Dysmetria and Errors in Predictions: The Role of Internal Forward Model


 and
and 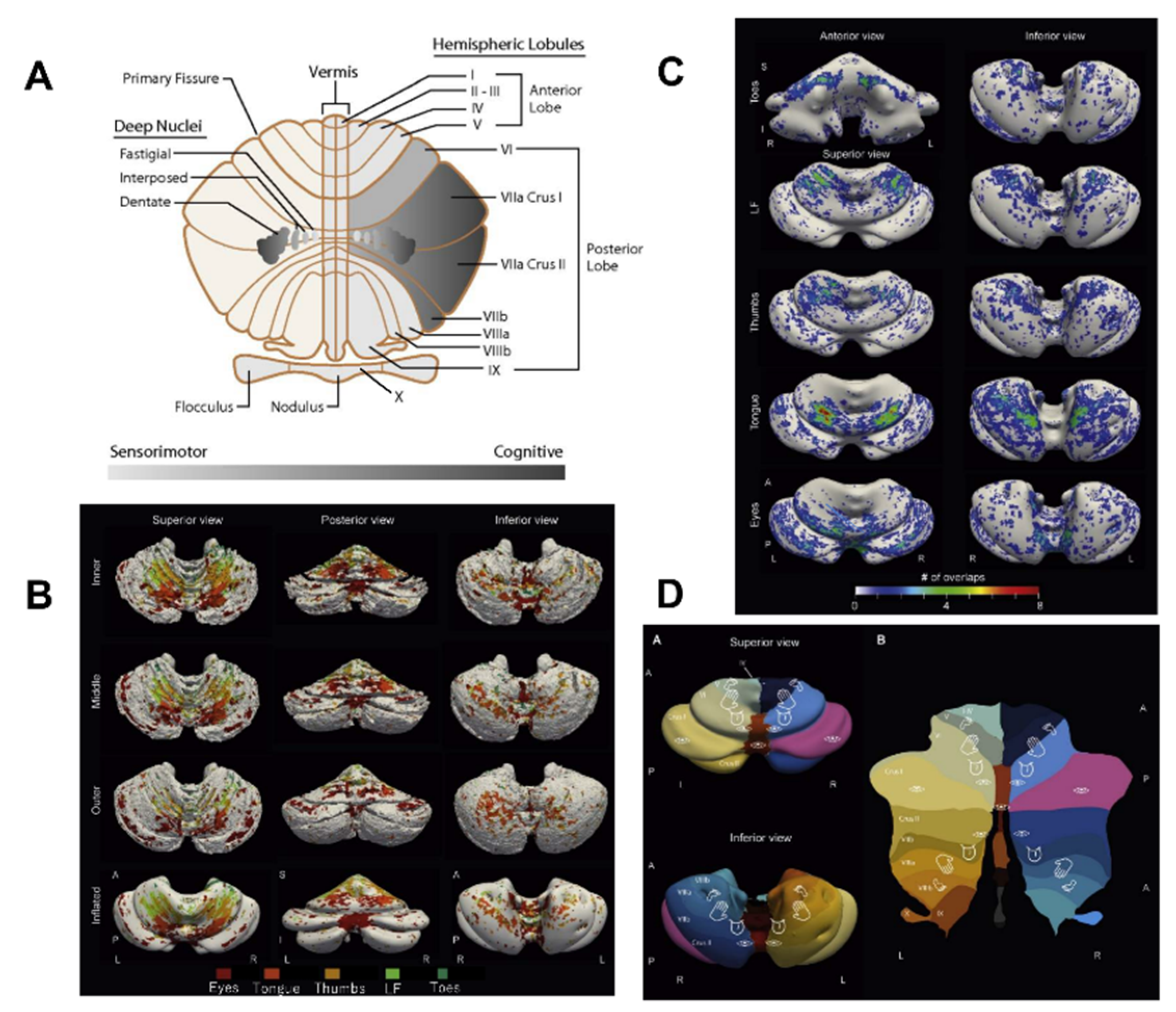{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Cerebellum and Predictions: Relevant Anatomy
