Tolerance of Stored Boar Spermatozoa to Autologous Seminal Plasma: A Proteomic and Lipidomic Approach

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
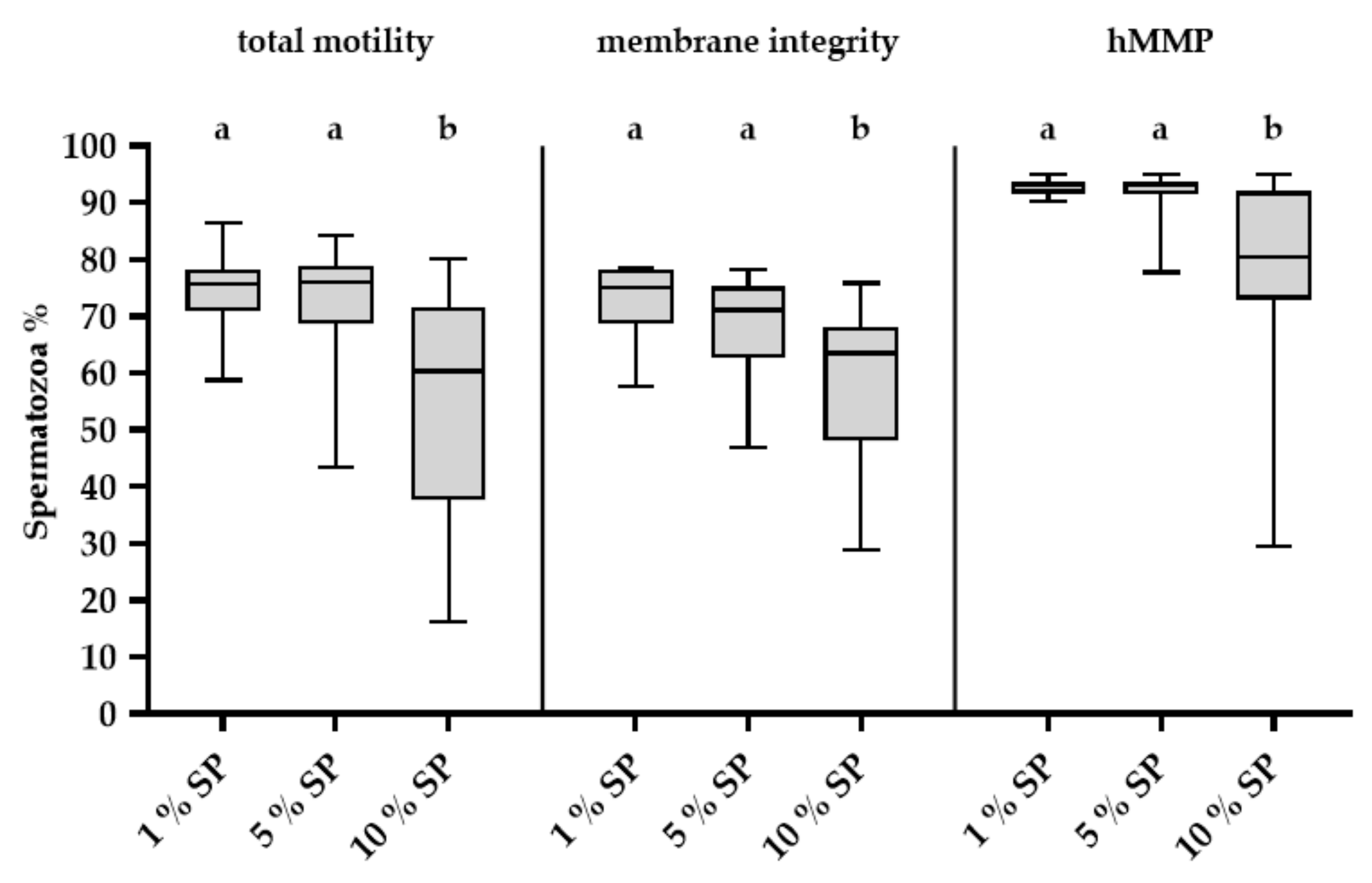
2.1. Effect of Different Concentrations of SP on Stored Spermatozoa
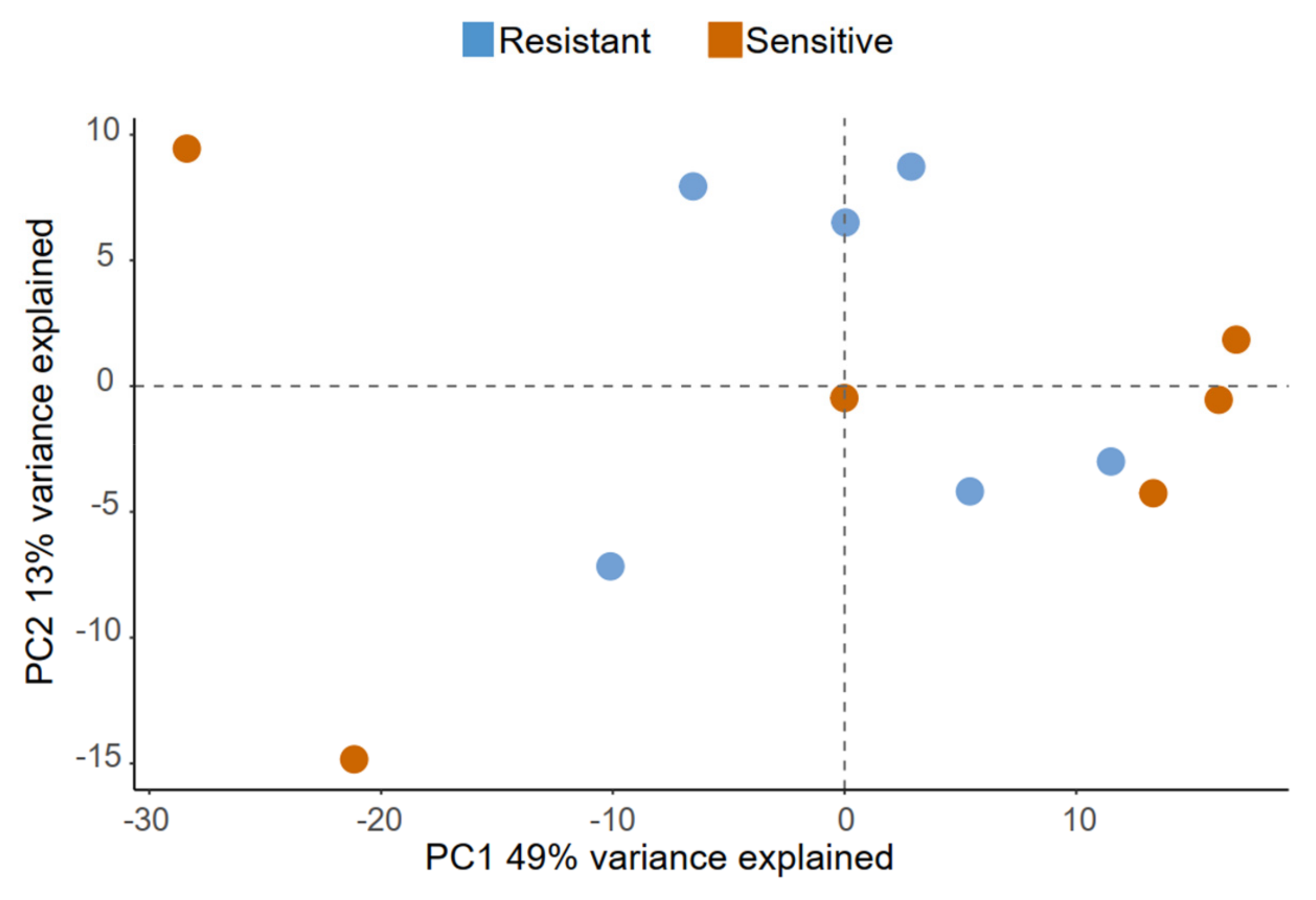
2.2. Boar Specificity and Ejaculate Effects
2.3. Proteomics of SP
2.4. Lipidomics of SP
3. Discussion
4. Materials and Methods
4.1. Spermatological Analysis
4.1.1. Chemicals and Media
4.1.2. Animals, Semen Processing and Seminal Plasma
4.1.3. Assessment of Sperm Motility
4.1.4. Assessment of Sperm Membrane Integrity
4.1.5. Assessment of Sperm Mitochondria Membrane Potential
4.1.6. Statistical Analysis
4.2. Proteome Analysis
4.3. Lipidome Analysis
4.3.1. Lipid Extraction for Mass Spectrometry Lipidomics
4.3.2. MS Data Acquisition
4.3.3. Data Analysis and Post-Processing
4.3.4. Lipidomics Data Downstream Analysis
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Waberski, D.; Riesenbeck, A.; Schulze, M.; Weitze, K.F.; Johnson, L. Application of preserved boar semen for artificial insemination: Past, present and future challenges. Theriogenology 2019, 137, 2–7. [Google Scholar] [CrossRef] [PubMed]
- Hunter, R.H.F. Sperm transport and reservoirs in the pig oviduct in relation to the time of ovulation. Reproduction 1981, 63, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Luther, A.-M.; Waberski, D. In vitro aging of boar spermatozoa: Role of sperm proximity and seminal plasma. Andrology 2019, 7, 382–390. [Google Scholar] [CrossRef]
- Höfner, L.; Luther, A.-M.; Waberski, D. The role of seminal plasma in the liquid storage of spermatozoa. Anim. Reprod. Sci. 2020, 106290. [Google Scholar] [CrossRef] [PubMed]
- Pavaneli, A.P.P.; Passarelli, M.D.S.; De Freitas, F.V.; Ravagnani, G.M.; Torres, M.A.; Martins, S.M.M.K.; Yeste, M.; Andrade, A. Removal of seminal plasma prior to liquid storage of boar spermatozoa: A practice that can improve their fertilizing ability. Theriogenology 2019, 125, 79–86. [Google Scholar] [CrossRef]
- Kommisrud, E.; Paulenz, H.; Sehested, E.; Grevle, I.S. Influence of Boar and Semen Parameters on Motility and Acrosome Integrity in Liquid Boar Semen Stored for Five Days. Acta Veter. Scand. 2002, 43, 49–55. [Google Scholar] [CrossRef] [PubMed]
- Parrilla, I.; Del Olmo, D.; Sijses, L.; Martinez-Alborcia, M.J.; Cuello, C.; Vazquez, J.M.; Martinez, E.A.; Roca, J. Differences in the ability of spermatozoa from individual boar ejaculates to withstand different semen-processing techniques. Anim. Reprod. Sci. 2012, 132, 66–73. [Google Scholar] [CrossRef] [PubMed]
