From Inflammation to the Onset of Fibrosis through A2A Receptors in Kidneys from Deceased Donors

 ,
,
Abstract
:1. Introduction
2. Results
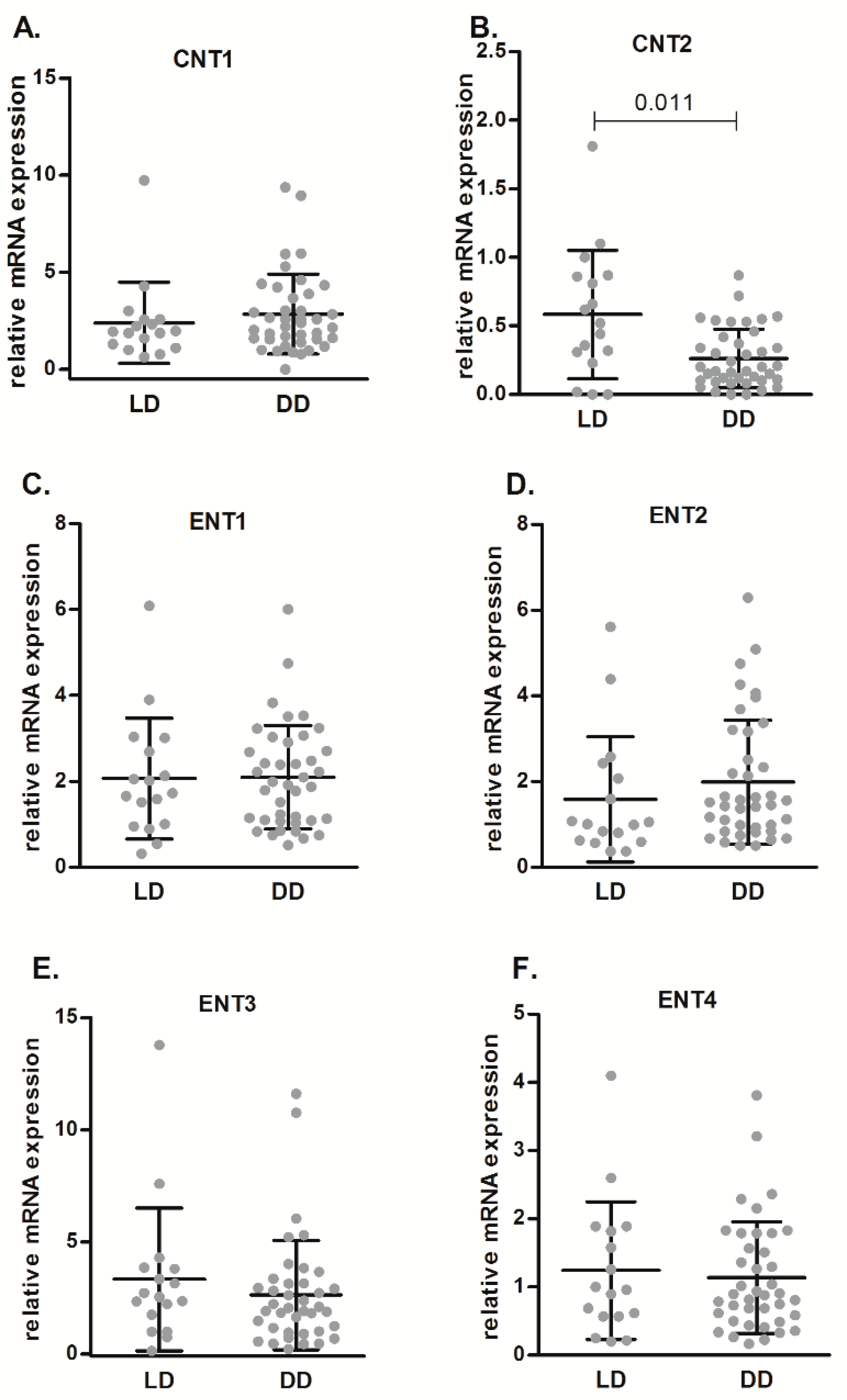
2.1. Differences in Purinome Gene Expression in Renal Biopsies from DD and LD
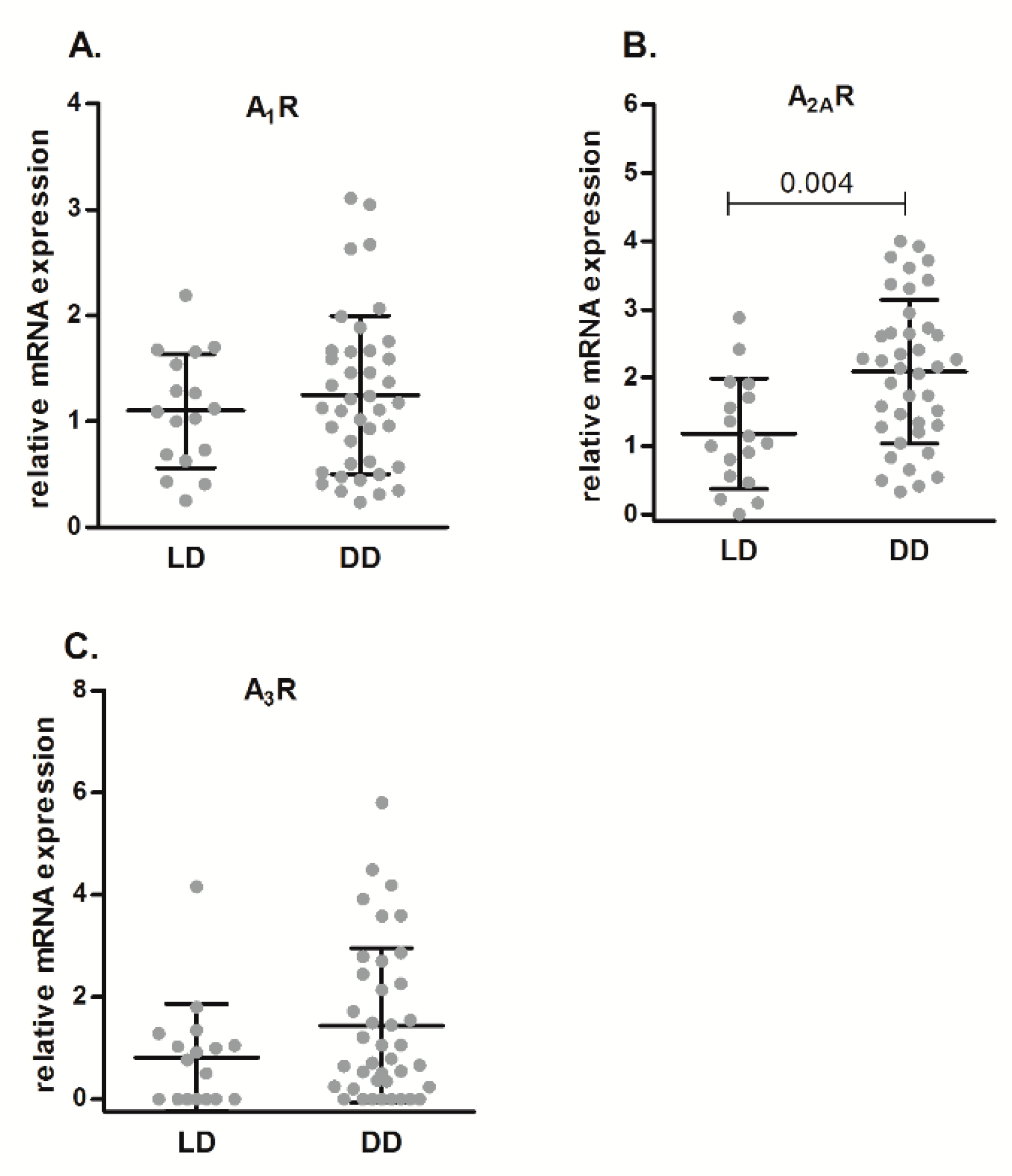
2.2. Correlations of A2AR mRNA Expression Levels
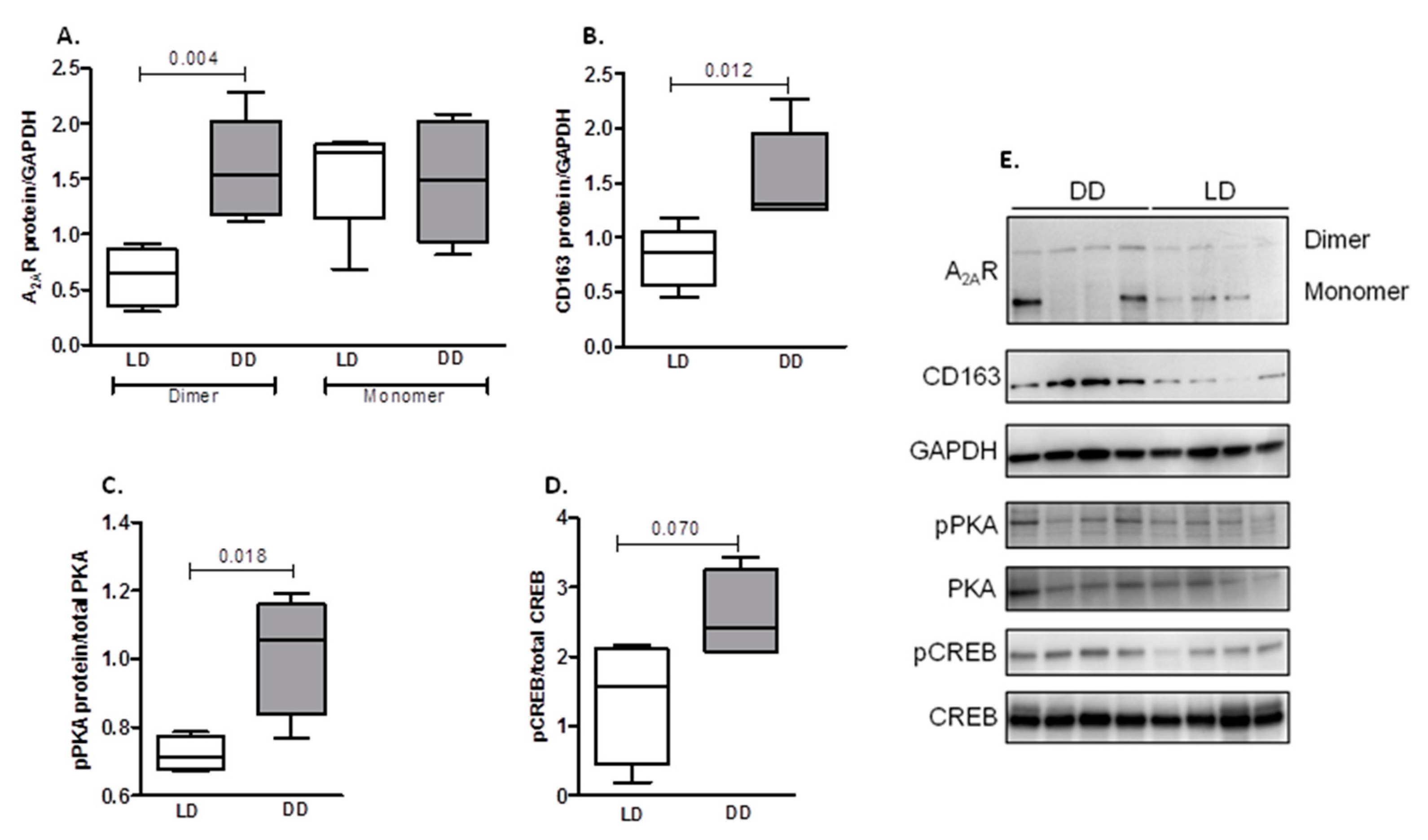
2.3. Protein Expression in Renal Biopsies from DD and LD
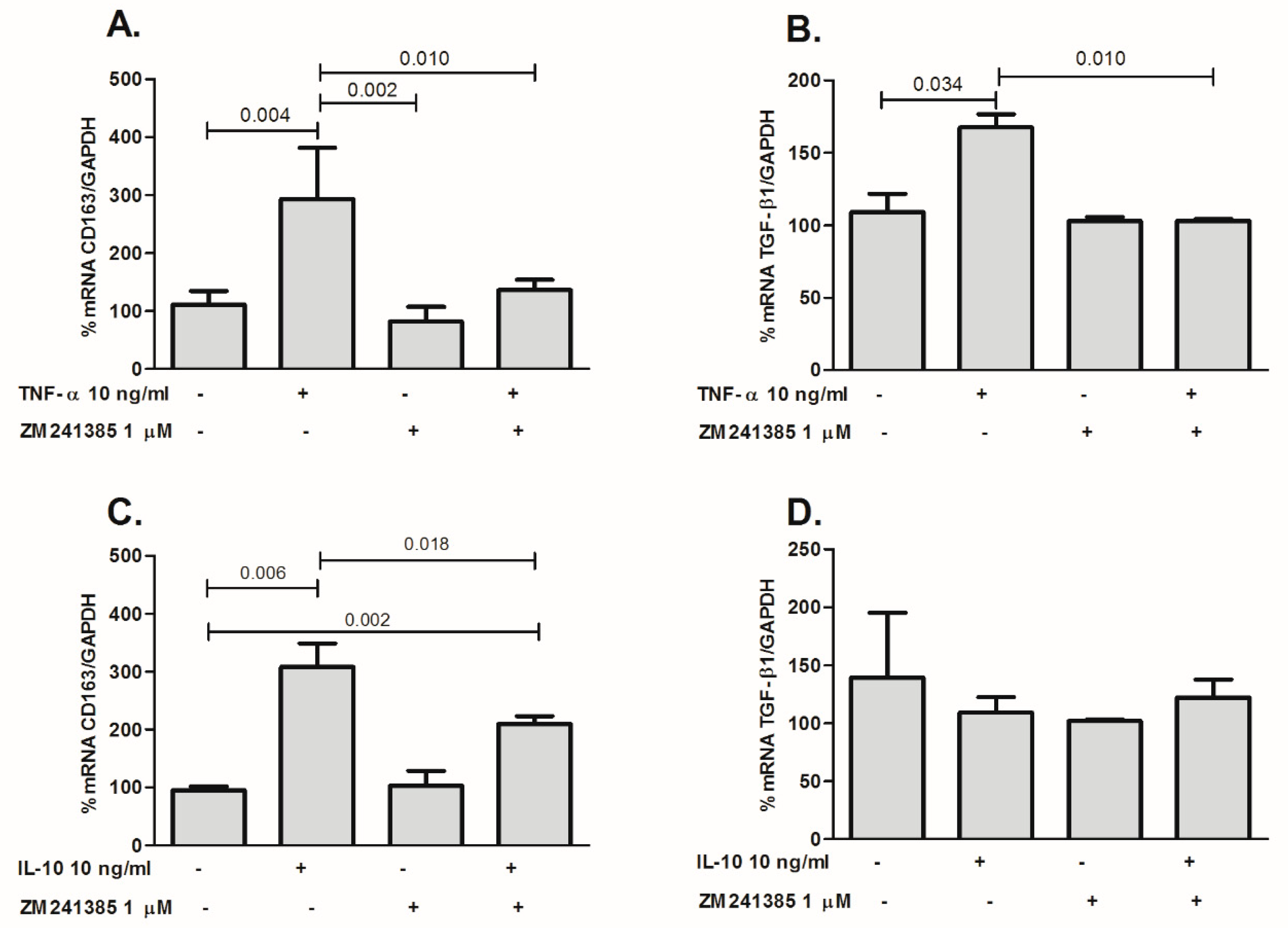
2.4. TNF-α Increases Expression of CD163 and TGF-β1 through A2AR in THP-1 Cells
3. Discussion
4. Materials and Methods
4.1. Donors, Patients, and Kidney Samples
4.2. Cell Culture and Treatments
4.3. Real Time PCR
4.3.1. Renal Tissue
4.3.2. THP-1 Cells
4.4. Western Blot
4.5. Statistics
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| DD | Deceased donor |
| LD | Living donor |
| A2AR | A2A adenosine receptor |
| KTR | Kidney transplant recipient |
| NT | Nucleoside transporter |
| SAHH | S-adenosyl-L-homocysteine hydrolase |
| AC | Adenylate cyclase |
| ADK | Adenosine kinase |
| PKA | Protein kinase A |
| SAH | S-adenosylhomocysteine |
| ENT | Equilibrative nucleoside transporter |
| CNT | Concentrative nucleoside transporter |
| hENT | Human ENT |
| hCNT | Human CNT |
References
- Opelz, G. Impact of HLA compatibility on survival of kidney transplants from unrelated live donors. Transplantation 1997, 64, 1473–1475. [Google Scholar] [CrossRef] [PubMed]
- Moreso, F.; Serón, D.; Gil-Vernet, S.; Riera, L.; Fulladosa, X.; Ramos, R.; Alsina, J.; Grinyó, J.M. Donor age and delayed graft function as predictors of renal allograft survival in rejection-free patients. Nephrol. Dial. Transpl. 1999, 14, 930–935. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snoeijs, M.G.J.; van Bijnen, A.; Swennen, E.; Haenen, G.R.M.M.; Roberts, L.J.; Christiaans, M.H.L.; Peppelenbosch, A.G.; Buurman, W.A.; Ernest van Heurn, L.W. Tubular epithelial injury and inflammation after ischemia and reperfusion in human kidney transplantation. Ann. Surg. 2011, 253, 598–604. [Google Scholar] [CrossRef] [PubMed]
- Pessione, F.; Cohen, S.; Durand, D.; Hourmant, M.; Kessler, M.; Legendre, C.; Mourad, G.; Noël, C.; Peraldi, M.-N.; Pouteil-Noble, C.; et al. Multivariate analysis of donor risk factors for graft survival in kidney transplantation. Transplantation 2003, 75, 361–367. [Google Scholar] [CrossRef]
- van der Hoeven, J.A.; Ploeg, R.J.; Postema, F.; Molema, I.; de Vos, P.; Girbes, A.R.; van Suylichem, P.T.; van Schilfgaarde, R.; Ter Horst, G.J. Induction of organ dysfunction and up-regulation of inflammatory markers in the liver and kidneys of hypotensive brain dead rats: A model to study marginal organ donors. Transplantation 1999, 68, 1884–1890. [Google Scholar] [CrossRef]
