Paclitaxel Induces Upregulation of Transient Receptor Potential Vanilloid 1 Expression in the Rat Spinal Cord

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of PTX on Mechanical Allodynia/Hyperalgesia
2.2. Effect of PTX on Thermal Hyperalgesia
2.3. Effect of PTX Treatment on TRPV1 Protein Expression in Spinal Cord
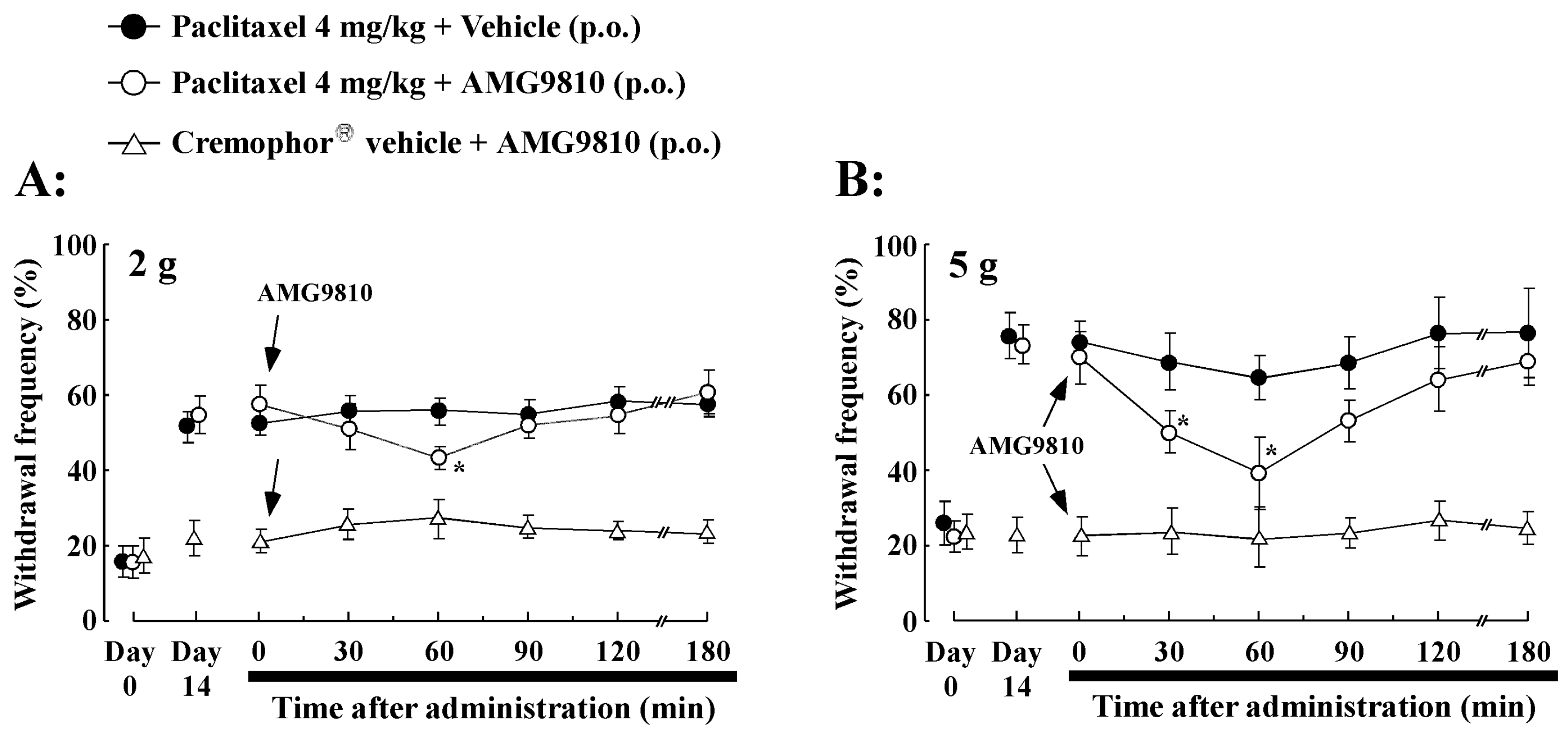
2.4. Effect of Oral Administration of TRPV1 Antagonist (AMG9810) on PTX-Induced Mechanical Allodynia/Hyperalgesia
2.5. Effect of Intrathecal Administration of TRPV1 Antagonist (AMG9810) on PTX-Induced Mechanical Allodynia/Hyperalgesia and Thermal Hyperalgesia
2.6. Effect of Intrathecal Administration of TRPV1 Small Interfering RNA (siRNA) on PTX-Induced Peripheral Neuropathy
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Drug Administration
4.3. Behavioral Observation (Mechanical or Thermal Stimulation)
4.4. Western Blot Analysis
4.5. Immunohistochemistry
4.6. Image Analysis
4.7. Intrathecal Injection of TRPV1 siRNA or TRPV1 Antagonist (AMG9810)
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Hagiwara, H.; Sunada, Y. Mechanism of taxane neurotoxicity. Breast Cancer 2004, 11, 82–85. [Google Scholar] [CrossRef]
- Chaudhry, V.; Rowinsky, E.K.; Sartorius, S.E.; Donehower, R.C.; Cornblath, D.R. Peripheral neuropathy from taxol and cisplatin combination chemotherapy: Clinical and electrophysiological studies. Ann. Neurol. 1994, 35, 304–311. [Google Scholar] [CrossRef]
- Sisignano, M.; Baron, R.; Scholich, K.; Geisslinger, G. Mechanism-based treatment for chemotherapy-induced peripheral neuropathic pain. Nat. Rev. Neurol. 2014, 10, 694–707. [Google Scholar] [CrossRef]
- Forsyth, P.A.; Balmaceda, C.; Peterson, K.; Seidman, A.D.; Brasher, P.; DeAngelis, L.M. Prospective study of paclitaxel-induced peripheral neuropathy with quantitative sensory testing. J. Neurooncol. 1997, 35, 47–53. [Google Scholar] [CrossRef]
- Dougherty, P.M.; Cata, J.P.; Cordella, J.V.; Burton, A.; Weng, H.R. Taxol-induced sensory disturbance is characterized by preferential impairment of myelinated fiber function in cancer patients. Pain 2004, 109, 132–142. [Google Scholar] [CrossRef]
- Wickham, R. Chemotherapy-induced peripheral neuropathy: A review and implications for oncology nursing practice. Clin. J. Oncol. Nurs. 2007, 11, 361–376. [Google Scholar] [CrossRef] [Green Version]
- Ledeboer, A.; Jekich, B.M.; Sloane, E.M.; Mahoney, J.H.; Langer, S.J.; Milligan, E.D.; Martin, D.; Maier, S.F.; Johnson, K.W.; Leinwand, L.A.; et al. Intrathecal interleukin-10 gene therapy attenuates paclitaxel-induced mechanical allodynia and proinflammatory cytokine expression in dorsal root ganglia in rats. Brain Behav. Immun. 2007, 21, 686–698. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Yang, C.; Wang, Z.J. Proteinase-activated receptor 2 sensitizes transient receptor potential vanilloid 1, transient receptor potential vanilloid 4, and transient receptor potential ankyrin 1 in paclitaxel-induced neuropathic pain. Neuroscience 2011, 193, 440–451. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, H.; Zhang, H.; Kosturakis, A.K.; Jawad, A.B.; Dougherty, P.M. Toll-like receptor 4 signaling contributes to paclitaxel-induced peripheral neuropathy. J. Pain 2014, 15, 712–725. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Bavencoffe, A.; Yang, P.; Feng, J.; Yin, S.; Qian, A.; Yu, W.; Liu, S.; Gong, X.; Cai, T.; et al. Zinc Inhibits TRPV1 to Alleviate Chemotherapy-Induced Neuropathic Pain. J. Neurosci. 2018, 38, 474–483. [Google Scholar] [CrossRef]
- Staaf, S.; Oerther, S.; Lucas, G.; Mattsson, J.P.; Ernfors, P. Differential regulation of TRP channels in a rat model of neuropathic pain. Pain 2009, 14, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, K.; Chiba, T.; Katagiri, N.; Saduka, M.; Abe, K.; Utsunomiya, I.; Hama, T.; Taguchi, K. Paclitaxel increases high voltage-dependent calcium channel current in dorsal root ganglion neurons of the rat. J. Pharmacol. Sci. 2012, 120, 187–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, C.M.; Jimenez-Andrade, J.M.; Jonas, B.M.; Sevcik, M.A.; Koewler, N.J.; Ghilardi, J.R.; Wong, G.Y.; Mantyh, P.W. Intravenous paclitaxel administration in the rat induces a peripheral sensory neuropathy characterized by macrophage infiltration and injury to sensory neurons and their supporting cells. Exp. Neurol. 2007, 203, 42–54. [Google Scholar] [CrossRef] [PubMed]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Sasamura, T.; Kuraishi, Y. Peripheral and central actions of capsaicin and VR1 receptor. Jpn. J. Pharmacol. 1999, 80, 275–280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyette-Davis, J.A.; Hou, S.; Abdi, S.; Dougherty, P.M. An updated understanding of the mechanisms involved in chemotherapy-induced neuropathy. Pain Manag. 2018, 8, 363–375. [Google Scholar] [CrossRef]
- Kissin, I. Vanilloid-induced conduction analgesia: Selective, dose-dependent, long-lasting, with a low level of potential neurotoxicity. Anesth. Analg. 2008, 107, 271–281. [Google Scholar] [CrossRef] [Green Version]
- Spicarova, D.; Nerandzic, V.; Palecek, J. Update on the role of spinal cord TRPV1 receptors in pain modulation. Physiol. Res. 2014, 63, S225–S236. [Google Scholar]
- Yu, L.; Yang, F.; Luo, H.; Liu, F.Y.; Han, J.S.; Xing, G.G.; Wan, Y. The role of TRPV1 in different subtypes of dorsal root ganglion neurons in rat chronic inflammatory nociception induced by complete Freund’s adjuvant. Mol. Pain 2008, 4. [Google Scholar] [CrossRef] [Green Version]
- Ta, L.E.; Bieber, A.J.; Carlton, S.M.; Loprinzi, C.L.; Low, P.A.; Windebank, A.J. Transient Receptor Potential Vanilloid 1 is essential for cisplatin-induced heat hyperalgesia in mice. Mol. Pain 2010, 6. [Google Scholar] [CrossRef] [Green Version]
- Tohda, C.; Sasaki, M.; Konemura, T.; Sasamura, T.; Itoh, M.; Kuraishi, Y. Axonal transport of VR1 capsaicin receptor mRNA in primary afferents and its participation in inflammation-induced increase in capsaicin sensitivity. J. Neurochem. 2001, 76, 1628–1635. [Google Scholar] [CrossRef]
