Desmin Interacts Directly with Mitochondria

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
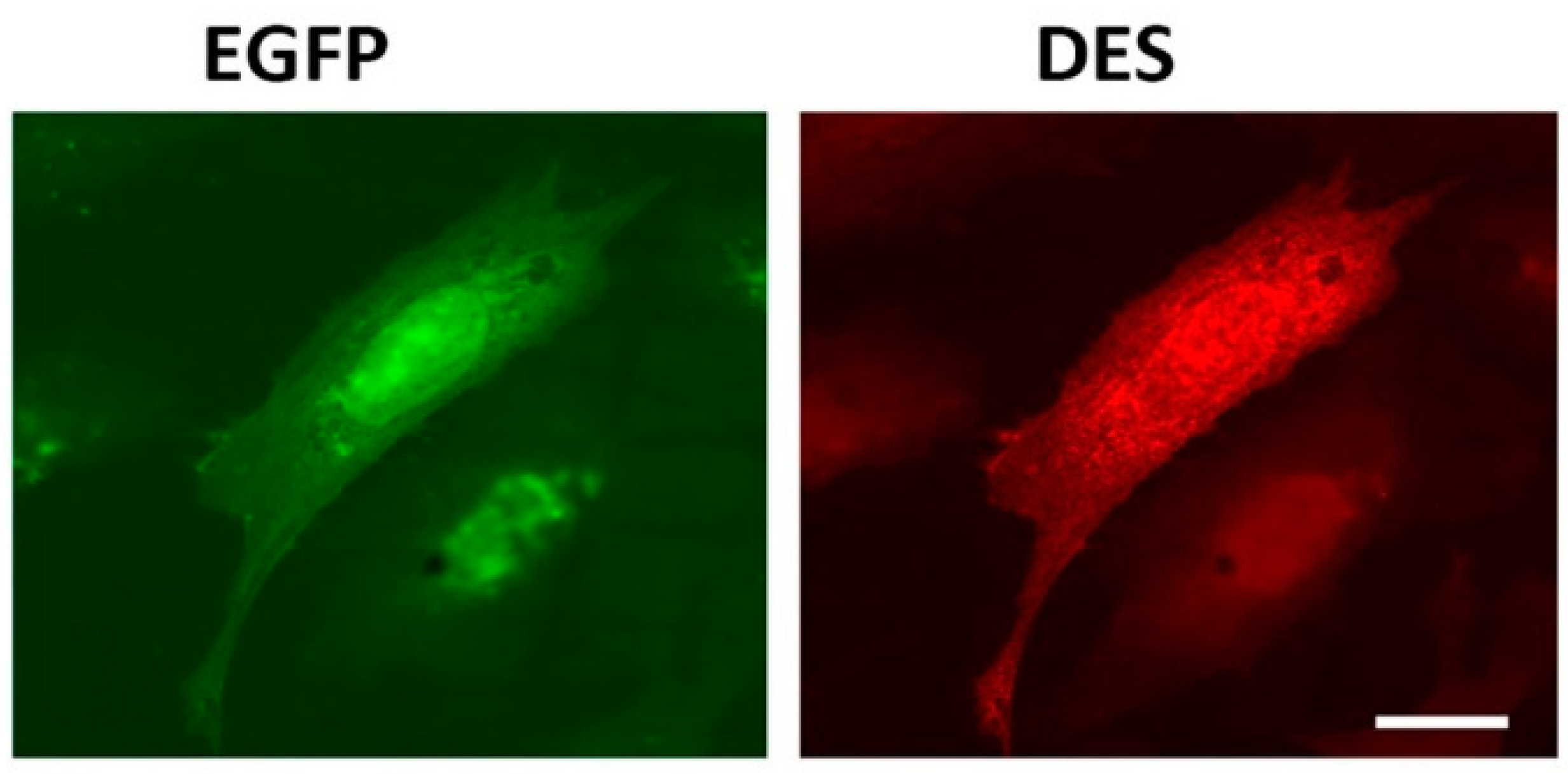
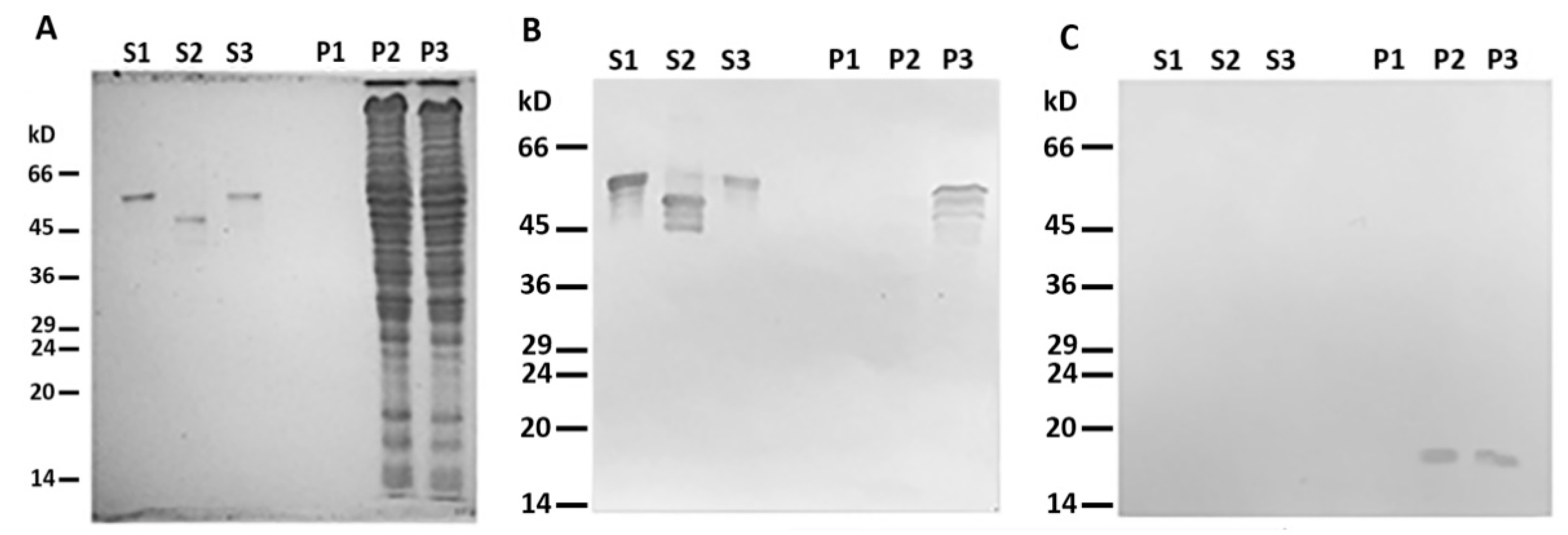
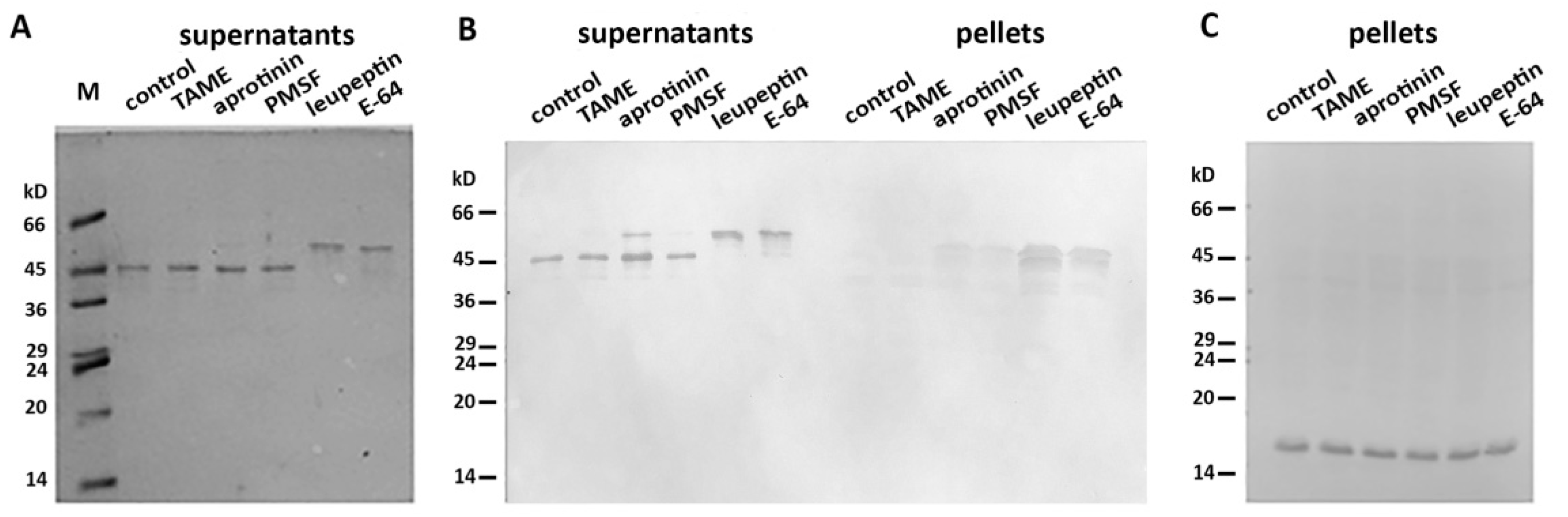
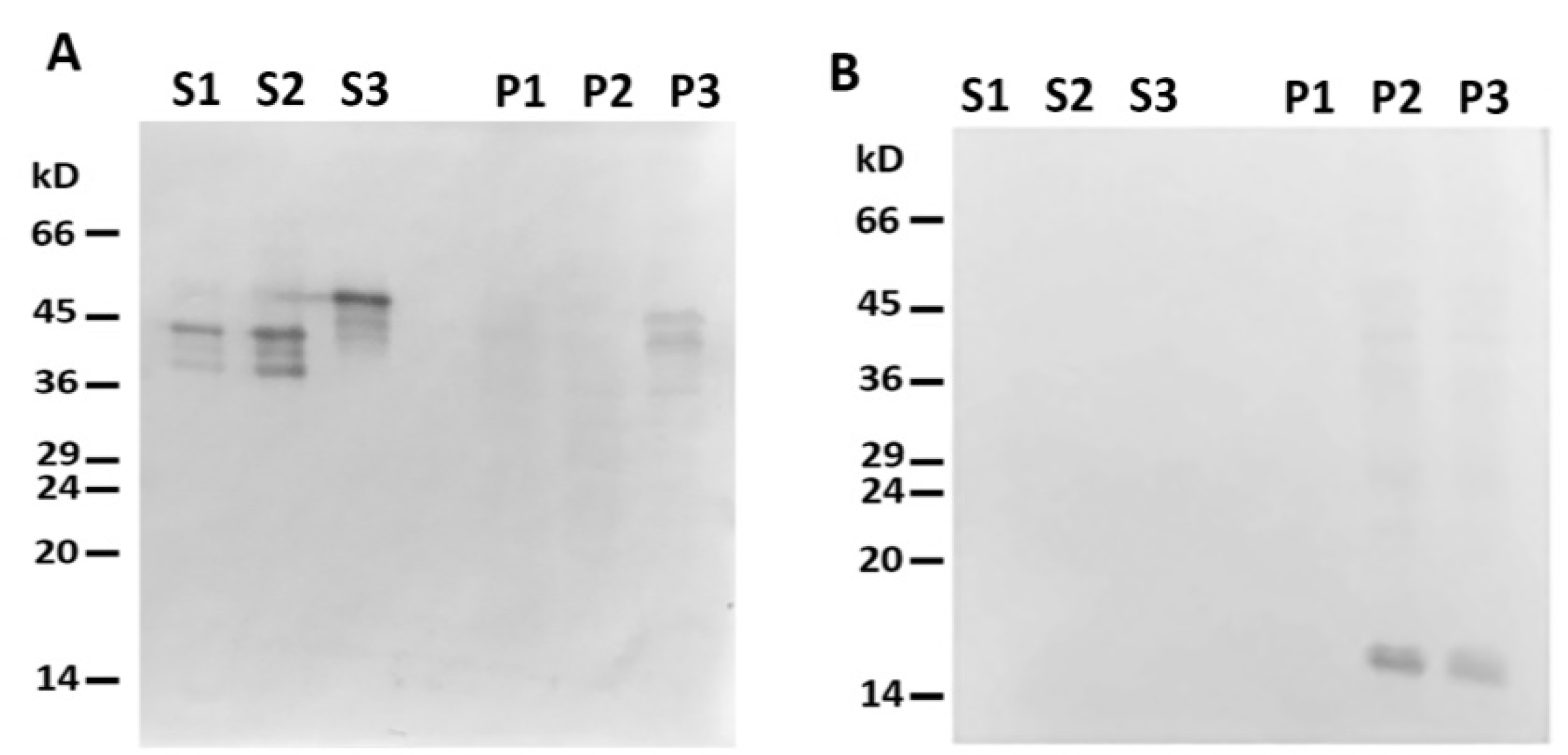
2.1. Desmin Binds to Mitochondria In Vitro
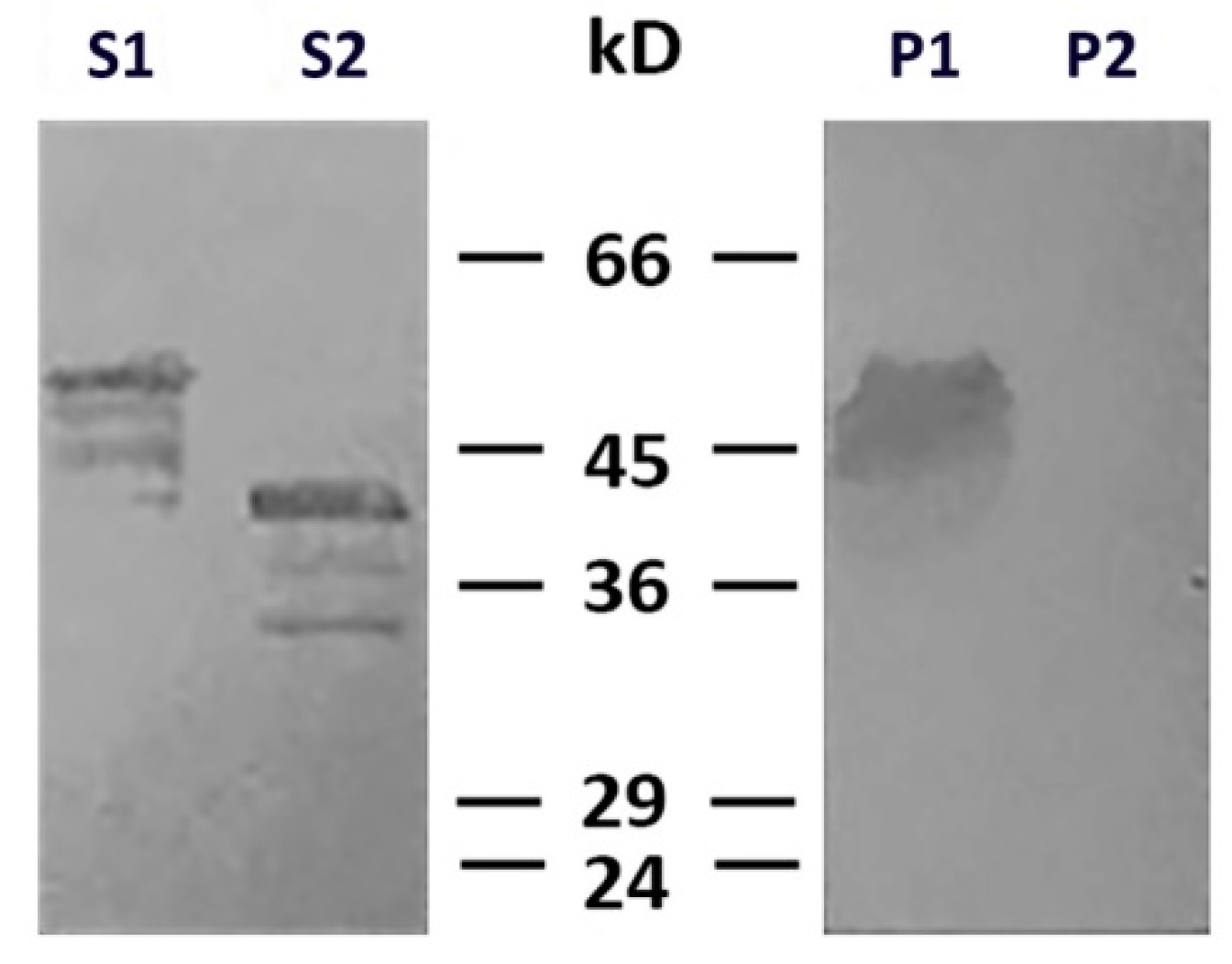
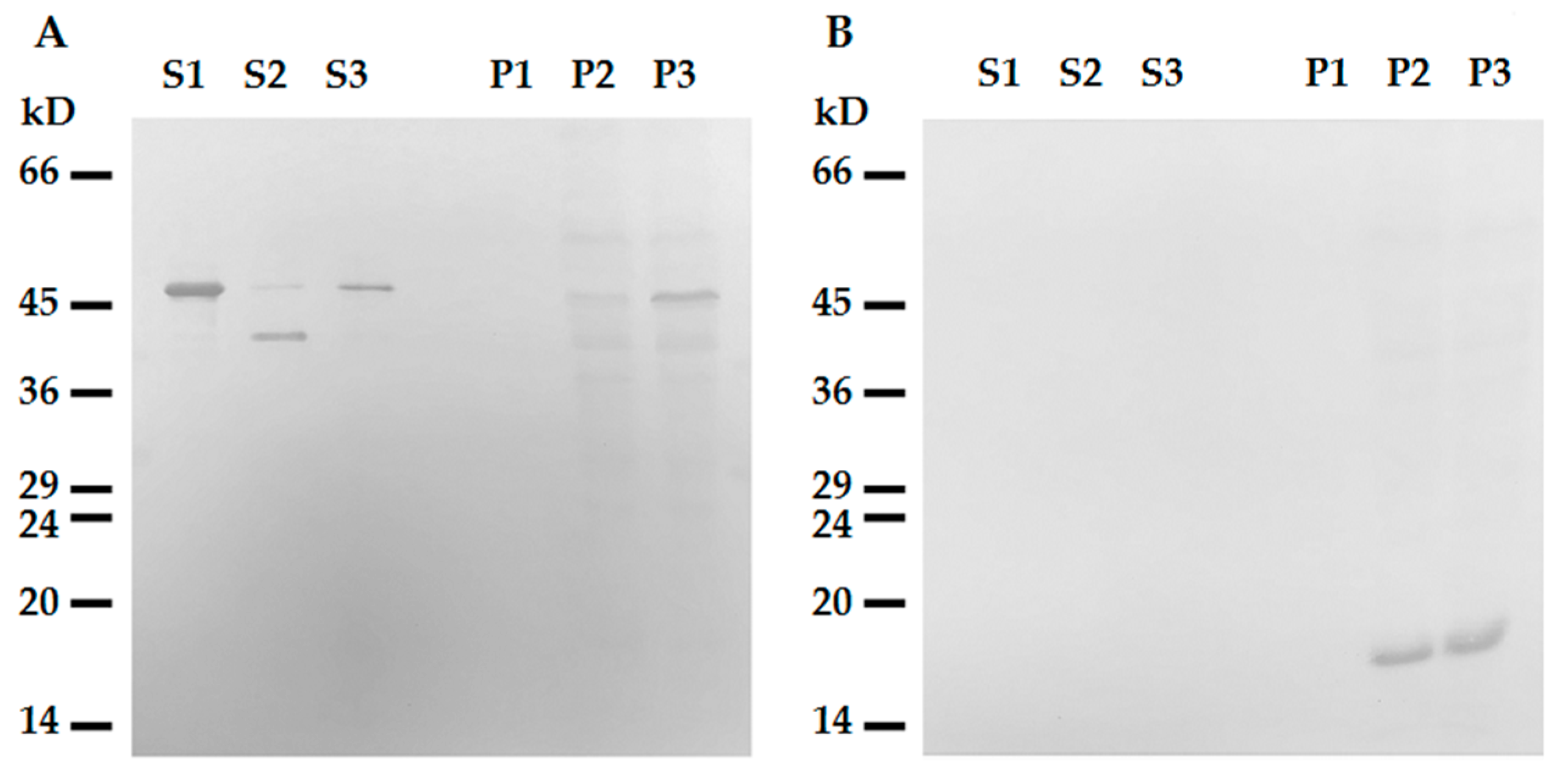
2.2. N-terminus of Desmin is Necessary for Binding to Mitochondria
3. Discussion
4. Materials and Methods
4.1. Cell Culture, Plasmids and Antibodies
4.2. Transfection of Cells and Immunofluorescence
4.3. Isolation of Mitochondria
4.4. Interaction of Mitochondria with Desmin
4.5. Detection of Mitochondrial Targeting Signals
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Etienne-Manneville, S. Cytoplasmic intermediate filaments in cell biology. Annu. Rev. Cell Dev. Biol. 2018, 34, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Steen, K.; Chen, D.; Wang, F.; Majumdar, R.; Chen, S.; Kumar, S.; Lombard, D.B.; Weigert, R.; Zieman, A.G.; Parent, C.A.; et al. A role for keratins in supporting mitochondrial organization and function in skin keratinocytes. Mol. Biol. Cell 2020. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, S.M.; Leube, R.E.; Schwarz, N. Keratin 6a mutations lead to impaired mitochondrial quality control. Br. J. Dermatol. 2020, 182, 636–647. [Google Scholar] [CrossRef]
- Milner, D.J.; Mavroidis, M.; Weisleder, N.; Capetanaki, Y. Desmin cytoskeleton linked to muscle mitochondrial distribution and respiratory function. J. Cell Biol. 2000, 150, 1283–1297. [Google Scholar] [CrossRef] [Green Version]
- Capetanaki, Y. Desmin cytoskeleton: A potential regulator of muscle mitochondrial behavior and function. Trends Cardiovasc. Med. 2002, 12, 339–348. [Google Scholar] [CrossRef]
- Chernoivanenko, I.S.; Matveeva, E.A.; Gelfand, V.I.; Goldman, R.D.; Minin, A.A. Mitochondrial membrane potential is regulated by vimentin intermediate filaments. FASEB J. 2015, 29, 820–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smolina, N.; Khudiakov, A.; Knyazeva, A.; Zlotina, A.; Sukhareva, K.; Kondratov, K.; Gogvadze, V.; Zhivotovsky, B.; Sejersen, T.; Kostareva, A. Desmin mutations result in mitochondrial dysfunction regardless of their aggregation properties. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2020, 1866, 165745. [Google Scholar] [CrossRef] [PubMed]
- Fountoulakis, M.; Soumaka, E.; Rapti, K.; Mavroidis, M.; Tsangaris, G.; Maris, A.; Weisleder, N.; Capetanaki, Y. Alterations in the heart mitochondrial proteome in a desmin null heart failure model. J. Mol. Cell Cardiol. 2005, 38, 461–474. [Google Scholar] [CrossRef]
- Cohen, S. Role of calpains in promoting desmin filaments depolymerization and muscle atrophy. BBA Mol. Cell Res. 2020. [Google Scholar] [CrossRef]
- Winter, L.; Abrahamsberg, C.; Wiche, G. Plectin isoform 1b mediates mitochondrion-intermediate filament network linkage and controls organelle shape. J. Cell. Biol. 2008, 181, 903–911. [Google Scholar] [CrossRef] [Green Version]
- Taylor, M.R.; Slavov, D.; Ku, L.; Di Lenarda, A.; Sinagra, G.; Carniel, E.; Haubold, K.; Boucek, M.M.; Ferguson, D.; Graw, S.L.; et al. Prevalence of desmin mutations in dilated cardiomyopathy. Circulation 2007, 115, 1244–1251. [Google Scholar] [CrossRef] [Green Version]