- van der Hoeven, J.A.B.; Molema, G.; Ter Horst, G.J.; Freund, R.L.; Wiersema, J.; van Schilfgaarde, R.; Leuvenink, H.G.D.; Ploeg, R.J. Relationship between duration of brain death and hemodynamic (in)stability on progressive dysfunction and increased immunologic activation of donor kidneys. Kidney Int. 2003, 64, 1874–1882. [Google Scholar] [CrossRef] [Green Version]
- Nijboer, W.N.; Schuurs, T.A.; van der Hoeven, J.A.B.; Fekken, S.; Wiersema-Buist, J.; Leuvenink, H.G.D.; Hofker, S.; Homan van der Heide, J.J.; van Son, W.J.; Ploeg, R.J. Effect of brain death on gene expression and tissue activation in human donor kidneys. Transplantation 2004, 78, 978–986. [Google Scholar] [CrossRef] [Green Version]
- Nagareda, T.; Kinoshita, Y.; Tanaka, A.; Takeda, M.; Sakano, T.; Yawata, K.; Sugimoto, T.; Nishizawa, Y.; Terada, N. Clinicopathology of kidneys from brain-dead patients treated with vasopressin and epinephrine. Kidney Int. 1993, 43, 1363–1370. [Google Scholar] [CrossRef] [Green Version]
- Kreisel, D.; Krupnick, A.S.; Gelman, A.E.; Engels, F.H.; Popma, S.H.; Krasinskas, A.M.; Balsara, K.R.; Szeto, W.Y.; Turka, L.A.; Rosengard, B.R. Non-hematopoietic allograft cells directly activate CD8+ T cells and trigger acute rejection: An alternative mechanism of allorecognition. Nat. Med. 2002, 8, 233–239. [Google Scholar] [CrossRef]
- Pratschke, J.; Wilhelm, M.J.; Laskowski, I.; Kusaka, M.; Beato, F.; Tullius, S.G.; Neuhaus, P.; Hancock, W.W.; Tilney, N.L. Influence of donor brain death on chronic rejection of renal transplants in rats. J. Am. Soc. Nephrol. 2001, 12, 2474–2481. [Google Scholar]
- Guillén-Gómez, E.; Guirado, L.; Belmonte, X.; Maderuelo, A.; Santín, S.; Juarez, C.; Ars, E.; Facundo, C.; Ballarín, J.A.; Vidal, S.; et al. Monocyte implication in renal allograft dysfunction. Clin. Exp. Immunol. 2014, 175, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Magil, A.B. Monocytes/macrophages in renal allograft rejection. Transpl. Rev. (Orlando) 2009, 23, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Serón, D.; O’Valle, F.; Moreso, F.; Gomà, M.; Hueso, M.; Grinyó, J.M.; Garcia del Moral, R. Immunophenotype of infiltrating cells in protocol renal allograft biopsies from tacrolimus-versus cyclosporine-treated patients. Transplantation 2007, 83, 649–652. [Google Scholar] [CrossRef] [PubMed]
- Guillén-Gómez, E.; Dasilva, I.; Silva, I.; Arce, Y.; Facundo, C.; Ars, E.; Breda, A.; Ortiz, A.; Guirado, L.; Ballarín, J.A.; et al. Early Macrophage Infiltration and Sustained Inflammation in Kidneys From Deceased Donors Are Associated With Long-Term Renal Function. Am. J. Transpl. 2017, 17, 733–743. [Google Scholar] [CrossRef]