2.1. Macroanatomy-Physiological Classification
2.1.1. Rostrocaudal Gradient
2.1.2. Medio-lateral Gradient
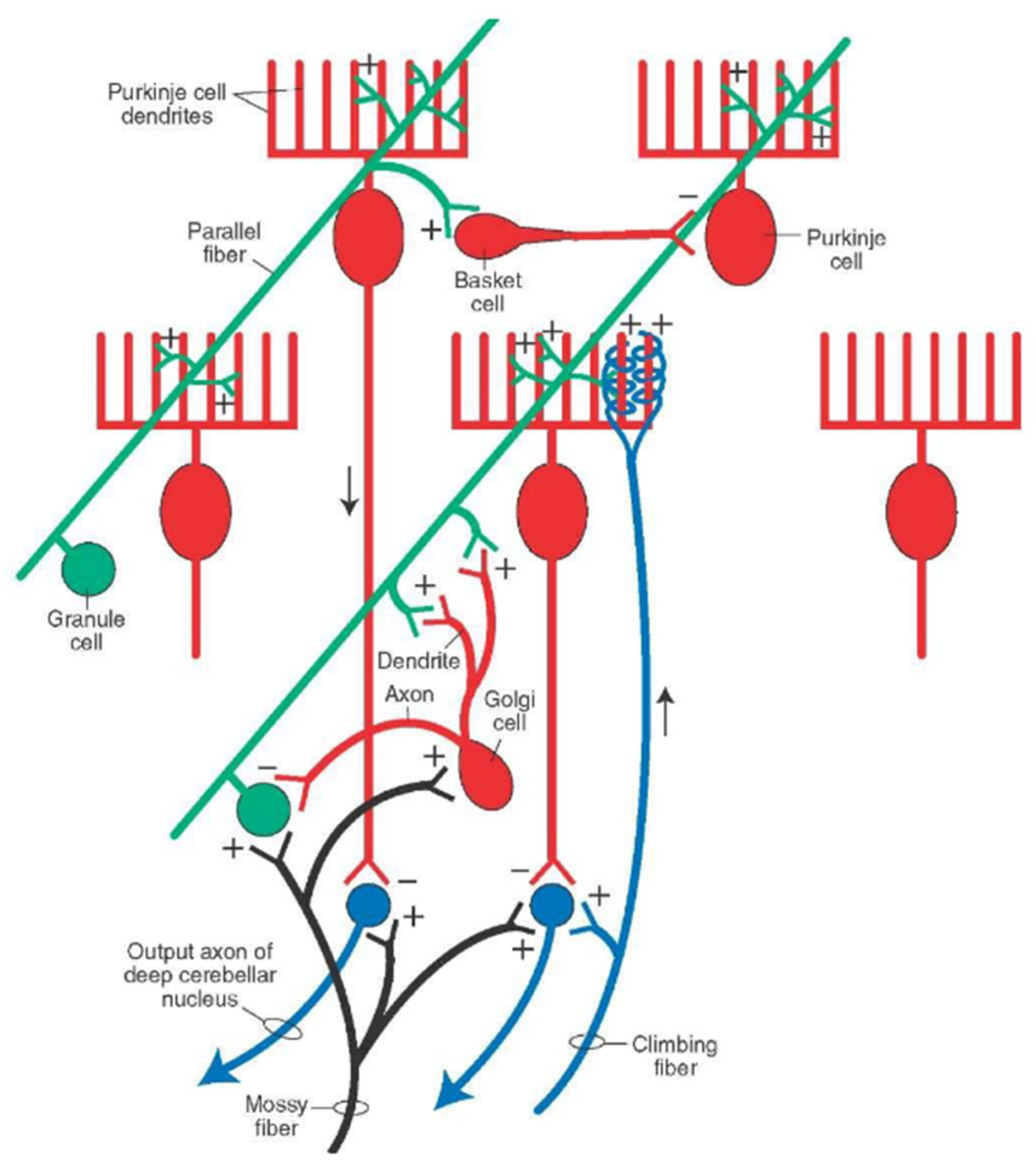
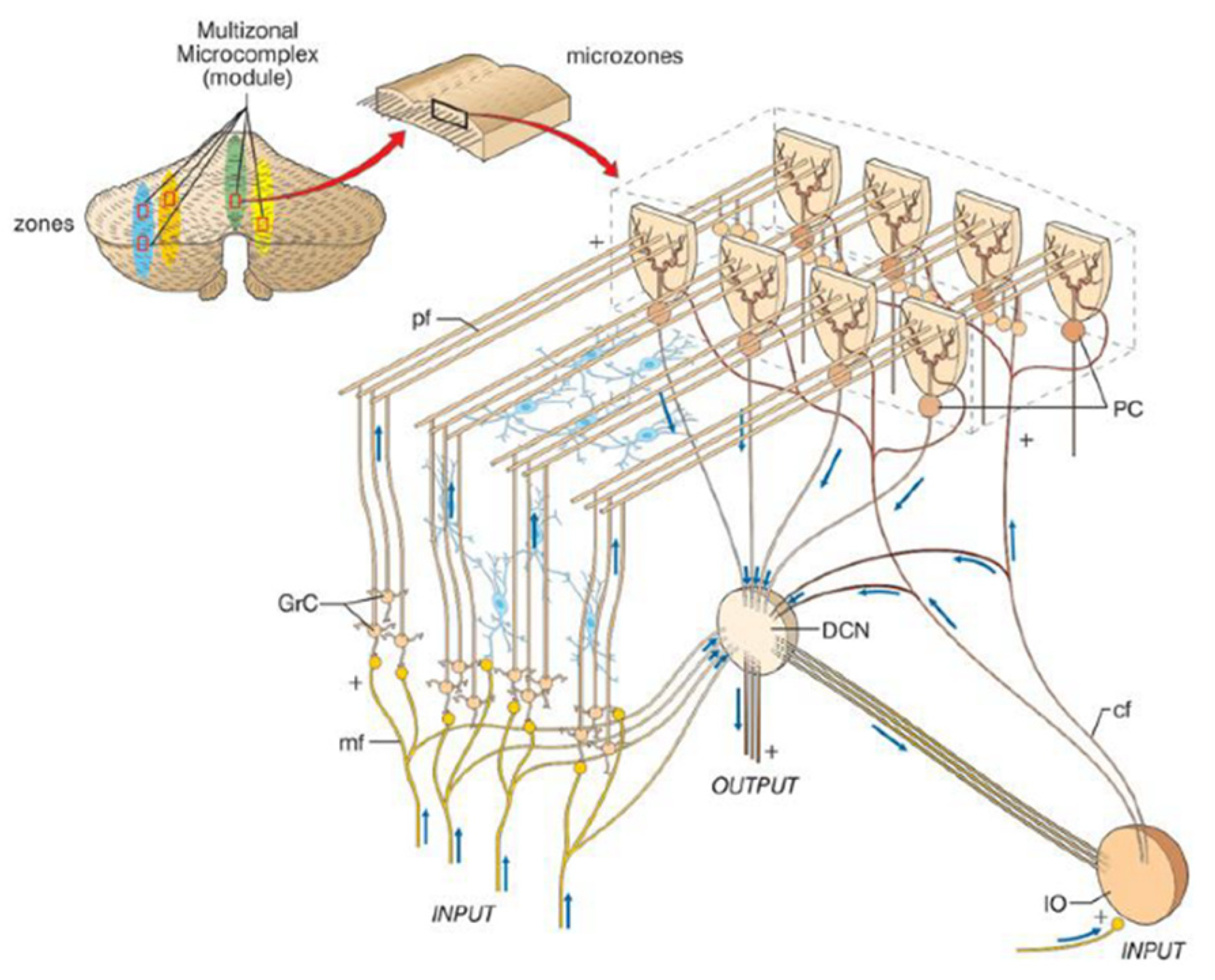
2.2. Microanatomy of the Cerebellar Cortex
2.2.1. Microzones
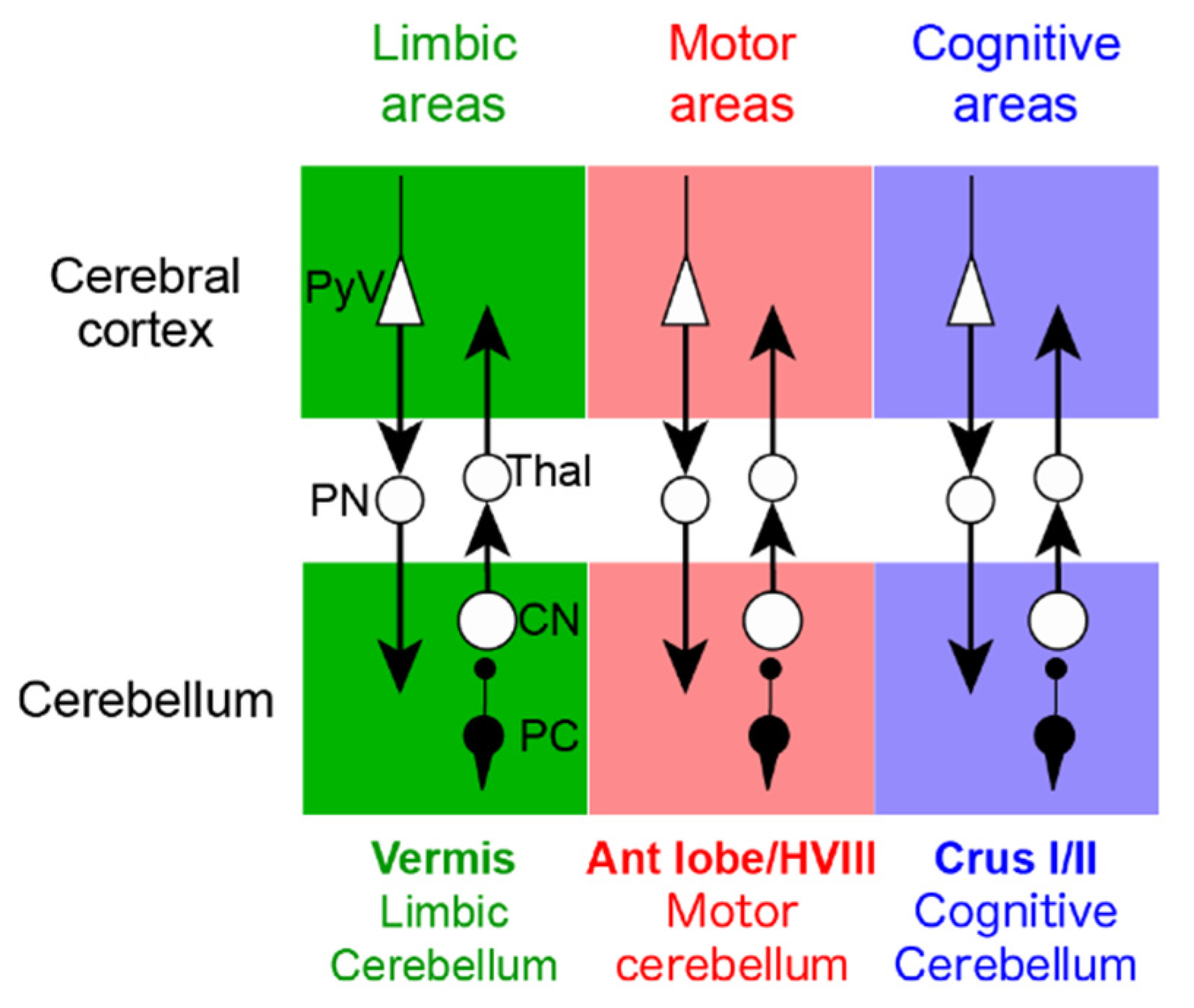
2.2.2. Cerebellar Modules
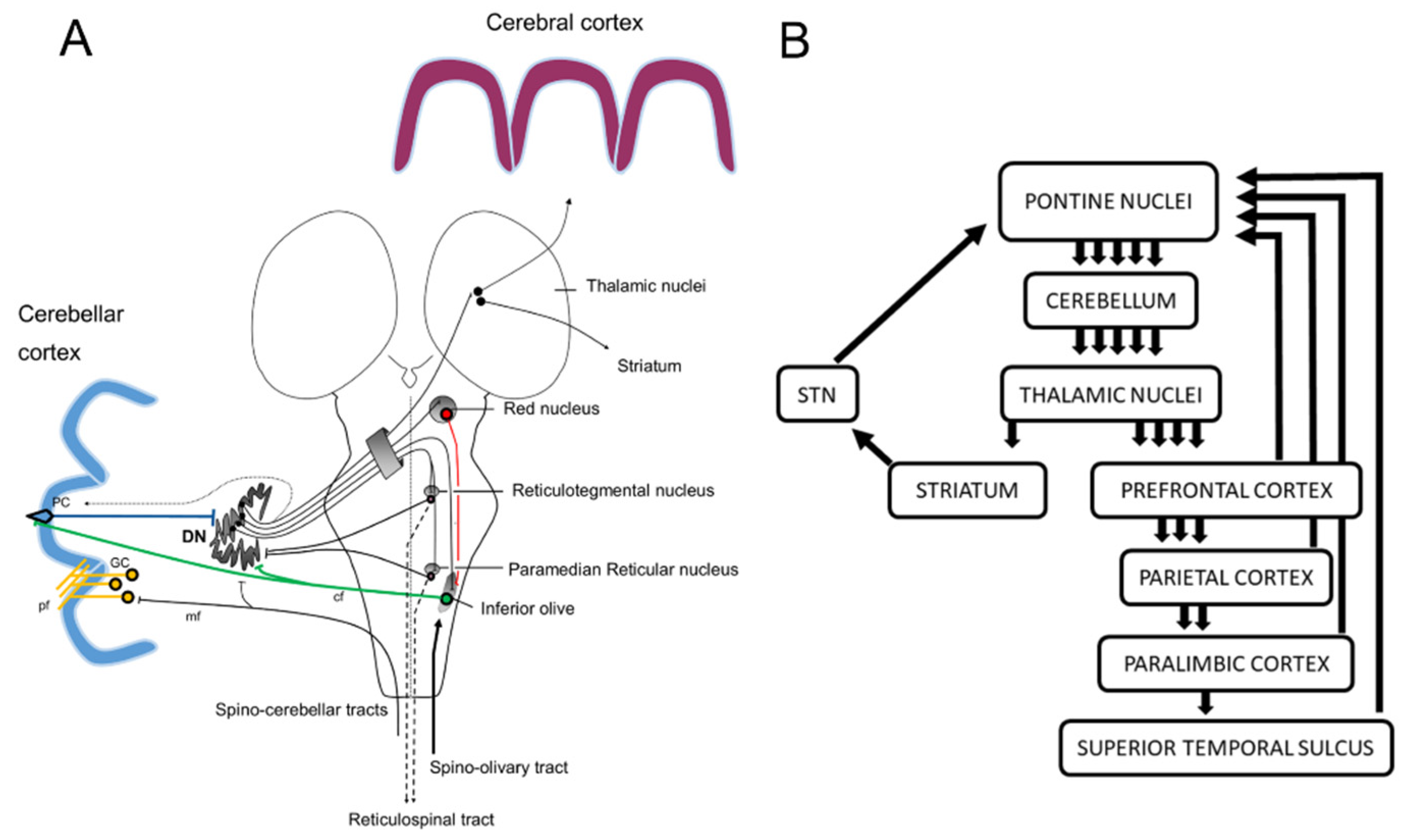
2.3. Cerebellar Projections to the Cerebral Cortex
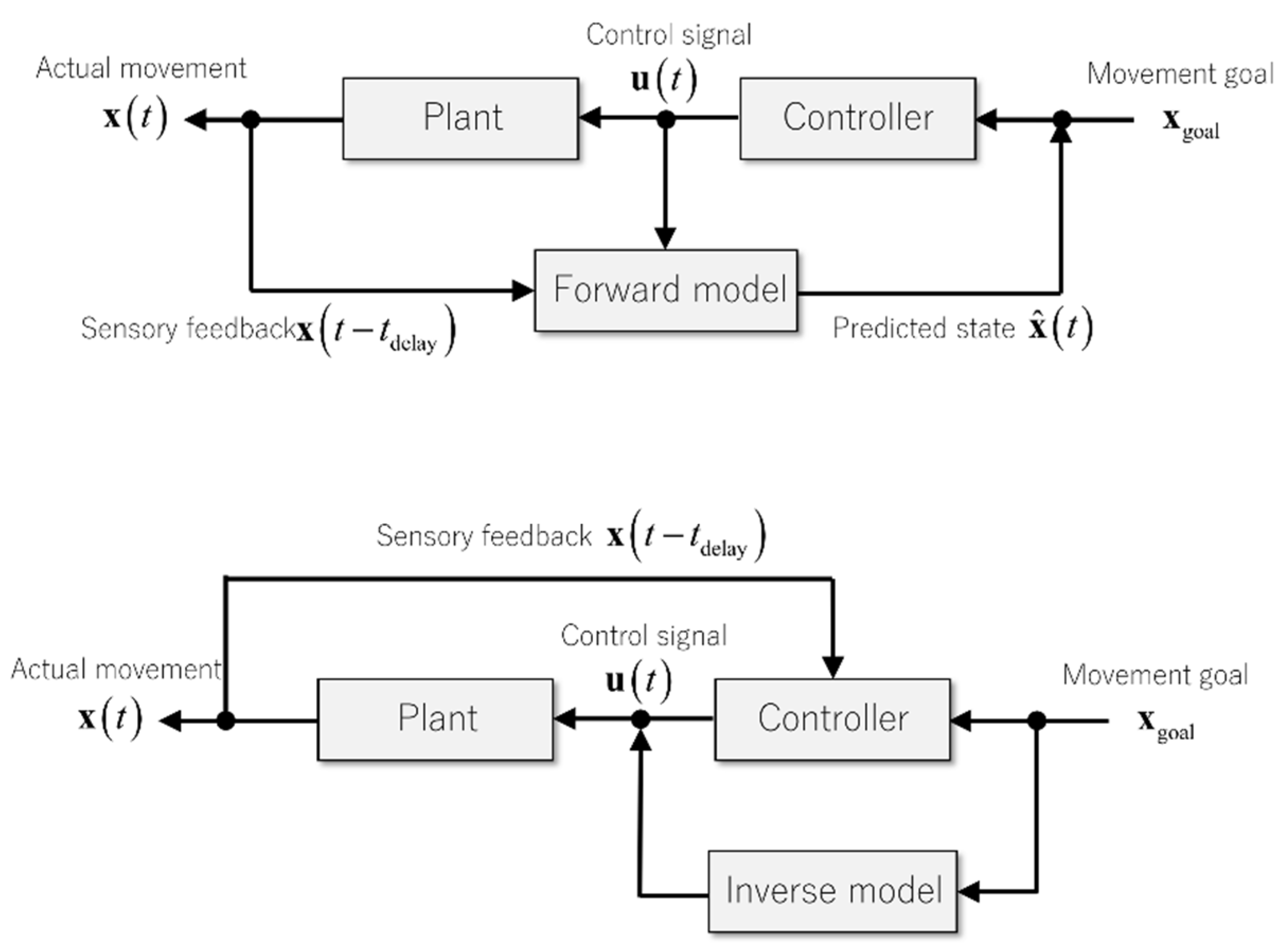
3. Internal Models for Motor Control and Motor Learning
3.1. Internal Model for Delay Compensation
3.2. Internal Model for Motor Learning
3.3. Internal Model for Interlimb Coordination
4. Patterns of Discharges in the Cerebellar Cortex
4.1. Simple Spikes
4.2. Complex Spikes
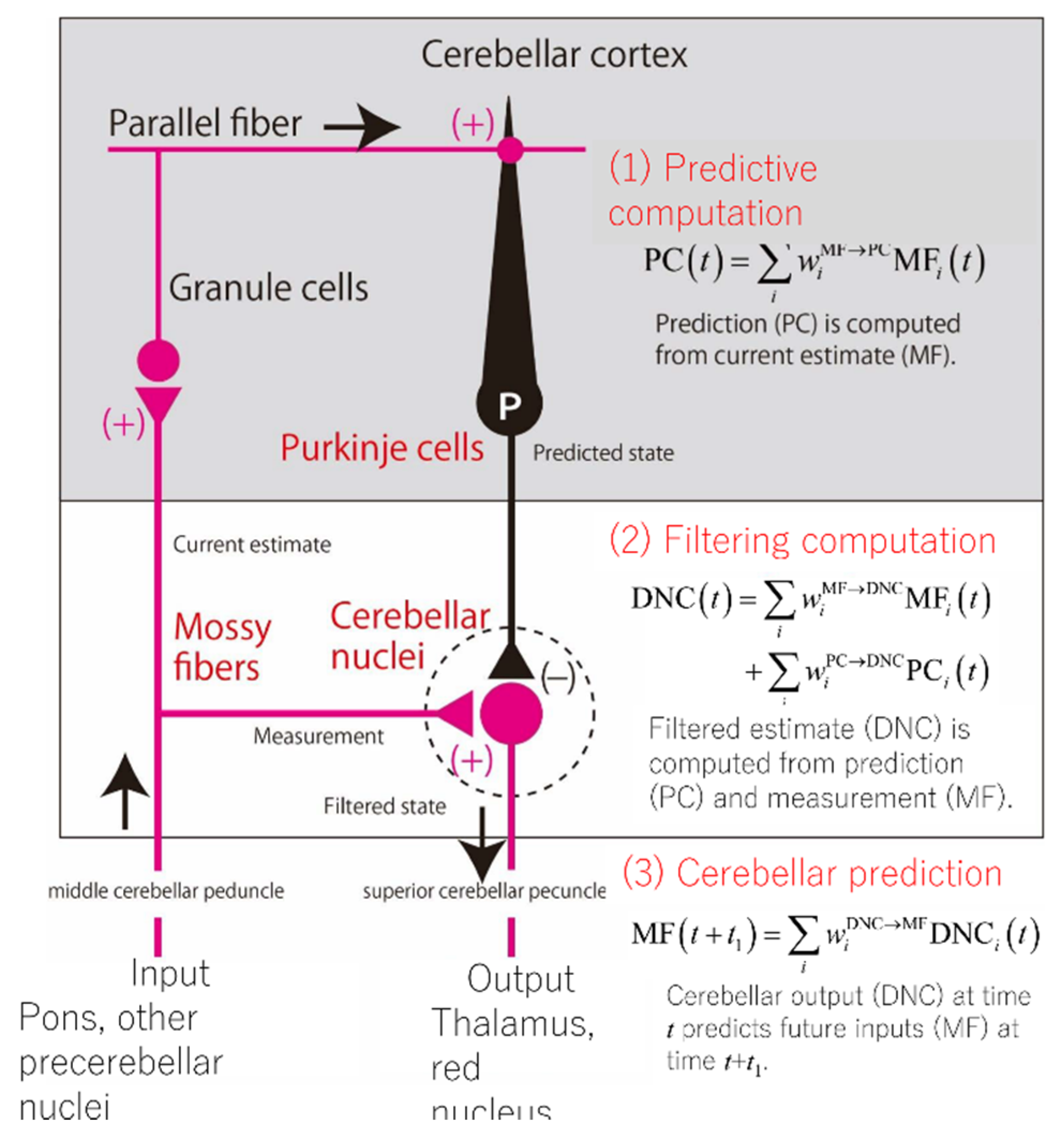
5. The Cerebellum as a Locus for Internal Forward Model
5.1. Behavioral Evidence for the Internal-Model Hypothesis of the Cerebellum
5.2. Neural Evidence for the Internal Forward Model Hypothesis of the Cerebellum
6. Cerebellar Dysmetria: Kinematic and EMG Features
6.1. EMG and Kinematic Features Associated with Dysmetria
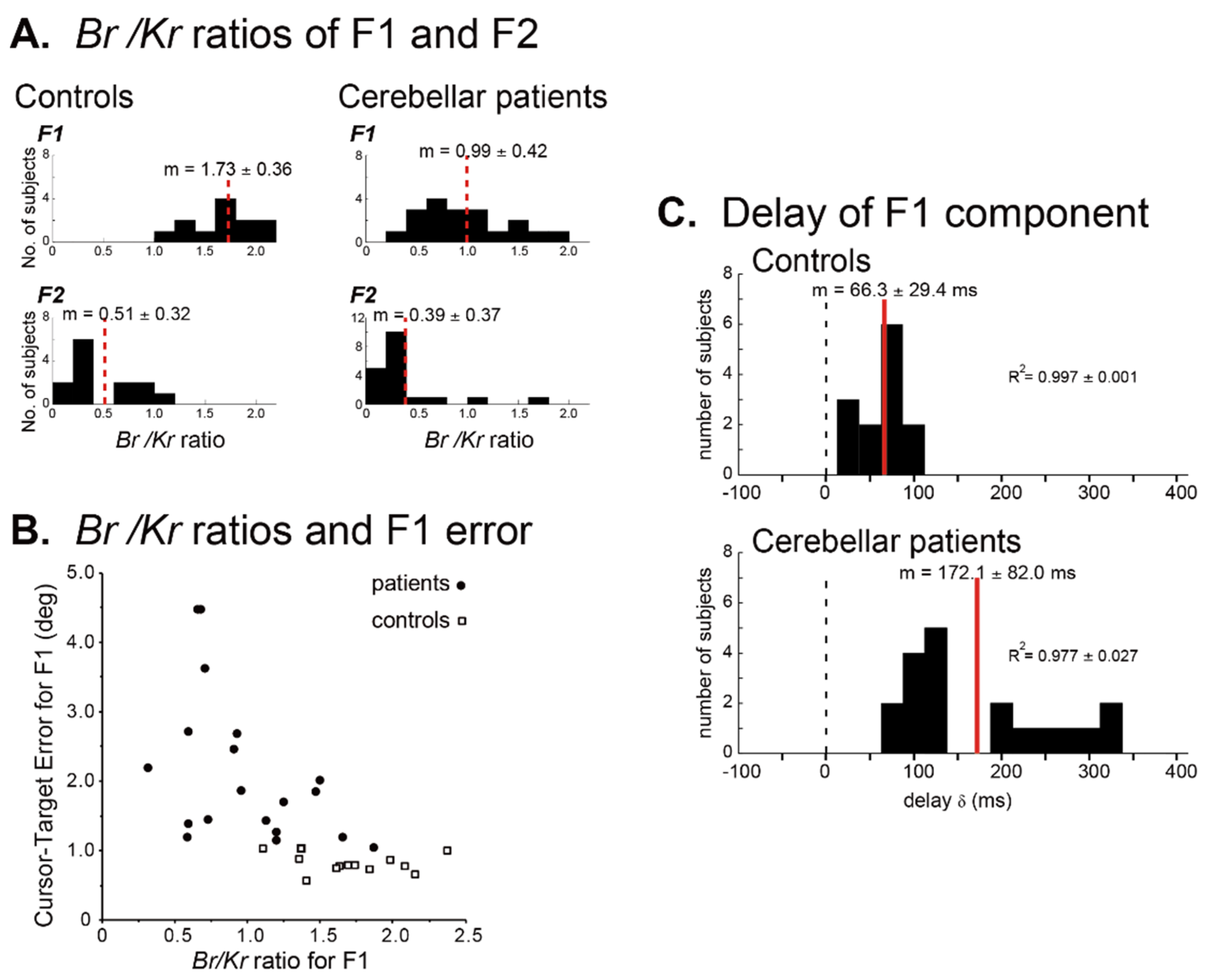
6.2. Poor Prediction and Compensatory Feedback Corrections
6.3. Algorithm of Predictive Control: Timing Versus Synergy?
6.3.1. Timing
6.3.2. Synergy
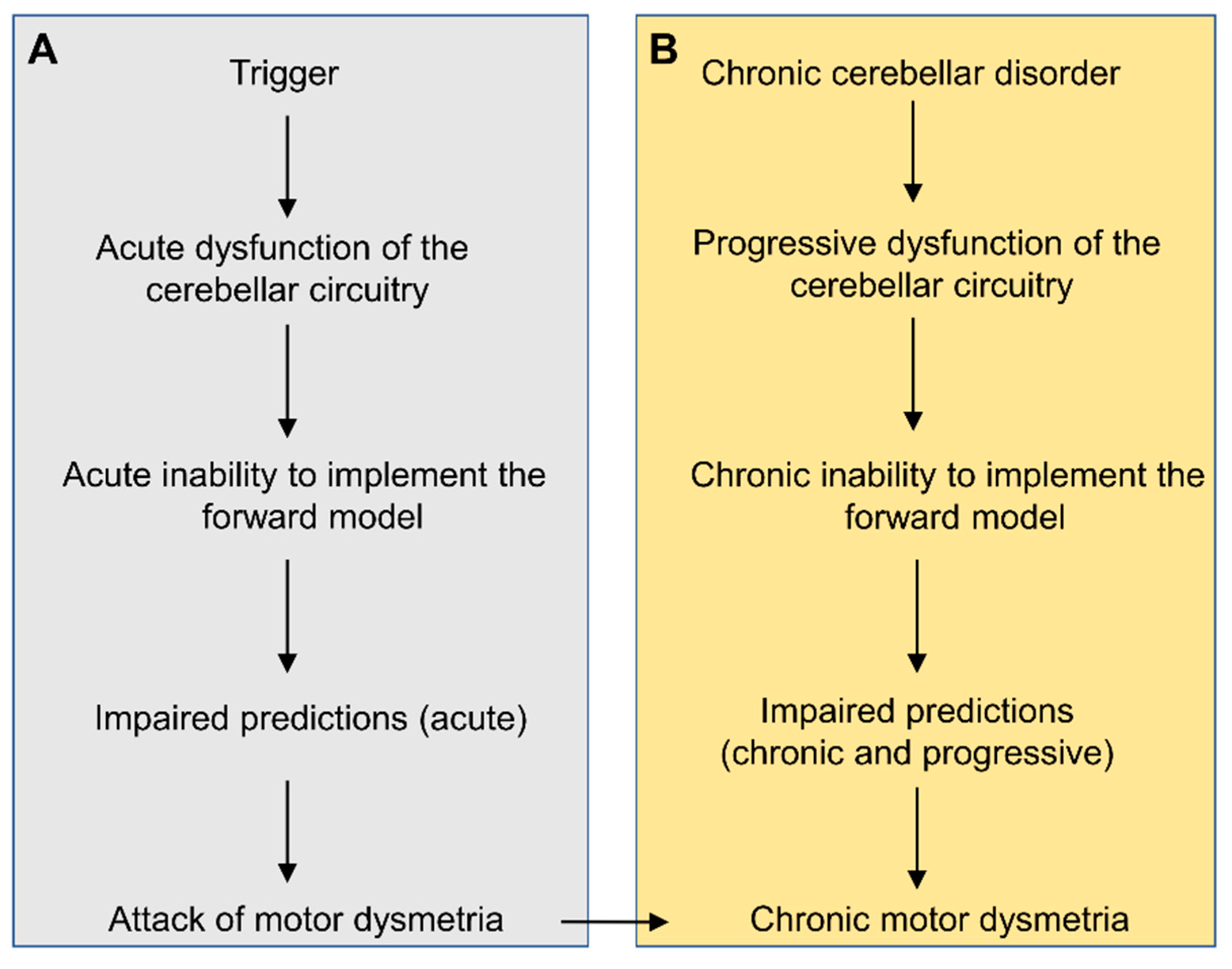
6.4. Dysmetria Spanning Multitudes of Time Scales
7. General Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Manto, M.; Bower, J.M.; Conforto, A.B.; Delgado-Garcia, J.M.; da Guarda, S.N.; Gerwig, M.; Habas, C.; Hagura, N.; Ivry, R.B.; Mariën, P.; et al. Consensus paper: Roles of the cerebellum in motor control-the diversity of ideas on cerebellar involvement in movement. Cerebellum 2012, 11, 457–487. [Google Scholar] [CrossRef]
- Garcin, R. The ataxias. In Handbook of Clinical Neurology. Volume 1 Disturbances of Nervous Function; Vinken, P.J., Bruyn, G.W., Eds.; North Holland Publishing Company: Amsterdam, The Netherland, 1969; pp. 309–335. [Google Scholar]
- Schmahmann, J.D.; Caplan, D. Cognition, emotion and the cerebellum. Brain 2006, 129, 290–292. [Google Scholar] [CrossRef]
- Manto, M. Mechanisms of human cerebellar dysmetria: Experimental evidence and current conceptual bases. J. Neuroeng. Rehabil. 2009, 6, 10. [Google Scholar] [CrossRef] [PubMed]
- Schmahmann, J.D. The cerebellum and cognition. Neurosci. Lett. 2019, 688, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Babinski, J. Exposé des Travaux Scientifiques; Masson: Paris, Frrance, 1913. [Google Scholar]
- Day, B.L.; Thompson, P.D.; Harding, A.E.; Marsden, C.D. Influence of vision on upper limb reaching movements in patients with cerebellar ataxia. Brain 1998, 121, 357–372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordan, M.I.; Rumelhart, D.E. Forward models: Supervised learning with a distal teacher. Cogn. Sci. 1992, 16, 307–354. [Google Scholar] [CrossRef]
- Wolpert, D.M.; Miall, R.C. Forward models for physiological motor control. Neural Netw. 1996, 9, 1265–1279. [Google Scholar] [PubMed]
