Alternative Options for Skin Cancer Therapy via Regulation of AKT and Related Signaling Pathways


Abstract
:1. Introduction
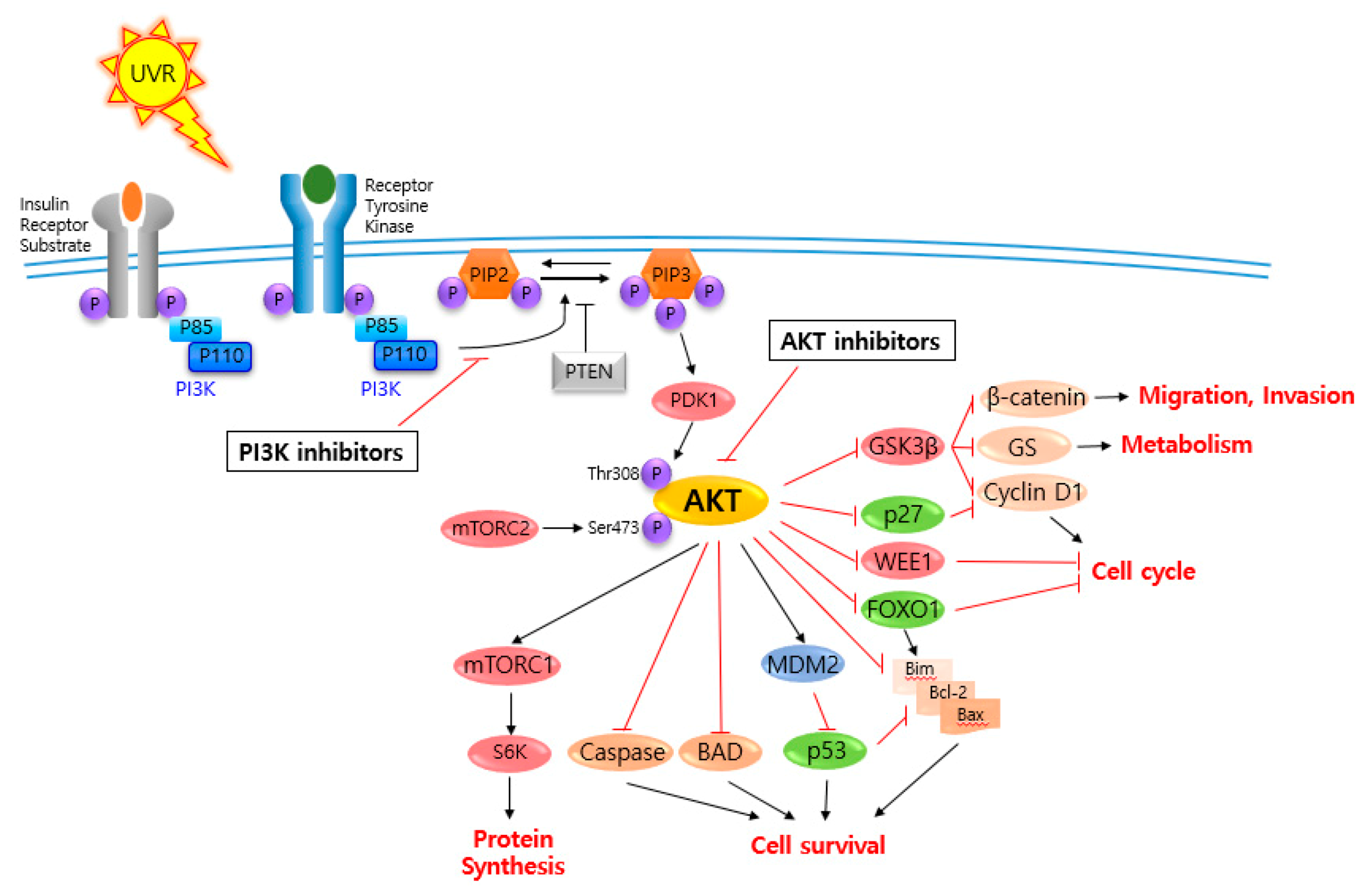
2. AKT and Related Signaling Pathways Are Important in Skin Cancer Regulation
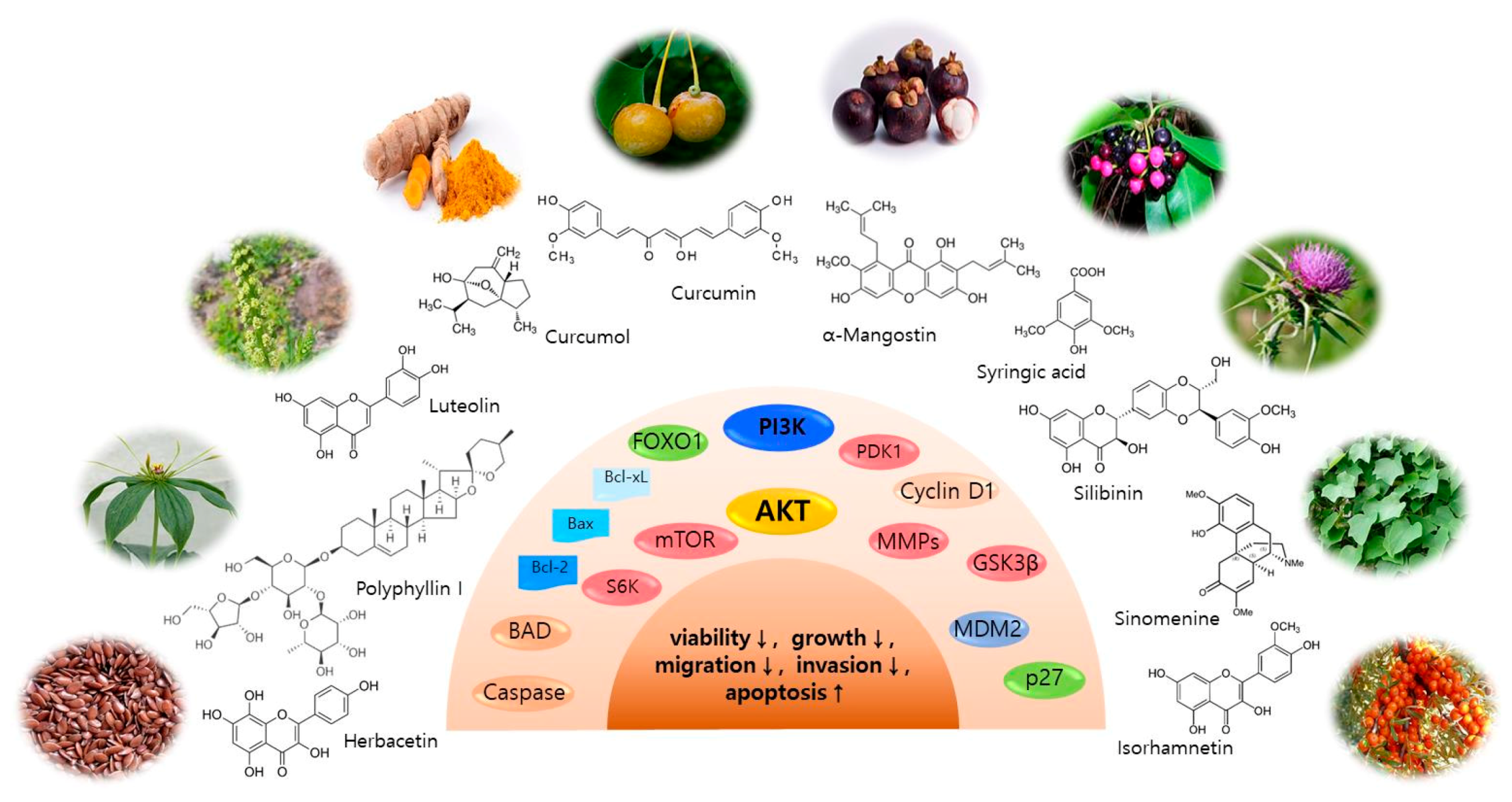
3. AKT and Related Signaling Pathway Inhibitors for Skin Cancer Regulation
3.1. Isorhamnetin
3.2. Curcumol
3.3. Polyphyllin I
3.4. Herbacetin
3.5. Luteolin
3.6. Sinomenine
3.7. Syringic Acid
3.8. Ginkgo Biloba Exocarp Extract
3.9. α-Mangostin
3.10. Silibinin (Silybin)
3.11. Curcumin
3.12. Other Bioactive Components
4. Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Search Strategy and Selection Criteria
References
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2020. CA Cancer J. Clin. 2020, 70, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Global Burden of Disease Cancer Collaboration; Fitzmaurice, C.; Abate, D.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdel-Rahman, O.; Abdelalim, A.; Abdoli, A.; Abdollahpour, I.; et al. Global, Regional, and National Cancer Incidence, Mortality, Years of Life Lost, Years Lived With Disability, and Disability-Adjusted Life-Years for 29 Cancer Groups, 1990 to 2017: A Systematic Analysis for the Global Burden of Disease Study. JAMA Oncol. 2019. [Google Scholar] [CrossRef] [Green Version]
- Tran, D.A.; Coronado, A.C.; Sarker, S.; Alvi, R. Estimating the health care costs of non-melanoma skin cancer in Saskatchewan using physician billing data. Curr. Oncol. 2019, 26, 114–118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guy, G.P., Jr.; Machlin, S.R.; Ekwueme, D.U.; Yabroff, K.R. Prevalence and costs of skin cancer treatment in the U.S., 2002–2006 and 2007–2011. Am. J. Prev. Med. 2015, 48, 183–187. [Google Scholar] [CrossRef]
- Apalla, Z.; Nashan, D.; Weller, R.B.; Castellsague, X. Skin Cancer: Epidemiology, Disease Burden, Pathophysiology, Diagnosis, and Therapeutic Approaches. Dermatol. Ther. 2017, 7, 5–19. [Google Scholar] [CrossRef] [Green Version]
- Wrobel, S.; Przybylo, M.; Stepien, E. The Clinical Trial Landscape for Melanoma Therapies. J. Clin. Med. 2019, 8, 368. [Google Scholar] [CrossRef] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Khavari, P.A. Modelling cancer in human skin tissue. Nat. Rev. Cancer 2006, 6, 270–280. [Google Scholar] [CrossRef]
- Roh, E.; Lee, M.H.; Zykova, T.A.; Zhu, F.; Nadas, J.; Kim, H.G.; Bae, K.B.; Li, Y.; Cho, Y.Y.; Curiel-Lewandrowski, C.; et al. Targeting PRPK and TOPK for skin cancer prevention and therapy. Oncogene 2018, 37, 5633–5647. [Google Scholar] [CrossRef]
- Lee, M.H.; Lim, D.Y.; Kim, M.O.; Lee, S.Y.; Shin, S.H.; Kim, J.Y.; Kim, S.H.; Kim, D.J.; Jung, S.K.; Yao, K.; et al. Genetic ablation of caspase-7 promotes solar-simulated light-induced mouse skin carcinogenesis: The involvement of keratin-17. Carcinogenesis 2015, 36, 1372–1380. [Google Scholar] [CrossRef]
- Kim, J.E.; Lee, K.W. Molecular Targets of Phytochemicals for Skin Inflammation. Curr. Pharm. Des. 2018, 24, 1533–1550. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Choi, B.Y.; Lee, M.H.; Bode, A.M.; Dong, Z. Implications of Genetic and Epigenetic Alterations of CDKN2A (p16(INK4a)) in Cancer. EBioMedicine 2016, 8, 30–39. [Google Scholar] [CrossRef] [Green Version]
- Pal, H.C.; Hunt, K.M.; Diamond, A.; Elmets, C.A.; Afaq, F. Phytochemicals for the Management of Melanoma. Mini Rev. Med. Chem. 2016, 16, 953–979. [Google Scholar] [PubMed]
- Lee, M.H.; Huang, Z.; Kim, D.J.; Kim, S.H.; Kim, M.O.; Lee, S.Y.; Xie, H.; Park, S.J.; Kim, J.Y.; Kundu, J.K.; et al. Direct targeting of MEK1/2 and RSK2 by silybin induces cell-cycle arrest and inhibits melanoma cell growth. Cancer Prev. Res. 2013, 6, 455–465. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nikas, I.P.; Paschou, S.A.; Ryu, H.S. The Role of Nicotinamide in Cancer Chemoprevention and Therapy. Biomolecules 2020, 10, 477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, M.; Bode, A.M.; Dong, Z.; Lee, M.H. AKT as a Therapeutic Target for Cancer. Cancer Res. 2019, 79, 1019–1031. [Google Scholar] [CrossRef] [Green Version]
