Interleukin-34, a Novel Paracrine/Autocrine Factor in Mouse Testis, and Its Possible Role in the Development of Spermatogonial Cells In Vitro

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
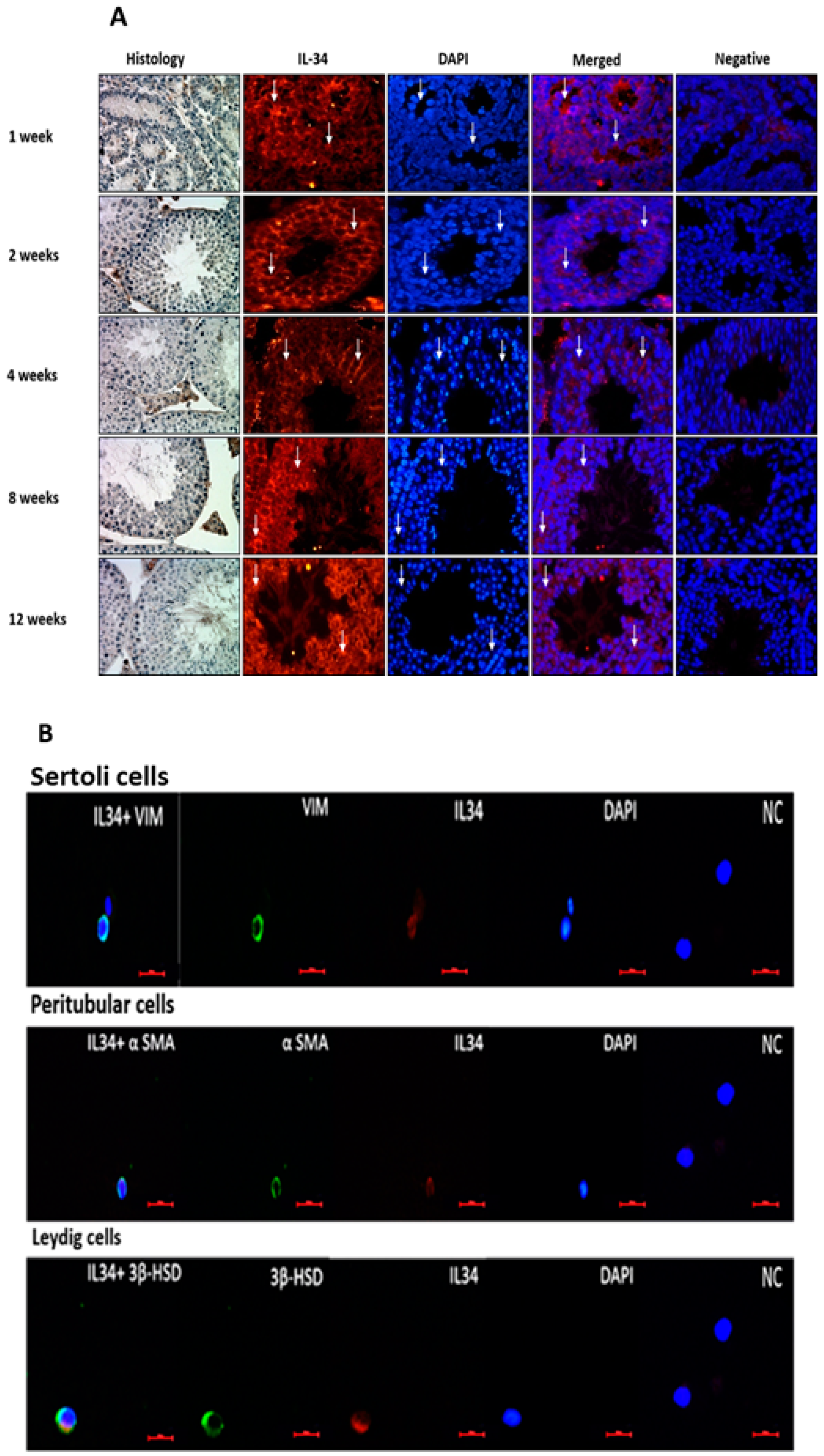
2.1. Localization and Expression Levels of IL-34 in Testicular Tissue
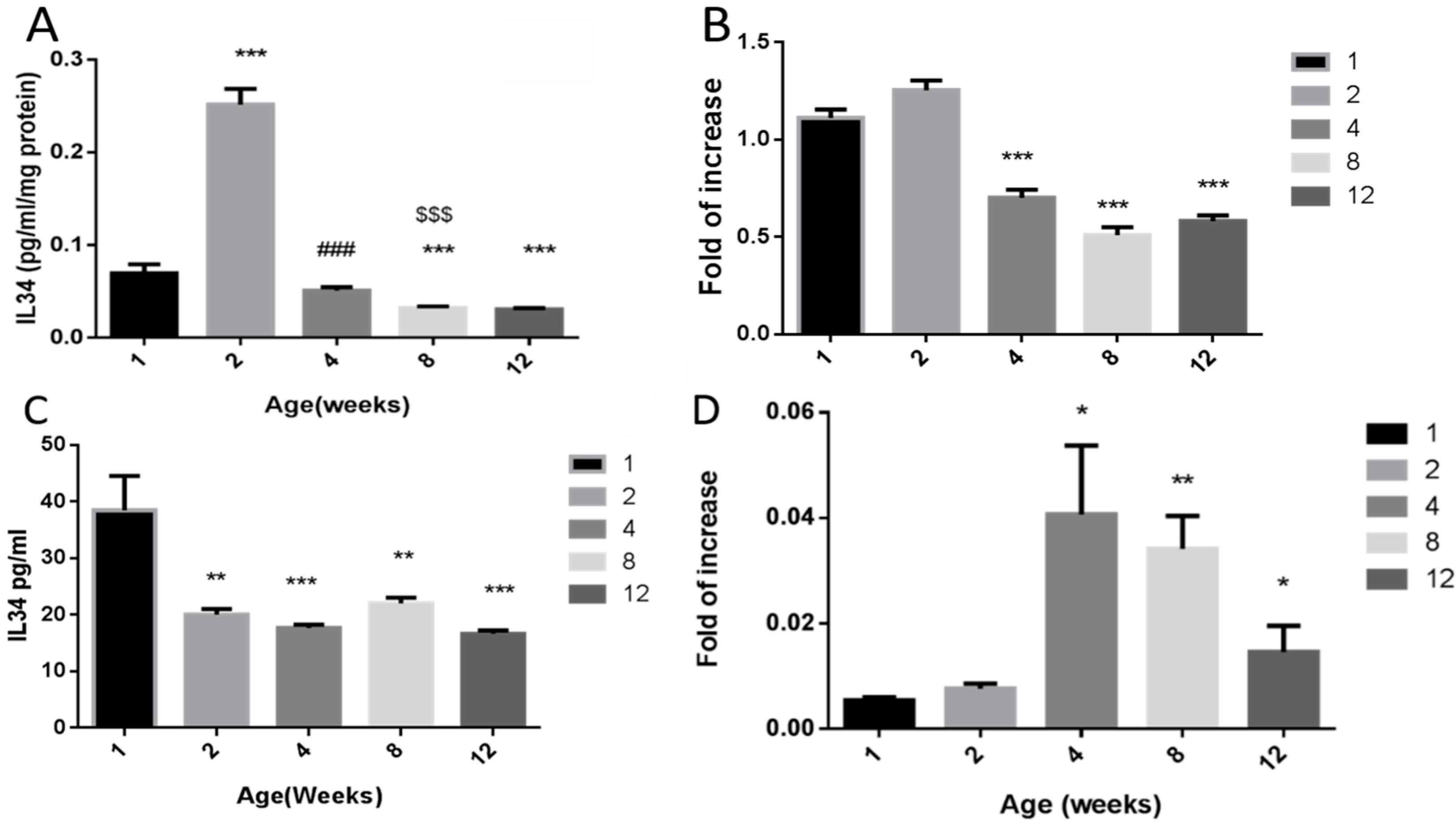
2.2. Effect of Age on the Levels of IL-34 in Testicular Homogenates and Sertoli Cell-Conditioned Media
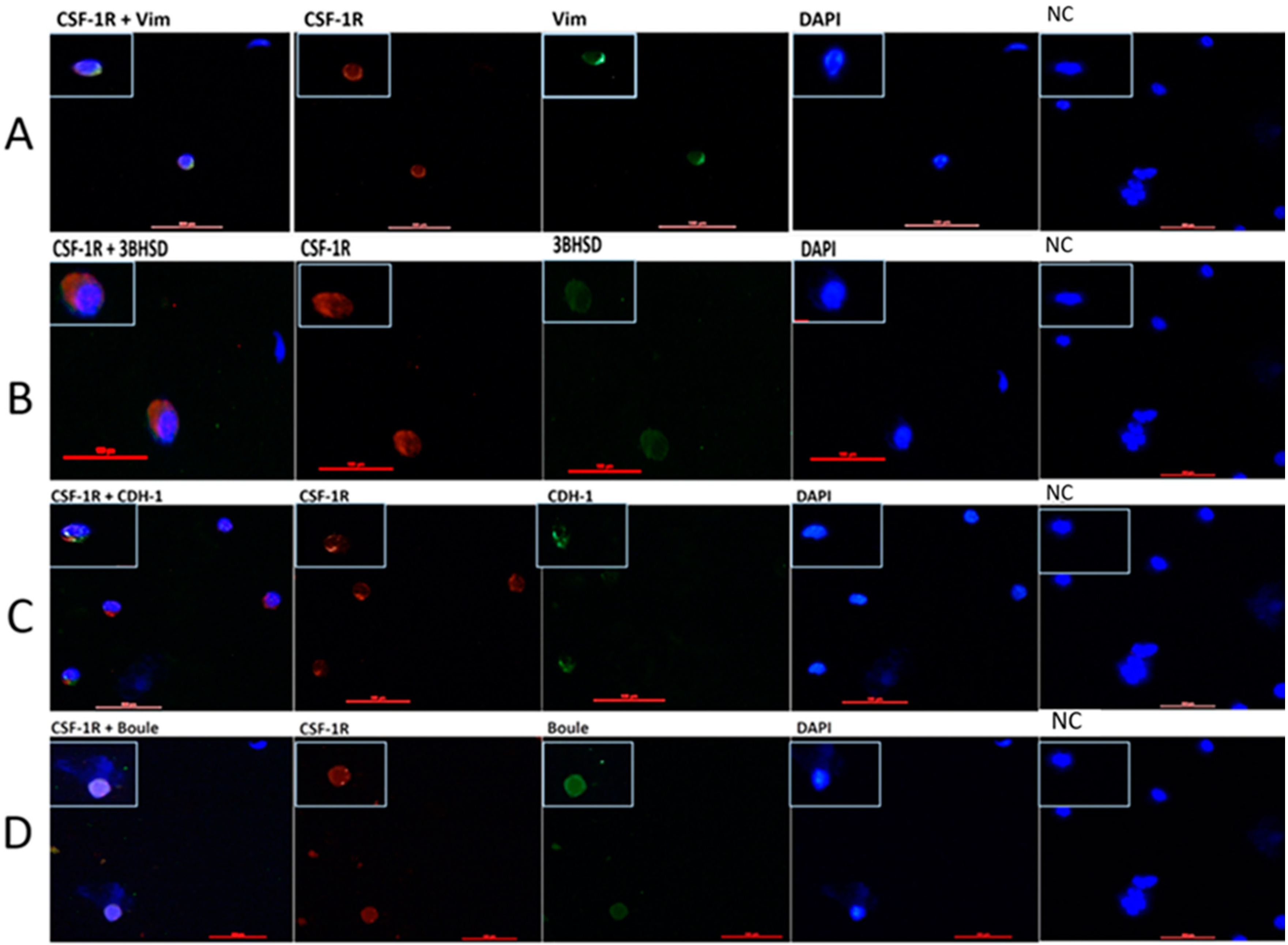
2.3. Localization of CSF-1R in Testicular Cells
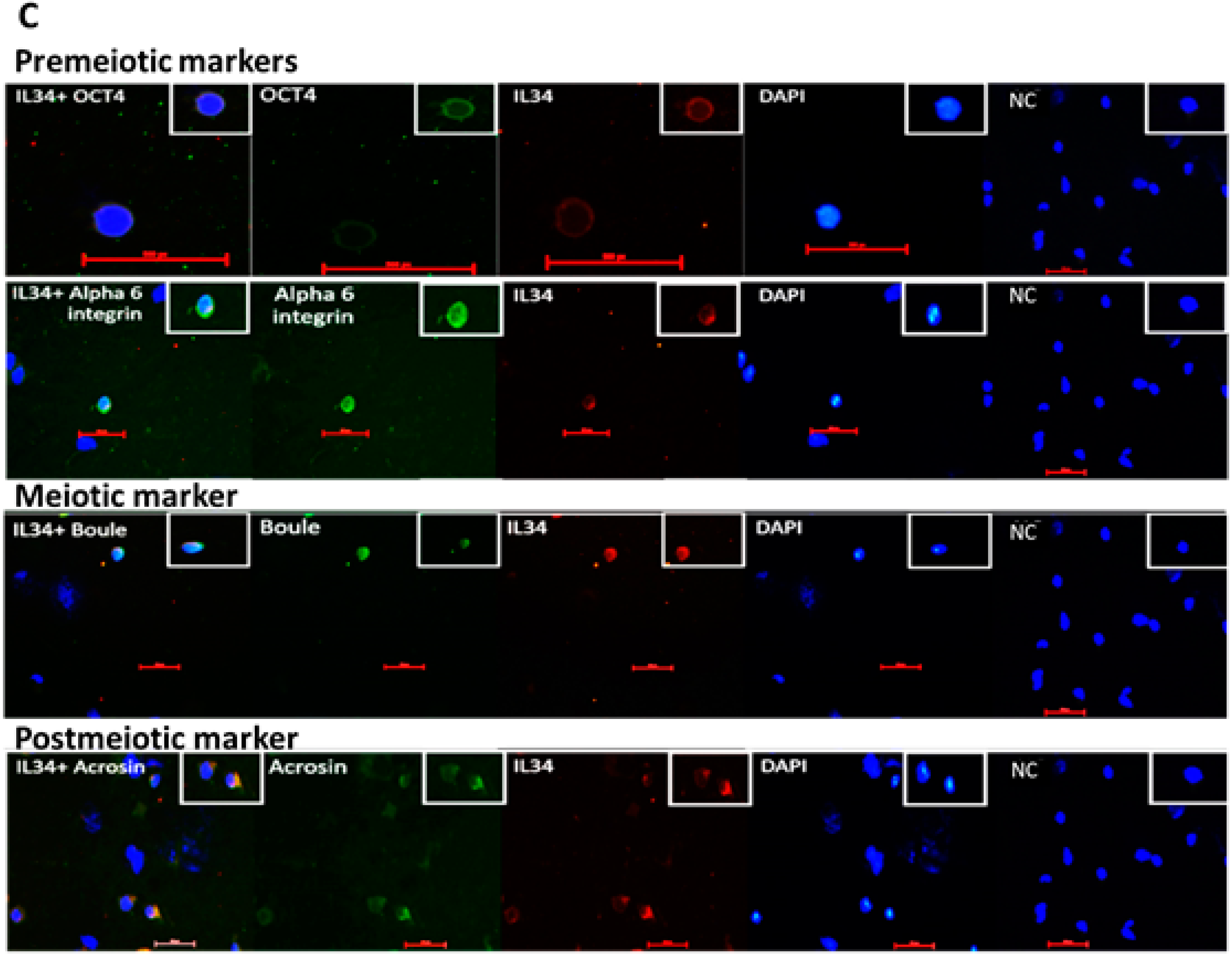
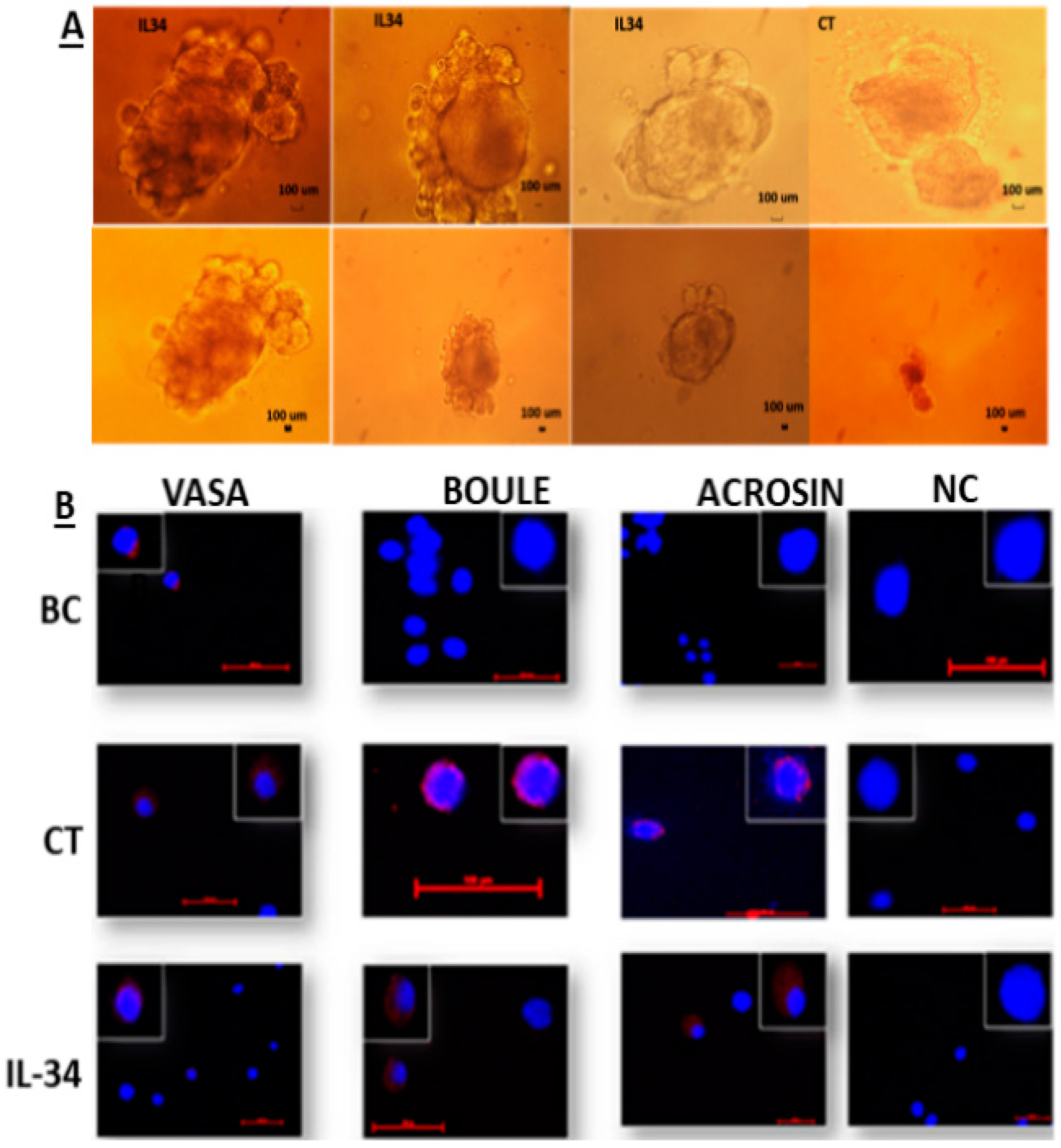
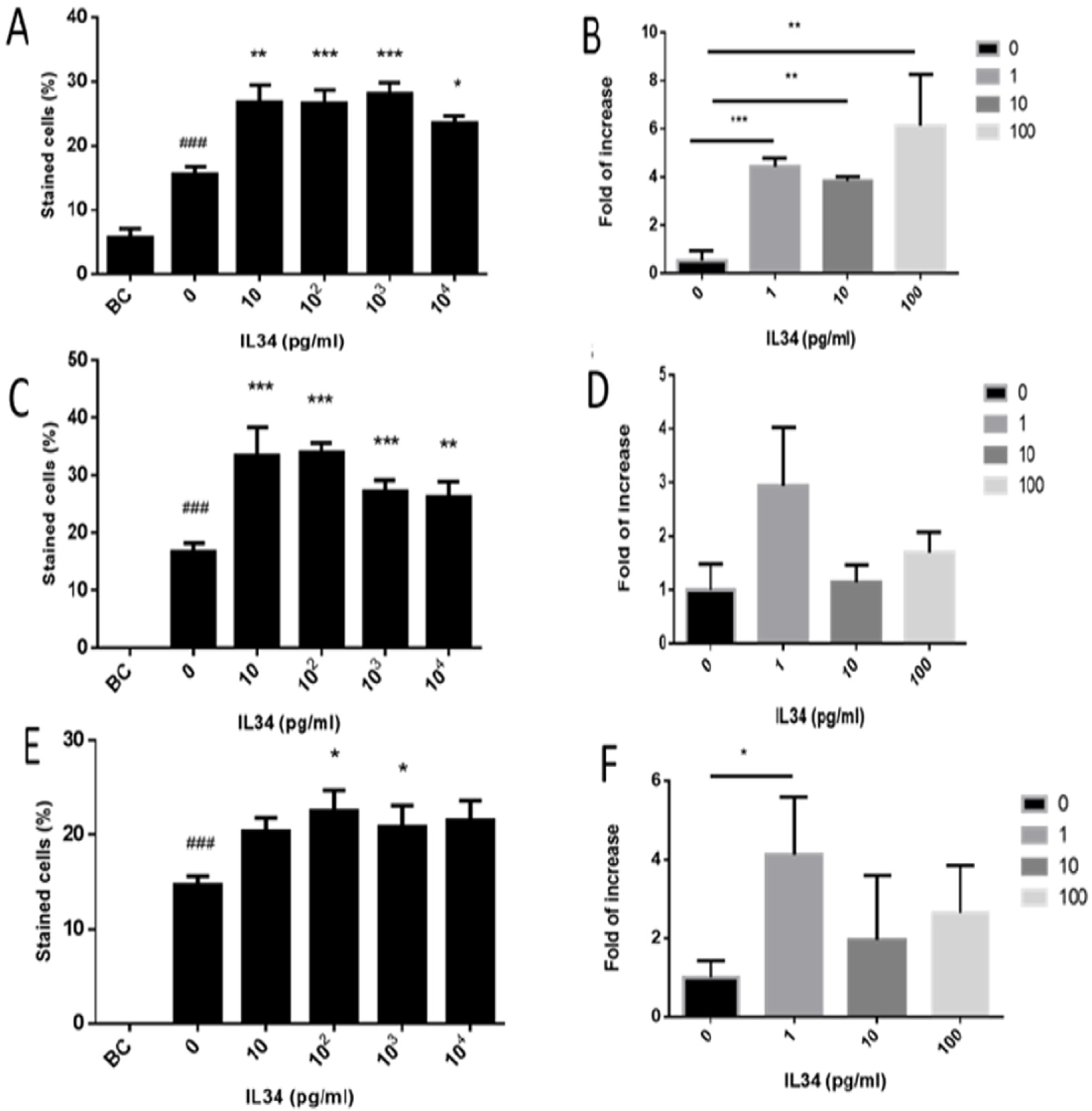
2.4. Involvement of IL-34 in the Development of Spermatogenesis In Vitro
2.5. Effect of IL-34 on the Levels of Developed Premeiotic, Meiotic, and Postmeiotic Cells In Vitro
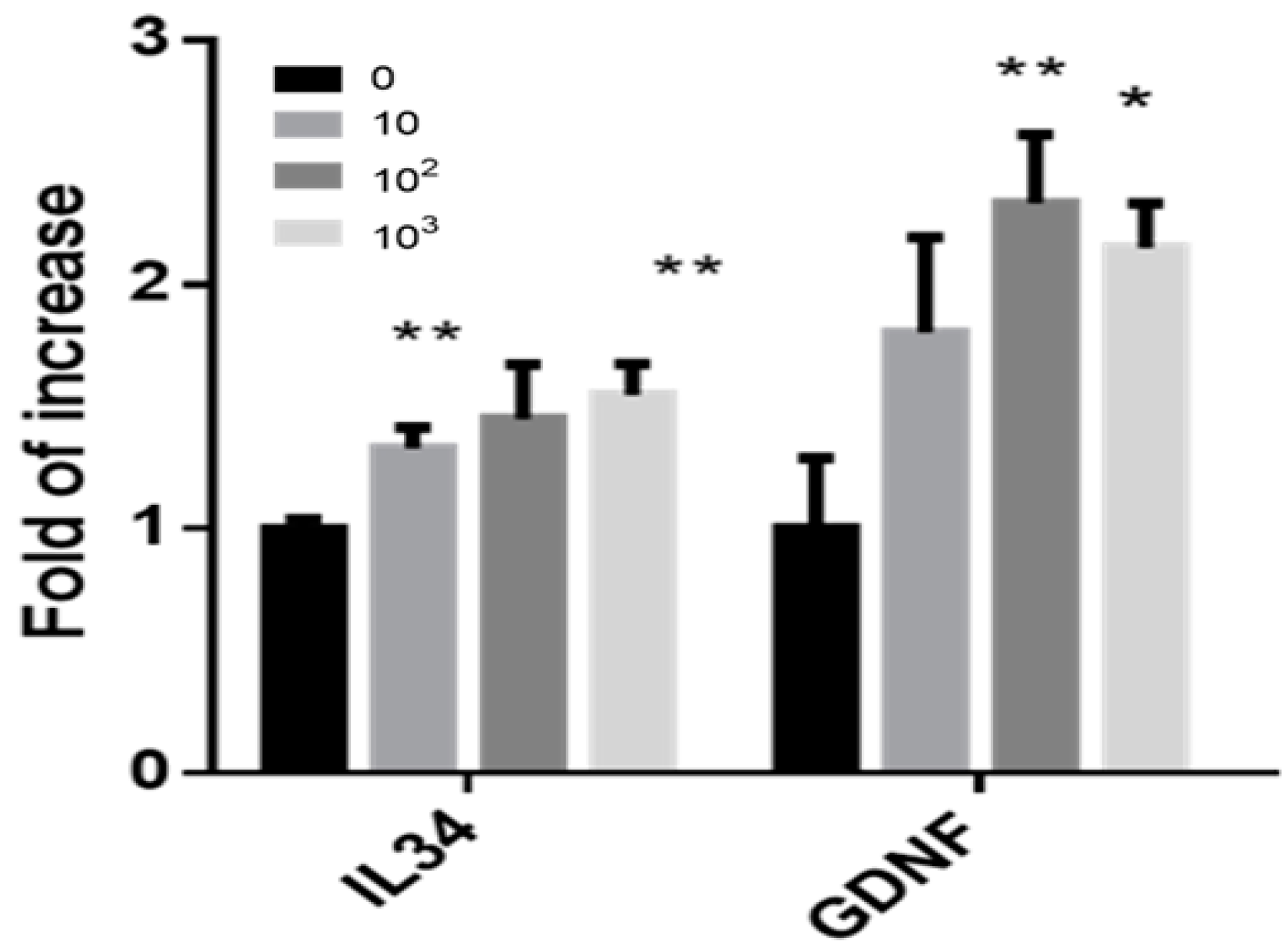
2.6. Effect of IL-34 on the Expression Levels of Growth Factors in Cells Developed in MCS
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Chemicals and Reagents
4.3. Isolation and Culture of Mouse Sertoli Cells
4.4. Immunofluorescence Staining of Mouse Testicular Tissue
4.5. Immunofluorescence Staining of Cells Isolated from Seminiferous Tubules
4.6. Preparation of Testicular Homogenates
4.7. Extraction of Total RNA for Real-Time PCR Analysis
