Cell Density-Dependent Fibroblast Growth Factor-2 Signaling Regulates Syndecan-4 Expression in Cultured Vascular Endothelial Cells

Abstract
:1. Introduction
2. Results
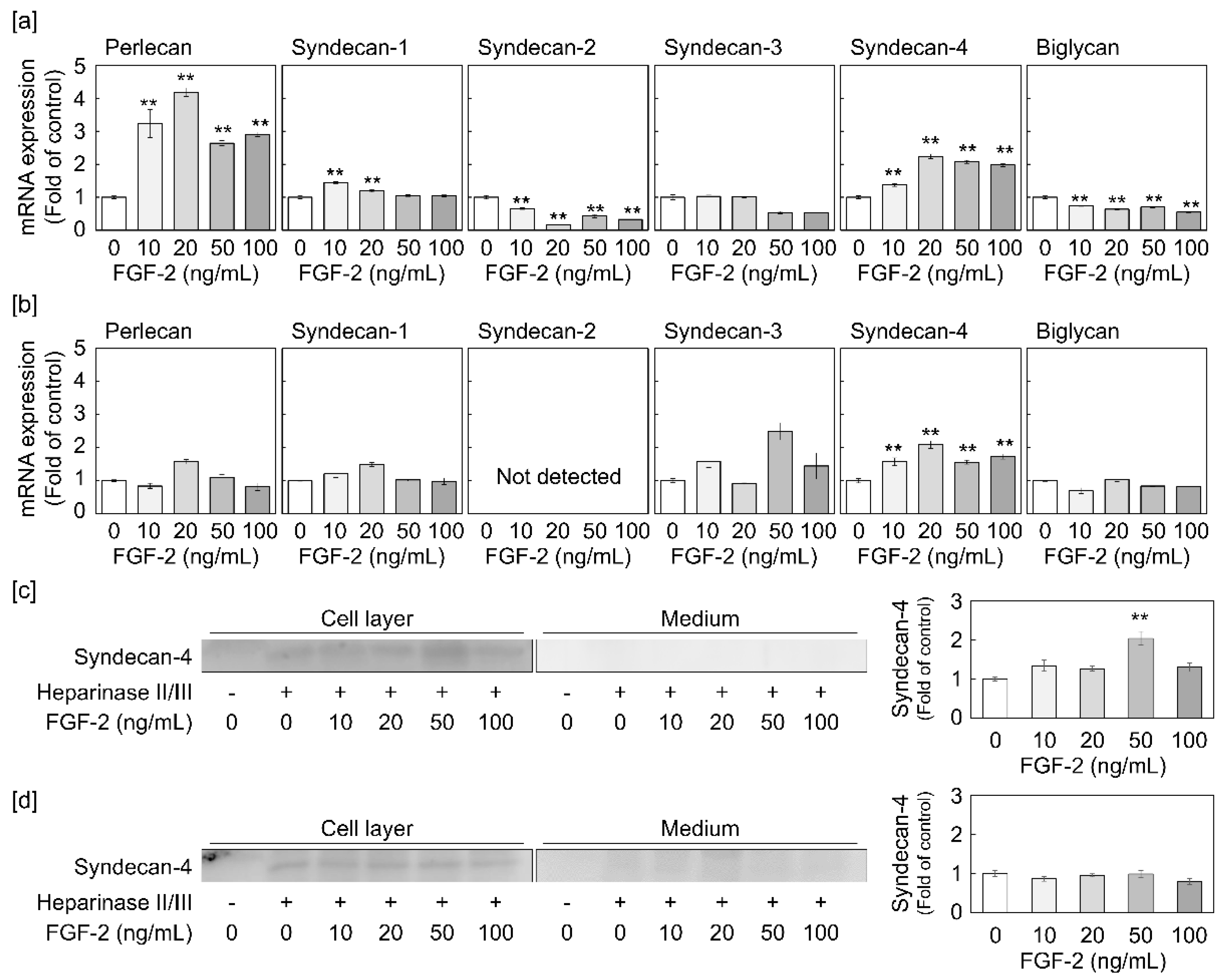
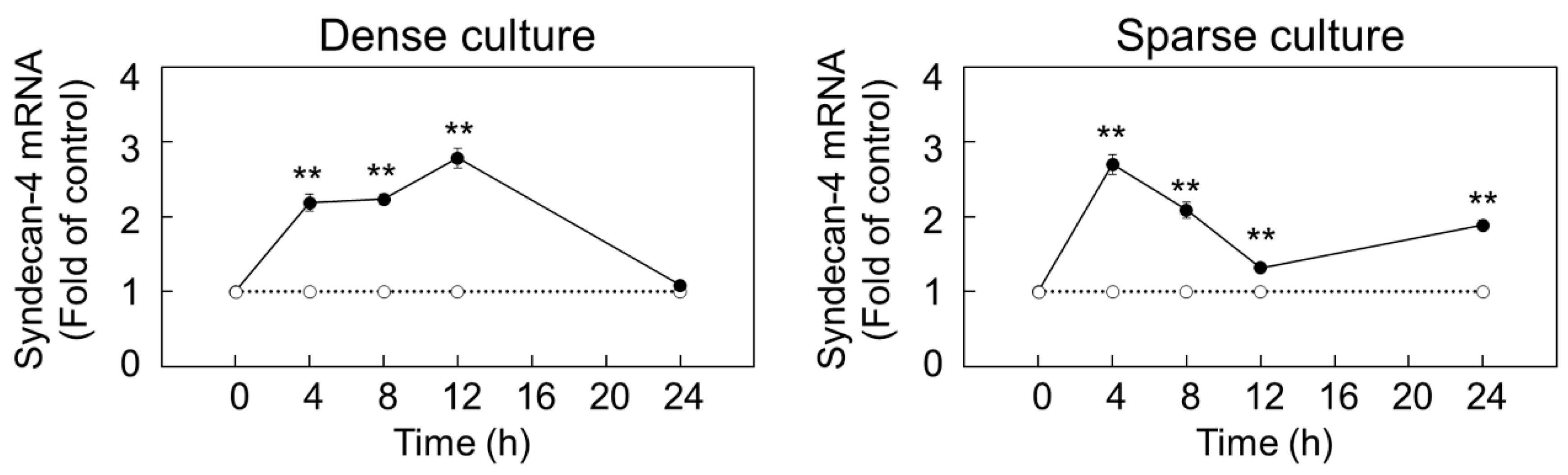
2.1. Syndecan-4 Expression is Induced by FGF-2 in Dense and Sparse Cultures of Vascular Endothelial Cells
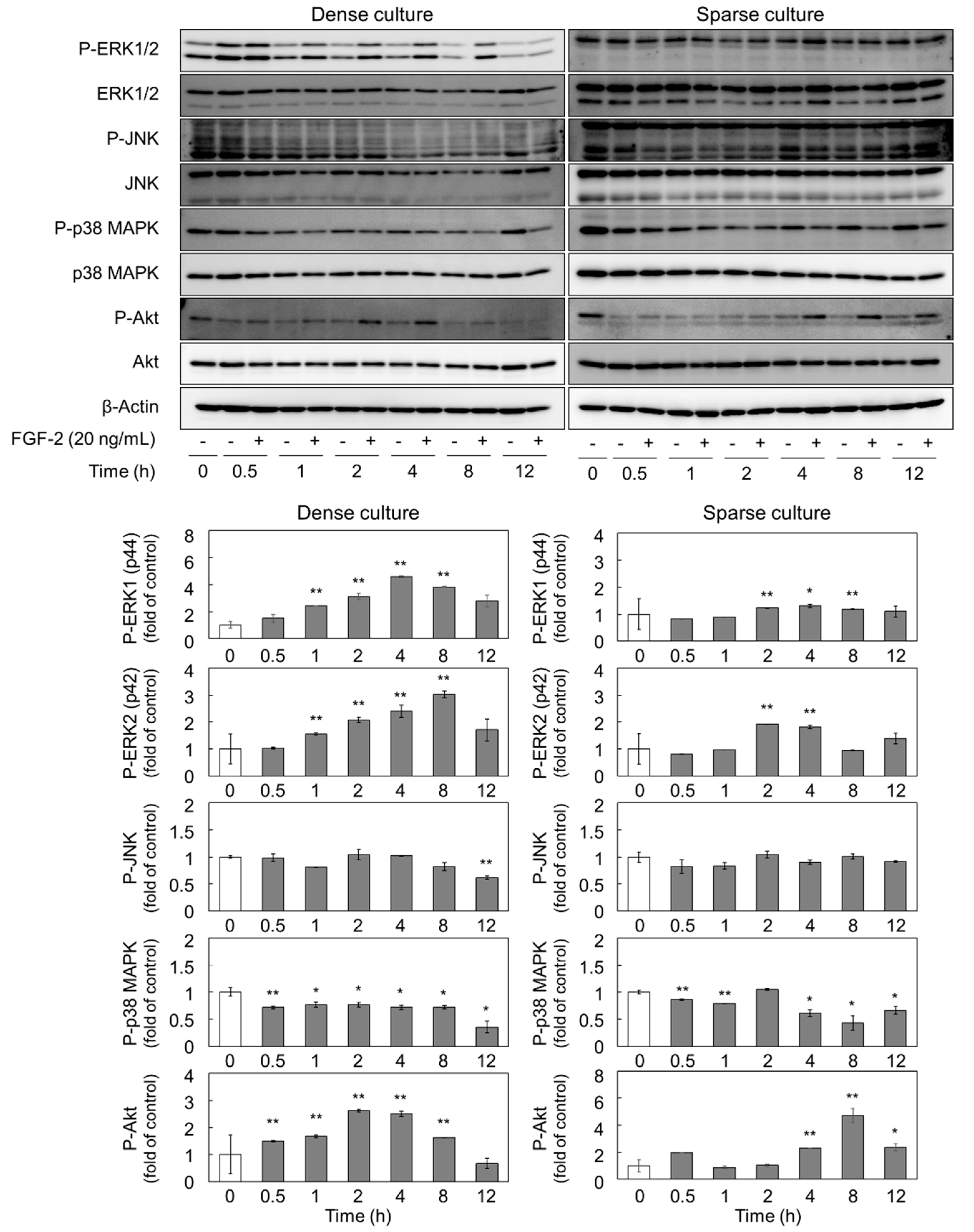
2.2. FGF-2 Activates ERK1/2 and Akt in Dense and Sparse Cultures of Vascular Endothelial Cells
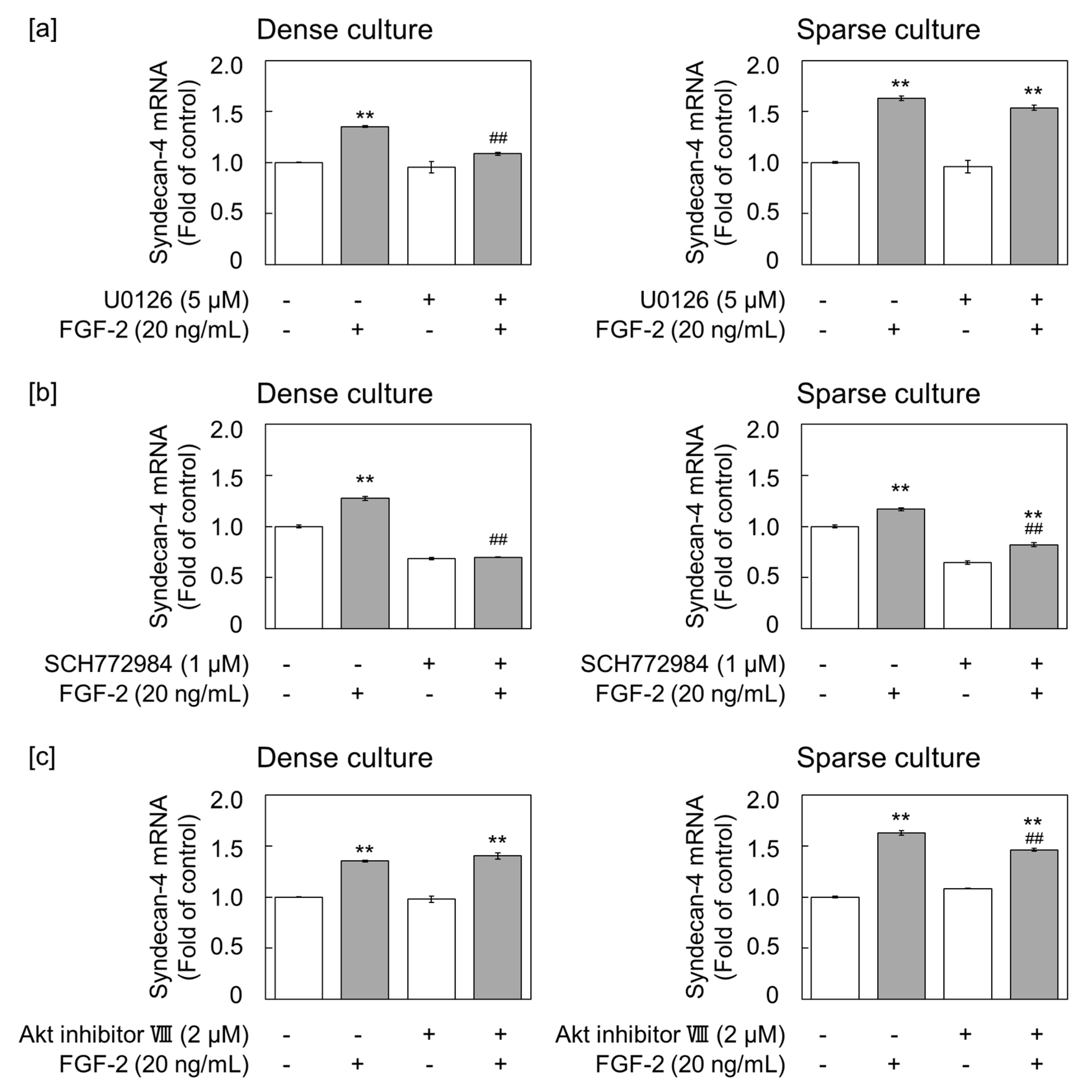
2.3. FGF-2 Induces Syndecan-4 via the ERK1/2 Pathway in Dense Cultures of Vascular Endothelial Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture and Treatments
4.3. Quantitative Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
4.4. Proteoglycan Core Protein Extraction and Western Blot Analysis
4.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| DEAE | Diethylaminoethyl |
| DMEM | Dulbecco’s modified Eagle’s medium |
| ERK | Extracellular signal-regulated kinase |
| FBS | Fetal bovine serum |
| FGF-2 | Fibroblast growth factor-2 |
| GAPDH | Glyceraldehyde 3-phosphate dehydrogenase |
| HRP | Horseradish peroxidase |
| JNK | c-Jun N-terminal kinase |
| MAPK | Mitogen-activated protein kinase |
| PVDF | Polyvinyl difluoride |
| qRT-PCR | Quantitative reverse transcription polymerase chain reaction |
| SDS | Sodium dodecyl sulfate |
| TGF-β | Transforming growth factor-β |
| VEGF | Vascular endothelial growth factor |
References
- Esmon, C.T. Regulation of blood coagulation. Biochim. Biophys. Acta 2000, 1477, 349–360. [Google Scholar] [CrossRef]
- Mehta, D.; Malik, A.B. Signaling mechanisms regulating endothelial permeability. Physiol. Rev. 2006, 86, 279–367. [Google Scholar] [CrossRef] [PubMed]