- Shariati, M.; Meric-Bernstam, F. Targeting AKT for cancer therapy. Expert Opin. Investig. Drugs 2019, 28, 977–988. [Google Scholar] [CrossRef]
- Kumar, C.C.; Madison, V. AKT crystal structure and AKT-specific inhibitors. Oncogene 2005, 24, 7493–7501. [Google Scholar] [CrossRef] [Green Version]
- Fayard, E.; Tintignac, L.A.; Baudry, A.; Hemmings, B.A. Protein kinase B/Akt at a glance. J. Cell Sci. 2005, 118, 5675–5678. [Google Scholar] [CrossRef] [Green Version]
- Song, M.; Liu, X.; Liu, K.; Zhao, R.; Huang, H.; Shi, Y.; Zhang, M.; Zhou, S.; Xie, H.; Chen, H.; et al. Targeting AKT with Oridonin Inhibits Growth of Esophageal Squamous Cell Carcinoma In Vitro and Patient-Derived Xenografts In Vivo. Mol. Cancer Ther. 2018, 17, 1540–1553. [Google Scholar] [CrossRef] [Green Version]
- Soares, C.D.; Borges, C.F.; Sena-Filho, M.; Almeida, O.P.; Stelini, R.F.; Cintra, M.L.; Graner, E.; Zecchin, K.G.; Jorge, J. Prognostic significance of cyclooxygenase 2 and phosphorylated Akt1 overexpression in primary nonmetastatic and metastatic cutaneous melanomas. Melanoma Res. 2017, 27, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Slipicevic, A.; Holm, R.; Nguyen, M.T.; Bohler, P.J.; Davidson, B.; Florenes, V.A. Expression of activated Akt and PTEN in malignant melanomas: Relationship with clinical outcome. Am. J. Clin. Pathol. 2005, 124, 528–536. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H.; Robinson, J.P.; Arave, R.A.; Burnett, W.J.; Kircher, D.A.; Chen, G.; Davies, M.A.; Grossmann, A.H.; VanBrocklin, M.W.; McMahon, M.; et al. AKT1 Activation Promotes Development of Melanoma Metastases. Cell Rep. 2015, 13, 898–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Yang, Z.; Song, W.; Zhou, L.; Li, Q.; Tao, K.; Zhou, J.; Wang, X.; Zheng, Z.; You, N.; et al. Overexpression of Bmi-1 contributes to the invasion and metastasis of hepatocellular carcinoma by increasing the expression of matrix metalloproteinase (MMP)2, MMP-9 and vascular endothelial growth factor via the PTEN/PI3K/Akt pathway. Int. J. Oncol. 2013, 43, 793–802. [Google Scholar] [CrossRef] [Green Version]
- Egeblad, M.; Werb, Z. New functions for the matrix metalloproteinases in cancer progression. Nat. Rev. Cancer 2002, 2, 161–174. [Google Scholar] [CrossRef]
- Song, N.R.; Lee, E.; Byun, S.; Kim, J.E.; Mottamal, M.; Park, J.H.; Lim, S.S.; Bode, A.M.; Lee, H.J.; Lee, K.W.; et al. Isoangustone A, a novel licorice compound, inhibits cell proliferation by targeting PI3K, MKK4, and MKK7 in human melanoma. Cancer Prev. Res. 2013, 6, 1293–1303. [Google Scholar] [CrossRef] [Green Version]
- Jung, S.K.; Kim, J.E.; Lee, S.Y.; Lee, M.H.; Byun, S.; Kim, Y.A.; Lim, T.G.; Reddy, K.; Huang, Z.; Bode, A.M.; et al. The P110 subunit of PI3-K is a therapeutic target of acacetin in skin cancer. Carcinogenesis 2014, 35, 123–130. [Google Scholar] [CrossRef] [Green Version]
- Antony, J.; Saikia, M.; Vinod, V.; Nath, L.R.; Katiki, M.R.; Murty, M.S.; Paul, A.; Shabna, A.; Chandran, H.; Joseph, S.M.; et al. DW-F5: A novel formulation against malignant melanoma from Wrightia tinctoria. Sci. Rep. 2015, 5, 11107. [Google Scholar] [CrossRef] [Green Version]
- Arcidiacono, P.; Stabile, A.M.; Ragonese, F.; Pistilli, A.; Calvieri, S.; Bottoni, U.; Crisanti, A.; Spaccapelo, R.; Rende, M. Anticarcinogenic activities of sulforaphane are influenced by Nerve Growth Factor in human melanoma A375 cells. Food Chem. Toxicol. 2018, 113, 154–161. [Google Scholar] [CrossRef]
- Olas, B.; Skalski, B.; Ulanowska, K. The Anticancer Activity of Sea Buckthorn [Elaeagnus rhamnoides (L.) A. Nelson]. Front. Pharmacol. 2018, 9, 232. [Google Scholar] [CrossRef] [Green Version]
