GANT61 and Lithium Chloride Inhibit the Growth of Head and Neck Cancer Cell Lines Through the Regulation of GLI3 Processing by GSK3β

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. The HH-GLI Signaling Pathway Is Active in HNSCC Cell Lines
2.2. HNSCC Cell Lines Respond to Downstream Inhibition More Efficiently Than to Upstream Inhibition
2.3. GANT61 and LiCl Regulate GLI3 Protein Levels in HNSCC
2.4. The Effect of LiCl Inhibition is Mediated by GLI3 Processing by GSK3β
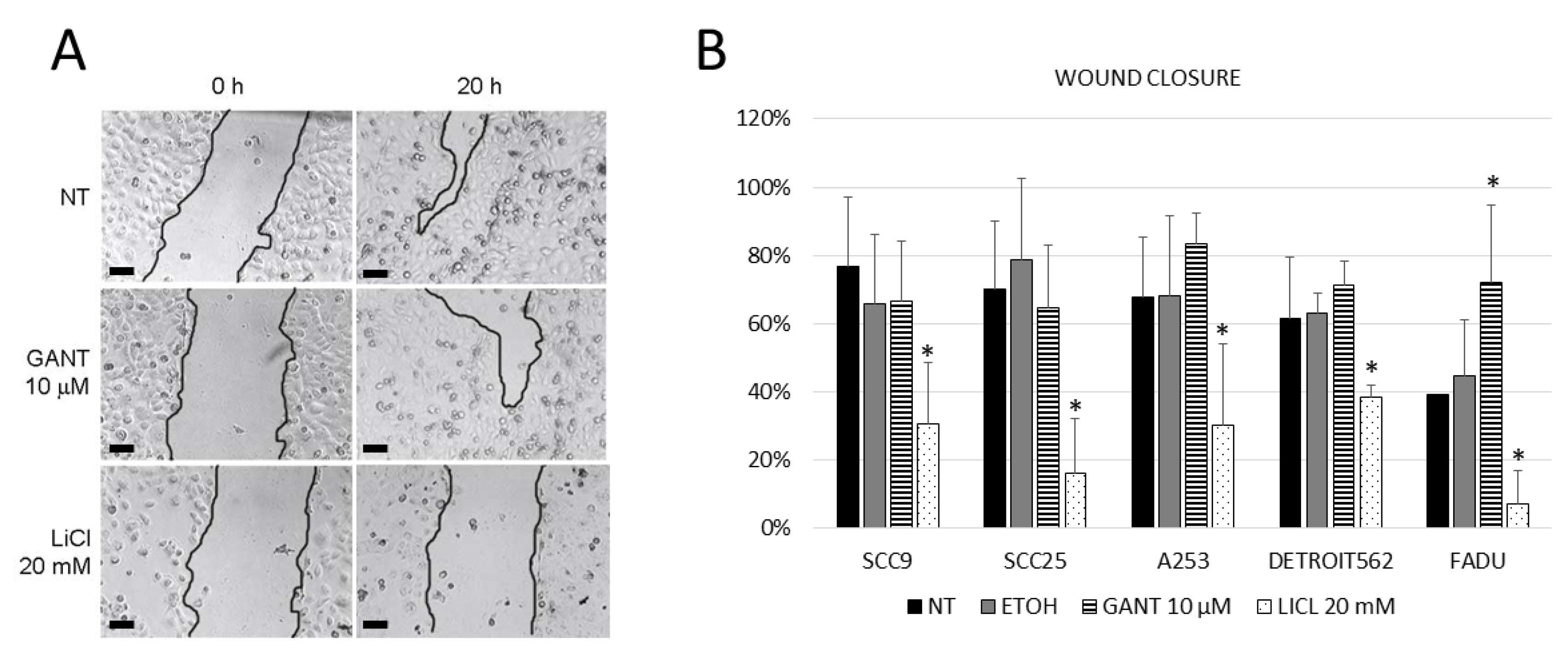
2.5. Downstream Inhibition Affects Colony Forming and Wound Closure Capabilities of HNSCC Cell Lines
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Cell Assays
4.2. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.3. Western Blotting
4.4. Immunoprecipitation and Coomassie Staining
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| HH-GLI | Hedgehog-Gli |
| LiCl | Lithium chloride |
| HNSCC | Head and neck squamous cell carcinoma |
| HPV16 | Human papilloma virus 16 |
| CSC | Cancer stem cells |
| EGFR | Epidermal growth factor receptor |
| PI3K | Phosphoinositol 3 kinase |
| VEGF | Vascular endothelial growth factor |
| PTCH1 | Patched 1 |
| HH | Hedgehog |
| SMO | Smoothened |
| βTrCP1 | Beta-transducin repeat containing protein 1 |
| GSK3β | Glycogen synthase 3 beta |
| PKA | Protein kinase A |
| SUFU | Suppressor of fused |
| CK1 | Casein kinase 1 |
| GLI | Glioma-associated oncogene homolog |
| EMT | Epithelial-mesenchymal transition |
| NQC | Nanoquinacrine |
| DMEM:HAM’S | Dulbecco’s Modified Eagle Medium: Ham’s F12 |
| EMEM | Eagle’s Minimum Essential Medium |
| FBS | Fetal bovine Serum |
| MTT | 3-(4,5-Dimethylthiazol-2-yl)-2,5-Diphenyltetrazolium Bromide |
| PBS | Phosphate Buffered Saline |
| RIPA | Radioimmunoprecipitation assay buffer |
| BCA | Bicinchonic Acid |
| PAA | Polyacrylamide |
| TBST | Tris-Buffered Saline, 0.1% Tween® 20 Detergent |
| HRP | Horseradish Peroxidase |
| TENN | 50 mM Tris, 5 mM EDTA, 150 mM NaCl, 0.5% NP-40, pH 8.0 |
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jou, A.; Hess, J. Epidemiology and Molecular Biology of Head and Neck Cancer. Oncol. Res. Treat. 2017, 40, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Elkashty, O.A.; Ashry, R.; Tran, S.D.; Elkashty, O. Head and neck cancer management and cancer stem cells implication. Saudi Dent. J. 2019, 31, 395–416. [Google Scholar] [CrossRef] [PubMed]
- Gillison, M.L.; D’Souza, G.; Westra, W.; Sugar, E.; Xiao, W.; Begum, S.; Viscidi, R. Distinct Risk Factor Profiles for Human Papillomavirus Type 16–Positive and Human Papillomavirus Type 16–Negative Head and Neck Cancers. J. Natl. Cancer Inst. 2008, 100, 407–420. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, A.K.; Engels, E.A.; Pfeiffer, R.M.; Hernandez, B.Y.; Xiao, W.; Kim, E.; Jiang, B.; Goodman, M.T.; Sibug-Saber, M.; Cozen, W.; et al. Human Papillomavirus and Rising Oropharyngeal Cancer Incidence in the United States. J. Clin. Oncol. 2011, 29, 4294–4301. [Google Scholar] [CrossRef]