- Harker, L.A.; Schwartz, S.M.; Ross, R. Endothelium and arteriosclerosis. Clin. Haematol. 1981, 10, 283–296. [Google Scholar] [PubMed]
- Berenson, G.S.; Radhakrishnamurthy, B.; Srinivasan, S.R.; Vijayagopal, P.; Dalferes, E.R., Jr.; Sharma, C. Recent advances in molecular pathology. Carbohydrate-protein macromolecules and arterial wall integrity-a role in atherogenesis. Exp. Mol. Pathol. 1984, 41, 267–287. [Google Scholar] [CrossRef]
- Camejo, G. The interaction of lipids and lipoproteins with the intercellular matrix of arterial tissue: Its possible role in atherogenesis. Adv. Lipid Res. 1982, 19, 1–53. [Google Scholar]
- Rifkin, D.B.; Moscatelli, D. Recent developments in the cell biology of basic fibroblast growth factor. J. Cell Biol. 1989, 109, 1–6. [Google Scholar] [CrossRef]
- Sato, Y.; Rifkin, D.B. Autocrine activities of basic fibroblast growth factor: Regulation of endothelial cell movement, plasminogen activator synthesis, and DNA synthesis. J. Cell Biol. 1988, 107, 1199–1205. [Google Scholar] [CrossRef] [Green Version]
- Assoian, R.K.; Sporn, M.B. Type beta transforming growth factor in human platelets: Release during platelet degranulation and action on vascular smooth muscle cells. J. Cell Biol. 1986, 102, 1217–1223. [Google Scholar] [CrossRef] [Green Version]
- Fràter-Schröder, M.; Müller, G.; Birchmeier, W.; Böhlen, P. Transforming growth factor-beta inhibits endothelial cell proliferation. Biochem. Biophys. Res. Commun. 1986, 137, 295–302. [Google Scholar] [CrossRef]
- Saksela, O.; Moscatelli, D.; Rifkin, D.B. The opposing effects of basic fibroblast growth factor and transforming growth factor beta on the regulation of plasminogen activator activity in capillary endothelial cells. J. Cell Biol. 1987, 105, 957–963. [Google Scholar] [CrossRef] [Green Version]
