Identification of Ovine Serum miRNAs Following Bacterial Lipopolysaccharide Challenge

 ,
,
Abstract
:
1. Introduction
2. Results
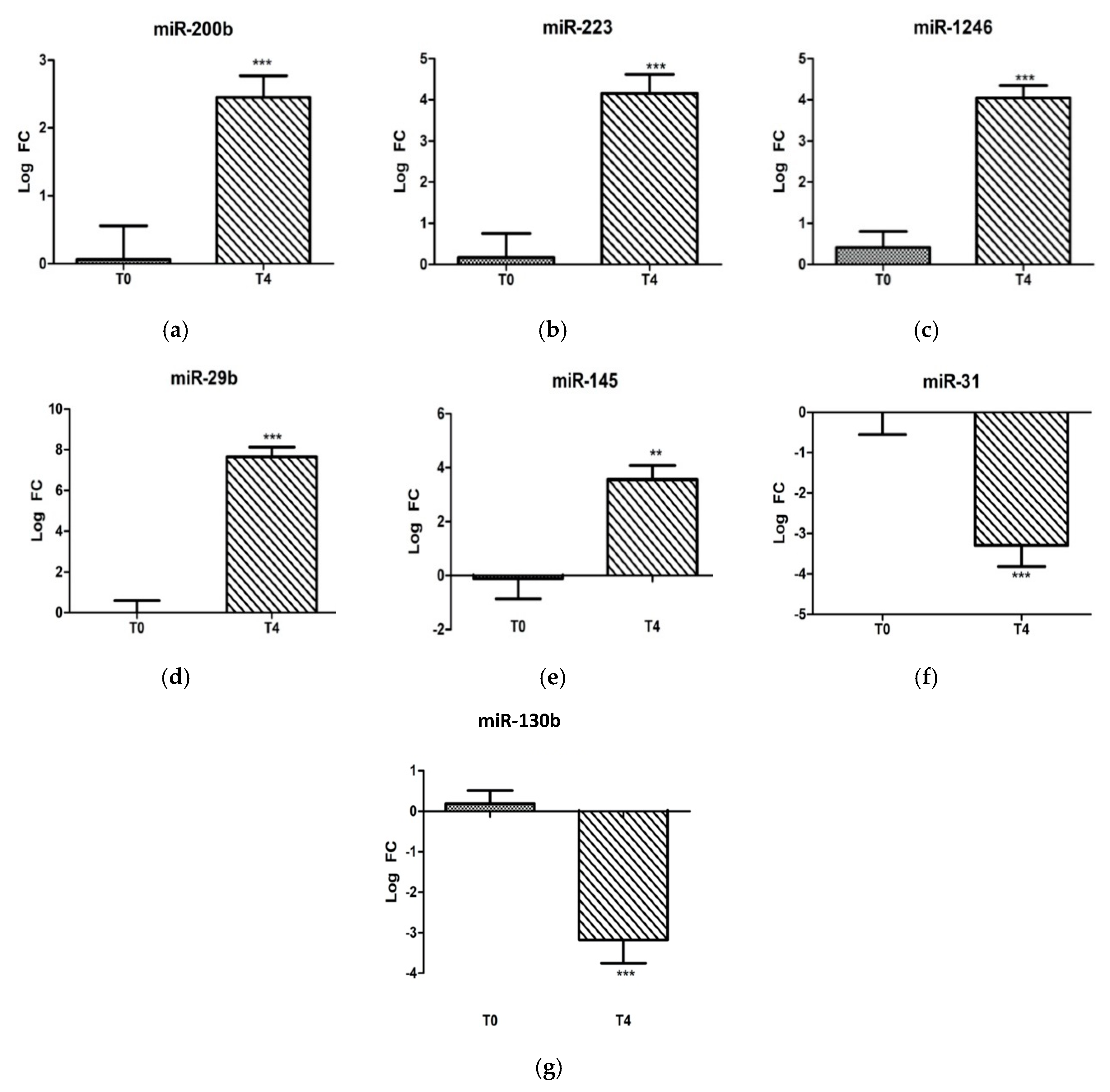
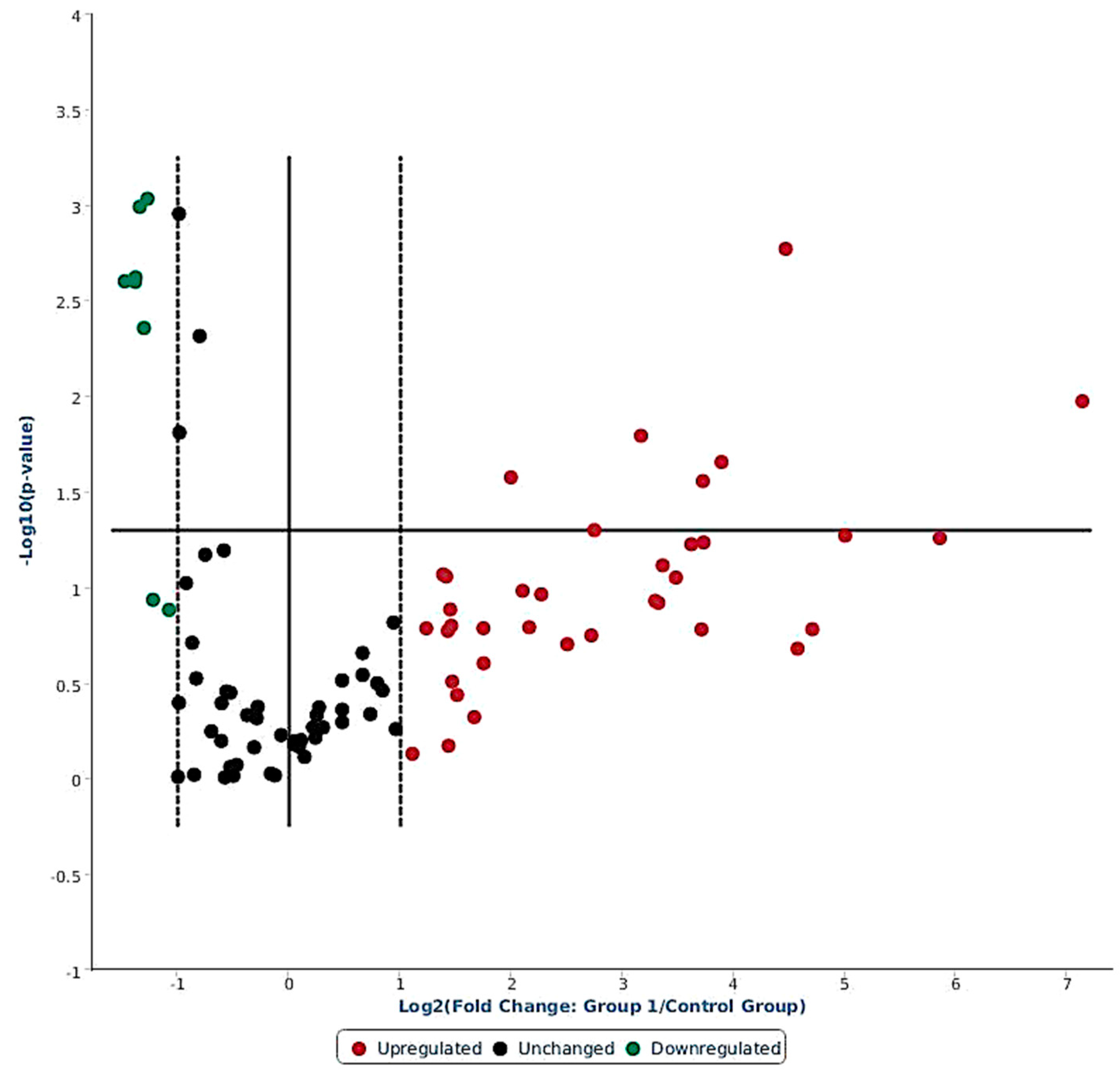
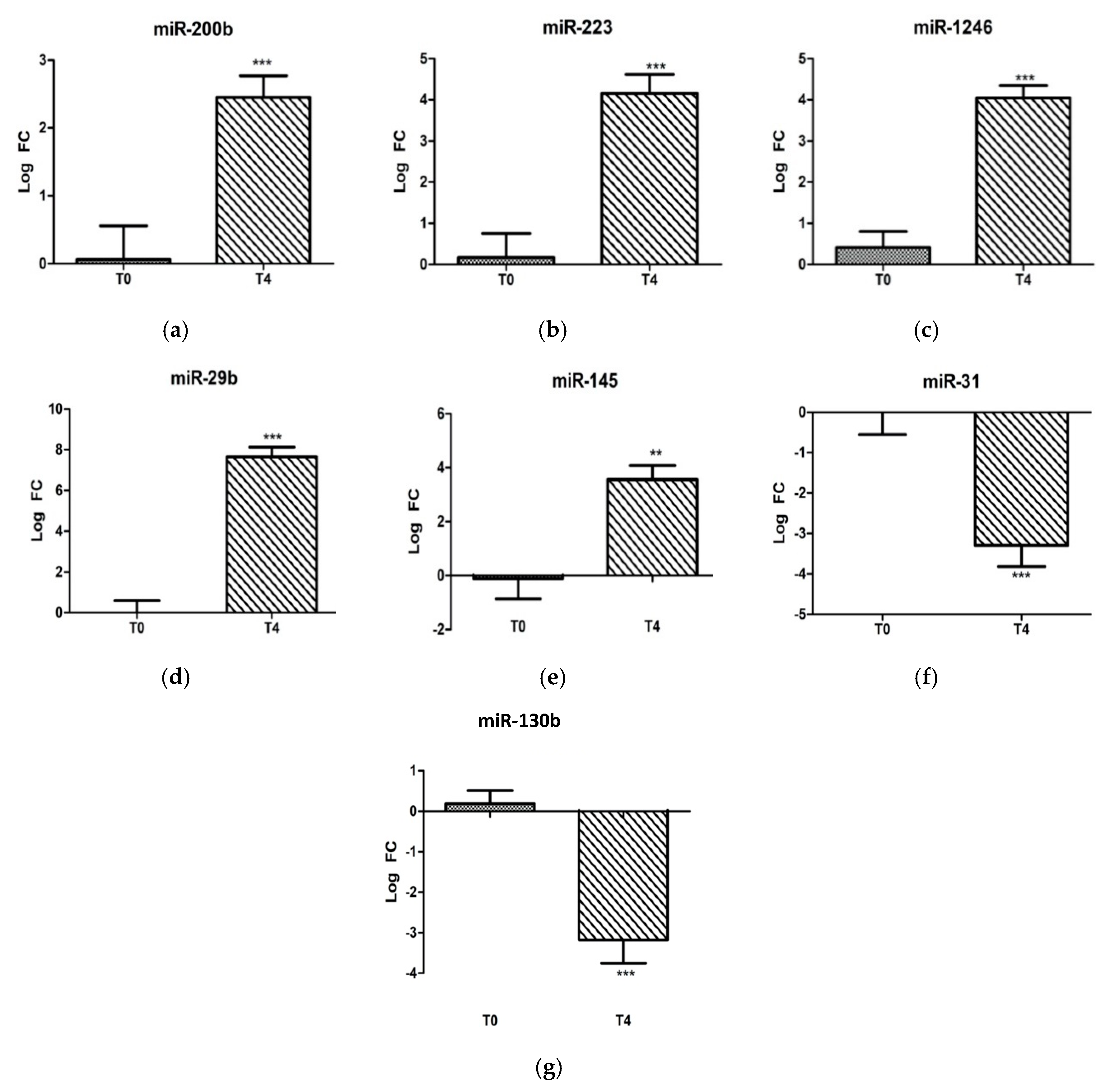
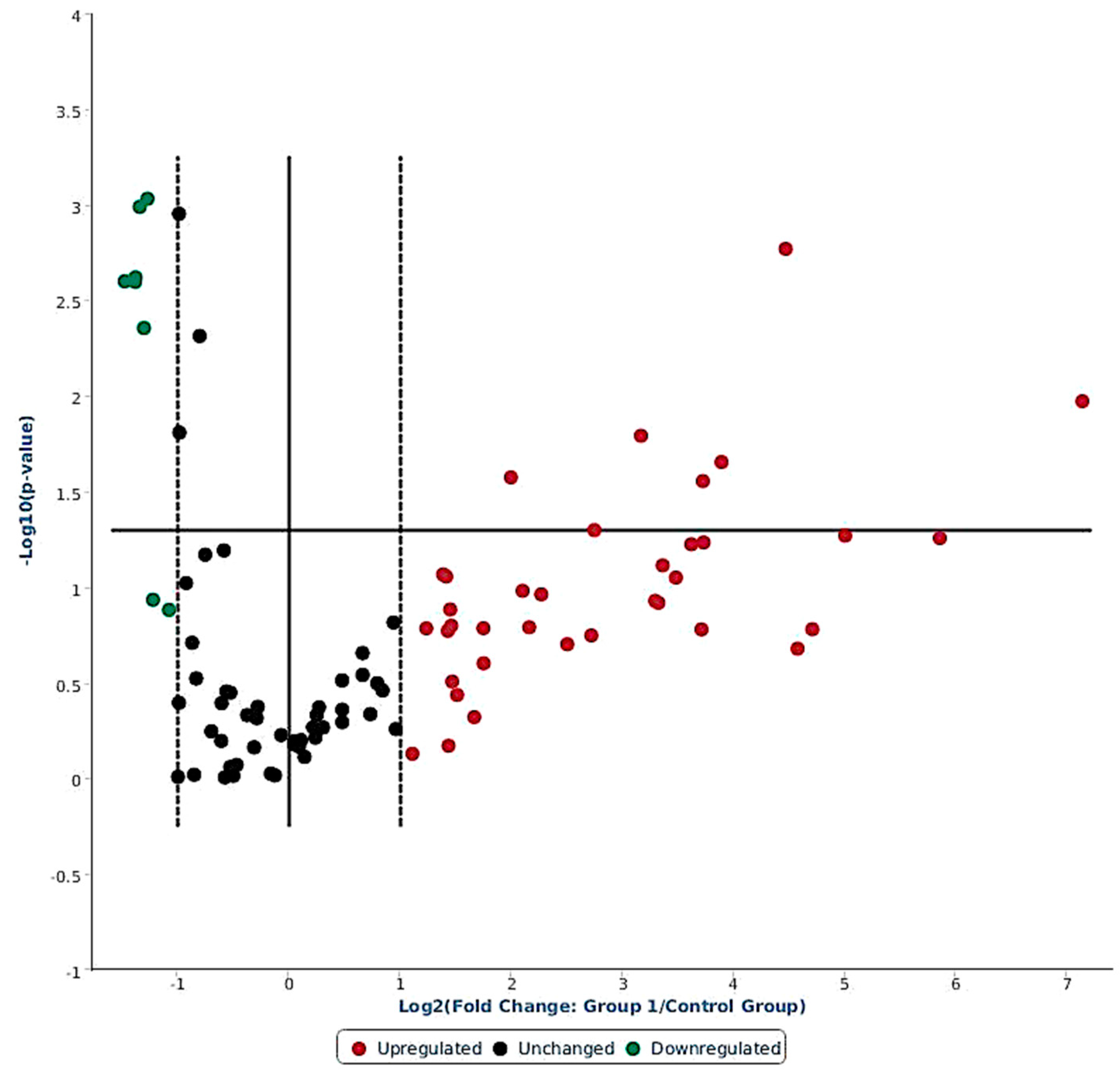
2.1. Ovine miRNAs Associated with LPS Stress Challenge
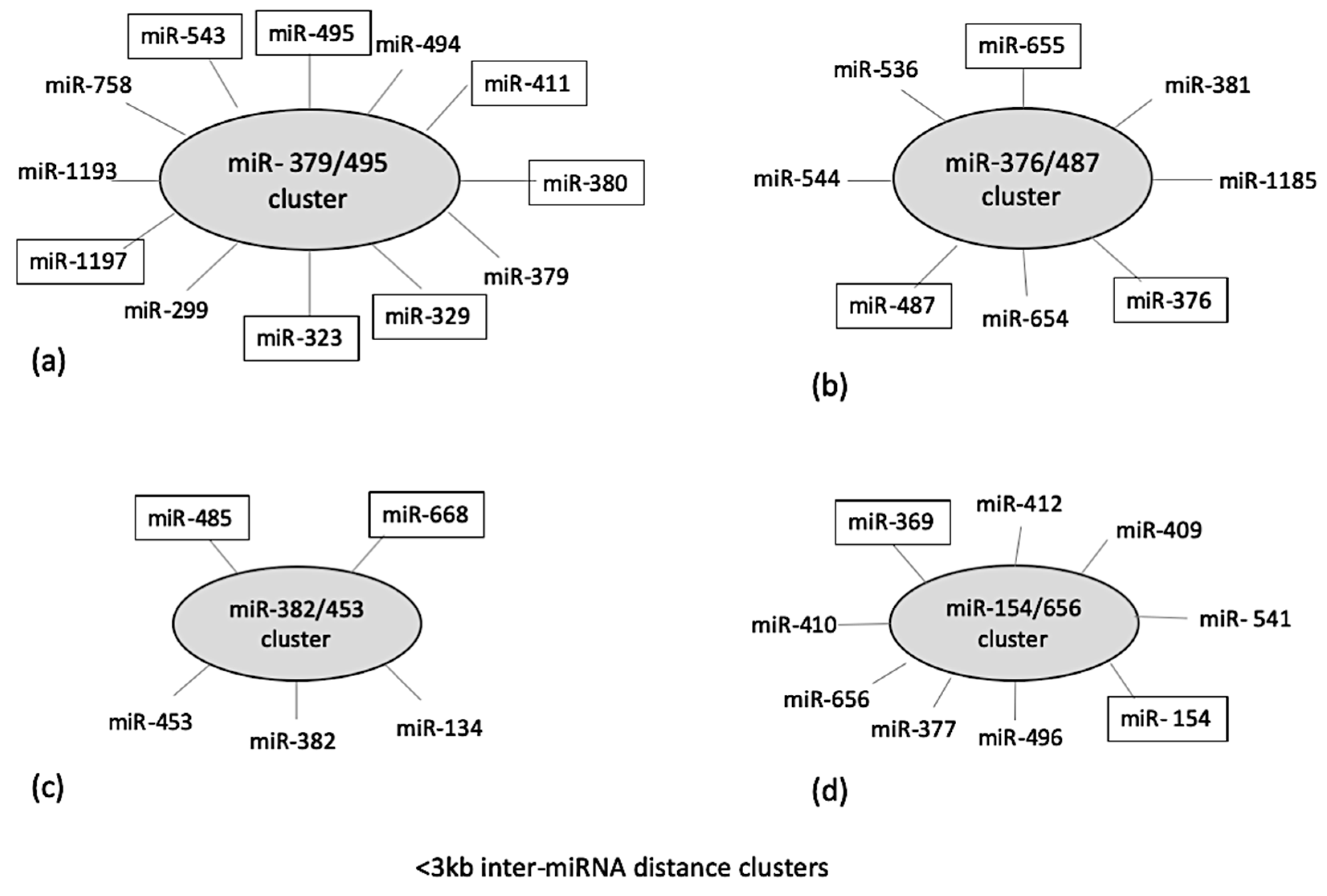
2.2. Genomic Localization and Cluster Analysis of Differentially Expressed (DE) miRNA
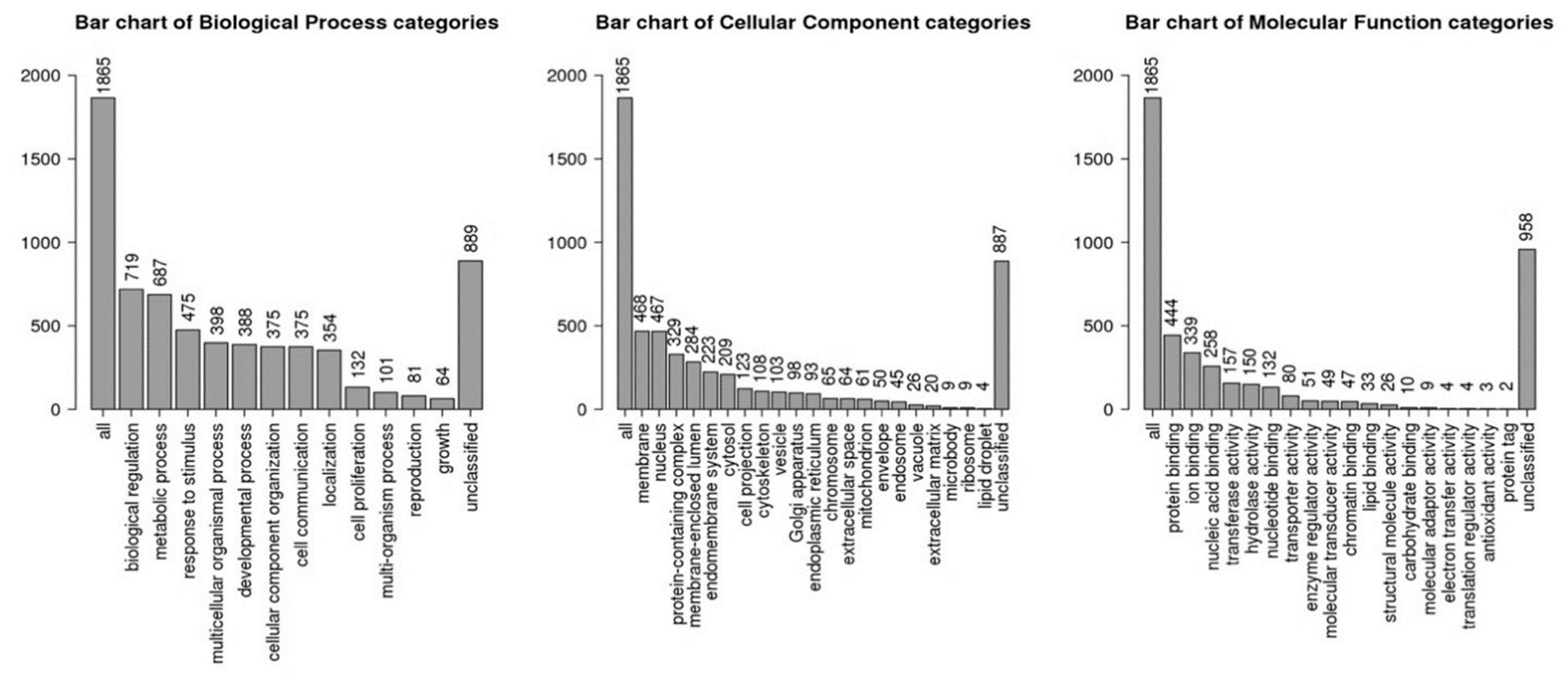
2.3. Gene Enrichment and Pathways Analysis
3. Discussion
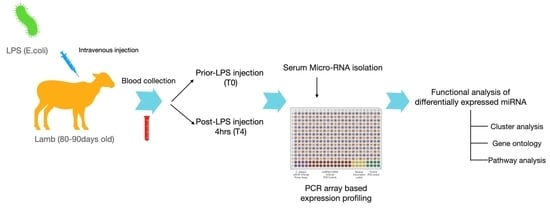
4. Materials and Methods
4.1. LPS Stress Challenge and Sample Collection
4.2. miRNA Isolation and cDNA Synthesis
4.3. miRNA Expression Analysis

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| miRNA | Forward Primer (5′-3′) | Annealing Temperature | References |
|---|---|---|---|
| miR-145a | GTCCAGTTTTCCCAGGAATCCCT | 60 °C | [58] |
| miR-130b | AGCAGGCAGTGCAATGATGA | 60 °C | |
| miR-145 | GTCCAGTTTTCCCAGGAATCC CT | 60 °C | |
| miR-223 | CCTGTCAGTTTGTCAAATACC CCA | 60 °C | |
| miR-1246 | GAATGGATTTTTGGAGCAGGA A | 60 °C | |
| miR-31 | GGAAGGCAAGATGCTGGCA | 60 °C | |
| miR-29b | GCGTAGCACCATTTGAAATC | 60 °C |
4.4. Target Gene Prediction and Pathway Analysis
4.5. Genomic Localization and Cluster Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| LPS | Lipopolysaccharide |
| IV | Intravenously |
| miR | MicroRNA |
| oar-mir | Ovine MicroRNA |
| DE | Differentially Expressed |
| FR | Fold regulation |
| MAPK | Mitogen-Activated Protein Kinase |
| PI3K | Phosphoinositide 3-kinases |
| FDR | False discovery rate |
References
- Herry, V.; Gitton, C.; Tabouret, G.; Répérant, M.; Forge, L.; Tasca, C.; Gilbert, F.B.; Guitton, E.; Barc, C.; Staub, C. Local immunization impacts the response of dairy cows to Escherichia coli mastitis. Sci. Rep. 2017, 7, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Abaker, J.A.; Xu, T.L.; Jin, D.; Chang, G.J.; Zhang, K.; Shen, X.Z. Lipopolysaccharide derived from the digestive tract provokes oxidative stress in the liver of dairy cows fed a high-grain diet. J. Dairy Sci. 2017, 100, 666–678. [Google Scholar] [CrossRef] [PubMed]
- Vargas, N.; Marino, F. Heat stress, gastrointestinal permeability and interleukin-6 signaling—Implications for exercise performance and fatigue. Temperature 2016, 3, 240–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denk, S.; Perl, M.; Huber-Lang, M. Damage-and pathogen-associated molecular patterns and alarmins: Keys to sepsis? Eur. Surg. Res. 2012, 48, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Kabaroff, L.C.; Rodriguez, A.; Quinton, M.; Boermans, H.; Karrow, N.A. Assessment of the ovine acute phase response and hepatic gene expression in response to Escherichia coli endotoxin. Vet. Immunol. Immunopathol. 2006, 113, 113–124. [Google Scholar] [CrossRef]