- Caballero, I.; Parrilla, I.; Almiñana, C.; Del Olmo, D.; Roca, J.; Martinez, E.A.; Vazquez, J.M. Seminal Plasma Proteins as Modulators of the Sperm Function and Their Application in Sperm Biotechnologies. Reprod. Domest. Anim. 2012, 47, 12–21. [Google Scholar] [CrossRef]
- Rodriguez-Martínez, H.; Kvist, U.; Ernerudh, J.; Sanz, L.; Calvete, J.J. Seminal Plasma Proteins: What Role Do They Play? Am. J. Reprod. Immunol. 2011, 66, 11–22. [Google Scholar] [CrossRef] [Green Version]
- Juyena, N.S.; Stelletta, C. Seminal Plasma: An Essential Attribute to Spermatozoa. J. Androl. 2011, 33, 536–551. [Google Scholar] [CrossRef] [Green Version]
- Barranco, I.; Rubér, M.; Perez-Patiño, C.; Atikuzzaman, M.; Martinez, E.A.; Roca, J.; Rodriguez-Martínez, H. The Seminal Plasma of the Boar is Rich in Cytokines, with Significant Individual and Intra-Ejaculate Variation. Am. J. Reprod. Immunol. 2015, 74, 523–532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Töpfer-Petersen, E.; Romero, A.; Varela, P.F.; Ekhlasi-Hundrieser, M.; Dostàlovà, Z.; Sanz, L.; Calvete, J.J. Spermadhesins: A new protein family. Facts, hypotheses and perspectives. Andrology 1998, 30, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Caballero, I.; Vazquez, J.M.; Garcia, E.; Parrilla, I.; Roca, J.; Calvete, J.J.; Sanz, L.; Martinez, E. Major proteins of boar seminal plasma as a tool for biotechnological preservation of spermatozoa. Theriogenology 2008, 70, 1352–1355. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, T.; Hiroaki, H.; Furuichi, Y.; Wada, K.; Satoh, M.; Satoh, M.; Osada, T.; Gagnon, C. Cloning of boar SPMI gene which is expressed specifically in seminal vesicle and codes for a sperm motility inhibitor protein. FEBS Lett. 1995, 368, 420–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Centurion, F.; Vazquez, J.M.; Calvete, J.J.; Roca, J.; Sanz, L.; Parrilla, I.; García, E.M.; Martinez, E.A. Influence of Porcine Spermadhesins on the Susceptibility of Boar Spermatozoa to High Dilution. Boil. Reprod. 2003, 69, 640–646. [Google Scholar] [CrossRef]
- Perez-Patiño, C.; Parrilla, I.; Barranco, I.; Vergara-Barberán, M.; Simó-Alfonso, E.F.; Herrero-Martínez, J.M.; Rodriguez-Martínez, H.; Martinez, E.A.; Roca, J. New In-Depth Analytical Approach of the Porcine Seminal Plasma Proteome Reveals Potential Fertility Biomarkers. J. Proteome Res. 2018, 17, 1065–1076. [Google Scholar] [CrossRef] [Green Version]