- Csoka, B.; Selmeczy, Z.; Koscso, B.; Nemeth, Z.H.; Pacher, P.; Murray, P.J.; Kepka-Lenhart, D.; Morris, S.M.; Gause, W.C.; Leibovich, S.J.; et al. Adenosine promotes alternative macrophage activation via A2A and A2B receptors. FASEB J. 2012, 26, 376–386. [Google Scholar] [CrossRef] [Green Version]
- Pinhal-Enfield, G.; Ramanathan, M.; Hasko, G.; Vogel, S.N.; Salzman, A.L.; Boons, G.-J.; Leibovich, S.J. An Angiogenic Switch in Macrophages Involving Synergy between Toll-Like Receptors 2, 4, 7, and 9 and Adenosine A2A Receptors. Am. J. Pathol. 2003, 163, 711–721. [Google Scholar] [CrossRef] [Green Version]
- Kurashima, Y.; Amiya, T.; Nochi, T.; Fujisawa, K.; Haraguchi, T.; Iba, H.; Tsutsui, H.; Sato, S.; Nakajima, S.; Iijima, H.; et al. Extracellular ATP mediates mast cell-dependent intestinal inflammation through P2 × 7 purinoceptors. Nat. Commun. 2012, 3, 1034. [Google Scholar] [CrossRef]
- Crespo Yanguas, S.; Willebrords, J.; Johnstone, S.R.; Maes, M.; Decrock, E.; De Bock, M.; Leybaert, L.; Cogliati, B.; Vinken, M. Pannexin1 as mediator of inflammation and cell death. Biochim. Biophys. Acta Mol. Cell Res. 2017, 1864, 51–61. [Google Scholar] [CrossRef]
- Hanner, F.; Lam, L.; Nguyen, M.T.X.; Yu, A.; Peti-Peterdi, J. Intrarenal localization of the plasma membrane ATP channel pannexin1. Am. J. Physiol. Ren. Physiol. 2012, 303. [Google Scholar] [CrossRef] [Green Version]
- Jankowski, J.; Perry, H.M.; Medina, C.B.; Huang, L.; Yao, J.; Bajwa, A.; Lorenz, U.M.; Rosin, D.L.; Ravichandran, K.S.; Isakson, B.E.; et al. Epithelial and Endothelial Pannexin1 Channels Mediate AKI. J. Am. Soc. Nephrol. 2018, 29, 1887–1899. [Google Scholar] [CrossRef] [Green Version]
- Cekic, C.; Linden, J. Purinergic regulation of the immune system. Nat. Rev. Immunol. 2016, 16, 177–192. [Google Scholar] [CrossRef] [PubMed]
- Pastor-Anglada, M.; Pérez-Torras, S. Who is who in Adenosine transport. Front. Pharmacol. 2018, 9, 627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, G.E.; Truong, L.D.; Li, P.; Zhang, P.; Du, J.; Chen, J.-F.; Feng, L. Adenosine A2A receptor activation and macrophage-mediated experimental glomerulonephritis. FASEB J. 2008, 22, 445–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khoa, N.D.; Montesinos, M.C.; Reiss, A.B.; Delano, D.; Awadallah, N.; Cronstein, B.N. Inflammatory cytokines regulate function and expression of adenosine A(2A) receptors in human monocytic THP-1 cells. J. Immunol. 2001, 167, 4026–4032. [Google Scholar] [CrossRef] [Green Version]
- Koo, D.D.H.; Welsh, K.I.; McLaren, A.J.; Roake, J.A.; Morris, P.J.; Fuggle, S.V. Cadaver versus living donor kidneys: Impact of donor factors on antigen induction before transplantation. Kidney Int. 1999, 56, 1551–1559. [Google Scholar] [CrossRef] [Green Version]