- Saku, T.; Furthmayr, H. Characterization of the major heparan sulfate proteoglycan secreted by bovine aortic endothelial cells in culture. Homology to the large molecular weight molecule of basement membranes. J. Biol. Chem. 1989, 264, 3514–3523. [Google Scholar] [PubMed]
- Kojima, T.; Shworak, N.W.; Rosenberg, R.D. Molecular cloning and expression of two distinct cDNA-encoding heparan sulfate proteoglycan core proteins from a rat endothelial cell line. J. Biol. Chem. 1992, 267, 4870–4877. [Google Scholar] [PubMed]
- Mertens, G.; Cassiman, J.J.; Van den Berghe, H.; Vermylen, J.; David, G. Cell surface heparan sulfate proteoglycans from human vascular endothelial cells. Core protein characterization and antithrombin III binding properties. J. Biol. Chem. 1992, 267, 20435–20443. [Google Scholar] [PubMed]
- Yamamoto, C.; Deng, X.; Fujiwara, Y.; Kaji, T. Proteoglycans predominantly synthesized by human brain microvascular endothelial cells in culture are perlecan and biglycan. J. Health Sci. 2005, 51, 576–583. [Google Scholar] [CrossRef] [Green Version]
- Couchman, J.R.; Pataki, C.A. An introduction to proteoglycans and their localization. J. Histochem. Cytochem. 2012, 60, 885–897. [Google Scholar] [CrossRef]
- Katsuda, S.; Kaji, T. Atherosclerosis and extracellular matrix. J. Atheroscler. Thromb. 2003, 10, 267–274. [Google Scholar] [CrossRef] [Green Version]
- Evanko, S.P.; Raines, E.W.; Ross, R.; Gold, L.I.; Wight, T.N. Proteoglycan distribution in lesions of atherosclerosis depends on lesion severity, structural characteristics, and the proximity of platelet-derived growth factor and transforming growth factor-beta. Am. J. Pathol. 1998, 152, 533–546. [Google Scholar]