4.8. Isolation of Tubular/Spermatogonial Cells
4.9. Culture of Isolated Spermatogonial Cells In Vitro in Methylcellulose Culture System (MCS)
4.10. Data Handling and Statistical Evaluation
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huleihel, M.; Lunenfeld, E. Regulation of spermatogenesis by paracrine/autocrine testicular factors. Asian J. Androl. 2004, 6, 259–268. [Google Scholar] [PubMed]
- Mahmoud, H. Concise review: Spermatogenesis in an artificial three-dimensional system. Stem Cells 2012, 30, 2355–2360. [Google Scholar] [CrossRef] [PubMed]
- Huleihel, M.; Nourashrafeddin, S.; Plant, T.M. Application of three-dimensional culture systems to study mammalian spermatogenesis, with an emphasis on the rhesus monkey (Macaca mulatta). Asian J. Aandrol. 2015, 17, 972. [Google Scholar] [CrossRef] [PubMed]
- França, L.R.; Hess, R.A.; Dufour, J.; Hofmann, M.-C.; Griswold, M. The Sertoli cell: One hundred fifty years of beauty and plasticity. Andrology 2016, 4, 189–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fayomi, A.P.; Orwig, K.E. Spermatogonial stem cells and spermatogenesis in mice, monkeys and men. Stem Cell Res. 2018, 29, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Sadri-Ardekani, H.; Atala, A. Testicular tissue cryopreservation and spermatogonial stem cell transplantation to restore fertility: From bench to bedside. Stem