- Naylor, D.; Sharma, A.; Li, Z.; Monteith, G.; Sullivan, T.; Canovas, A.; Mallard, B.A.; Baes, C.; Karrow, N.A. Characterizing ovine serum stress biomarkers during endotoxemia. J. Dairy Sci. 2020, 103, 5501–5508. [Google Scholar] [CrossRef]
- Pant, S.D.; You, Q.; Schenkel, L.C.; Vander Voort, G.; Schenkel, F.S.; Wilton, J.; Cain, L.; Karrow, N.A. A genome-wide association study to identify chromosomal regions influencing ovine cortisol response. Livest. Sci. 2016, 187, 40–47. [Google Scholar] [CrossRef]
- Begg, D.J.; Purdie, A.C.; de Silva, K.; Dhand, N.K.; Plain, K.M.; Whittington, R.J. Variation in susceptibility of different breeds of sheep to Mycobacterium avium subspecies paratuberculosis following experimental inoculation. Vet. Res. 2017, 48, 36. [Google Scholar] [CrossRef] [Green Version]
- Bowdridge, S.A.; Zajac, A.M.; Notter, D.R. St. Croix sheep produce a rapid and greater cellular immune response contributing to reduced establishment of Haemonchus contortus. Vet. Parasitol. 2015, 208, 204–210. [Google Scholar] [CrossRef]
- Hadfield, J.M.; Bowdridge, E.C.; Holásková, I.; Elsasser, T.H.; Dailey, R.A. Breed-specific differences in the immune response to lipopolysaccharide in ewes. J. Anim. Sci. 2018, 96, 4220–4228. [Google Scholar] [CrossRef] [PubMed]
- Leung, A.K.; Sharp, P.A. MicroRNA functions in stress responses. Mol. Cell 2010, 40, 205–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, X.; Ranade, K.; Talker, R.; Jallal, B.; Shen, N.; Yao, Y. MicroRNA-mediated regulation of innate immune response in rheumatic diseases. Arthritis Res. Ther. 2013, 15, 210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, P.; Cho, S.K.; Thulstrup, P.W.; Bjerrum, M.J.; Lee, P.H.; Kang, J.-H.; Bhang, Y.-J.; Yang, S.W. MicroRNA biomarkers in neurodegenerative diseases and emerging nanosensors technology. J. Mov. Disord. 2017, 10, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paul, P.; Chakraborty, A.; Sarkar, D.; Langthasa, M.; Rahman, M.; Bari, M.; Singha, R.S.; Malakar, A.K.; Chakraborty, S. Interplay between MiRNAs and human diseases. J. Cell. Physiol. 2018, 233, 2007–2018. [Google Scholar] [CrossRef]
- Witwer, K.W. Circulating MicroRNA biomarker studies: Pitfalls and potential solutions. Clin. Chem. 2015, 61, 56–63. [Google Scholar] [CrossRef]