- Kaji, T.; Yamada, A.; Miyajima, S.; Yamamoto, C.; Fujiwara, Y.; Wight, T.N.; Kinsella, M.G. Cell density-dependent regulation of proteoglycan synthesis by transforming growth factor-beta1 in cultured bovine aortic endothelial cells. J. Biol. Chem. 2000, 275, 1463–1470. [Google Scholar] [CrossRef] [Green Version]
- Kaji, T.; Yamamoto, C.; Oh-I, M.; Nishida, T.; Takigawa, M. Differential regulation of biglycan and decorin synthesis by connective tissue growth factor in cultured vascular endothelial cells. Biochem. Biophys. Res. Commun. 2004, 322, 22–28. [Google Scholar] [CrossRef]
- Javerzat, S.; Auguste, P.; Bikfalvi, A. The role of fibroblast growth factors in vascular development. Trends Mol. Med. 2002, 8, 483–489. [Google Scholar] [CrossRef]
- Lim, W.; Bae, H.; Bazer, F.W.; Song, G. Fibroblast growth factor 2 induces proliferation and distribution of G2 /M phase of bovine endometrial cells involving activation of PI3K/AKT and MAPK cell signaling and prevention of effects of ER stress. J. Cell. Physiol. 2018, 233, 3295–3305. [Google Scholar] [CrossRef] [PubMed]
- Kaji, T.; Yamamoto, C.; Oh-I, M.; Fujiwara, Y.; Yamazaki, Y.; Morita, T.; Plaas, A.H.; Wight, T.N. The vascular endothelial growth factor VEGF165 induces perlecan synthesis via VEGF receptor-2 in cultured human brain microvascular endothelial cells. Biochim. Biophys. Acta 2006, 1760, 1465–1474. [Google Scholar] [CrossRef] [PubMed]
- Hara, T.; Yoshida, E.; Shinkai, Y.; Yamamoto, C.; Fujiwara, Y.; Kumagai, Y.; Kaji, T. Biglycan intensifies ALK5-Smad2/3 signaling by TGF-beta1 and downregulates syndecan-4 in cultured vascular endothelial cells. J. Cell. Biochem. 2017, 118, 1087–1096. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, T.; Yoshida, E.; Fujiwara, Y.; Yamamoto, C.; Kaji, T. Transforming growth factor-beta1 modulates the expression of syndecan-4 in cultured vascular endothelial cells in a biphasic manner. J. Cell. Biochem. 2017, 118, 2009–2017. [Google Scholar] [CrossRef] [Green Version]
- Zehe, C.; Engling, A.; Wegehingel, S.; Schafer, T.; Nickel, W. Cell-surface heparan sulfate proteoglycans are essential components of the unconventional export machinery of FGF-2. Proc. Natl. Acad. Sci. USA 2006, 103, 15479–15484. [Google Scholar] [CrossRef] [Green Version]
- Baeyens, N.; Mulligan-Kehoe, M.J.; Corti, F.; Simon, D.D.; Ross, T.D.; Rhodes, J.M.; Wang, T.Z.; Mejean, C.O.; Simons, M.; Humphrey, J.; et al. Syndecan 4 is required for endothelial alignment in flow and atheroprotective signaling. Proc. Natl. Acad. Sci. USA 2014, 111, 17308–17313. [Google Scholar] [CrossRef] [Green Version]
- Woods, A.; Couchman, J.R. Syndecan-4 and focal adhesion function. Curr. Opin. Cell Biol. 2001, 13, 578–583. [Google Scholar] [CrossRef]
- Colliec-Jouault, S.; Shworak, N.W.; Liu, J.; de Agostini, A.I.; Rosenberg, R.D. Characterization of a cell mutant specifically defective in the synthesis of anticoagulantly active heparan sulfate. J. Biol. Chem. 1994, 269, 24953–24958. [Google Scholar]
- Shworak, N.W.; Kojima, T.; Rosenberg, R.D. Isolation and characterization of ryudocan and syndecan heparan sulfate proteoglycans, core proteins, and cDNAs from a rat endothelial cell line. Haemostasis 1993, 23, 161–176. [Google Scholar] [CrossRef]
- Dimou, E.; Cosentino, K.; Platonova, E.; Ros, U.; Sadeghi, M.; Kashyap, P.; Katsinelos, T.; Wegehingel, S.; Noe, F.; Garcia-Saez, A.J.; et al. Single event visualization of unconventional secretion of FGF2. J. Cell Biol. 2018, 218, 683–699. [Google Scholar] [CrossRef] [Green Version]