- Parrilla, I.; Perez-Patiño, C.; Li, J.; Barranco, I.; Padilla, L.; Rodriguez-Martinez, H.; Martinez, E.; Roca, J. Boar semen proteomics and sperm preservation. Theriogenology 2019, 137, 23–29. [Google Scholar] [CrossRef]
- Rodríguez-Martínez, H.; Kvist, U.; Saravia, F.; Wallgren, M.; Johannisson, A.; Sanz, L.; Peña, F.J.; Martínez, E.A.; Roca, J.; Vázquez, J.M.; et al. The physiological roles of the boar ejaculate. Soc. Reprod. Fertil. Suppl. 2009, 66, 1–21. [Google Scholar]
- Perez-Patiño, C.; Barranco, I.; Parrilla, I.; Valero, M.L.; Martinez, E.A.; Rodriguez-Martínez, H.; Roca, J. Characterization of the porcine seminal plasma proteome comparing ejaculate portions. J. Proteom. 2016, 142, 15–23. [Google Scholar] [CrossRef] [Green Version]
- González-Cadavid, V.; Martins, J.A.; Moreno, F.B.; Andrade, T.S.; Santos, A.C.; Monteiro-Moreira, A.C.O.; Moreira, R.A.; Moura, A.A. Seminal plasma proteins of adult boars and correlations with sperm parameters. Theriogenology 2014, 82, 697–707. [Google Scholar] [CrossRef] [Green Version]
- Vilagran, I.; Castillo-Martín, M.; Prieto-Martínez, N.; Bonet, S.; Yeste, M.; Information, P.E.K.F.C. Triosephosphate isomerase (TPI) and epididymal secretory glutathione peroxidase (GPX5) are markers for boar sperm quality. Anim. Reprod. Sci. 2016, 165, 22–30. [Google Scholar] [CrossRef] [PubMed]
- De Lazari, F.L.; Sontag, E.R.; Schneider, A.; Moura, A.A.; Vasconcelos, F.R.; Nagano, C.S.; Mattos, R.C.; Jobim, M.I.M.; Bustamante-Filho, I.C. Seminal plasma proteins and their relationship with sperm motility and morphology in boars. Andrology 2018, 51, e13222. [Google Scholar] [CrossRef] [PubMed]
- Vilagran, I.; Yeste, M.; Sancho, S.; Castillo, J.; Oliva, R.; Bonet, S. Comparative analysis of boar seminal plasma proteome from different freezability ejaculates and identification of Fibronectin 1 as sperm freezability marker. Andrology 2015, 3, 345–356. [Google Scholar] [CrossRef] [PubMed]
- Novak, S.; Dixon, W.T.; Foxcroft, G.R.; Dyck, M.; Ruiz-Sánchez, A. Seminal Plasma Proteins as Potential Markers of Relative Fertility in Boars. J. Androl. 2009, 31, 188–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, W.-S.; Oh, S.-A.; Kim, Y.-J.; Rahman, S.; Park, Y.-J.; Pang, M.-G. Proteomic approaches for profiling negative fertility markers in inferior boar spermatozoa. Sci. Rep. 2015, 5, 13821. [Google Scholar] [CrossRef] [Green Version]