- Zhou, M.; Yu, G.; Yang, X.; Zhu, C.; Zhang, Z.; Zhan, X. Circulating MicroRNAs as biomarkers for the early diagnosis of childhood tuberculosis infection. Mol. Med. Rep. 2016, 13, 4620–4626. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Shen, X.; Xie, L.; Chu, M.; Ma, Y. MicroRNA-181b regulates endotoxin tolerance by targeting IL-6 in macrophage RAW264.7 cells. J. Inflamm. 2015, 12, 18. [Google Scholar] [CrossRef] [Green Version]
- Kabekkodu, S.P.; Shukla, V.; Varghese, V.K.; D’Souza, J.; Chakrabarty, S.; Satyamoorthy, K. Clustered MiRNAs and their role in biological functions and diseases. Biol. Rev. 2018, 93, 1955–1986. [Google Scholar] [CrossRef]
- Ross, M.; Atalla, H.; Karrow, N.A.; Mallard, B. Bioactivity of bovine colostrum and milk exosomes from high, average, and low immune responders on human intestinal epithelial cells. J. Dairy Sci. 2020. (Accepted). [Google Scholar]
- He, X.; Jing, Z.; Cheng, G. MicroRNAs: New regulators of toll-like receptor signalling pathways. BioMed Res. Int. 2014, 2014, 945169. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, C.-H.; Rau, C.-S.; Jeng, J.C.; Chen, Y.-C.; Lu, T.-H.; Wu, C.-J.; Wu, Y.-C.; Tzeng, S.-L.; Yang, J.C.-S. Whole blood-derived MicroRNA signatures in mice exposed to lipopolysaccharides. J. Biomed. Sci. 2012, 19, 69. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Yang, L.; Zhao, Z.; Wang, J.; Zhang, X. Signature MiRNAs involved in the innate immunity of invertebrates. PLoS ONE 2012, 7, e39015. [Google Scholar] [CrossRef] [PubMed]
- Vasilescu, C.; Dragomir, M.; Tanase, M.; Giza, D.; Purnichescu-Purtan, R.; Chen, M.; Yeung, S.-C. J.; Calin, G.A. Circulating MiRNAs in sepsis—A network under attack: An in-silico prediction of the potential existence of MiRNA sponges in sepsis. PLoS ONE 2017, 12, e0183334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dumache, R.; Rogobete, A.F.; Bedreag, O.H.; Sarandan, M.; Cradigati, A.C.; Papurica, M.; Dumbuleu, C.M.; Nartita, R.; Sandesc, D. Use of MiRNAs as biomarkers in sepsis. Anal. Cell. Pathol. 2015, 2015, 186716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawless, N.; Reinhardt, T.A.; Bryan, K.; Baker, M.; Pesch, B.; Zimmerman, D.; Zuelke, K.; Sonstegard, T.; O’Farrelly, C.; Lippolis, J.D. MicroRNA regulation of bovine monocyte inflammatory and metabolic networks in an in vivo infection model. G3 Genes Genomes Genet. 2014, 4, 957–971. [Google Scholar] [CrossRef] [Green Version]
- Luoreng, Z.-M.; Wang, X.-P.; Mei, C.-G.; Zan, L.-S. Comparison of MicroRNA profiles between bovine mammary glands infected with Staphylococcus aureus and Escherichia coli. Int. J. Biol. Sci. 2018, 14, 87. [Google Scholar] [CrossRef]