- Wolpert, D.M.; Miall, R.C.; Kawato, M. Internal models in the cerebellum. Trends Cogn. Sci. 1998, 2, 338–347. [Google Scholar] [CrossRef]
- Allen, G.I.; Tsukahara, N. Cerebrocerebellar communication systems. Physiol. Rev. 1974, 54, 957–1006. [Google Scholar] [CrossRef]
- Bostan, A.C.; Dum, R.P.; Strick, P.L. Cerebellar networks with the cerebral cortex and basal ganglia. Trends. Cogn. Sci. 2013, 17, 241–254. [Google Scholar] [CrossRef] [Green Version]
- Strick, P.L.; Dum, R.P.; Fiez, J.A. Cerebellum and nonmotor function. Annu. Rev. Neurosci. 2009, 32, 413–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolliat, Y.; Bazin, P.L.; van der Zung, W. Whole-body somatotopic maps in the cerebelum revealed with 7T fMRI. Neuroimage 2020, 211, 116624. [Google Scholar] [CrossRef] [PubMed]
- Guell, X.; Gabrieli, J.D.E.; Schmahmann, J.D. Triple representation of language, working memory, social and emotion processing in the cerebellum: Convergent evidence from task and seed-based resting-state fMRI analyses in a single large cohort. Neuroimage 2018, 172, 437–449. [Google Scholar] [CrossRef]
- Stoodley, C.J.; Schmahmann, J.D. Functional topography of the human cerebellum. Handb. Clin. Neurol. 2018, 154, 59–70. [Google Scholar] [CrossRef]
- Timmann, D.; Drepper, J.; Frings, M.; Maschke, M.; Richter, S.; Gerwig, M.; Kolb, F.P. The human cerebellum contributes to motor, emotional and cognitive associative learning. A review. Cortex 2010, 46, 845–857. [Google Scholar] [CrossRef] [PubMed]
- Apps, R.; Hawkes, R.; Aoki, S.; Bengtsson, F.; Brown, A.M.; Chen, G.; Ebner, T.J.; Isope, P.; Jörntell, H.; Lackey, E.P.; et al. Cerebellar modules and their role as operational cerebellar processing units: A consensus paper [corrected]. Cerebellum 2018, 17, 654–682. [Google Scholar] [CrossRef] [Green Version]
- Ito, M. The Cerebellum and Neural Control.; Raven Press: New York, NY, USA, 1984. [Google Scholar]
- Shinoda, Y.; Sugiuchi, Y.; Futami, T.; Izawa, R. Axon collaterals of mossy fibers from the pontine nucleus in the cerebellar dentate nucleus. J. Neurophysiol. 1992, 67, 547–560. [Google Scholar] [CrossRef]
- Wu, H.S.; Sugihara, I.; Shinoda, Y. Projection patterns of single mossy fibers originating from the lateral reticular nucleus in the rat cerebellar cortex and nuclei. J. Comp. Neurol. 1999, 411, 97–118. [Google Scholar] [CrossRef]
- Ishikawa, T.; Tomatsu, S.; Izawa, J.; Kakei, S. The cerebro-cerebellum: Could it be loci of forward models? Neurosci. Res. 2016, 104, 72–79. [Google Scholar] [CrossRef] [Green Version]
- D’Angelo, E.; Casali, S. Seeking a unified framework for cerbellar function and dysfunction: From circuit operations to cognition. Front. Neural Circuit 2013, 6, 116. [Google Scholar] [CrossRef] [Green Version]
- Apps, R.; Hwaks, R. Cerebellar cortical organization: One map hypothesis. Nat. Rev. Neurosci. 2009, 10, 670–681. [Google Scholar] [CrossRef] [PubMed]
- Buckner, R.L.; Krienen, F.M.; Castellanos, A.; Diaz, J.C.; Yeo, B.T. The organization of the human cerebellum estimated by intrinsic functional connectivity. J. Neurophysiol. 2011, 106, 2322–2345. [Google Scholar] [CrossRef] [PubMed]