- Duan, R.; Liang, X.; Chai, B.; Zhou, Y.; Du, H.; Suo, Y.; Chen, Z.; Li, Q.; Huang, X. Isorhamnetin Induces Melanoma Cell Apoptosis via the PI3K/Akt and NF-kappaB Pathways. Biomed. Res. Int. 2020, 2020, 1057943. [Google Scholar] [CrossRef] [PubMed]
- Guo, P.; Wang, Y.W.; Weng, B.X.; Li, X.K.; Yang, S.L.; Ye, F.Q. Synthesis, anti-tumor activity, and structure-activity relationships of curcumol derivatives. J. Asian Nat. Prod. Res. 2014, 16, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Ning, N.; Liu, S.; Liu, X.; Tian, Z.; Jiang, Y.; Yu, N.; Tan, B.; Feng, H.; Feng, X.; Zou, L. Curcumol inhibits the proliferation and metastasis of melanoma via the miR-152-3p/PI3K/AKT and ERK/NF-kappaB signaling pathways. J. Cancer 2020, 11, 1679–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, R.; Guo, J.; Zhang, Z.; Zhou, Y.; Hua, Y. Polyphyllin I inhibits gastric cancer cell proliferation by downregulating the expression of fibroblast activation protein alpha (FAP) and hepatocyte growth factor (HGF) in cancer-associated fibroblasts. Biochem. Biophys. Res. Commun. 2018, 497, 1129–1134. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Li, Y.; Wang, X.; Hu, S.; Wang, H.; Shi, Q.; Wang, Y.; Yang, Y. Polyphyllin I suppresses human osteosarcoma growth by inactivation of Wnt/beta-catenin pathway in vitro and in vivo. Sci. Rep. 2017, 7, 7605. [Google Scholar] [CrossRef] [Green Version]
- Long, J.; Pi, X. Polyphyllin I Promoted Melanoma Cells Autophagy and Apoptosis via PI3K/Akt/mTOR Signaling Pathway. Biomed. Res. Int. 2020, 2020, 5149417. [Google Scholar] [CrossRef]
- Kim, D.J.; Roh, E.; Lee, M.H.; Oi, N.; Lim, D.Y.; Kim, M.O.; Cho, Y.Y.; Pugliese, A.; Shim, J.H.; Chen, H.; et al. Herbacetin Is a Novel Allosteric Inhibitor of Ornithine Decarboxylase with Antitumor Activity. Cancer Res. 2016, 76, 1146–1157. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.J.; Lee, M.H.; Liu, K.; Lim, D.Y.; Roh, E.; Chen, H.; Kim, S.H.; Shim, J.H.; Kim, M.O.; Li, W.; et al. Herbacetin suppresses cutaneous squamous cell carcinoma and melanoma cell growth by targeting AKT and ODC. Carcinogenesis 2017, 38, 1136–1146. [Google Scholar] [CrossRef] [Green Version]
- Potocnjak, I.; Simic, L.; Gobin, I.; Vukelic, I.; Domitrovic, R. Antitumor activity of luteolin in human colon cancer SW620 cells is mediated by the ERK/FOXO3a signaling pathway. Toxicol. In Vitro 2020, 66, 104852. [Google Scholar] [CrossRef]
- Masraksa, W.; Tanasawet, S.; Hutamekalin, P.; Wongtawatchai, T.; Sukketsiri, W. Luteolin attenuates migration and invasion of lung cancer cells via suppressing focal adhesion kinase and non-receptor tyrosine kinase signaling pathway. Nutr. Res. Pract. 2020, 14, 127–133. [Google Scholar] [CrossRef]
- Byun, S.; Lee, K.W.; Jung, S.K.; Lee, E.J.; Hwang, M.K.; Lim, S.H.; Bode, A.M.; Lee, H.J.; Dong, Z. Luteolin inhibits protein kinase C(epsilon) and c-Src activities and UVB-induced skin cancer. Cancer Res. 2010, 70, 2415–2423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yao, X.; Jiang, W.; Yu, D.; Yan, Z. Luteolin inhibits proliferation and induces apoptosis of human melanoma cells in vivo and in vitro by suppressing MMP-2 and MMP-9 through the PI3K/AKT pathway. Food Funct. 2019, 10, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Xia, X.; May, B.H.; Zhang, A.L.; Guo, X.; Lu, C.; Xue, C.C.; Huang, Q. Chinese Herbal Medicines for Rheumatoid Arthritis: Text-Mining the Classical Literature for Potentially Effective Natural Products. Evid. Based Complement Alternat Med. 2020, 2020, 7531967. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Liu, C.; Tan, T.; Li, S.; Tang, S.; Chen, X. Sinomenine sensitizes human gastric cancer cells to cisplatin through negative regulation of PI3K/AKT/Wnt signaling pathway. Anticancer Drugs 2019, 30, 983–990. [Google Scholar] [CrossRef]