- Barranco, I.; Padilla, L.; Pérez-Patiño, C.; Vazquez, J.M.; Martínez, E.A.; Rodríguez-Martínez, H.; Roca, J.; Parrilla, I. Seminal Plasma Cytokines Are Predictive of the Outcome of Boar Sperm Preservation. Front. Veter. Sci. 2019, 6, 436. [Google Scholar] [CrossRef]
- Mann, T.; Lutwak-Mann, C.; Frs, T.M. Male Reproductive Function and Semen; Springer Science and Business Media LLC: Berlin, Germany, 1981; pp. 306–312. [Google Scholar]
- Wood, P.L.; Scoggin, K.; Ball, B.A.; Troedsson, M.H.; Squires, E.L. Lipidomics of equine sperm and seminal plasma: Identification of amphiphilic (O-acyl)-ω-hydroxy-fatty acids. Theriogenology 2016, 86, 1212–1221. [Google Scholar] [CrossRef]
- Engel, K.M.; Baumann, S.; Rolle-Kampczyk, U.; Schiller, J.; Von Bergen, M.; Grunewald, S. Metabolomic profiling reveals correlations between spermiogram parameters and the metabolites present in human spermatozoa and seminal plasma. PLoS ONE 2019, 14, e0211679. [Google Scholar] [CrossRef] [Green Version]
- Caballero, I.; Vazquez, J.M.; Centurión, F.; Rodriguez-Martínez, H.; Parrilla, I.; Roca, J.; Cuello, C.; Martinez, E.A. Comparative Effects of Autologous and Homologous Seminal Plasma on the Viability of Largely Extended Boar Spermatozoa. Reprod. Domest. Anim. 2004, 39, 370–375. [Google Scholar] [CrossRef]
- Druart, X.; Rickard, J.P.; Tsikis, G.; De Graaf, S.P. Seminal plasma proteins as markers of sperm fertility. Theriogenology 2019, 137, 30–35. [Google Scholar] [CrossRef]
- Rodriguez, A.L.; Van Soom, A.; Arsenakis, I.; Maes, D. Boar management and semen handling factors affect the quality of boar extended semen. Porc. Health Manag. 2017, 3, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Yu, M.; Liu, B.; Zhu, M.; Li, K. Porcine granulin gene (GRN): Molecular cloning, polymorphism and chromosomal localization. DNA Seq. 2006, 17, 350–354. [Google Scholar] [CrossRef] [PubMed]
- Baba, T.; Hoff, H.B.; Nemoto, H.; Lee, H.; Orth, J.; Arai, Y.; Gerton, G.L. Acrogranin, an acrosomal cysteine-rich glycoprotein, is the precursor of the growth-modulating peptides, granulins, and epithelins, and is expressed in somatic as well as male germ cells. Mol. Reprod. Dev. 1993, 34, 233–243. [Google Scholar] [CrossRef] [PubMed]