Cell Res. Ther. 2014, 5, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castrillon, D.H.; Quade, B.J.; Wang, T.; Quigley, C.; Crum, C.P. The human VASA gene is specifically expressed in the germ cell lineage. Proc. Nat. Acad. Sci. USA 2000, 97, 9585–9590. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Jung, H.; Yoon, M. VASA (DDX4) is a putative marker for spermatogonia, spermatocytes and round spermatids in stallions. Reprod. Domes. Anim. 2015, 50, 1032–1038. [Google Scholar] [CrossRef]
- Kee, K.; Angeles, V.T.; Flores, M.; Nguyen, H.N.; Pera, R.A.R. Human DAZL, DAZ and BOULE genes modulate primordial germ-cell and haploid gamete formation. Nature 2009, 462, 222–225. [Google Scholar] [CrossRef] [PubMed]
- Flörke-Gerloff, S.; Töpfer-Petersen, E.; Müller-Esterl, W.; Schill, W.-B.; Engel, W. Acrosin and the acrosome in human spermatogenesis. Hum. Genet. 1983, 65, 61–67. [Google Scholar] [CrossRef]
- Xu, E.Y.; Lee, D.F.; Klebes, A.; Turek, P.J.; Kornberg, T.B.; Reijo Pera, R.A. Human BOULE gene rescues meiotic defects in infertile flies. Hum. Mol. Genet. 2003, 12, 169–175. [Google Scholar] [CrossRef] [PubMed]
- Airaksinen, M.S.; Saarma, M. The GDNF family: Signalling, biological functions and therapeutic value. Nat. Rev. Neurosci. 2002, 3, 383–394. [Google Scholar] [CrossRef] [PubMed]