- Sun, Z.; Zheng, L.; Liu, X.; Xing, W.; Liu, X. Sinomenine inhibits the growth of melanoma by enhancement of autophagy via PI3K/AKT/mTOR inhibition. Drug Des. Devel. Ther. 2018, 12, 2413–2421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Periyannan, V.; Annamalai, V.; Veerasamy, V. Syringic acid modulates molecular marker-involved cell proliferation, survival, apoptosis, inflammation, and angiogenesis in DMBA-induced oral squamous cell carcinoma in Syrian hamsters. J. Biochem. Mol. Toxicol. 2020, e22574. [Google Scholar] [CrossRef]
- Cho, H.D.; Kim, J.H.; Won, Y.S.; Moon, K.D.; Seo, K.I. Inhibitory Effects of Pectinase-Treated Prunus Mume Fruit Concentrate on Colorectal Cancer Proliferation and Angiogenesis of Endothelial Cells. J. Food Sci. 2019, 84, 3284–3295. [Google Scholar] [CrossRef]
- Ha, S.J.; Lee, J.; Park, J.; Kim, Y.H.; Lee, N.H.; Kim, Y.E.; Song, K.M.; Chang, P.S.; Jeong, C.H.; Jung, S.K. Syringic acid prevents skin carcinogenesis via regulation of NoX and EGFR signaling. Biochem. Pharmacol. 2018, 154, 435–445. [Google Scholar] [CrossRef]
- Cao, C.; Han, D.; Su, Y.; Ge, Y.; Chen, H.; Xu, A. Ginkgo biloba exocarp extracts induces autophagy in Lewis lung cancer cells involving AMPK/mTOR/p70S6k signaling pathway. Biomed. Pharmacother. 2017, 93, 1128–1135. [Google Scholar] [CrossRef]
- Han, D.; Cao, C.; Su, Y.; Wang, J.; Sun, J.; Chen, H.; Xu, A. Ginkgo biloba exocarp extracts inhibits angiogenesis and its effects on Wnt/beta-catenin-VEGF signaling pathway in Lewis lung cancer. J. Ethnopharmacol. 2016, 192, 406–412. [Google Scholar] [CrossRef]
- Cao, C.; Su, Y.; Gao, Y.; Luo, C.; Yin, L.; Zhao, Y.; Chen, H.; Xu, A. Ginkgo biloba Exocarp Extract Inhibits the Metastasis of B16-F10 Melanoma Involving PI3K/Akt/NF-kappaB/MMP-9 Signaling Pathway. Evid. Based Complement Alternat Med. 2018, 2018, 4969028. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Tan, Y.P.; Zhao, L.; Wang, L.; Fu, N.J.; Zheng, S.P.; Shen, X.F. Anticancer activity of dietary xanthone alpha-mangostin against hepatocellular carcinoma by inhibition of STAT3 signaling via stabilization of SHP1. Cell Death Dis. 2020, 11, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien, H.J.; Ying, T.H.; Hsieh, S.C.; Lin, C.L.; Yu, Y.L.; Kao, S.H.; Hsieh, Y.H. alpha-Mangostin attenuates stemness and enhances cisplatin-induced cell death in cervical cancer stem-like cells through induction of mitochondrial-mediated apoptosis. J. Cell. Physiol. 2020, 235, 5590–5601. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Ma, H.; Liu, Z.; Huang, W.; Xu, X.; Zhang, X. alpha-Mangostin inhibits DMBA/TPA-induced skin cancer through inhibiting inflammation and promoting autophagy and apoptosis by regulating PI3K/Akt/mTOR signaling pathway in mice. Biomed. Pharmacother. 2017, 92, 672–680. [Google Scholar] [CrossRef]
- Sellam, L.S.; Zappasodi, R.; Chettibi, F.; Djennaoui, D.; Yahi-Ait Mesbah, N.; Amir-Tidadini, Z.C.; Touil-Boukoffa, C.; Ouahioune, W.; Merghoub, T.; Bourouba, M. Silibinin down-regulates PD-L1 expression in nasopharyngeal carcinoma by interfering with tumor cell glycolytic metabolism. Arch. Biochem. Biophys. 2020, 690, 108479. [Google Scholar] [CrossRef]
- Tilley, C.; Deep, G.; Agarwal, C.; Wempe, M.F.; Biedermann, D.; Valentova, K.; Kren, V.; Agarwal, R. Silibinin and its 2,3-dehydro-derivative inhibit basal cell carcinoma growth via suppression of mitogenic signaling and transcription factors activation. Mol. Carcinog. 2016, 55, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Dheeraj, A.; Rigby, C.M.; O’Bryant, C.L.; Agarwal, C.; Singh, R.P.; Deep, G.; Agarwal, R. Silibinin Treatment Inhibits the Growth of Hedgehog Inhibitor-Resistant Basal Cell Carcinoma Cells via Targeting EGFR-MAPK-Akt and Hedgehog Signaling. Photochem. Photobiol. 2017, 93, 999–1007. [Google Scholar] [CrossRef]