- Bours, M.J.L.; Swennen, E.L.R.; Di Virgilio, F.; Cronstein, B.N.; Dagnelie, P.C. Adenosine 5???-triphosphate and adenosine as endogenous signaling molecules in immunity and inflammation. Pharm. Ther. 2006, 112, 358–404. [Google Scholar] [CrossRef]
- Lawrence, T.; Willoughby, D.A.; Gilroy, D.W. Anti-inflammatory lipid mediators and insights into the resolution of inflammation. Nat. Rev. Immunol. 2002, 2, 787–795. [Google Scholar] [CrossRef]
- Haskó, G.; Pacher, P.; Deitch, E.A.; Vizi, E.S. Shaping of monocyte and macrophage function by adenosine receptors. Pharm. Ther. 2007, 113, 264–275. [Google Scholar] [CrossRef] [Green Version]
- Haskó, G.; Kuhel, D.G.; Chen, J.F.; Schwarzschild, M.A.; Deitch, E.A.; Mabley, J.G.; Marton, A.; Szabó, C. Adenosine inhibits IL-12 and TNF-[alpha] production via adenosine A2a receptor-dependent and independent mechanisms. FASEB J. 2000, 14, 2065–2074. [Google Scholar] [CrossRef] [Green Version]
- Buenestado, A.; Grassin Delyle, S.; Arnould, I.; Besnard, F.; Naline, E.; Blouquit-Laye, S.; Chapelier, A.; Bellamy, J.F.; Devillier, P. The role of adenosine receptors in regulating production of tumour necrosis factor-alpha and chemokines by human lung macrophages. Br. J. Pharm. 2010, 159, 1304–1311. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.B.; Kalluri, R. Mechanistic connection between inflammation and fibrosis. Kidney Int. Suppl. 2010, 78, S22–S26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morello, S.; Ito, K.; Yamamura, S.; Lee, K.-Y.; Jazrawi, E.; DeSouza, P.; Barnes, P.; Cicala, C.; Adcock, I.M. IL-1 and TNF- Regulation of the Adenosine Receptor (A2A) Expression: Differential Requirement for NF- B Binding to the Proximal Promoter. J. Immunol. 2006, 177, 7173–7183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Capecchi, P.L.; Camurri, A.; Pompella, G.; Mazzola, A.; Maccherini, M.; Diciolla, F.; Lazzerini, P.E.; Abbracchio, M.P.; Laghi-Pasini, F. Upregulation of A2A adenosine receptor expression by TNF-alpha in PBMC of patients with CHF: A regulatory mechanism of inflammation. J. Card. Fail. 2005, 11, 67–73. [Google Scholar] [CrossRef] [PubMed]
- St Hilaire, C.; Carroll, S.H.; Chen, H.; Ravid, K. Mechanisms of induction of adenosine receptor genes and its functional significance. J. Cell. Physiol. 2009, 218, 35–44. [Google Scholar] [CrossRef] [Green Version]
- Khoa, N.D.; Postow, M.; Danielsson, J.; Cronstein, B.N. Tumor necrosis factor-alpha prevents desensitization of Galphas-coupled receptors by regulating GRK2 association with the plasma membrane. Mol. Pharm. 2006, 69, 1311–1319. [Google Scholar] [CrossRef] [PubMed]