- Luo, H.; Cheng, J.; Han, J.S.; Wan, Y. Change of vanilloid receptor 1 expression in dorsal root ganglion and spinal dorsal horn during inflammatory nociception induced by complete Freund’s adjuvant in rats. Neuroreport 2004, 15, 655–658. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalynovska, N.; Adamek, P.; Palecek, J. TRPV1 receptors contribute to paclitaxel-induced c-Fos expression in spinal cord dorsal horn neurons. Physiol. Res. 2017, 66, 549–552. [Google Scholar] [CrossRef] [PubMed]
- Christoph, T.; Grünweller, A.; Mika, J.; Schäfer, M.K.; Wade, E.J.; Weihe, E.; Erdmann, V.A.; Frank, R.; Gillen, C.; Kurreck, J. Silencing of vanilloid receptor TRPV1 by RNAi reduces neuropathic and visceral pain in vivo. Biochem. Biophys. Res. Commun. 2006, 350, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Matta, J.A.; Ahern, G.P. Voltage is a partial activator of rat thermosensitive TRP channels. J. Physiol. 2007, 585, 469–482. [Google Scholar] [CrossRef]
- Hara, T.; Chiba, T.; Abe, K.; Makabe, A.; Ikeno, S.; Kawakami, K.; Utsunomiya, I.; Hama, T.; Taguchi, K. Effect of paclitaxel on transient receptor potential vanilloid 1 in rat dorsal root ganglion. Pain 2013, 154, 882–889. [Google Scholar] [CrossRef]
- Spicarov, D.; Palecek, J. The Role of Spinal Cord Vanilloid (TRPV1) Receptors in Pain Modulation. Physiol. Res. 2008, 57, S69–S77. [Google Scholar]
- Chiba, T.; Oka, Y.; Kambe, T.; Koizumi, N.; Abe, K.; Kawakami, K.; Utsunomiya, I.; Taguchi, K. Paclitaxel-induced peripheral neuropathy increases substance P release in rat spinal cord. Eur. J. Pharmacol. 2016, 770, 46–51. [Google Scholar] [CrossRef]
- Matsumoto, M.; Inoue, M.; Hald, A.; Xie, W.; Ueda, H. Inhibition of paclitaxel-induced A-fiber hypersensitization by gabapentin. J. Pharmacol. Exp. Ther. 2006, 318, 735–740. [Google Scholar] [CrossRef]
- Segat, G.C.; Manjavachi, M.N.; Matias, D.O.; Passos, G.F.; Freitas, C.S.; Costa, R.; Calixto, J.B. Antiallodynic effect of β-caryophyllene on paclitaxel-induced peripheral neuropathy in mice. Neuropharmacology 2017, 125, 207–219. [Google Scholar] [CrossRef]
- Caterina, M.J. Vanilloid receptors take a TRP beyond the sensory afferent. Pain 2003, 105, 5–9. [Google Scholar] [CrossRef]
- Kissin, I.; Szallasi, A. Therapeutic Targeting of TRPV1 by Resiniferatoxin, from Preclinical Studies to Clinical Trials. Curr. Top. Med. Chem. 2011, 11, 2159–2170. [Google Scholar] [CrossRef] [PubMed]
- Spicarova, D.; Palecek, J. The role of the TRPV1 endogenous agonist. N-Oleoyldopamine in modulation of nociceptive signaling at the spinal cord level. J. Neurophysiol. 2009, 102, 234–243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Adamek, P.; Zhang, H.; Tatsui, C.E.; Rhines, L.D.; Mrozkova, P.; Li, Q.; Kosturakis, A.K.; Cassidy, R.M.; Harrison, D.S.; et al. The cancer chemotherapeutic paclitaxel increases human and rodent sensory neuron responses to TRPV1 by activation of TLR4. J. Neurosci. 2015, 35, 13487–13500. [Google Scholar] [CrossRef]
- Valtschanoff, J.G.; Rustioni, A.; Guo, A.; Hwang, S.J. Vanilloid receptor VR1 is both presynaptic and postsynaptic in the superficial laminae of the rat dorsal horn. J. Comp. Neurol. 2001, 436, 225–235. [Google Scholar] [CrossRef]