- Ibrahim, S.; Szóstek-Mioduchowska, A.; Skarzynski, D. Expression profiling of selected MiRNAs in equine endometrium in response to LPS challenge in vitro: A new understanding of the inflammatory immune response. Vet. Immunol. Immunopathol. 2019, 209, 37–44. [Google Scholar] [CrossRef]
- Dorhoi, A.; Iannaccone, M.; Farinacci, M.; Faé, K.C.; Schreiber, J.; Moura-Alves, P.; Nouailles, G.; Mollenkopf, H.-J.; Oberbeck-Müller, D.; Jörg, S. MicroRNA-223 controls susceptibility to tuberculosis by regulating lung neutrophil recruitment. J. Clin. Investig. 2013, 123, 4836–4848. [Google Scholar] [CrossRef] [Green Version]
- Benz, F.; Roy, S.; Trautwein, C.; Roderburg, C.; Luedde, T. Circulating MicroRNAs as biomarkers for sepsis. Int. J. Mol. Sci. 2016, 17, 78. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Yu, M.; Yu, G.; Bian, J.; Deng, X.; Wan, X.; Zhu, K. Serum MiR-146a and MiR-223 as potential new biomarkers for sepsis. Biochem. Biophys. Res. Commun. 2010, 394, 184–188. [Google Scholar] [CrossRef]
- Fang, Y.; Gao, F.; Hao, J.; Liu, Z. MicroRNA-1246 mediates lipopolysaccharide-induced pulmonary endothelial cell apoptosis and acute lung injury by targeting angiotensin-converting enzyme 2. Am. J. Transl. Res. 2017, 9, 1287. [Google Scholar] [PubMed]
- Bott, A.; Erdem, N.; Lerrer, S.; Hotz-Wagenblatt, A.; Breunig, C.; Abnaof, K.; Wörner, A.; Wilhelm, H.; Münstermann, E.; Ben-Baruch, A. MiRNA-1246 induces pro-inflammatory responses in mesenchymal stem/stromal cells by regulating PKA and PP2A. Oncotarget 2017, 8, 43897. [Google Scholar] [CrossRef] [PubMed]
- Singh, J.; Mukhopadhyay, C.S.; Kaur, S.; Malhotra, P.; Sethi, R.S.; Choudhary, R.K. Identification of the MicroRNA repertoire in TLR-ligand challenged bubaline PBMCs as a model of bacterial and viral infection. PLoS ONE 2016, 11, e0156598. [Google Scholar] [CrossRef] [PubMed]
- Sharbati, J.; Lewin, A.; Kutz-Lohroff, B.; Kamal, E.; Einspanier, R.; Sharbati, S. Integrated MicroRNA-MRNA-analysis of human monocyte derived macrophages upon Mycobacterium avium subsp. hominissuis infection. PLoS ONE 2011, 6, e20258. [Google Scholar] [CrossRef] [PubMed]
- Lucotti, S.; Rainaldi, G.; Evangelista, M.; Rizzo, M. Fludarabine treatment favors the retention of MiR-485-3p by prostate cancer cells: Implications for survival. Mol. Cancer 2013, 12, 52. [Google Scholar] [CrossRef] [Green Version]
- Kálmán, S.; Garbett, K.A.; Vereczkei, A.; Shelton, R.C.; Korade, Ž.; Mirnics, K. Metabolic stress-induced MicroRNA and MRNA expression profiles of human fibroblasts. Exp. Cell Res. 2014, 320, 343–353. [Google Scholar] [CrossRef] [Green Version]