- Pourhanifeh, M.H.; Mottaghi, R.; Razavi, Z.S.; Shafiee, A.; Hajighadimi, S.; Mirzaei, H. Therapeutic Applications of Curcumin and its Novel Formulations in the Treatment of Bladder Cancer: A Review of Current Evidence. Anticancer Agents Med. Chem. 2020. [Google Scholar] [CrossRef]
- Avila-Galvez, M.A.; Gimenez-Bastida, J.A.; Espin, J.C.; Gonzalez-Sarrias, A. Dietary Phenolics against Breast Cancer. A Critical Evidence-Based Review and Future Perspectives. Int. J. Mol. Sci. 2020, 21, 5718. [Google Scholar] [CrossRef]
- Zhao, G.; Han, X.; Zheng, S.; Li, Z.; Sha, Y.; Ni, J.; Sun, Z.; Qiao, S.; Song, Z. Curcumin induces autophagy, inhibits proliferation and invasion by downregulating AKT/mTOR signaling pathway in human melanoma cells. Oncol. Rep. 2016, 35, 1065–1074. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Liao, R.; Silva, M.; Li, S.; Jiang, Y.; Peng, T.; Lazarovici, P.; Zheng, W. Pristimerin-induced uveal melanoma cell death via inhibiting PI3K/Akt/FoxO3a signalling pathway. J. Cell. Mol. Med. 2020, 24, 6208–6219. [Google Scholar] [CrossRef] [PubMed]
- Li, C.Y.; Wang, Q.; Wang, X.M.; Li, G.X.; Shen, S.; Wei, X.L. Gambogic acid exhibits anti-metastatic activity on malignant melanoma mainly through inhibition of PI3K/Akt and ERK signaling pathways. Eur. J. Pharmacol. 2019, 864, 172719. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.N.; Baek, S.B.; Jung, H.J. Bee Venom and Its Peptide Component Melittin Suppress Growth and Migration of Melanoma Cells via Inhibition of PI3K/AKT/mTOR and MAPK Pathways. Molecules 2019, 24, 929. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.; Xiao, P.; Sun, J.; Guo, L. Anticancer effects of kaempferol in A375 human malignant melanoma cells are mediated via induction of apoptosis, cell cycle arrest, inhibition of cell migration and downregulation of m-TOR/PI3K/AKT pathway. J. BUON 2018, 23, 218–223. [Google Scholar]
- Carpi, S.; Polini, B.; Poli, G.; Alcantara Barata, G.; Fogli, S.; Romanini, A.; Tuccinardi, T.; Guella, G.; Frontini, F.P.; Nieri, P.; et al. Anticancer Activity of Euplotin C, Isolated from the Marine Ciliate Euplotes crassus, Against Human Melanoma Cells. Mar. Drugs 2018, 16, 166. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Q.Q.; Liu, W.B. Lycorine inhibits melanoma A375 cell growth and metastasis through the inactivation of the PI3K/AKT signaling pathway. Med. Sci. 2018, 34, 33–38. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.; Wang, Z.; Liu, Y.; Peng, X.; Liu, Y.; Zhu, S.; Zhang, Z.; Qiu, Y.; Jin, M.; Wang, R.; et al. Oxyfadichalcone C inhibits melanoma A375 cell proliferation and metastasis via suppressing PI3K/Akt and MAPK/ERK pathways. Life Sci. 2018, 206, 35–44. [Google Scholar] [CrossRef]
- Chen, X.Y.; Li, D.F.; Han, J.C.; Wang, B.; Dong, Z.P.; Yu, L.N.; Pan, Z.H.; Qu, C.J.; Chen, Y.; Sun, S.G.; et al. Reprogramming induced by isoliquiritigenin diminishes melanoma cachexia through mTORC2-AKT-GSK3beta signaling. Oncotarget 2017, 8, 34565–34575. [Google Scholar] [CrossRef]
- Zou, N.; Wei, Y.; Li, F.; Yang, Y.; Cheng, X.; Wang, C. The inhibitory effects of compound Muniziqi granule against B16 cells and harmine induced autophagy and apoptosis by inhibiting Akt/mTOR pathway. BMC Complement Altern Med. 2017, 17, 517. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Han, X.; Cheng, W.; Ni, J.; Zhang, Y.; Lin, J.; Song, Z. Apigenin inhibits proliferation and invasion, and induces apoptosis and cell cycle arrest in human melanoma cells. Oncol. Rep. 2017, 37, 2277–2285. [Google Scholar] [CrossRef] [Green Version]
- Shih, Y.L.; Chou, H.M.; Chou, H.C.; Lu, H.F.; Chu, Y.L.; Shang, H.S.; Chung, J.G. Casticin impairs cell migration and invasion of mouse melanoma B16F10 cells via PI3K/AKT and NF-kappaB signaling pathways. Environ. Toxicol. 2017, 32, 2097–2112. [Google Scholar] [CrossRef]