- Dimou, E.; Nickel, W. Unconventional mechanisms of eukaryotic protein secretion. Curr. Biol. 2018, 28, R406–R410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, Y.; Murphy, P.R.; Sato, R.; Friesen, H.G. Fibroblast growth factor release by bovine endothelial cells and human astrocytoma cells in culture is density dependent. Mol. Endocrinol. 1989, 3, 744–748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richardson, T.P.; Trinkaus-Randall, V.; Nugent, M.A. Regulation of basic fibroblast growth factor binding and activity by cell density and heparan sulfate. J. Biol. Chem. 1999, 274, 13534–13540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsui, Y.; Ikesue, M.; Danzaki, K.; Morimoto, J.; Sato, M.; Tanaka, S.; Kojima, T.; Tsutsui, H.; Uede, T. Syndecan-4 prevents cardiac rupture and dysfunction after myocardial infarction. Circ. Res. 2011, 108, 1328–1339. [Google Scholar] [CrossRef] [Green Version]
- Ikesue, M.; Matsui, Y.; Ohta, D.; Danzaki, K.; Ito, K.; Kanayama, M.; Kurotaki, D.; Morimoto, J.; Kojima, T.; Tsutsui, H.; et al. Syndecan-4 deficiency limits neointimal formation after vascular injury by regulating vascular smooth muscle cell proliferation and vascular progenitor cell mobilization. Arter. Thromb. Vasc. Biol. 2011, 31, 1066–1074. [Google Scholar] [CrossRef]
- Jee, S.H.; Chu, C.Y.; Chiu, H.C.; Huang, Y.L.; Tsai, W.L.; Liao, Y.H.; Kuo, M.L. Interleukin-6 induced basic fibroblast growth factor-dependent angiogenesis in basal cell carcinoma cell line via JAK/STAT3 and PI3-kinase/Akt pathways. J. Invest. Derm. 2004, 123, 1169–1175. [Google Scholar] [CrossRef] [Green Version]