- Meier, M.; Padilla, G.P.; Herrmann, H.; Wedig, T.; Hergt, M.; Patel, T.R.; Burkhard, P. Vimentin coil 1A-A molecular switch involved in the initiation of filament elongation. J. Mol. Biol. 2009, 390, 245–261. [Google Scholar] [CrossRef] [PubMed]
- Nekrasova, O.E.; Mendez, M.G.; Chernoivanenko, I.S.; Tyurin-Kuzmin, P.A.; Kuczmarski, E.R.; Gelfand, V.I.; Goldman, R.D.; Minin, A.A. Vimentin intermediate filaments modulate the motility of mitochondria. Mol. Biol. Cell 2011, 22, 2282–2289. [Google Scholar] [CrossRef] [PubMed]
- Rapaport, D. Finding the right organelle. Targeting signals in mitochondrial outer-membrane proteins. EMBO Rep. 2003, 4, 948–952. [Google Scholar] [CrossRef] [Green Version]
- Emanuelsson, O.; Brunak, S.; von Heijne, G.; Nielsen, H. Locating proteins in the cell using TargetP, SignalP and related tools. Nat. Protoc. 2007, 2, 953–971. [Google Scholar] [CrossRef] [PubMed]
- Quirós, P.M.; Langer, T.; López-Otín, C. New roles for mitochondrial proteases in health, ageing and disease. Nat. Rev. Mol. Cell Biol. 2015, 16, 345–359. [Google Scholar] [CrossRef]
