida, T. Removal of estrogenic activities of bisphenol A and nonylphenol by oxidative enzymes from lignin-degrading basidiomycetes. Chemosphere 2001, 42, 271–276. [Google Scholar] [CrossRef] [Green Version]
- Galliker, P.; Hommes, G.; Schlosser, D.; Corvini, P.F.; Shahgaldian, P. Laccase-modified silica nanoparticles efficiently catalyze the transformation of phenolic compounds. J. Colloid Interf. Sci. 2010, 349, 98–105. [Google Scholar] [CrossRef]
- Cajthaml, T. Biodegradation of endocrine-disrupting compounds by ligninolytic fungi: Mechanisms involved in the degradation. Environ. Microbiol. 2015, 17, 4822–4834. [Google Scholar] [CrossRef]
- Telke, A.A.; Kalyani, D.C.; Jadhav, U.U.; Parshetti, G.K.; Govindwar, S.P. Purification and characterization of an extracellular laccase from a Pseudomonas sp. LBC1 and its application for the removal of bisphenol A. J. Mol. Catal. B Enzym. 2009, 61, 252–260. [Google Scholar] [CrossRef]
- Roh, H.; Subramanya, N.; Zhao, F.; Yu, C.P.; Sandt, J.; Chu, K.H. Biodegradation potential of wastewater micropollutants by ammonia-oxidizing bacteria. Chemosphere 2009, 77, 1084–1089. [Google Scholar] [CrossRef]
- Sakai, K.; Yamanaka, H.; Moriyoshi, K.; Ohmoto, T.; Ohe, T. Biodegradation of bisphenol a and related compounds by Sphingomonas sp. strain BP-7 isolated from seawater. Biosci. Biotech. Bioch. 2007, 71, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Zhou, N.A.; Lutovsky, A.C.; Andaker, G.L.; Gough, H.L.; Ferguson, J.F. Cultivation and characterization of bacterial isolates capable of degrading pharmaceutical and personal care products for improved removal in activated sludge wastewater treatment. Biodegradation 2013, 24, 813–827. [Google Scholar] [CrossRef] [PubMed]
- Sun, Q.; Li, Y.; Chou, P.H.; Peng, P.Y.; Yu, C.P. Transformation of bisphenol A and alkylphenols by ammonia-oxidizing bacteria through nitration. Environ. Sci. Technol. 2012, 46, 4442–4448. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Jia, Y.; Ruth, N.; Shi, Y.; Wang, J.; Qiao, C.; Yan, Y. Biotransformations of bisphenols mediated by a novel Arthrobacter sp. strain YC-RL1. Appl. Microbiol. Biotechnol. 2016, 100, 1967–1976. [Google Scholar] [CrossRef] [PubMed]
- Nahurira, R.; Ren, L.; Song, J.; Jia, Y.; Wang, J.; Fan, S.; Wang, H.; Yan, Y. Degradation of Di(2-Ethylhexyl) Phthalate by a Novel Gordonia alkanivorans Strain YC-RL2. Curr. Microbiol. 2017, 74, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Ren, L. Biodegradation of Di-(2-ethylhexyl) Phthalate by Rhodococcus ruber YC-YT1 in Contaminated Water and Soil. Int. J. Environ. Res. Public Health 2018, 15, 964. [Google Scholar] [CrossRef] [Green Version]
- Zeng, J.; Zhu, Q.; Wu, Y.; Lin, X. Oxidation of polycyclic aromatic hydrocarbons using Bacillus subtilis CotA with high laccase activity and copper independence. Chemosphere 2016, 148, 1–7. [Google Scholar] [CrossRef]
- Wei, K.; Yin, H.; Peng, H.; Lu, G.; Dang, Z. Bioremediation of triphenyl phosphate by Brevibacillus brevis: Degradation characteristics and role of cytochrome P450 monooxygenase. Sci. Total Environ. 2018, 627, 1389–1395. [Google Scholar] [CrossRef]
- Kang, J.H.; Kondo, F. Effects of bacterial counts and temperature on the biodegradation of bisphenol A in river water. Chemosphere 2002, 49, 493–498. [Google Scholar] [CrossRef]
- Boonyaroj, V.; Chiemchaisri, C.; Chiemchaisri, W.; Theepharaksapan, S.; Yamamoto, K. Toxic organic micro-pollutants removal mechanisms in long-term operated membrane bioreactor treating municipal solid waste leachate. Bioresour. Technol. 2012, 113, 174–180. [Google Scholar] [CrossRef]
- McCormick, J.M.; Van Es, T.; Cooper, K.R.; White, L.A.; Haggblom, M.M. Microbially mediated O-methylation of bisphenol A results in metabolites with increased toxicity to the developing zebrafish (Danio rerio) embryo. Environ. Sci. Technol. 2011, 45, 6567–6574. [Google Scholar] [CrossRef]