- Solé, C.; Domingo, S.; Ferrer, B.; Moliné, T.; Ordi-Ros, J.; Cortés-Hernández, J. MicroRNA expression profiling identifies MiR-31 and MiR-485-3p as regulators in the pathogenesis of discoid cutaneous lupus. J. Investig. Dermatol. 2019, 139, 51–61. [Google Scholar] [CrossRef] [Green Version]
- Majumder, M.; Dunn, L.; Liu, L.; Hasan, A.; Vincent, K.; Brackstone, M.; Hess, D.; Lala, P.K. COX-2 induces oncogenic Micro RNA MiR655 in human breast cancer. Sci. Rep. 2018, 8, 327. [Google Scholar] [CrossRef] [Green Version]
- Naqvi, A.R.; Zhong, S.; Dang, H.; Fordham, J.B.; Nares, S.; Khan, A. Expression profiling of LPS responsive MiRNA in primary human macrophages. J. Microb. Biochem. Technol. 2016, 8, 136–143. [Google Scholar] [CrossRef]
- Sun, F.; Li, S.-G.; Zhang, H.-W.; Hua, F.-W.; Sun, G.-Z.; Huang, Z. MiRNA-411 attenuates inflammatory damage and apoptosis following spinal cord injury. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 491–498. [Google Scholar] [CrossRef]
- Xiang, Y.; Eyers, F.; Herbert, C.; Tay, H.L.; Foster, P.S.; Yang, M. MicroRNA-487b is a negative regulator of macrophage activation by targeting IL-33 production. J. Immunol. Baltim. Md 1950 2016, 196, 3421–3428. [Google Scholar] [CrossRef] [PubMed]
- Suárez, Y.; Wang, C.; Manes, T.D.; Pober, J.S. Cutting edge: TNF-induced MicroRNAs regulate TNF-induced expression of E-selectin and intercellular adhesion molecule-1 on human endothelial cells: Feedback control of inflammation. J. Immunol. Baltim. Md 1950 2010, 184, 21–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, N.; Meisgen, F.; Butler, L.M.; Han, G.; Wang, X.-J.; Söderberg-Nauclér, C.; Ståhle, M.; Pivarcsi, A.; Sonkoly, E. MicroRNA-31 is overexpressed in psoriasis and modulates inflammatory cytokine and chemokine production in keratinocytes via targeting serine/threonine kinase 40. J. Immunol. 2013, 190, 678–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magni, S.; Comani, G.B.; Elli, L.; Vanessi, S.; Ballarini, E.; Nicolini, G.; Rusconi, M.; Castoldi, M.; Meneveri, R.; Muckenthaler, M.U.; et al. MiRNAs affect the expression of innate and adaptive immunity proteins in celiac disease. Off. J. Am. Coll. Gastroenterol. ACG 2014, 109, 1662–1674. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Liu, P.; Wu, H.; Cui, P.; Li, Y.; Liu, Y.; Liu, Z.; Gou, S. MicroRNA-323-3p inhibits cell invasion and metastasis in pancreatic ductal adenocarcinoma via direct suppression of SMAD2 and SMAD3. Oncotarget 2016, 7, 14912–14924. [Google Scholar] [CrossRef] [Green Version]
- Travis, M.A.; Sheppard, D. TGF-β activation and function in immunity. Annu. Rev. Immunol. 2014, 32, 51–82. [Google Scholar] [CrossRef] [Green Version]