- Bani-Hani, A.H.; Campbell, M.T.; Meldrum, D.R.; Meldrum, K.K. Cytokines in epithelial-mesenchymal transition: A new insight into obstructive nephropathy. J. Urol. 2008, 180, 461–468. [Google Scholar] [CrossRef]
- Bates, R.C.; Mercurio, A.M. Tumor necrosis factor-alpha stimulates the epithelial-to-mesenchymal transition of human colonic organoids. Mol. Biol. Cell 2003, 14, 1790–1800. [Google Scholar] [CrossRef] [Green Version]
- Meldrum, K.K.; Misseri, R.; Metcalfe, P.; Dinarello, C.A.; Hile, K.L.; Meldrum, D.R. TNF-alpha neutralization ameliorates obstruction-induced renal fibrosis and dysfunction. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R1456–R1464. [Google Scholar] [CrossRef]
- Therrien, F.J.F.J.; Agharazii, M.; Lebel, M.; Larivière, R.; Larivire, R. Neutralization of tumor necrosis factor-alpha reduces renal fibrosis and hypertension in rats with renal failure. Am. J. Nephrol. 2012, 36, 151–161. [Google Scholar] [CrossRef]
- Haskó, G.; Pacher, P. Regulation of macrophage function by adenosine. Arter. Thromb. Vasc. Biol. 2012, 32, 865–869. [Google Scholar] [CrossRef] [Green Version]
- Garcia, G.E.; Truong, L.D.; Chen, J.-F.; Johnson, R.J.; Feng, L. Adenosine A(2A) receptor activation prevents progressive kidney fibrosis in a model of immune-associated chronic inflammation. Kidney Int. 2011, 80, 378–388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Veledo, S.; Huber-Ruano, I.; Aymerich, I.; Duflot, S.; Casado, F.J.; Pastor-Anglada, M. Bile acids alter the subcellular localization of CNT2 (concentrative nucleoside cotransporter) and increase CNT2-related transport activity in liver parenchymal cells. Biochem. J. 2006, 395, 337–344. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Torras, S.; Iglesias, I.; Llopis, M.; Lozano, J.J.; Antolín, M.; Guarner, F.; Pastor-Anglada, M. Transportome Profiling Identifies Profound Alterations in Crohn’s Disease Partially Restored by Commensal Bacteria. J. Crohn’s Colitis 2016, 10, 850–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Medina-Pulido, L.; Molina-Arcas, M.; Justicia, C.; Soriano, E.; Burgaya, F.; Planas, A.M.; Pastor-Anglada, M. Hypoxia and P1 receptor activation regulate the high-affinity concentrative adenosine transporter CNT2 in differentiated neuronal PC12 cells. Biochem. J. 2013, 454, 437–445. [Google Scholar] [CrossRef]
- Ohta, A.; Sitkovsky, M. Role of G-protein-coupled adenosine receptors in downregulation of inflammation and protection from tissue damage. Nature 2001, 414, 916–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sitkovsky, M.V.; Lukashev, D.; Apasov, S.; Kojima, H.; Koshiba, M.; Caldwell, C.; Ohta, A.; Thiel, M. Physiological control of immune response and inflammatory tissue damage by hypoxia-inducible factors and adenosine A2A receptors. Annu. Rev. Immunol. 2004, 22, 657–682. [Google Scholar] [CrossRef]