- Kelly, R.M.; Strick, P.L. Cerebellar loops with motor cortex and prefrontal cortex of a nonhuman primate. J. Neurosci. 2003, 23, 8432–8444. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmolesky, M.T.; Wang, Y.; Hanes, D.P.; Thompson, K.G.; Leutgeb, S.; Schall, J.D.; Leventhal, A.G. Signal timing across the macaque visual system. J. Neurophysiol. 1998, 79, 3272–3278. [Google Scholar] [CrossRef]
- More, H.L.; Donelan, J.M. Scaling of sensorimotor delays in terrestrial mammals. Proc. Biol. Sci. 2018, 285, 20180613. [Google Scholar] [CrossRef] [Green Version]
- More, H.L.; Hutchinson, J.R.; Collins, D.F.; Weber, D.J.; Aung, S.K.; Donelan, J.M. Scaling of sensorimotor control in terrestrial mammals. Proc. Biol. Sci. 2010, 277, 3563–3568. [Google Scholar] [CrossRef] [Green Version]
- Wolpert, D.M.; Ghahramani, Z.; Jordan, M.I. An internal model for sensorimotor integration. Science 1995, 269, 1880–1882. [Google Scholar] [CrossRef]
- Todorov, E. Optimality principles in sensorimotor control. Nat. Neurosci. 2004, 7, 907–915. [Google Scholar] [CrossRef] [Green Version]
- Atkeson, C.G. Learning arm kinematics and dynamics. Annu. Rev. Neurosci. 1989, 12, 157–183. [Google Scholar] [CrossRef]
- Kawato, M.; Furukawa, K.; Suzuki, R. A hierarchical neural-network model for control and learning of voluntary movement. Biol. Cybern. 1987, 57, 169–185. [Google Scholar] [CrossRef]
- Mazzoni, P.; Krakauer, J.W. An implicit plan overrides an explicit strategy during visuomotor adaptation. J. Neurosci. 2006, 26, 3642–3645. [Google Scholar] [CrossRef]
- Tseng, Y.W.; Diedrichsen, J.; Krakauer, J.W.; Shadmehr, R.; Bastian, A.J. Sensory prediction errors drive cerebellum-dependent adaptation of reaching. J. Neurophysiol. 2007, 98, 54–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shadmehr, R.; Smith, M.A.; Krakauer, J.W. Error correction, sensory prediction, and adaptation in motor control. Annu. Rev. Neurosci. 2010, 33, 89–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rescorla, R.A.; Wagner, A.R. A theory of Pavlovian conditioning: Variations in the effectiveness of reinforcement and nonreinforcement. In Classical conditioning II. Current Research and Theory; Black, A.H., Prokasy, W.F., Eds.; Appleton-Century-Crofts: New York, NY, USA, 1972; pp. 64–99. [Google Scholar]
- Barto, A.G.; Sutton, R.S.; Anderson, C.W. Neuronlike adaptive elements that can solve difficult learning control problems. IEEE Trans. Syst. Man Cybern. 1983, SMC-14, 834–846. [Google Scholar] [CrossRef]
- Nowak, D.A.; Hermsdorfer, J.; Rost, K.; Timmann, D.; Topka, H. Predictive and reactive finger force control during catching in cerebellar degeneration. Cerebellum 2004, 3, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Nowak, D.A.; Timmann, D.; Hermsdorfer, J. Dexterity in cerebellar agenesis. Neuropsychologia 2007, 45, 696–703. [Google Scholar] [CrossRef] [PubMed]