- Bateman, A.; Bennett, H.P. Granulins: The structure and function of an emerging family of growth factors. J. Endocrinol. 1998, 158, 145–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, S.; Kratzsch, D.; Schaab, M.; Scholz, M.; Grunewald, S.; Thiery, J.; Paasch, U.; Kratzsch, J. Seminal plasma adipokine levels are correlated with functional characteristics of spermatozoa. Fertil. Steril. 2013, 99, 1256–1263. [Google Scholar] [CrossRef] [PubMed]
- Viana, A.G.A.; Martins, A.M.A.; Pontes, A.H.; Fontes, W.; Castro, M.S.; Ricart, C.A.O.; Sousa, M.V.; Kaya, A.; Topper, E.; Memili, E.; et al. Proteomic landscape of seminal plasma associated with dairy bull fertility. Sci. Rep. 2018, 8, 16323. [Google Scholar] [CrossRef]
- Dall, E.; Brandstetter, H. Structure and function of legumain in health and disease. Biochimie 2016, 122, 126–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacyno, E.; Kołodziej, A.; Kawęcka, M.; Pietruszka, A.; Matysiak, B.; Kamyczek, M. The relationship between blood serum and seminal plasma cholesterol content in young boars and their semen qualitative traits and testes size. Arch. Anim. Breed. 2009, 52, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Am-In, N.; Kirkwood, R.; Techakumphu, M.; Tantasuparuk, W. Lipid profiles of sperm and seminal plasma from boars having normal or low sperm motility. Theriogenology 2011, 75, 897–903. [Google Scholar] [CrossRef]
- Žaja, I.; Žura, S.M.; Vince, S.; Vilić, M.; Majić-Balić, I.; Đuričić, D.; Milinković-Tur, S. Differences in seminal plasma and spermatozoa antioxidative systems and seminal plasma lipid and protein levels among boar breeds and hybrid genetic traits. Anim. Reprod. Sci. 2016, 170, 75–82. [Google Scholar] [CrossRef]
- Vadnais, M.L.; Althouse, G.C. Characterization of capacitation, cryoinjury, and the role of seminal plasma in porcine sperm. Theriogenology 2011, 76, 1508–1516. [Google Scholar] [CrossRef] [PubMed]
- Johnson, L.A.; Gerrits, R.J.; Young, E.P. Quantitiative analysis of porcine spermatozoa and seminal plasma phospholipids as affected by frequency of ejaculation. Reprod. Fertil. 1969, 19, 95–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schulze, M.; Jakop, U.; Jung, M.; Cabezón, F. Influences on thermo-resistance of boar spermatozoa. Theriogenology 2019, 127, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Sampaio, J.L.; Gerl, M.J.; Klose, C.; Ejsing, C.S.; Beug, H.; Simons, K.; Shevchenko, A. Membrane lipidome of an epithelial cell line. Proc. Natl. Acad. Sci. USA 2011, 108, 1903–1907. [Google Scholar] [CrossRef] [Green Version]