- Meng, X.; Lindahl, M.; Hyvönen, M.E.; Parvinen, M.; de Rooij, D.G.; Hess, M.W.; Raatikainen-Ahokas, A.; Sainio, K.; Rauvala, H.; Lakso, M. Regulation of cell fate decision of undifferentiated spermatogonia by GDNF. Science 2000, 287, 1489–1493. [Google Scholar] [CrossRef]
- Bokemeyer, C.; Kuczyk, M.A.; Dunn, T.; Serth, J.; Hartmann, K.; Jonasson, J.; Pietsch, T.; Jonas, U.; Schmoll, H.-J. Expression of stem-cell factor and its receptor c-kit protein in normal testicular tissue and malignant germ-cell tumours. J. Cancer Res. Clin. Oncol. 1996, 122, 301–306. [Google Scholar] [CrossRef] [PubMed]
- Oatley, J.M.; Oatley, M.J.; Avarbock, M.R.; Tobias, J.W.; Brinster, R.L. Colony stimulating factor 1 is an extrinsic stimulator of mouse spermatogonial stem cell self-renewal. Development 2009, 136, 1191–1199. [Google Scholar] [CrossRef] [Green Version]
- Prasanth, S.G.; Giran, H.M.; Ali, S. Biology of protooncogene c-kit receptor and spermatogenesis. Curr. Pharm. 2004, 2, 47–60. [Google Scholar] [CrossRef]
- Su, H.; Wang, Y.; Söder, O.; Hou, M. Identification of an Isoform of Colony-Stimulating Factor 1 Receptor mRNA in the Rat Testis. Biochem. Genet. 2014, 52, 310–319. [Google Scholar] [CrossRef]
- Kokkinaki, M.; Lee, T.-L.; He, Z.; Jiang, J.; Golestaneh, N.; Hofmann, M.-C.; Chan, W.-Y.; Dym, M. The molecular signature of spermatogonial stem/progenitor cells in the 6-day-old mouse testis. Biol. Reprod. 2009, 80, 707–717. [Google Scholar] [CrossRef] [Green Version]