- Swarbrick, A.; Woods, S.L.; Shaw, A.; Balakrishnan, A.; Phua, Y.; Nguyen, A.; Chanthery, Y.; Lim, L.; Ashton, L.J.; Judson, R.L.; et al. MiR-380-5p represses P53 to control cellular survival and is associated with poor outcome in MYCN-amplified neuroblastoma. Nat. Med. 2010, 16, 1134–1140. [Google Scholar] [CrossRef]
- Zhang, S.; Zhao, F.; Wei, C.; Sheng, X.; Ren, H.; Xu, L.; Lu, J.; Liu, J.; Zhang, L.; Du, L. Identification and characterization of the MiRNA transcriptome of Ovis aries. PLoS ONE 2013, 8, e58905. [Google Scholar] [CrossRef] [Green Version]
- Hou, L.; Ji, Z.; Wang, G.; Wang, J.; Chao, T.; Wang, J. Identification and characterization of MicroRNAs in the intestinal tissues of sheep (Ovis aries). PLoS ONE 2018, 13, e0193371. [Google Scholar] [CrossRef] [Green Version]
- Laganà, A.; Veneziano, D.; Spata, T.; Tang, R.; Zhu, H.; Mohler, P.J.; Kilic, A. Identification of general and heart-specific MiRNAs in sheep (Ovis aries). PLoS ONE 2015, 10, e0143313. [Google Scholar] [CrossRef] [Green Version]
- Saini, H.K.; Griffiths-Jones, S.; Enright, A.J. Genomic analysis of human MicroRNA transcripts. Proc. Natl. Acad. Sci. USA 2007, 104, 17719–17724. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, K.; Seike, M.; Takeuchi, S.; Soeno, C.; Miyanaga, A.; Noro, R.; Minegishi, Y.; Kubota, K.; Gemma, A. MiR-379/411 cluster regulates IL-18 and contributes to drug resistance in malignant pleural mesothelioma. Oncol. Rep. 2014, 32, 2365–2372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Mei, C.; Su, X.; Wang, H.; Yang, W.; Zan, L. MEF2A Regulates the MEG3-DIO3 MiRNA mega cluster-targeted PP2A signaling in bovine skeletal myoblast differentiation. Int. J. Mol. Sci. 2019, 20, 2748. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Angelo, B.; Astarita, C.; Boffo, S.; Massaro-Giordano, M.; Antonella Ianuzzi, C.; Caporaso, A.; Macaluso, M.; Giordano, A. LPS-induced inflammatory response triggers cell cycle reactivation in murine neuronal cells through retinoblastoma proteins induction. Cell Cycle Georget. Tex 2017, 16, 2330–2336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ono, Y.; Sakamoto, K. Lipopolysaccharide inhibits myogenic differentiation of C2C12 myoblasts through the toll-like receptor 4-nuclear factor-ΚB signaling pathway and myoblast-derived tumor necrosis factor-α. PLoS ONE 2017, 12, e0182040. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT–PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- D’haene, B.; Mestdagh, P.; Hellemans, J.; Vandesompele, J. MiRNA expression profiling: From reference genes to global mean normalization. In Next-Generation MicroRNA Expression Profiling Technology; Springer: Berlin/Heidelberg, Germany, 2012; pp. 261–272. [Google Scholar]