- Jiang, S.; Li, Y.; Lin, T.; Yuan, L.; Li, Y.; Wu, S.; Xia, L.; Shen, H.; Lu, J. IL-35 inhibits angiogenesis through VEGF/Ang2/Tie2 pathway in rheumatoid arthritis. Cell. Physiol. Biochem. 2016, 40, 1105–1116. [Google Scholar] [CrossRef]
- Buscà, R.; Pouysségur, J.; Lenormand, P. ERK1 and ERK2 map kinases: Specific roles or functional redundancy? Front. Cell Dev. Biol. 2016, 4, 53. [Google Scholar] [CrossRef] [Green Version]
- Kinsella, M.G.; Tsoi, C.K.; Jarvelainen, H.T.; Wight, T.N. Selective expression and processing of biglycan during migration of bovine aortic endothelial cells. The role of endogenous basic fibroblast growth factor. J. Biol. Chem. 1997, 272, 318–325. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Wu, H.; Xie, J.; Li, G.; Gu, R.; Kang, L.; Wang, L.; Xu, B. Syndecan-4 regulates the bFGF-induced chemotactic migration of endothelial cells. J. Mol. Histol. 2016, 47, 503–509. [Google Scholar] [CrossRef]
- Das, S.; Monteforte, A.J.; Singh, G.; Majid, M.; Sherman, M.B.; Dunn, A.K.; Baker, A.B. Syndecan-4 enhances therapeutic angiogenesis after hind limb ischemia in mice with type 2 diabetes. Adv. Healthc. Mater. 2016, 5, 1008–1013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, T.; Kojima, T.; Matsuzaki, H.; Nakamura, T.; Yoshida, E.; Fujiwara, Y.; Yamamoto, C.; Saito, S.; Kaji, T. Induction of syndecan-4 by organic-inorganic hybrid molecules with a 1,10-phenanthroline structure in cultured vascular endothelial cells. Int. J. Mol. Sci. 2017, 18, 352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hara, T.; Tatsuishi, H.; Banno, T.; Fujie, T.; Yamamoto, C.; Naka, H.; Kaji, T. Copper (II) bis (diethyldithiocarbamate) induces the expression of syndecan-4, a transmembrane heparan sulfate proteoglycan, via p38 MAPK activation in vascular