- Ejsing, C.S.; Sampaio, J.L.; Surendranath, V.; Duchoslav, E.; Ekroos, K.; Klemm, R.W.; Simons, K.; Shevchenko, A. Global analysis of the yeast lipidome by quantitative shotgun mass spectrometry. Proc. Natl. Acad. Sci. USA 2009, 106, 2136–2141. [Google Scholar] [CrossRef] [Green Version]
- Surma, M.A.; Herzog, R.; Vasilj, A.; Klose, C.; Christinat, N.; Morin-Rivron, D.; Simons, K.; Masoodi, M.; Sampaio, J.L. An automated shotgun lipidomics platform for high throughput, comprehensive, and quantitative analysis of blood plasma intact lipids. Eur. J. Lipid Sci. Technol. 2015, 117, 1540–1549. [Google Scholar] [CrossRef] [Green Version]
- Liebisch, G.; Binder, M.; Schifferer, R.; Langmann, T.; Schulz, B.; Schmitz, G. High throughput quantification of cholesterol and cholesteryl ester by electrospray ionization tandem mass spectrometry (ESI-MS/MS). Biochim. et Biophys. Acta (BBA). Mol. Cell Boil. Lipids 2006, 1761, 121–128. [Google Scholar] [CrossRef]
- Herzog, R.; Schwudke, D.; Schuhmann, K.; Sampaio, J.L.; Bornstein, S.R.; Schroeder, M.; Shevchenko, A. A novel informatics concept for high-throughput shotgun lipidomics based on the molecular fragmentation query language. Genome Boil. 2011, 12. [Google Scholar] [CrossRef] [Green Version]
- Herzog, R.; Schuhmann, K.; Schwudke, D.; Sampaio, J.L.; Bornstein, S.R.; Schroeder, M.; Shevchenko, A. LipidXplorer: A Software for Consensual Cross-Platform Lipidomics. PLoS ONE 2012, 7, e29851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anonymous. The R Project for Statistical Computing. Available online: http://www.r-project.org/ (accessed on 13 February 2012).








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Höfner, L.; Luther, A.-M.; Palladini, A.; Fröhlich, T.; Waberski, D. Tolerance of Stored Boar Spermatozoa to Autologous Seminal Plasma: A Proteomic and Lipidomic Approach. Int. J. Mol. Sci. 2020, 21, 6474. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186474
Höfner L, Luther A-M, Palladini A, Fröhlich T, Waberski D. Tolerance of Stored Boar Spermatozoa to Autologous Seminal Plasma: A Proteomic and Lipidomic Approach. International Journal of Molecular Sciences. 2020; 21(18):6474. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186474
Chicago/Turabian StyleHöfner, Lisa, Anne-Marie Luther, Alessandra Palladini, Thomas Fröhlich, and Dagmar Waberski. 2020. "Tolerance of Stored Boar Spermatozoa to Autologous Seminal Plasma: A Proteomic and Lipidomic Approach" International Journal of Molecular Sciences 21, no. 18: 6474. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186474