- Zelante, T.; Ricciardi-Castagnoli, P. The yin-yang nature of CSF1R-binding cytokines. Nat. Immunol. 2012, 13, 717–719. [Google Scholar] [CrossRef]
- Chitu, V.; Gokhan, Ş.; Nandi, S.; Mehler, M.F.; Stanley, E.R. Emerging roles for CSF-1 receptor and its ligands in the nervous system. Trends Neurosci. 2016, 39, 378–393. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Szretter, K.J.; Vermi, W.; Gilfillan, S.; Rossini, C.; Cella, M.; Barrow, A.D.; Diamond, M.S.; Colonna, M. IL-34 is a tissue-restricted ligand of CSF1R required for the development of Langerhans cells and microglia. Nat. Immunol. 2012, 13, 753–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Colonna, M. Interkeukin-34, a cytokine crucial for the differentiation and maintenance of tissue resident macrophages and Langerhans cells. Eur. J. Immunol. 2014, 44, 1575–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wynn, T.A.; Chawla, A.; Pollard, J.W. Macrophage biology in development, homeostasis and disease. Nature 2013, 496, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Baghdadi, M.; Ishikawa, K.; Endo, H.; Umeyama, Y.; Ataka, T.; Wada, H.; Oyamada, Y.; Hyakushima, N.; Seino, K.-I. Enhanced expression of IL-34 in an inflammatory cyst of the submandibular gland: A case report. Inflamm. Regen. 2018, 38, 12. [Google Scholar] [CrossRef] [Green Version]