- Thiel, M.; Chouker, A.; Ohta, A.; Jackson, E.; Caldwell, C.; Smith, P.; Lukashev, D.; Bittmann, I.; Sitkovsky, M. V Oxygenation inhibits the physiological tissue-protecting mechanism and thereby exacerbates acute inflammatory lung injury. PLoS Biol. 2005, 3, e174. [Google Scholar] [CrossRef] [Green Version]
- Beavis, P.A.; Stagg, J.; Darcy, P.K.; Smyth, M.J. CD73: A potent suppressor of antitumor immune responses. Trends Immunol. 2012, 33, 231–237. [Google Scholar] [CrossRef]
- Regateiro, F.S.; Howie, D.; Nolan, K.F.; Agorogiannis, E.I.; Greaves, D.R.; Cobbold, S.P.; Waldmann, H. Generation of anti-inflammatory adenosine by leukocytes is regulated by TGF-β. Eur. J. Immunol. 2011, 41, 2955–2965. [Google Scholar] [CrossRef]
- Zanin, R.F.; Braganhol, E.; Bergamin, L.S.L.S.; Campesato, L.F.I.L.F.I.; Filho, A.Z.; Moreira, J.C.F.J.C.F.; Morrone, F.B.; Sévigny, J.; Schetinger, M.R.C.; de Souza Wyse, A.T.; et al. Differential macrophage activation alters the expression profile of NTPDase and Ecto-5’-nucleotidase. PLoS ONE 2012, 7, e31205. [Google Scholar] [CrossRef] [Green Version]
- Kalsi, K.; Lawson, C.; Dominguez, M.; Taylor, P.; Yacoub, M.H.; Smolenski, R.T. Regulation of ecto-5′-nucleotidase by TNF-alpha in human endothelial cells. Mol. Cell. Biochem. 2002, 232, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Blume, C.; Felix, A.; Shushakova, N.; Gueler, F.; Falk, C.S.; Haller, H.; Schrader, J. Autoimmunity in CD73/Ecto-5′-nucleotidase deficient mice induces renal injury. PLoS ONE 2012, 7, e37100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, F.; Dahl, G. A permeant regulating its permeation pore: Inhibition of pannexin 1 channels by ATP. Am. J. Physiol. Cell Physiol. 2009, 296, C250. [Google Scholar] [CrossRef] [PubMed]
- Whyte-Fagundes, P.; Zoidl, G. Mechanisms of pannexin1 channel gating and regulation. Biochim. Biophys. Acta Biomembr. 2018, 1860, 65–71. [Google Scholar] [CrossRef]
- Barroso, M.; Kao, D.; Blom, H.J.; Tavares de Almeida, I.; Castro, R.; Loscalzo, J.; Handy, D.E. S-adenosylhomocysteine induces inflammation through NFkB: A possible role for EZH2 in endothelial cell activation. Biochim. Biophys. Acta Mol. Basis Dis. 2016, 1862, 82–92. [Google Scholar] [CrossRef] [Green Version]
- Ferrante, C.J.; Pinhal-Enfield, G.; Elson, G.; Cronstein, B.N.; Hasko, G.; Outram, S.; Leibovich, S.J. The Adenosine-Dependent Angiogenic Switch of Macrophages to an M2-Like Phenotype is Independent of Interleukin-4 Receptor Alpha (IL-4Rα) Signaling. Inflammation 2013, 36, 921–931. [Google Scholar] [CrossRef]