- Chen, H.Y.; Jiang, Y.W.; Kuo, C.L.; Way, T.D.; Chou, Y.C.; Chang, Y.S.; Chung, J.G. Chrysin inhibit human melanoma A375.S2 cell migration and invasion via affecting MAPK signaling and NF-kappaB signaling pathway in vitro. Environ. Toxicol. 2019, 34, 434–442. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.F.; Lai, K.C.; Peng, S.F.; Maraming, P.; Huang, Y.P.; Huang, A.C.; Chueh, F.S.; Huang, W.W.; Chung, J.G. Berberine Inhibits Human Melanoma A375.S2 Cell Migration and Invasion via Affecting the FAK, uPA, and NF-kappaB Signaling Pathways and Inhibits PLX4032 Resistant A375.S2 Cell Migration In Vitro. Molecules 2018, 23, 2019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, N.; Hongbo, T.; Xu, Y.; Wu, M.; Wu, Y. Anticancer activity of caffeic acid nbutyl ester against A431 skin carcinoma cell line occurs via induction of apoptosis and inhibition of the mTOR/PI3K/AKT signaling pathway. Mol. Med. Rep. 2018, 17, 5652–5657. [Google Scholar] [PubMed]
- Basu, A.K. DNA Damage, Mutagenesis and Cancer. Int. J. Mol. Sci. 2018, 19, 970. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Lin, L.; Choi, Y.; Michniak-Kohn, B. Development and in-vitro evaluation of co-loaded berberine chloride and evodiamine ethosomes for treatment of melanoma. Int. J. Pharm. 2020, 581, 119278. [Google Scholar] [CrossRef] [PubMed]
- Moolakkadath, T.; Aqil, M.; Ahad, A.; Imam, S.S.; Praveen, A.; Sultana, Y.; Mujeeb, M.; Iqbal, Z. Fisetin loaded binary ethosomes for management of skin cancer by dermal application on UV exposed mice. Int. J. Pharm. 2019, 560, 78–91. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Compounds | Plants | Cancer Types | Cell Lines | Mechanisms | Ref. |
|---|---|---|---|---|---|
| Isorhamnetin | Persicaria thunbergii H., Elaeagnus rhamnoides (L.) | melanoma | B16F10 | proliferation↓, migration↓ pAKT↓, PFKFB4↓ | [31] |
| Curcumol | Curcuma wenyujin | melanoma | B16F10 | viability↓, colony formation↓ migration↓, pAKT↓, c-MET↓, miR-152-3p↓ | [33] |
| Polyphyllin I | Paris polyphylla | melanoma | A375 | cells growth↓, migration↓, invasion↓, cell cycle progression↓, apoptosis↑, Bax↑, cleaved caspases-3↑, Bcl-2↓ autophagy↑, Beclin 1↑, LC3II ↑, p62 ↓ pPI3K↓, pAKT↓, pmTOR↓ | [36] |
| Herbacetin | Flaxseed, ramose scouring rush herb | SCC, melanoma | JB6, A431, SK-MEL-5, SK-MEL-28 | AKT1/2 activity↓, ODC activity↓, growth↓, neoplastic transformation↓, pGSK3β↓, ODC activity↓, AP1 activity↓, NF-κB activity↓, pERK1/2↓, p65↓ | [38] |
| melanoma | A375, Hs294T | tumor growth↓, angiogenesis↓, pEGFR↓, pAKT↓, pERK ↓ pGSK3β↓, ODC activity↓, AP1 activity↓, NF-κB activity↓ | |||
| Luteolin | Reseda luteola | melanoma | A375.S2 | proliferation↓, migration↓, invasion↓, apoptosis↑, MMP-2↓, MMP-9↓, TIMP-1↑, TIMP-2↑, pAKT1↓, pPI3K↓ | [41] |
| Sinomenine | Sinomenium acutum | melanoma | B16F10 | cell viability↓, apoptosis↑, Bax↑, Bcl-2↓, caspase-3 activity↑, autophagy↑, Beclin-1↑, LC3II/LC3I ratio↑, pp62/SQSTM1↓, pAKT↓, pmTOR↓ | [45] |
| Syringic acid | Euterpe oleracea, Rhus javanica | non-melanoma | HaCaT | UVB-induced COX-2↓, UVB-induced MMP-1↓, UVB-induced PGE2 generation↓, UVB-induced AP-1 activity↓, pERK1/2↓, pJNK1/2↓, pp38↓, pMEK1/2↓, p-MKK4/7↓, pMKK3/6↓, pB-Raf↓, pAKT↓, pSrc↓, EGFR↓, UVB-induced cyclooxygenase-2↓, matrix metalloproteinase-1↓, prostaglandin E2↓ | [48] |
| Ginkgo biloba Exocarp Extract | Ginkgo biloba L. | melanoma | B16F10 | proliferation↓, migration↓, heterogeneous adhesion↓, pPI3K↓, pAKT↓, NF-κB↓, MMP-9↓ | [51] |
| Silibinin | Milk thistle plant (Silybum marianum) | BCC | ASZ001, Sant-1, GDC-0449 resistance ASZ001 | growth↓, colony formation ↓, pEGFR↓, pAKT↓, cyclin D1↓, Gli-1↓, SMO↓, SUFU↓, apoptosis↑, caspase-3↑, Bcl-2↓ | [56] |
| Silybum marianum (L.) Gaertn., Asteraceae | BCC | ASZ, BSZ | cell growth↓, clonogenicity↓, apoptosis↑, pEGFR↓, pERK1/2↓, pAKT↓, pSTAT3↓ | [57] | |
| Curcumin | rhizome of Curcuma longa | melanoma | A375 and C8161 | proliferation↓, invasion↓, G2/M phase cell-cycle arrest↑, autophagy↑, pAKT↓, pmTORC1↓, pp70S6K↓ | [60] |