- Ebisui, C.; Tsujinaka, T.; Kido, Y.; Iijima, S.; Yano, M.; Shibata, H.; Tanaka, T.; Mori, T. Role of intracellular proteases in differentiation of L6 myoblast cells. Biochem. Mol. Biol. Int. 1994, 32, 515–521. [Google Scholar] [PubMed]
- Siklos, M.; BenAissa, M.; Thatcher, G.R. Cysteine proteases as therapeutic targets: Does selectivity matter? A systematic review of calpain and cathepsin inhibitors. Acta Pharm. Sin. B 2015, 5, 506–519. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.K.; Nath, R.; Posner, A.; Raser, K.J.; Buroker-Kilgore, M.; Hajimohammadreza, I. An alpha-mercaptoacrylic acid derivative is a selective nonpeptide cell-permeable calpain inhibitor and is neuroprotective. Proc. Natl. Acad. Sci. USA 1996, 93, 6687–6692. [Google Scholar] [CrossRef] [Green Version]
- Arrington, D.D.; Van Vleet, T.R.; Schnellmann, R.G. Calpain 10: A mitochondrial calpain and its role in calcium-induced mitochondrial dysfunction. Am. J. Physiol. Cell Physiol. 2006, 291, C1159–C1171. [Google Scholar] [CrossRef]