- Tominaga, M.; Caterina, M.J.; Malmberg, A.B.; Rosen, T.A.; Gilbert, H.; Skinner, K.; Raumann, B.E.; Basbaum, A.I.; Julius, D. The cloned capsaicin receptor integrates multiple pain-producing stimuli. Neuron 1998, 21, 531–543. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.R.; Kim, S.U.; Oh, U.; Jin, B.K. Transient receptor potential vanilloid subtype 1 mediates microglial cell death in vivo and in vitro via Ca2+-mediated mitochondrial damage and cytochrome c release. J. Immunol. 2006, 177, 4322–4329. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Willcockson, H.H.; Valtschanoff, J.G. Influence of the vanilloid receptor TRPV1 on the activation of spinal cord glia in mouse models of pain. Exp. Neurol. 2009, 220, 383–390. [Google Scholar] [CrossRef] [Green Version]
- Guo, A.; Vulchanova, L.; Wang, J.; Li, X.; Elde, R. Immunocytochemical localization of the vanilloid receptor 1 (VR1): Relationship to neuropeptides, the P2X3 purinoceptor and IB4 binding sites. Eur. J. Neurosci. 1999, 11, 946–958. [Google Scholar] [CrossRef]
- Mantyh, P.W.; Rogers, S.D.; Honore, P.; Allen, B.J.; Ghilardi, J.R.; Li, J.; Daughters, R.S.; Lappi, D.A.; Wiley, R.G.; Simone, D.A. Inhibition of hyperalgesia by ablation of lamina I spinal neurons expressing the substance P receptor. Science 1997, 278, 275–279. [Google Scholar] [CrossRef]
- Hudson, L.J.; Bevan, S.; Wotherspoon, G.; Gentry, C.; Fox, A.; Winter, J. VR1 protein expression increases in undamaged DRG neurons after partial nerve injury. Eur. J. Neurosci. 2001, 13, 2105–2114. [Google Scholar] [CrossRef] [PubMed]
- Rashid, M.H.; Inoue, M.; Kondo, S.; Kawashima, T.; Bakoshi, S.; Ueda, H. Novel expression of vanilloid receptor 1 on capsaicin-insensitive fibers accounts for the analgesic effect of capsaicin cream in neuropathic pain. J. Pharmacol. Exp. Ther. 2003, 304, 940–948. [Google Scholar] [CrossRef] [PubMed]
- Gavva, N.R.; Tamir, R.; Qu, Y.; Klionsky, L.; Zhang, T.J.; Immke, D.; Wang, J.; Zhu, D.; Vanderah, T.W.; Porreca, F.; et al. AMG 9810 [(E)-3-(4-t-butylphenyl)-N-(2,3-dihydrobenzo[b][1,4] dioxin-6-yl)acrylamide], a novel vanilloid receptor 1 (TRPV1) antagonist with antihyperalgesic properties. J. Pharmacol. Exp. Ther. 2005, 313, 474–484. [Google Scholar] [CrossRef] [PubMed]
- Kanai, Y.; Nakazato, E.; Fujiuchi, A.; Hara, T.; Imai, A. Involvement of an increased spinal TRPV1 sensitization through its up-regulation in mechanical allodynia of CCI rats. Neuropharmacology 2005, 49, 977–984. [Google Scholar] [CrossRef]
- Kanai, Y.; Hara, T.; Imai, A.; Sakakibara, A. Differential involvement of TRPV1 receptors at the central and peripheral nerves in CFA-induced mechanical and thermal hyperalgesia. J. Pharm. Pharmacol. 2007, 59, 733–738. [Google Scholar] [CrossRef]