endothelial cells. Int. J. Mol. Sci. 2018, 19, 3302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.; Kwon, M.J.; Lim, Y.; Yun, J.H.; Lee, W.; Oh, E.S. Trans-regulation of syndecan functions by hetero-oligomerization. J. Biol. Chem. 2015, 290, 16943–16953. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′–3′) | Reverse Primer (5′–3′) |
|---|---|---|
| Perlecan | GCTGAGGGCGTACGATGG | TGCCCAGGCGTCGGAACT |
| Syndecan-1 | CAGTCAGGAGACAGCATCAG | CCGACAGACATTCCATACC |
| Syndecan-2 | CCAGATGAAGAGGACACAAACG | CCAATAACTCCGCCAGCAA |
| Syndecan-3 | CAAGCAGGCGAGCGTC | GGTGGCAGAGATGAAGTGG |
| Syndecan-4 | TTGCCGTCTTCCTCGTGC | AGGCGTAGAACTCATTGGTGG |
| Biglycan | GCTGCCACTGCCATCTGAG | CGAGGACCAAGGCGTAG |
| GAPDH | AACACCCTCAAGATTGTCAGCAA | ACAGTCTTCTGGGTGGCAGTGA |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hara, T.; Yabushita, S.; Yamamoto, C.; Kaji, T. Cell Density-Dependent Fibroblast Growth Factor-2 Signaling Regulates Syndecan-4 Expression in Cultured Vascular Endothelial Cells. Int. J. Mol. Sci. 2020, 21, 3698. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103698
Hara T, Yabushita S, Yamamoto C, Kaji T. Cell Density-Dependent Fibroblast Growth Factor-2 Signaling Regulates Syndecan-4 Expression in Cultured Vascular Endothelial Cells. International Journal of Molecular Sciences. 2020; 21(10):3698. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103698
Chicago/Turabian StyleHara, Takato, Shiori Yabushita, Chika Yamamoto, and Toshiyuki Kaji. 2020. "Cell Density-Dependent Fibroblast Growth Factor-2 Signaling Regulates Syndecan-4 Expression in Cultured Vascular Endothelial Cells" International Journal of Molecular Sciences 21, no. 10: 3698. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21103698