- Montesinos, M.C.; Gadangi, P.; Longaker, M.; Sung, J.; Levine, J.; Nilsen, D.; Reibman, J.; Li, M.; Jiang, C.K.; Hirschhorn, R.; et al. Wound healing is accelerated by agonists of adenosine A2 (G alpha s-linked) receptors. J. Exp. Med. 1997, 186, 1615–1620. [Google Scholar] [CrossRef]
- Leibovich, S.J.; Chen, J.-F.; Pinhal-Enfield, G.; Belem, P.C.; Elson, G.; Rosania, A.; Ramanathan, M.; Montesinos, C.; Jacobson, M.; Schwarzschild, M.A.; et al. Synergistic up-regulation of vascular endothelial growth factor expression in murine macrophages by adenosine A(2A) receptor agonists and endotoxin. Am. J. Pathol. 2002, 160, 2231–2244. [Google Scholar] [CrossRef] [Green Version]
- Olah, M.E.; Caldwell, C.C. Adenosine receptors and mammalian toll-like receptors: Synergism in macrophages. Mol. Interv. 2003, 3, 370–374. [Google Scholar] [CrossRef]
- Csóka, B.; Németh, Z.H.; Virág, L.; Gergely, P.; Leibovich, S.J.; Pacher, P.; Sun, C.-X.X.; Blackburn, M.R.; Vizi, E.S.; Deitch, E.A.; et al. A2A adenosine receptors and C/EBP?? are crucially required for IL-10 production by macrophages exposed to Escherichia coli. Blood 2007, 110, 2685–2695. [Google Scholar] [CrossRef] [Green Version]
- Ruffell, D.; Mourkioti, F.; Gambardella, A.; Kirstetter, P.; Lopez, R.G.; Rosenthal, N.; Nerlov, C. A CREB-C/EBPbeta cascade induces M2 macrophage-specific gene expression and promotes muscle injury repair. Proc. Natl. Acad. Sci. USA 2009, 106, 17475–17480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Natsuka, S.; Akira, S.; Nishio, Y.; Hashimoto, S.; Sugita, T.; Isshiki, H.; Kishimoto, T. Macrophage differentiation-specific expression of NF-IL6, a transcription factor for interleukin-6. Blood 1992, 79, 460–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, B.; Ullenbruch, M.R.; Jin, H.; Gharaee-Kermani, M.; Phan, S.H. An essential role for CCAAT/enhancer binding protein beta in bleomycin-induced pulmonary fibrosis. J. Pathol. 2007, 211, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Wu, Z.; Jin, H.; Hashimoto, N.; Liu, T.; Phan, S.H. CCAAT/enhancer-binding protein beta isoforms and the regulation of alpha-smooth muscle actin gene expression by IL-1 beta. J. Immunol. 2004, 173, 4661–4668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
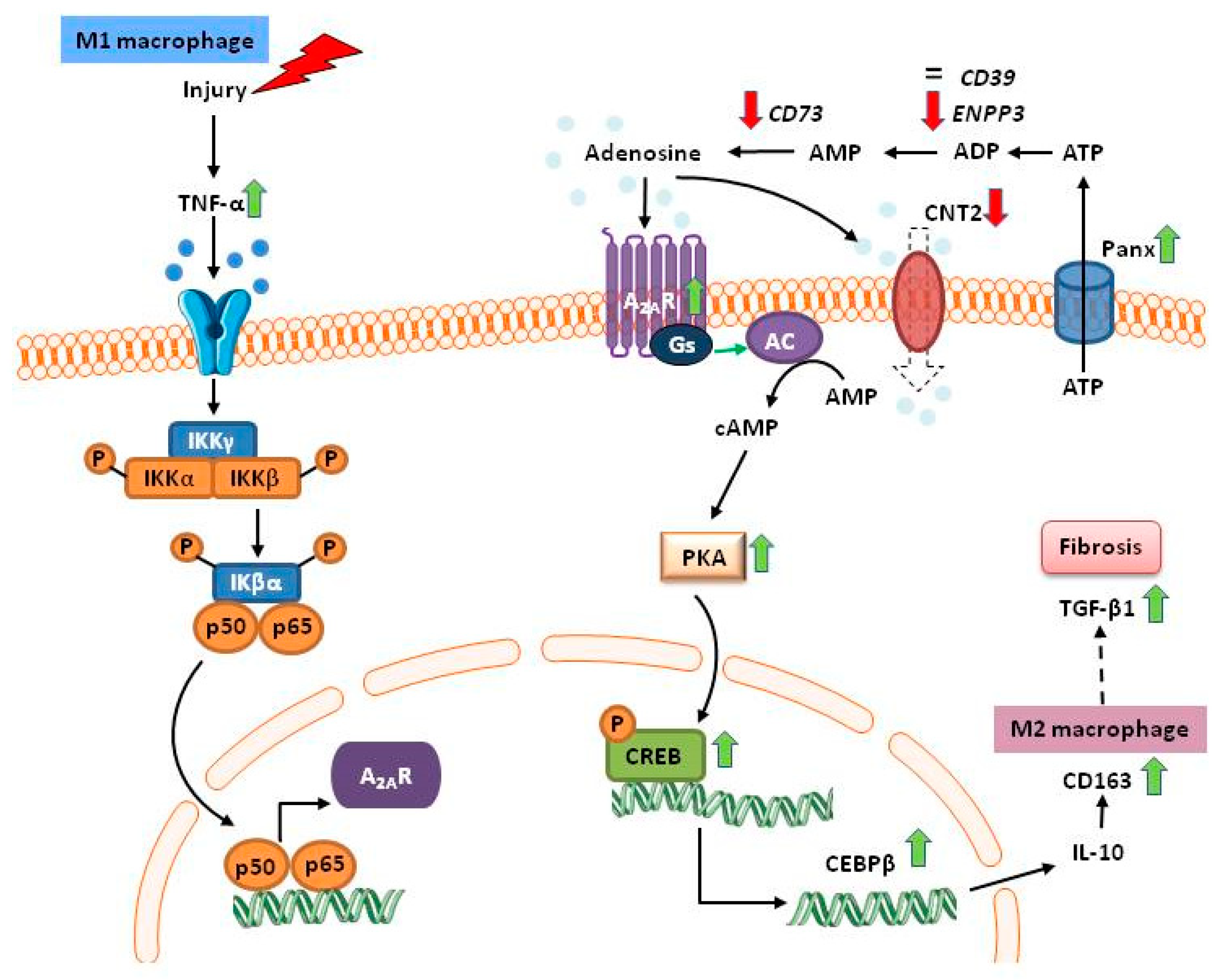{kind=link}
| A2AR Correlations | |||||
|---|---|---|---|---|---|
| DD | LD | ||||
| p | Rho Spearman | p | Rho Spearman | ||
| Enzymes | CD39 | 0.065 | 0.272 | 0.311 | 0.246 |
| CD73 | <0.001 | 0.635 | 0.705 | −0.093 | |
| ADK | <0.001 | 0.609 | 0.433 | 0.191 | |
| SAHH | <0.001 | 0.501 | 0.697 | 0.096 | |
| ADCY | <0.001 | 0.521 | 0.271 | 0.266 | |
| Nucleoside transporters | ENT1 | <0.001 | 0.603 | 0.170 | 0.328 |