- Cui, M.; Honore, P.; Zhong, C.; Gauvin, D.; Mikusa, J.; Hernandez, G.; Chandran, P.; Gomtsyan, A.; Brown, B.; Bayburt, E.K.; et al. TRPV1 receptors in the CNS play a key role in broad-spectrum analgesia of TRPV1 antagonists. J. Neurosci. 2006, 26, 9385–9393. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Yang, Q.; Crook, R.J.; O’Neil, R.G.; Walters, E.T. TRPV1 channels make major contributions to behavioral hypersensitivity and spontaneous activity in nociceptors after spinal cord injury. Pain 2013, 154, 2130–2141. [Google Scholar] [CrossRef]
- Luo, M.C.; Zhang, D.Q.; Ma, S.W.; Huang, Y.Y.; Shuster, S.J.; Porreca, F.; Lai, J. An efficient intrathecal delivery of small interfering RNA to the spinal cord and peripheral neurons. Mol. Pain 2005, 1, 29. [Google Scholar] [CrossRef] [Green Version]
- Wu, F.X.; Bian, J.J.; Miao, X.R.; Huang, S.D.; Xu, X.W.; Gong, D.J.; Sun, Y.M.; Lu, Z.J.; Yu, W.F. Intrathecal siRNA Against Toll-like Receptor 4 Reduces Nociception in a Rat Model of Neuropathic Pain. Int. J. Med. Sci. 2010, 7, 251–259. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Chiba, N.; Chiba, T.; Kambe, T.; Abe, K.; Kawakami, K.; Utsunomiya, I.; Taguchi, K. Transient receptor potential ankyrin 1 that is induced in dorsal root ganglion neurons contributes to acute cold hypersensitivity after oxaliplatin administration. Mol. Pain 2015, 11, 69. [Google Scholar] [CrossRef] [Green Version]
- Caterina, M.J.; Leffler, A.; Malmberg, A.B.; Martin, W.J.; Trafton, J.; Petersen-Zeitz, K.R.; Koltzenburg, M.; Basbaum, A.I.; Julius, D. Impaired nociception and pain sensation in mice lacking the capsaicin receptor. Science 2000, 288, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.B.; Gray, J.; Gunthorpe, M.J.; Hatcher, J.P.; Davey, P.T.; Overend, P.; Harries, M.H.; Latcham, J.; Clapham, C.; Atkinson, K.; et al. Vanilloid receptor-1 is essential for inflammatory thermal hyperalgesia. Nature 2000, 405, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Christoph, T.; Gillen, C.; Mika, J.; Grünweller, A.; Schäfer, M.K.; Schiene, K.; Frank, R.; Jostock, R.; Bahrenberg, G.; Weihe, E.; et al. Antinociceptive effect of antisense oligonucleotides against the vanilloid receptor VR1/TRPV1. Neurochem. Int. 2007, 50, 281–290. [Google Scholar] [CrossRef] [PubMed]






© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kamata, Y.; Kambe, T.; Chiba, T.; Yamamoto, K.; Kawakami, K.; Abe, K.; Taguchi, K. Paclitaxel Induces Upregulation of Transient Receptor Potential Vanilloid 1 Expression in the Rat Spinal Cord. Int. J. Mol. Sci. 2020, 21, 4341. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124341
Kamata Y, Kambe T, Chiba T, Yamamoto K, Kawakami K, Abe K, Taguchi K. Paclitaxel Induces Upregulation of Transient Receptor Potential Vanilloid 1 Expression in the Rat Spinal Cord. International Journal of Molecular Sciences. 2020; 21(12):4341. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124341
Chicago/Turabian StyleKamata, Yukako, Toshie Kambe, Terumasa Chiba, Ken Yamamoto, Kazuyoshi Kawakami, Kenji Abe, and Kyoji Taguchi. 2020. "Paclitaxel Induces Upregulation of Transient Receptor Potential Vanilloid 1 Expression in the Rat Spinal Cord" International Journal of Molecular Sciences 21, no. 12: 4341. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21124341