| Pristimerin | Celastraceae, Hippocrateacea | uveal melanoma | UM-1 | apoptosis↑, viability↓, colony formation↓, mitochondrial membrane potential↓, ROS level↑, G0/G1 phase arrest↑ migration↓, invasion↓ pAKT↓, pFoxO3a ↓, Bim↑, p27Kip1↑, cleaved caspase-3↑, PARP↑, Bax↑, Cyclin D1↓, Bcl-2↓ | [61] |
| Gambogic acid | resin of Garciania hanburyi | melanoma | A375, B16F10, | proliferation↓, migration↓, invasion↓, adhesion↓, EMT↓, angiogenesis processes↓ MMP-2 and MMP-9 activities↓ PI3K–AKT–mTOR signaling pathway↓ | [62] |
| Melittin/Bee Venom | honey bees (Apis mellifera) | melanoma | B16F10, A375SM, SK-MEL-28 | growth↓, colony-forming ability↓, migration↓, invasion↓, apoptosis↑, cleaved caspase-3 and -9↑, pPI3K↓, pAKT↓, mTOR↓, ERK↓, p38↓ | [63] |
| Kaempferol | piper | melanoma | A375 | proliferation↓, migration↓, colony formation↓, apoptosis↑, G2/M cell cycle arrest↑, pmTOR↓, pPI3K↓, pAKT↓ | [64] |
| Euplotin C | Euplotes crassus | melanoma | A375, 501Mel, MeWo, HDFa | viability↓, apoptosis↑, migration↓, B-Raf↓, pERK 1/2↓, pAKT↓ | [65] |
| Lycorine | Lycoris radiate spider lilies (Lycoris), daffodils (Narcissus) and snowdrops (Galanthus) | malignant melanoma | HEMa, A375 | proliferation↓, cell migration↓, invasion↓, apoptosis↑, caspase-3↑, Bax↑, Bcl-2↓, pAKT↓, pmTOR↓, 4EBP1↓ | [66] |
| Oxyfadichalcone C | Oxytropis falcate | melanoma | A375 | proliferation↓, G1 phase arrest↑, apoptosis↑, migration↓, invasion↓, p27↑, cyclin D1↓, ppRb↓, pIntegrin β1↓, MMP-2/9↓, metastasis↓, pPDK1↓, pAKT↓, pGSK-3β↓, pmTOR↓, pp70s6k↓, pERK↓ | [67] |
| Isoliquiritigenin | Glycyrrhizae Radix | melanoma | A375 | proliferation↓, G2/M cell cycle arrest↑, mTOR↓, RICTOR↓, pAKT↓, pGSK-3β↓ | [68] |
| Muniziqi granule/harmine | Peganum harmala, Cichorium intybus, Dracocephalum moldavica, Ocimum basilicum, Althaea rosea, and Nigella glandulifera | melanoma | B16F10 | proliferation↓, autophagy, autophagosome formation↑, LC3-II↑, P62↓, apoptosis↑, G1 cell cycle arrest↑, pAKT↓, pmTOR↓, pERK1/2↓ | [69] |
| Apigenin | Various fruits and vegetables | A375, C8161 | proliferation↓, migration↓, invasion↓, apoptosis↑, G2/M cell cycle arrest↑, cleaved caspase-3↑, cleaved PARP↑, pERK1/2↓, pAKT↓, pmTOR↓ | [70] | |
| Casticin | Fructus viticis | melanoma | B16F10 | migration↓, invasion↓, MMP-9↓, MMP-2↓, MMP-1↓, FAK↓, 14-3-3↓, GRB2↓, AKT↓, NF-κB↓, p65↓, SOS-1↓, p-EGFR↓, p-JNK 1/2↓, uPA↓, Rho A↓ | [71] |
| Chrysin | passionflower, silver linden, honey, propolis | melanoma | A375.S2 | mobility↓, migration↓, invasion↓, MMP-2 activity↓, GRB2↓, SOS-1↓, PKC↓, pAKT (Thr308)↓, NF-κBp65↓, NF-κBp50↓ uPA↓, N-cadherin↓, MMP-1↓, MMP-2↓, VEGF↓, E-cadherin↑, NF-κBp65↓ | [72] |
| Berberine | the roots and bark of Berberis genus | melanoma | A375.S2 | morphological changes↑, viability↓, mobility↓, migration↓, invasion↓, MMP-9 activity↓, MMP-1↓, MMP-13↓, E-cadherin↑, N-cadherin↓, RhoA↓, ROCK1↓, SOS-1↓, GRB2↓, Ras↓, pERK1/2↓, pc-Jun↓, p-FAK↓, pAKT↓, NF-κB↓, uPA↓, PKC↓, PI3K↓ | [73] |
| caffeic acid n-butyl ester | skin carcinoma | A431 | Apoptosis↑, Bax↑, Bcl-2↓, ROS↑, MMP↓, G2 phase arrest↑, migration↓, pmTOR↓, pPI3K, pAKT↓ | [74] |
| Compounds | Plants | Cancer Types | Model | Treatment | Mechanisms | Ref. |
|---|---|---|---|---|---|---|
| Isorhamnetin | Hippophae rhamnoides L. | melanoma | C57BL/6 mice injected with B16F10 cells, 1 × 105 | 20 mg/kg per day; for 7 days | Proliferation↓, Ki67↓ | [31] |
| Curcumol | Curcuma wenyujin | melanoma | C57BL/6 mice injected (s.c. into the right lower paw and i.v. into the tail vein) with B16 cells, 2 × 106 | 20 mg/kg, i.p.; 3 times per week; for 30 days | proliferation↓, growth↓ invasion↓, metastasis↓ | [33] |
| Polyphyllin I | Paris polyphylla | melanoma | male BALB/c -nude mice with A375 cells, 2 × 106 | Polyphyllin I 5 mg/kg; i.p.; once a day for 35 days | tumor weight↓, tumor size↓ apoptosis↑, TUNEL positive cells↑, Ki67↓ | [36] |