- Das, R.; Li, G.; Mai, B.; An, T. Spore cells from BPA degrading bacteria Bacillus sp. GZB displaying high laccase activity and stability for BPA degradation. Sci. Total Environ. 2018, 640-641, 798–806. [Google Scholar] [CrossRef] [PubMed]
- Rappert, S.; Li, R.; Kokova, M.; Antholz, M.; Nagorny, S.; Francke, W.; Muller, R. Degradation of 2,5-dimethylpyrazine by Rhodococcus erythropolis strain DP-45 isolated from a waste gas treatment plant of a fishmeal processing company. Biodegradation 2007, 18, 585–596. [Google Scholar] [CrossRef] [PubMed]
- Ning, D.; Wang, H.; Zhuang, Y. Induction of functional cytochrome P450 and its involvement in degradation of benzoic acid by Phanerochaete chrysosporium. Biodegradation 2010, 21, 297–308. [Google Scholar] [CrossRef] [PubMed]
- Mtibaa, R.; Olicon-Hernandez, D.R.; Pozo, C.; Nasri, M.; Mechichi, T.; Gonzalez, J.; Aranda, E. Degradation of bisphenol A and acute toxicity reduction by different thermo-tolerant ascomycete strains isolated from arid soils. Ecotoxicol. Environ. Saf. 2018, 156, 87–96. [Google Scholar] [CrossRef]
- Yu, K.; Yi, S.; Li, B.; Guo, F.; Peng, X.; Wang, Z.; Wu, Y.; Alvarez-Cohen, L.; Zhang, T. An integrated meta-omics approach reveals substrates involved in synergistic interactions in a bisphenol A (BPA)-degrading microbial community. Microbiome 2019, 7, 16. [Google Scholar] [CrossRef] [Green Version]
- Matsumura, Y.; Akahira-Moriya, A.; Sasaki-Mori, M. Bioremediation of bisphenol-A polluted soil by Sphingomonas bisphenolicum AO1 and the microbial community existing in the soil. Biocontrol Sci. 2015, 20, 35–42. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microorganism | Resources | Degradation Efficiency | References |
|---|---|---|---|
| Sphingomonas sp. strain MV1 | sludge | 100%, 10 g/L, 4 days | [16] |
| Sphingomonas bisphenolicum strain AO1 | soil | 100%, 115 mg/L, 117 h (without other carbon sources); 100%, 115 mg/L, 6 h (with glucose) | [18] |
| Achromobacter xylosoxidans B-16 | wastes compost leachate | 100%, 3 mg/L, 4.5 days | [21] |
| Sphingomonas sp. strain BP-7 | offshore seawater | >95%, 114 mg/L, 40 days | [30] |
| Cupriavidus basilensis JF1 | fixed bed reactor | 50%, 34.2 mg/L, 144 days | [23] |
| Bacillus sp. GZB | sediment | 92.9%, 5 mg/L, 96 h | [9] |
| Sphingobium sp. BiD32 | sludge | 100%, 1 mg/L, 4.2 h | [31] |
| Nitrosomonas europaea ATCC 19718 | NR 1 | >50%, 1 mg/L, 5 days | [32] |
| Proposed Products | RT (min) | M (m/z) | MS/MS (m/z) | Molecular Formula | Structure |
|---|---|---|---|---|---|
| 1,2-Bis(4-hydroxyphenyl)-2-propanol | 7.171~7.212 | 243 | 135, 210, 225 | C15H16O3 |  |
| 4,4′-Dihydroxy-α-methylstilbene | 15.557~15.565 | 225 | 69 | C15H14O2 |  |
| 4-Hydroxybenzaldehyde | 5.700~5.709 | 121 | 92 | C7H6O2 |  |
| 4′-Hydroxyacetophenone | 5.931~5.947 | 135 | 92, 120 | C8H8O2 |  |
| 2,3-Bis(4-hydroxyphenyl)-1,2-propanediol | 3.839~3.873 | 259 | 62, 135 | C15H16O4 |  |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jia, Y.; Eltoukhy, A.; Wang, J.; Li, X.; Hlaing, T.S.; Aung, M.M.; Nwe, M.T.; Lamraoui, I.; Yan, Y. Biodegradation of Bisphenol A by Sphingobium sp. YC-JY1 and the Essential Role of Cytochrome P450 Monooxygenase. Int. J. Mol. Sci. 2020, 21, 3588. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103588
Jia Y, Eltoukhy A, Wang J, Li X, Hlaing TS, Aung MM, Nwe MT, Lamraoui I, Yan Y. Biodegradation of Bisphenol A by Sphingobium sp. YC-JY1 and the Essential Role of Cytochrome P450 Monooxygenase. International Journal of Molecular Sciences. 2020; 21(10):3588. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103588
Chicago/Turabian StyleJia, Yang, Adel Eltoukhy, Junhuan Wang, Xianjun Li, Thet Su Hlaing, Mar Mar Aung, May Thet Nwe, Imane Lamraoui, and Yanchun Yan. 2020. "Biodegradation of Bisphenol A by Sphingobium sp. YC-JY1 and the Essential Role of Cytochrome P450 Monooxygenase" International Journal of Molecular Sciences 21, no. 10: 3588. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103588