- Llinas, R.; Sugimori, M. Electrophysiological properties of in vitro Purkinje cell somata in mammalian cerebellar slices. J. Physiol. 1980, 305, 171–195. [Google Scholar] [CrossRef]
- Palmer, L.M.; Clark, B.A.; Grundemann, J.; Roth, A.; Stuart, G.J.; Hausser, M. Initiation of simple and complex spikes in cerebellar Purkinje cells. J. Physiol. 2010, 588, 1709–1717. [Google Scholar] [CrossRef] [Green Version]
- Voogd, J. Cerebellar zones: A personal history. Cerebellum 2011, 10, 334–350. [Google Scholar] [CrossRef] [Green Version]
- Manto, M.; Habas, C. Le Cervelet, de L’anatomie et la Physiologie à la Clinique Humaine; Springer-Verlag: Paris, France, 2013. [Google Scholar]
- Popa, L.S.; Hewitt, A.L.; Ebner, T.J. Purkinje cell simple spike discharge encodes error signals consistent with a forward internal model. Cerebellum 2013, 12, 331–333. [Google Scholar] [CrossRef]
- Popa, L.S.; Hewitt, A.L.; Ebner, T.J. The cerebellum for jocks and nerds alike. Front. Syst. Neurosci. 2014, 8, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brunel, N.; Hakim, V.; Isope, P.; Nadal, J.P.; Barbour, B. Optimal information storage and the distribution of synaptic weights: Perceptron versus Purkinje cell. Neuron 2004, 43, 745–757. [Google Scholar] [CrossRef] [PubMed]
- Bauswein, E.; Kolb, F.P.; Leimbeck, B.; Rubia, F.J. Simple and complex spike activity of cerebellar Purkinje cells during active and passive movements in the awake monkey. J. Physiol. 1983, 339, 379–394. [Google Scholar] [CrossRef] [PubMed]
- Popa, L.S.; Hewitt, A.L.; Ebner, T.J. Predictive and feedback performance errors are signaled in the simple spike discharge of individual Purkinje cells. J. Neurosci. 2012, 32, 15345–15358. [Google Scholar] [CrossRef]
- Hewitt, A.L.; Popa, L.S.; Pasalar, S.; Hendrix, C.M.; Ebner, T.J. Representation of limb kinematics in Purkinje cell simple spike discharge is conserved across multiple tasks. J. Neurophysiol. 2011, 106, 2232–2247. [Google Scholar] [CrossRef]
- Popa, L.S.; Streng, M.L.; Ebner, T.J. Long-Term Predictive and Feedback Encoding of Motor Signals in the Simple Spike Discharge of Purkinje Cells. eNeuro 2017, 4. [Google Scholar] [CrossRef]
- Otis, T.S. Simple Spikes. In Essentials of Cerebellum and Cerebellar Disorders; Gruol, D.L., Koibuchi, N., Manto, M., Molinari, M., Schmahmann, J.D., Shen, Y., Eds.; Springer Nature: Cham, Switzlerland, 2016; pp. 299–303. [Google Scholar]
- Larry, N.; Yarkoni, M.; Lixenberg, A.; Joshua, M. Cerebellar climbing fibers encode expected reward size. Elife 2019, 8. [Google Scholar] [CrossRef]
- Ito, M. The Cerebellum Brain for an Implicit Self; FT Press: Upper Saddle River, NJ, USA, 2012. [Google Scholar]
- Ito, M.; Sakurai, M.; Tongroach, P. Climbing fibre induced depression of both mossy fibre responsiveness and glutamate sensitivity of cerebellar Purkinje cells. J. Physiol. 1982, 324, 113–134. [Google Scholar] [CrossRef]
- Kitazawa, S.; Kimura, T.; Yin, P.B. Cerebellar complex spikes encode both destinations and errors in arm movements. Nature 1998, 392, 494–497. [Google Scholar] [CrossRef]