| Herbacetin | Flaxseed, ramose scouring rush herb | SCC | -DMBA/TPA model; Hairless SKH:HR-1-hrBr (SKH-1) (8–9 weeks old), initiation with DMBA (200 nmol), and promotion with 17 nmol of TPA in acetone, topically applied twice weekly for 20 weeks -solar–UV induced-skin carcinogenesis model; exposed to solar–UV (48 kJ/UVA/2.9 kJ/UVB) three times weekly for 12 weeks -xenograft model; Athymic mice (Cr:NIH(S), NIH Swiss nude, 6–9-wk-old) with SK-MEL-5 cells, 3 × 106 | -DMBA/TPA model; 100 or 500 nmol of herbacetin applied to dorsal mouse skin at 30 min before TPA treatment. -solar–UV induced-skin tumor mouse model; after 20 weeks later, herbacetin 100 or 500 nmol for an additional 7 weeks -xenograft model; herbacetin 0.2 and 1 mg/kg; i.p. injected three times per week for 15 days | skin papillomas↓, tumor volume↓, Ki67↓, pAKT↓, pGSK3β↓, pRSK↓, ODC↓ | [38] |
| Luteolin | Reseda luteola | Melanoma | Female BALB/c -nude mice with A375 cells, 1 × 107 | 100 mg/kg/day, i.p. for 22 days | tumor growth↓ PI3K/AKT↓, MMP-2↓, MMP-9↓ | [41] |
| Sinomenine | Sinomenium acutum | Melanoma | xenograft model; BALB/c nude mice (6-week-old) by subcutaneously injection with B16-F10 cells | 100 mg/kg/day; s.c., daily for 35 days. | tumor weight↓, tumor volume↓, Ki67↓, PCNA↓ | [45] |
| Syringic acid | Euterpe oleracea, Rhus javanica | non-melanoma | SKH-1 hairless mouse, UVB (0.2 J/cm2) exposure (three times per week for 22 weeks) | 0.2 or 1 mM per mouse in 200 μL acetone on the dorsal surface 1 h before UVB irradiation | UVB-induced skin tumor↓, COX-2↓, MMP-13↓, | [48] |
| Ginkgo biloba Exocarp Extract | C57BL/6J female mice (6-week-old) by subcutaneously injection with B16-F10, 2.0 × 106 cells | 50, 100, 200 mg/kg by intragestic gavage, once a day for 17 days | tumor growth↓, lung metastasis↓, MMP-9↓ | [51] | ||
| α-Mangostin | pericarp of mangosteen | Skin cancer | DMBA (60 μg)/TPA (4 μg) induced skin carcinogenesis model in ICR female mice, once a week for 20 weeks | 5 and 20 mg/kg, (dissolved in 0.2 mL olive oil) once a day, starting from the day after TPA was topically applied, i.p. for 20 weeks | Skin papilloma↓, growth↓, LC3↑, LC3-II↑, Beclin1↑, LC3-I↓, p62↓, Bax↑, cleaved caspase-3↑, cleaved PARP↑, Bad↑, Bcl-2↓, Bcl-xl↓, apoptosis↑, p-PI3K↓, p-AKT↓, p-mTOR↓ | [54] |
| Silibinin and its 2,3-dehydro-derivative | Silybum marianum (L.) Gaertn., Asteraceae | BCC | ectopic allograft model; five weeks old nude mice (Foxn1nu/nu) by subcutaneously injection with 1 × 106 ASZ cells | silibinin (200 mg/kg in 0.5% CMC) or DHS (200 mg/kg); oral administration, 6 days per week for a total of 7 weeks | tumor growth↓, PCNA↓, cyclin D1↓, proliferation↓, NF-κB↓, AP-1↓, c-Fos↓ | [56] |
| Curcumin | rhizome of Curcuma longa | Melanoma | BALB/c nude female mice (6-week-old) by subcutaneously injection with A375 cells (1 × 107/mL) | 25 mg/kg by i.p. injections, every day for 3 weeks | growth↓ | [60] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, S.-Y.; Chae, J.-I.; Kwak, A.-W.; Lee, M.-H.; Shim, J.-H. Alternative Options for Skin Cancer Therapy via Regulation of AKT and Related Signaling Pathways. Int. J. Mol. Sci. 2020, 21, 6869. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186869
Hwang S-Y, Chae J-I, Kwak A-W, Lee M-H, Shim J-H. Alternative Options for Skin Cancer Therapy via Regulation of AKT and Related Signaling Pathways. International Journal of Molecular Sciences. 2020; 21(18):6869. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186869
Chicago/Turabian StyleHwang, Sun-Young, Jung-Il Chae, Ah-Won Kwak, Mee-Hyun Lee, and Jung-Hyun Shim. 2020. "Alternative Options for Skin Cancer Therapy via Regulation of AKT and Related Signaling Pathways" International Journal of Molecular Sciences 21, no. 18: 6869. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186869