- Božinović, K.; Sabol, I.; Rakusic, Z.; Jakovčević, A.; Šekerija, M.; Lukinović, J.; Prgomet, D.; Grce, M. HPV-driven oropharyngeal squamous cell cancer in Croatia — Demography and survival. PLoS ONE 2019, 14, e0211577. [Google Scholar] [CrossRef] [Green Version]
- Leović, D.; Sabol, M.; Ozretic, P.; Musani, V.; Car, D.; Marjanovic, K.; Zubcic, V.; Sabol, I.; Sikora, M.; Grce, M.; et al. Hh-Gli signaling pathway activity in oral and oropharyngeal squamous cell carcinoma. Head Neck 2011, 34, 104–112. [Google Scholar] [CrossRef]
- Chen, D.; Wang, C.-Y. Targeting cancer stem cells in squamous cell carcinoma. Precis. Clin. Med. 2019, 2, 152–165. [Google Scholar] [CrossRef]
- Cristina, V.; Herrera-Gómez, R.G.; Szturz, P.; Espeli, V.; Siano, M. Immunotherapies and Future Combination Strategies for Head and Neck Squamous Cell Carcinoma. Int. J. Mol. Sci. 2019, 20, 5399. [Google Scholar] [CrossRef] [Green Version]
- Bowles, D.W.; McDermott, J.D.; Jimeno, A. Novel treatments for head and neck squamous cell carcinoma: Preclinical identification and clinical investigation. Future Oncol. 2014, 10, 1065–1080. [Google Scholar] [CrossRef]
- Sabol, M.; Trnski, D.; Musani, V.; Ozretic, P.; Levanat, S. Role of GLI Transcription Factors in Pathogenesis and Their Potential as New Therapeutic Targets. Int. J. Mol. Sci. 2018, 19, 2562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, C.H.; Dignam, J.J.; Hammond, M.E.; Klimowicz, A.C.; Petrillo, S.K.; Magliocco, A.; Jordan, R.; Trotti, A.; Spencer, S.; Cooper, J.S.; et al. Glioma-Associated Oncogene Family Zinc Finger 1 Expression and Metastasis in Patients With Head and Neck Squamous Cell Carcinoma Treated With Radiation Therapy (RTOG 9003). J. Clin. Oncol. 2011, 29, 1326–1334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fan, H.-X.; Wang, S.; Zhao, H.; Liu, N.; Chen, D.; Sun, M.; Zheng, J. Sonic hedgehog signaling may promote invasion and metastasis of oral squamous cell carcinoma by activating MMP-9 and E-cadherin expression. Med. Oncol. 2014, 31, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.-F.; Chang, C.-J.; Lin, C.-P.; Chang, S.-Y.; Chu, P.-Y.; Tai, S.-K.; Li, W.-Y.; Chao, K.C.; Chen, Y.-J. Expression of hedgehog signaling molecules as a prognostic indicator of oral squamous cell carcinoma. Head Neck 2012, 34, 1556–1561. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Cui, Y.; Ni, W.; Kim, S.-H.; Xuan, Y. Gli1, a potential regulator of esophageal cancer stem cell, is identified as an independent adverse prognostic factor in esophageal squamous cell carcinoma. J. Cancer Res. Clin. Oncol. 2016, 143, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Wadhwa, R.; Wang, X.; Baladandayuthapani, V.; Liu, B.; Shiozaki, H.; Shimodaira, Y.; Lin, Q.; Elimova, E.; Hofstetter, W.L.; Swisher, S.G.; et al. Nuclear expression of Gli-1 is predictive of pathologic complete response to chemoradiation in trimodality treated oesophageal cancer patients. Br. J. Cancer 2017, 117, 648–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Yan, M.; Li, R.R.; Chen, W.T. Sonic Hedgehog Signalling Activation Contributes to ALCAM Over-Expression and Poor Clinical Outcome in Patients with Oral Squamous Cell Carcinoma. Chin. J. Dent. Res. 2018, 21, 31–40. [Google Scholar]