- Herzfeld, D.J.; Kojima, Y.; Soetedjo, R.; Shadmehr, R. Encoding of action by the Purkinje cells of the cerebellum. Nature 2015, 526, 439–442. [Google Scholar] [CrossRef] [Green Version]
- Ohmae, S.; Medina, J.F. Climbing fibers encode a temporal-difference prediction error during cerebellar learning in mice. Nat. Neurosci. 2015, 18, 1798–1803. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kagamihara, Y.; Kakei, S. A new method for functional evaluation of motor commands in patients with cerebellar ataxia. PLoS ONE 2015, 10, e0132983. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kagamihara, Y.; Tomatsu, S.; Kakei, S. The functional role of the cerebellum in visually guided tracking movement. Cerebellum 2012, 11, 426–433. [Google Scholar] [CrossRef] [PubMed]
- Mitoma, H.; Adhikari, K.; Aeschlimann, D.; Chattopadhyay, P.; Hadjivassiliou, M.; Hampe, C.S.; Honnorat, J.; Joubert, B.; Kakei, S.; Lee, J.; et al. Consensus Paper: Neuroimmune Mechanisms of Cerebellar Ataxias. Cerebellum 2016, 15, 213–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakei, S.; Lee, J.; Mitoma, H.; Tanaka, H.; Manto, M.; Hampe, C.S. Contribution of the cerebellum to predictive motor control and its evaluation in ataxic patients. Front. Hum. Neurosci. 2019, 13, 216. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, H.; Ishikawa, T.; Kakei, S. Neural evidence of the cerebellum as a state predictor. Cerebellum 2019, 18, 349–371. [Google Scholar] [CrossRef] [Green Version]
- Ishikawa, T.; Tomatsu, S.; Tsunoda, Y.; Lee, J.; Hoffman, D.S.; Kakei, S. Releasing dentate nucleus cells from Purkinje cell inhibition generates output from the cerebrocerebellum. PLoS ONE 2014, 9, e108774. [Google Scholar] [CrossRef] [Green Version]
- Tomatsu, S.; Ishikawa, T.; Tsunoda, Y.; Lee, J.; Hoffman, D.S.; Kakei, S. Information processing in the hemisphere of the cerebellar cortex for control of wrist movement. J. Neurophysiol. 2016, 115, 255–270. [Google Scholar] [CrossRef]
- Tanaka, H.; Ishikawa, T.; Lee, J.; Kakei, S. The cerebre-cerebellum as a locus of forward model; a review. Front. Syst. Neurosci. 2020, 14, 19. [Google Scholar] [CrossRef] [Green Version]
- Hallett, M.; Shahani, B.T.; Young, R.R. EMG analysis of patients with cerebellar deficits. J. Neurol. Neurosurg. Psychiatry 1975, 38, 1163–1169. [Google Scholar] [CrossRef] [Green Version]
- Hallett, M.; Shahani, B.T.; Young, R.R. EMG analysis of stereotyped voluntary movements in man. J. Neurol. Neurosurg. Psychiatry 1975, 38, 1154–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flament, D.; Hore, J. Movement and electromyographic disorders associated with cerebellar dysmetria. J. Neurophysiol. 1986, 55, 1221–1233. [Google Scholar] [CrossRef] [PubMed]
- Hore, J.; Wild, B.; Diener, H.C. Cerebellar dysmetria at the elbow, wrist, and fingers. J. Neurophysiol. 1991, 65, 563–571. [Google Scholar] [CrossRef] [PubMed]