| ENT2 | 0.003 | 0.426 | 0.507 | 0.162 | |
| ENT3 | <0.001 | 0.563 | 0.673 | 0.104 | |
| ENT4 | 0.002 | 0.441 | 0.459 | 0.181 | |
| CNT1 | 0.049 | 0.288 | 0.792 | 0.065 | |
| CNT2 | 0.017 | 0.347 | 0.665 | −0.106 | |
| A2AR Correlations | |||||||
|---|---|---|---|---|---|---|---|
| RNA Expression | DD | LD | |||||
| DD vs. LD | Significance | p | Rho Spearman | p | Rho Spearman | ||
| Inflammatory & M1 markers | TNF | ↑ | [14] * | <0.001 | 0.504 | 0.022 | 0.520 |
| NFKB | ↑ | p = 0.033 | <0.001 | 0.626 | 0.997 | 0.001 | |
| CD16 | ↑ | [14] * | <0.001 | 0.507 | 0.131 | 0.359 | |
| CD86 | = | [14] * | <0.001 | 0.478 | 0.303 | 0.250 | |
| IL-1β | ↑ | [14] * | 0.003 | 0.432 | 0.983 | 0.005 | |
| Anti-inflammatory & M2 markers | IL10 | = | n.s. | <0.001 | 0.492 | 0.265 | 0.269 |
| CD206 | = | [14] * | <0.001 | 0.577 | 0.132 | 0.358 | |
| CD163 | ↑ | [14] * | <0.001 | 0.502 | 0.209 | 0.302 | |
| IL13RA2 | = | n.s. | 0.002 | 0.437 | 0.204 | 0.305 | |
| CD209 | ↑ | [14] * | 0.056 | 0.281 | 0.557 | 0.144 | |
| CEBPB | ↑ | p < 0.001 | <0.001 | 0.482 | 0.446 | 0.186 | |
| EMT & Fibrosis markers | TGFB1 | ↑ | [14] * | <0.001 | 0.712 | 0.061 | 0.437 |
| FIBRONECTIN | = | [14] * | <0.001 | 0.652 | 0.545 | 0.148 | |
| ACTA2 | = | [14] * | <0.001 | 0.572 | 0.123 | 0.366 | |
| VIMENTIN | = | [14] * | 0.002 | 0.447 | 0.718 | 0.089 | |
| COL1A1 | ↑ | p = 0.006 | 0.005 | 0.407 | 0.102 | 0.386 | |
| COL1A2 | = | n.s. | <0.001 | 0.511 | 0.158 | 0.337 | |
| COL3A1 | = | n.s. | 0.015 | 0.354 | 0.051 | 0.454 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guillén-Gómez, E.; Silva, I.; Serra, N.; Caballero, F.; Leal, J.; Breda, A.; San Martín, R.; Pastor-Anglada, M.; Ballarín, J.A.; Guirado, L.; et al. From Inflammation to the Onset of Fibrosis through A2A Receptors in Kidneys from Deceased Donors. Int. J. Mol. Sci. 2020, 21, 8826. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228826
Guillén-Gómez E, Silva I, Serra N, Caballero F, Leal J, Breda A, San Martín R, Pastor-Anglada M, Ballarín JA, Guirado L, et al. From Inflammation to the Onset of Fibrosis through A2A Receptors in Kidneys from Deceased Donors. International Journal of Molecular Sciences. 2020; 21(22):8826. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228826
Chicago/Turabian StyleGuillén-Gómez, Elena, Irene Silva, Núria Serra, Francisco Caballero, Jesús Leal, Alberto Breda, Rody San Martín, Marçal Pastor-Anglada, José A. Ballarín, Lluís Guirado, and et al. 2020. "From Inflammation to the Onset of Fibrosis through A2A Receptors in Kidneys from Deceased Donors" International Journal of Molecular Sciences 21, no. 22: 8826. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21228826