- Li, Q.; Yang, C.; Du, J.; Zhang, B.; He, Y.; Hu, Q.; Li, M.; Zhang, Y.; Wang, C.; Zhong, J. Characterization of MiRNA profiles in the mammary tissue of dairy cattle in response to heat stress. BMC Genomics 2018, 19, 975. [Google Scholar] [CrossRef] [Green Version]
- Chan, W.C.; Lin, W.C. MetaMirClust: Discovery and exploration of evolutionarily conserved miRNA clusters. In Microarray Data Analysis; Humana Press: New York, NY, USA, 2015; pp. 75–89. [Google Scholar]

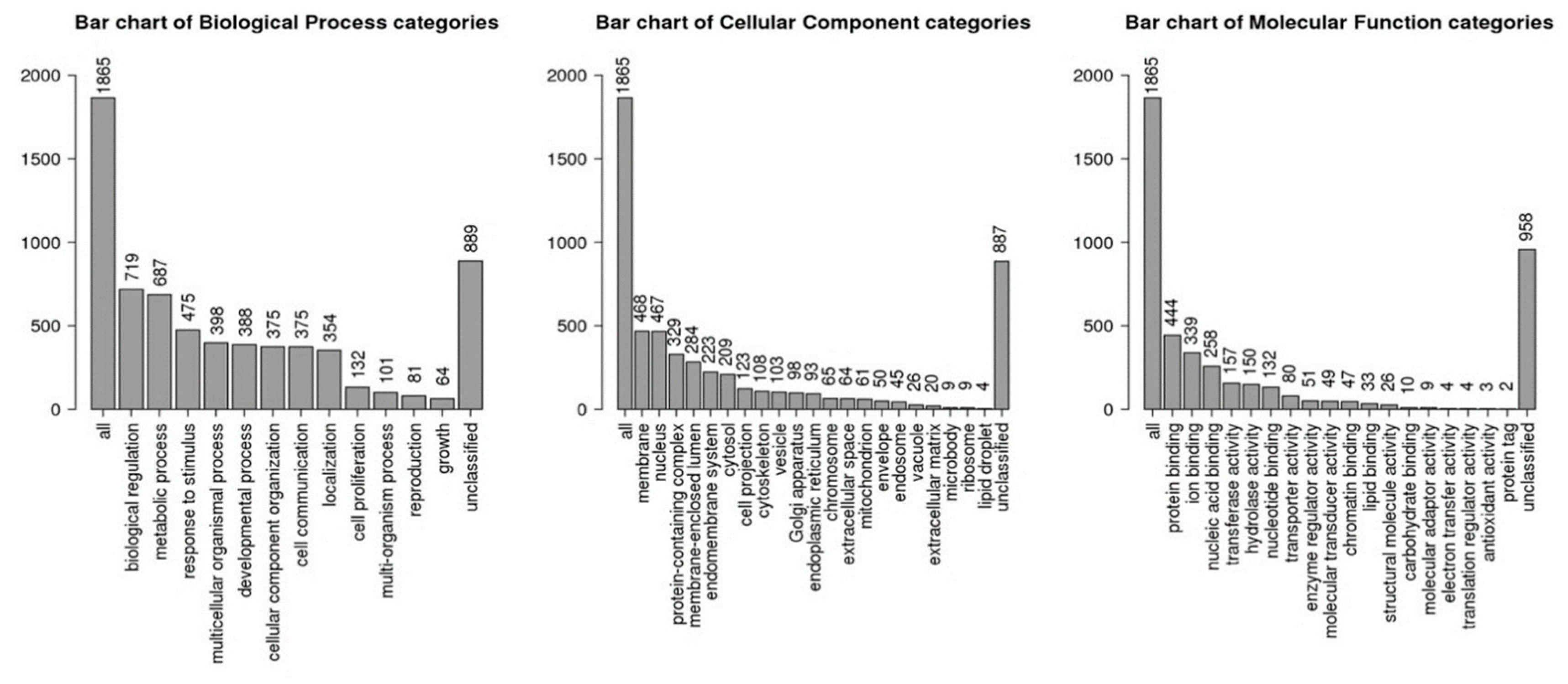
| Mature ID | Fold-Regulation | p-Value | Mature ID | Fold-Regulation | p-Value |
|---|---|---|---|---|---|
| Upregulated | Downregulated | ||||
| oar-miR-369-3p | 140.89 | 0.011 * | oar-miR-154b-3p | −2.41 | <0.001 * |
| oar-miR-495-3p | 57.94 | 0.055 * | oar-miR-323b | −2.6 | 0.002 * |
| oar-miR-376a-3p | 32.09 | 0.050 * | oar-miR-323c | −2.47 | 0.004 * |
| oar-miR-543-3p | 22.15 | 0.002 * | oar-miR-380-5p | −2.78 | 0.002 * |
| oar-miR-668-3p | 14.85 | 0.022 * | oar-miR-665-5p | −2.54 | 0.001 * |
| oar-miR-329a-3p | 13.22 | 0.028 * | oar-miR-1197-5p | −2.61 | 0.002 * |
| oar-miR-487b-3p | 12.29 | 0.060 | |||
| oar-miR-485-3p | 10.29 | 0.077 | |||
| oar-miR-655-3p | 8.99 | 0.016 * | |||
| oar-miR-411a-5p | 6.72 | 0.050 * | |||
| oar-miR-154a-3p | 3.99 | 0.027 * | |||
| Gene Set | Description | FDR |
|---|---|---|
| Biological Processes | ||
| GO:0010468 | Regulation of gene expression | 1.48 × 10−12 |
| GO:0051252 | Regulation of RNA metabolic process | 1.68 × 10−11 |
| GO:0010556 | Regulation of macromolecule biosynthetic process | 1.68 × 10−11 |
| GO:0019219 | Regulation of nucleobase-containing compound metabolic process | 1.93 × 10−11 |
| GO:0009653 | Anatomical structure morphogenesis | 8.08 × 10−11 |
| GO:2000112 | Regulation of cellular macromolecule biosynthetic process | 8.08 × 10−11 |
| GO:0009889 | Regulation of biosynthetic process | 9.23 × 10−11 |
| GO:0031326 | Regulation of cellular biosynthetic process | 1.06 × 10−10 |
| GO:0072359 | Circulatory system development | 1.16 × 10−10 |
| GO:0051254 | Positive regulation of RNA metabolic process | 1.31 × 10−10 |
| Cellular Component | ||
| GO:0031981 | Nuclear lumen | 0.00 |
| GO:0005654 | Nucleoplasm | 0.00 |
| GO:0097458 | Neuron part | 2.47 × 10−7 |
| GO:0044459 | Plasma membrane part | 8.69 × 10−7 |
| GO:0005829 | Cytosol | 2.78 × 10−6 |
| GO:0120025 | Plasma membrane bounded cell projection | 4.86 × 10−6 |
| GO:0042995 | Cell projection | 4.86 × 10−6 |