- Manto, M.; Godaux, E.; Jacquy, J. Cerebellar hypermetria is larger when the inertial load is artificially increased. Ann. Neurol. 1994, 35, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Manto, M.; Godaux, E.; Jacquy, J.; Hildebrand, J. Cerebellar hypermetria associated with a selective decrease in the rate of rise of antagonist activity. Ann. Neurol. 1996, 39, 271–274. [Google Scholar] [CrossRef] [PubMed]
- Manto, M.U.; Hildebrand, J.; Jacquy, J. Shift from hypermetria to hypometria in an aberrant recovery following cerebellar infarction. J. Neurol. Sci. 1998, 157, 42–51. [Google Scholar] [CrossRef]
- Hewer, R.L.; Cooper, R.; Morgan, M.H. An investigation into the value of treating intention tremor by weighting the affected limb. Brain 1972, 95, 579–590. [Google Scholar] [CrossRef]
- Hallett, M.; Marsden, C.D. Ballistic flexion movements of the human thumb. J. Physiol. 1979, 294, 33–50. [Google Scholar] [CrossRef]
- Hannaford, B.; Stark, L. Roles of the elements of the triphasic control signal. Exp. Neurol. 1985, 90, 619–634. [Google Scholar] [CrossRef]
- Berardelli, A.; Rothwell, J.C.; Day, B.L.; Kachi, T.; Marsden, C.D. Duration of the first agonist EMG burst in ballistic arm movements. Brain Res. 1984, 304, 183–187. [Google Scholar] [CrossRef]
- Holmes, G. The symptoms of acute cerebellar injuries due to gunshot injuries. Brain 1917, 40, 461–535. [Google Scholar] [CrossRef]
- Angel, R.W. Antagonist muscle activity during rapid arm movements: Central versus proprioceptive influences. J. Neurol. Neurosurg. Psychiatry 1977, 40, 683–686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffman, D.S.; Strick, P.L. Step-tracking movements of the wrist in humans. II. EMG analysis. J. Neurosci. 1990, 10, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Benecke, R.; Meinck, H.M.; Conrad, B. Rapid goal-directed elbow flexion movements: Limitations of the speed control system due to neural constraints. Exp. Brain Res. 1985, 59, 470–477. [Google Scholar] [CrossRef]
- Forget, R.; Lamarre, Y. Rapid elbow flexion in the absence of proprioceptive and cutaneous feedback. Hum. Neurobiol. 1987, 6, 27–37. [Google Scholar]
- Hore, J.; Flament, D. Evidence that a disordered servo-like mechanism contributes to tremor in movements during cerebellar dysfunction. J. Neurophysiol. 1986, 56, 123–136. [Google Scholar] [CrossRef]
- Elble, R.J.; Schieber, M.H.; Thach, W.T., Jr. Activity of muscle spindles, motor cortex and cerebellar nuclei during action tremor. Brain Res. 1984, 323, 330–334. [Google Scholar] [CrossRef]
- Hallett, M. Overview of human tremor physiology. Mov. Disord. 1998, 13, 434–438. [Google Scholar] [CrossRef]
- Massaquoi, S.G.; Slotine, J.J. The intermediate cerebellum may function as a wave-variable processor. Neurosci. Lett. 1996, 215, 60–64. [Google Scholar] [CrossRef]
- Casamento-Moran, A.; Yacoubi, B.; Wilkes, B.J.; Hess, C.W.; Foote, K.D.; Okun, M.S.; Wagle Shukla, A.; Vaillancourt, D.E.; Christoum, E.A. Quantitative separation of tremor and ataxia in essential tremor. Ann. Neurol. 2020. [Google Scholar] [CrossRef]
- Holmes, G. The cerebellum of man. Brain 1939, 62, 1–30. [Google Scholar] [CrossRef]