- Dimitrova, K.; Stoehr, M.; Dehghani, F.; Dietz, A.; Wichmann, G.; Bertolini, J.; Mozet, C. Overexpression of the Hedgehog Signalling Pathway in Head and Neck Squamous Cell Carcinoma. Onkologie 2013, 36, 1. [Google Scholar] [CrossRef]
- Richtig, G.; Aigelsreiter, A.M.; Asslaber, M.; Weiland, T.; Pichler, M.; Eberhard, K.; Sygulla, S.; Schauer, S.; Hoefler, G.; Aigelsreiter, A. Hedgehog pathway proteins SMO and GLI expression as prognostic markers in head and neck squamous cell carcinoma. Histopathology 2019, 75, 118–127. [Google Scholar] [CrossRef] [Green Version]
- Schneider, S.; Thurnher, D.; Kloimstein, P.; Leitner, V.; Petzelbauer, P.; Pammer, J.; Brunner, M.; Erovic, B.M. Expression of the Sonic hedgehog pathway in squamous cell carcinoma of the skin and the mucosa of the head and neck. Head Neck 2011, 33, 244–250. [Google Scholar] [CrossRef]
- Gonzalez, A.C.; Ferreira, M.; Ariel, T.; Reis, S.R.A.; Andrade, Z.; Medrado, A.P. Immunohistochemical evaluation of hedgehog signalling in epithelial/mesenchymal interactions in squamous cell carcinoma transformation: A pilot study. J. Oral Pathol. Med. 2015, 45, 173–179. [Google Scholar] [CrossRef]
- Yu, J.; Wu, R.; Wang, Z.; Chen, S.; Chen, S.; Guo, G.; Liu, Z. Cyclopamine Suppresses Human Esophageal Carcinoma Cell Growth by Inhibiting Glioma-Associated Oncogene Protein-1, a Marker of Human Esophageal Carcinoma Progression. Med Sci. Monit. 2019, 25, 1518–1525. [Google Scholar] [CrossRef]
- Mozet, C.; Stoehr, M.; Dimitrova, K.; Dietz, A.; Wichmann, G. Hedgehog targeting by cyclopamine suppresses head and neck squamous cell carcinoma and enhances chemotherapeutic effects. Anticancer. Res. 2013, 33, 2415–2424. [Google Scholar] [PubMed]
- Hehlgans, S.; Booms, P.; Güllülü, Ö.; Sader, R.; Rödel, C.; Balermpas, P.; Rödel, F.; Ghanaati, S. Radiation Sensitization of Basal Cell and Head and Neck Squamous Cell Carcinoma by the Hedgehog Pathway Inhibitor Vismodegib. Int. J. Mol. Sci. 2018, 19, 2485. [Google Scholar] [CrossRef] [Green Version]
- Essid, N.; Chambard, J.C.; Elgaaïed, A.B. Induction of epithelial-mesenchymal transition (EMT) and Gli1 expression in head and neck squamous cell carcinoma (HNSCC) spheroid cultures. Bosn. J. Basic Med Sci. 2018, 18, 336–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, M.F.S.D.; Miguita, L.; De Andrade, N.P.; Heguedusch, D.; Rodini, C.O.; Moyses, R.A.; Toporcov, T.N.; Gama, R.R.; Tajara, E.E.; Nunes, F.D. GLI3 knockdown decreases stemness, cell proliferation and invasion in oral squamous cell carcinoma. Int. J. Oncol. 2018, 53, 2458–2472. [Google Scholar] [CrossRef] [PubMed]
- Luo, J. Glycogen synthase kinase 3β (GSK3β) in tumorigenesis and cancer chemotherapy. Cancer Lett. 2009, 273, 194–200. [Google Scholar] [CrossRef] [Green Version]