- Nandi, S.; Cioce, M.; Yeung, Y.-G.; Nieves, E.; Tesfa, L.; Lin, H.; Hsu, A.W.; Halenbeck, R.; Cheng, H.-Y.; Gokhan, S. Receptor-type protein-tyrosine phosphatase ζ is a functional receptor for interleukin-34. J. Biol. Chem. 2013, 288, 21972–21986. [Google Scholar] [CrossRef] [Green Version]
- Elhija, M.A.; Lunenfeld, E.; Schlatt, S.; Huleihel, M. Differentiation of murine male germ cells to spermatozoa in a soft agar culture system. Asian J. Androl. 2012, 14, 285. [Google Scholar] [CrossRef] [Green Version]
- Stukenborg, J.-B.; Schlatt, S.; Simoni, M.; Yeung, C.-H.; Elhija, M.A.; Luetjens, C.M.; Huleihel, M.; Wistuba, J. New horizons for in vitro spermatogenesis? An update on novel three-dimensional culture systems as tools for meiotic and post-meiotic differentiation of testicular germ cells. Mol. Hum. Reprod. 2009, 15, 521–529. [Google Scholar] [CrossRef] [Green Version]
- Abofoul-Azab, M.; AbuMadighem, A.; Lunenfeld, E.; Kapelushnik, J.; Shi, Q.; Pinkas, H.; Huleihel, M. Development of postmeiotic cells in vitro from spermatogonial cells of prepubertal cancer patients. Stem Cells Dev. 2018, 27, 1007–1020. [Google Scholar] [CrossRef]