| GO:0044456 | Synapse part | 6.34 × 10−6 |
| GO:0098978 | Glutamatergic synapse | 1.23 × 10−5 |
| GO:0036477 | Somatodendritic compartment | 1.38 × 10-5 |
| Molecular Functions | ||
| GO:0140110 | Transcription regulator activity | 1.57 × 10−8 |
| GO:0003700 | DNA-binding transcription factor activity | 1.64 × 10−8 |
| GO:0000981 | DNA-binding transcription factor activity, RNA polymerase II-specific | 1.64 × 10−8 |
| GO:0000977 | RNA polymerase II regulatory region sequence-specific DNA binding | 2.41 × 10−8 |
| GO:0043565 | Sequence-specific DNA binding | 2.41 × 10−8 |
| GO:0001012 | RNA polymerase II regulatory region DNA binding | 2.41 × 10−8 |
| GO:0000976 | Transcription regulatory region sequence-specific DNA binding | 3.16 × 10−8 |
| GO:0001067 | Regulatory region nucleic acid binding | 3.16 × 10−8 |
| GO:0044212 | Transcription regulatory region DNA binding | 1.57 × 10−8 |
| GO:0140110 | Transcription regulator activity | 1.64 × 10−8 |
| S.No | #Term | Corrected p-Value (FDR) | Database |
|---|---|---|---|
| 1 | PI3K-Akt signaling pathway | 9.99 × 10−13 | KEGG Pathway |
| 2 | Immune System | 1.41 × 10−12 | Reactome |
| 3 | Pathways in cancer | 3.50 × 10−11 | KEGG Pathway |
| 4 | MAPK signaling pathway | 1.02 × 10−10 | KEGG Pathway |
| 5 | Focal adhesion | 1.87 × 10−10 | KEGG Pathway |
| 6 | Ras signaling pathway | 8.38 × 10−10 | KEGG Pathway |
| 7 | FoxO signaling pathway | 8.83 × 10−8 | KEGG Pathway |
| 8 | Rap1 signaling pathway | 1.13 × 10−7 | KEGG Pathway |
| 9 | Post-translational protein modification | 1.25 × 10−7 | Reactome |
| 10 | Signal Transduction | 1.92 × 10−7 | Reactome |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sharma, A.; Shandilya, U.K.; Sullivan, T.; Naylor, D.; Canovas, A.; Mallard, B.A.; Karrow, N.A. Identification of Ovine Serum miRNAs Following Bacterial Lipopolysaccharide Challenge. Int. J. Mol. Sci. 2020, 21, 7920. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217920
Sharma A, Shandilya UK, Sullivan T, Naylor D, Canovas A, Mallard BA, Karrow NA. Identification of Ovine Serum miRNAs Following Bacterial Lipopolysaccharide Challenge. International Journal of Molecular Sciences. 2020; 21(21):7920. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217920
Chicago/Turabian StyleSharma, Ankita, Umesh K. Shandilya, Tianna Sullivan, Danielle Naylor, Angela Canovas, Bonnie A. Mallard, and Niel A. Karrow. 2020. "Identification of Ovine Serum miRNAs Following Bacterial Lipopolysaccharide Challenge" International Journal of Molecular Sciences 21, no. 21: 7920. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21217920