- Trnski, D.; Sabol, M.; Gojevic, A.; Martinić, M.; Ozretic, P.; Musani, V.; Ramić, S.; Levanat, S. GSK3β and Gli3 play a role in activation of Hedgehog-Gli pathway in human colon cancer—Targeting GSK3β downregulates the signaling pathway and reduces cell proliferation. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2015, 1852, 2574–2584. [Google Scholar] [CrossRef] [Green Version]
- Kinzler, K.W.; Ruppert, J.M.; Bigner, S.H.; Vogelstein, B. The GLI gene is a member of the Kruppel family of zinc finger proteins. Nature 1988, 332, 371–374. [Google Scholar] [CrossRef]
- Kinzler, K.W.; Vogelstein, B. The GLI gene encodes a nuclear protein which binds specific sequences in the human genome. Mol. Cell. Boil. 1990, 10, 634–642. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Harrington, N.; Moraes, R.C.; Wu, M.-F.; Hilsenbeck, S.G.; Lewis, M.T. Cyclopamine inhibition of human breast cancer cell growth independent of Smoothened (Smo). Breast Cancer Res. Treat. 2008, 115, 505–521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nayak, A.; Siddharth, S.; Das, S.; Nayak, D.; Sethy, C.; Kundu, C.N. Nanoquinacrine caused apoptosis in oral cancer stem cells by disrupting the interaction between GLI1 and β catenin through activation of GSK3β. Toxicol. Appl. Pharmacol. 2017, 330, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Schindelin, J.; Arganda-Carreras, I.; Frise, E.; Kaynig, V.; Longair, M.; Pietzch, T.; Preibisch, S.; Rueden, C.; Saalfeld, S.; Schmid, B.; et al. Fiji: An open source platform for biological-image analysis. Nature Meth. 2012, 9, 679–682. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Line | Tissue | Cyclopamine (µM) | LiCl (mM) | GANT61 (μM) |
|---|---|---|---|---|
| SCC9 | Tongue | >10 | 11.45 | >25 |
| SCC25 | Tongue | 9.51 | 15.24 | 6.83 |
| A253 | Salivary gland | 6.48 | 14.56 | 8.36 |
| DETROIT562 | Pharynx | >10 | 11.4 | 9.54 |
| FADU | Hypopharynx | >10 | 5.53 | 19.7 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zubčić, V.; Rinčić, N.; Kurtović, M.; Trnski, D.; Musani, V.; Ozretić, P.; Levanat, S.; Leović, D.; Sabol, M. GANT61 and Lithium Chloride Inhibit the Growth of Head and Neck Cancer Cell Lines Through the Regulation of GLI3 Processing by GSK3β. Int. J. Mol. Sci. 2020, 21, 6410. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176410
Zubčić V, Rinčić N, Kurtović M, Trnski D, Musani V, Ozretić P, Levanat S, Leović D, Sabol M. GANT61 and Lithium Chloride Inhibit the Growth of Head and Neck Cancer Cell Lines Through the Regulation of GLI3 Processing by GSK3β. International Journal of Molecular Sciences. 2020; 21(17):6410. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176410
Chicago/Turabian StyleZubčić, Vedran, Nikolina Rinčić, Matea Kurtović, Diana Trnski, Vesna Musani, Petar Ozretić, Sonja Levanat, Dinko Leović, and Maja Sabol. 2020. "GANT61 and Lithium Chloride Inhibit the Growth of Head and Neck Cancer Cell Lines Through the Regulation of GLI3 Processing by GSK3β" International Journal of Molecular Sciences 21, no. 17: 6410. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21176410