- AbuMadighem, A.; Solomon, R.; Stepanovsky, A.; Kapelushnik, J.; Shi, Q.; Meese, E.; Lunenfeld, E.; Huleihel, M. Development of spermatogenesis in vitro in three-dimensional culture from spermatogonial cells of busulfan-treated immature mice. Int. J. Mol. Sci. 2018, 19, 3804. [Google Scholar] [CrossRef] [Green Version]
- Gan, H.; Cai, T.; Lin, X.; Wu, Y.; Wang, X.; Yang, F.; Han, C. Integrative Proteomic and Transcriptomic Analyses Reveal Multiple Post-transcriptional Regulatory Mechanisms of Mouse Spermatogenesis. Mol. Cell. Proteom. 2013, 12, 1144–1157. [Google Scholar] [CrossRef] [Green Version]
- Wei, S.; Nandi, S.; Chitu, V.; Yeung, Y.-G.; Yu, W.; Huang, M.; Williams, L.T.; Lin, H.; Stanley, E.R. Functional overlap but differential expression of CSF-1 and IL-34 in their CSF-1 receptor-mediated regulation of myeloid cells. J. Leukoc. Biol. 2010, 88, 495–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, D.; Ma, A.; Huang, Z.; Wang, G.; Wang, T.; Xia, D.; Ma, B. Transcriptome Analysis for Identification of Genes Related to Gonad Differentiation, Growth, Immune Response and Marker Discovery in The Turbot (Scophthalmus maximus). PLoS ONE 2016, 11, e0149414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huleihel, M.; Fadlon, E.; Abuelhija, A.; Haber, E.P.; Lunenfeld, E. Glial cell line-derived neurotrophic factor (GDNF) induced migration of spermatogonial cells in vitro via MEK and NF-kB pathways. Differentiation 