- Manni, E.; Petrosini, L. Luciani’s work on the cerebellum a century later. Trends Neurosci. 1997, 20, 112–116. [Google Scholar] [CrossRef]
- Thach, W.T.; Goodkin, H.P.; Keating, J.G. The cerebellum and the adaptive coordination of movement. Annu. Rev. Neurosci. 1992, 15, 403–442. [Google Scholar] [CrossRef] [PubMed]
- Bares, M.; Apps, R.; Avanzino, L.; Breska, A.; D’Angelo, E.; Filip, P.; Gerwig, M.; Ivry, R.B.; Lawrenson, C.L.; Louis, E.D.; et al. Consensus paper: Decoding the Contributions of the Cerebellum as a Time Machine. From Neurons to Clinical Applications. Cerebellum 2019, 18, 266–286. [Google Scholar] [CrossRef] [Green Version]
- Schlerf, J.E.; Spencer, R.M.; Zelaznik, H.N.; Ivry, R.B. Timing of rhythmic movements in patients with cerebellar degeneration. Cerebellum 2007, 6, 221–231. [Google Scholar] [CrossRef]
- Gerwig, M.; Hajjar, K.; Dimitrova, A.; Maschke, M.; Kolb, F.P.; Frings, M.; Thilmann, A.F.; Forsting, M.; Diener, H.C.; Timmannm, D. Timing of conditioned eyeblink responses is impaired in cerebellar patients. J. Neurosci. 2005, 25, 3919–3931. [Google Scholar] [CrossRef]
- Thach, W.T. Does the cerebellum initiate movement? Cerebellum 2014, 13, 139–150. [Google Scholar] [CrossRef]
- Babinski, J. De l’asynergie cérébelleuse. Rev. Neurol. 1899, 7, 806–816. [Google Scholar]
- Goodkin, H.P.; Thach, W.T. Cerebellar control of constrained and unconstrained movements. II. EMG and nuclear activity. J. Neurophysiol. 2003, 89, 896–908. [Google Scholar] [CrossRef] [Green Version]
- Panyakaew, P.; Cho, H.J.; Srivanitchapoom, P.; Popa, T.; Wu, T.; Hallett, M. Cerebellar brain inhibition in the target and surround muscles during voluntary tonic activation. Eur. J. Neurosci. 2016, 43, 1075–1081. [Google Scholar] [CrossRef] [Green Version]
- Grimaldi, G.; Oulad Ben Taib, N.; Manto, M.; Bodranghien, F. Marked reduction of cerebellar deficits in upper limbs following transcranial cerebello-cerebral DC stimulation: Tremor reduction and re-programming of the timing of antagonist commands. Front. Syst. Neurosci. 2014, 8, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]









© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cabaraux, P.; Gandini, J.; Kakei, S.; Manto, M.; Mitoma, H.; Tanaka, H. Dysmetria and Errors in Predictions: The Role of Internal Forward Model. Int. J. Mol. Sci. 2020, 21, 6900. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186900
Cabaraux P, Gandini J, Kakei S, Manto M, Mitoma H, Tanaka H. Dysmetria and Errors in Predictions: The Role of Internal Forward Model. International Journal of Molecular Sciences. 2020; 21(18):6900. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186900
Chicago/Turabian StyleCabaraux, Pierre, Jordi Gandini, Shinji Kakei, Mario Manto, Hiroshi Mitoma, and Hirokazu Tanaka. 2020. "Dysmetria and Errors in Predictions: The Role of Internal Forward Model" International Journal of Molecular Sciences 21, no. 18: 6900. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21186900