- Baron, C.P.; Jacobsen, S.; Purslow, P.P. Cleavage of desmin by cysteine proteases: Calpains and cathepsin B. Meat Sci. 2004, 68, 447–456. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, H.; Hofmann, I.; Franke, W.W. Identification of a nonapeptide motif in the vimentin head domain involved in intermediate filament assembly. J. Mol. Biol. 1992, 223, 637–650. [Google Scholar] [CrossRef]
- Pfanner, N.; Warscheid, B.; Wiedemann, N. Mitochondrial proteins: From biogenesis to functional networks. Nat. Rev. Mol. Cell Biol. 2019, 20, 267–284. [Google Scholar] [CrossRef]
- Liu, Z.; Yu, K.; Dong, J.; Zhao, L.; Liu, Z.; Zhang, Q.; Li, S.; Du, Y.; Cheng, H. Precise prediction of calpain cleavage sites and their aberrance caused by mutations in cancer. Front. Genet. 2019, 10, 715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vakhrusheva, A.; Endzhievskaya, S.; Zhuikov, V.; Nekrasova, T.; Parshina, E.; Ovsiannikova, N.; Popov, V.; Bagrov, D.; Minin, A.A.; Sokolova, O.S. The Role of Vimentin in Directional Migration of Rat Fibroblasts. Cytoskeleton 2019, 76, 467–476. [Google Scholar] [CrossRef]
- Erster, O.; Liscovitch, M. A modified inverse PCR procedure for insertion, deletion, or replacement of a DNA fragment in a target sequence and its application in the ligand interaction scan method for generation of ligand-regulated proteins. Methods Mol. Biol. 2010, 634, 157–174. [Google Scholar]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dayal, A.A.; Medvedeva, N.V.; Nekrasova, T.M.; Duhalin, S.D.; Surin, A.K.; Minin, A.A. Desmin Interacts Directly with Mitochondria. Int. J. Mol. Sci. 2020, 21, 8122. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218122
Dayal AA, Medvedeva NV, Nekrasova TM, Duhalin SD, Surin AK, Minin AA. Desmin Interacts Directly with Mitochondria. International Journal of Molecular Sciences. 2020; 21(21):8122. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218122
Chicago/Turabian StyleDayal, Alexander A., Natalia V. Medvedeva, Tatiana M. Nekrasova, Sergey D. Duhalin, Alexey K. Surin, and Alexander A. Minin. 2020. "Desmin Interacts Directly with Mitochondria" International Journal of Molecular Sciences 21, no. 21: 8122. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218122