2013, 86, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Abofoul-Azab, M.; Lunenfeld, E.; Levitas, E.; Zeadna, A.; Younis, J.S.; Bar-Ami, S.; Huleihel, M. Identification of Premeiotic, Meiotic, and Postmeiotic Cells in Testicular Biopsies Without Sperm from Sertoli Cell-Only Syndrome Patients. Int. J. Mol. Sci. 2019, 20, 470. [Google Scholar] [CrossRef] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sawaied, A.; Lunenfeld, E.; Huleihel, M. Interleukin-34, a Novel Paracrine/Autocrine Factor in Mouse Testis, and Its Possible Role in the Development of Spermatogonial Cells In Vitro. Int. J. Mol. Sci. 2020, 21, 8143. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218143
Sawaied A, Lunenfeld E, Huleihel M. Interleukin-34, a Novel Paracrine/Autocrine Factor in Mouse Testis, and Its Possible Role in the Development of Spermatogonial Cells In Vitro. International Journal of Molecular Sciences. 2020; 21(21):8143. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218143
Chicago/Turabian StyleSawaied, Alaa, Eitan Lunenfeld, and Mahmoud Huleihel. 2020. "Interleukin-34, a Novel Paracrine/Autocrine Factor in Mouse Testis, and Its Possible Role in the Development of Spermatogonial Cells In Vitro" International Journal of Molecular Sciences 21, no. 21: 8143. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21218143