Proregenerative Activity of IL-33 in Gastric Tissue Cells Undergoing Helicobacter Pylori-Induced Apoptosis


Abstract
:1. Introduction
2. Results
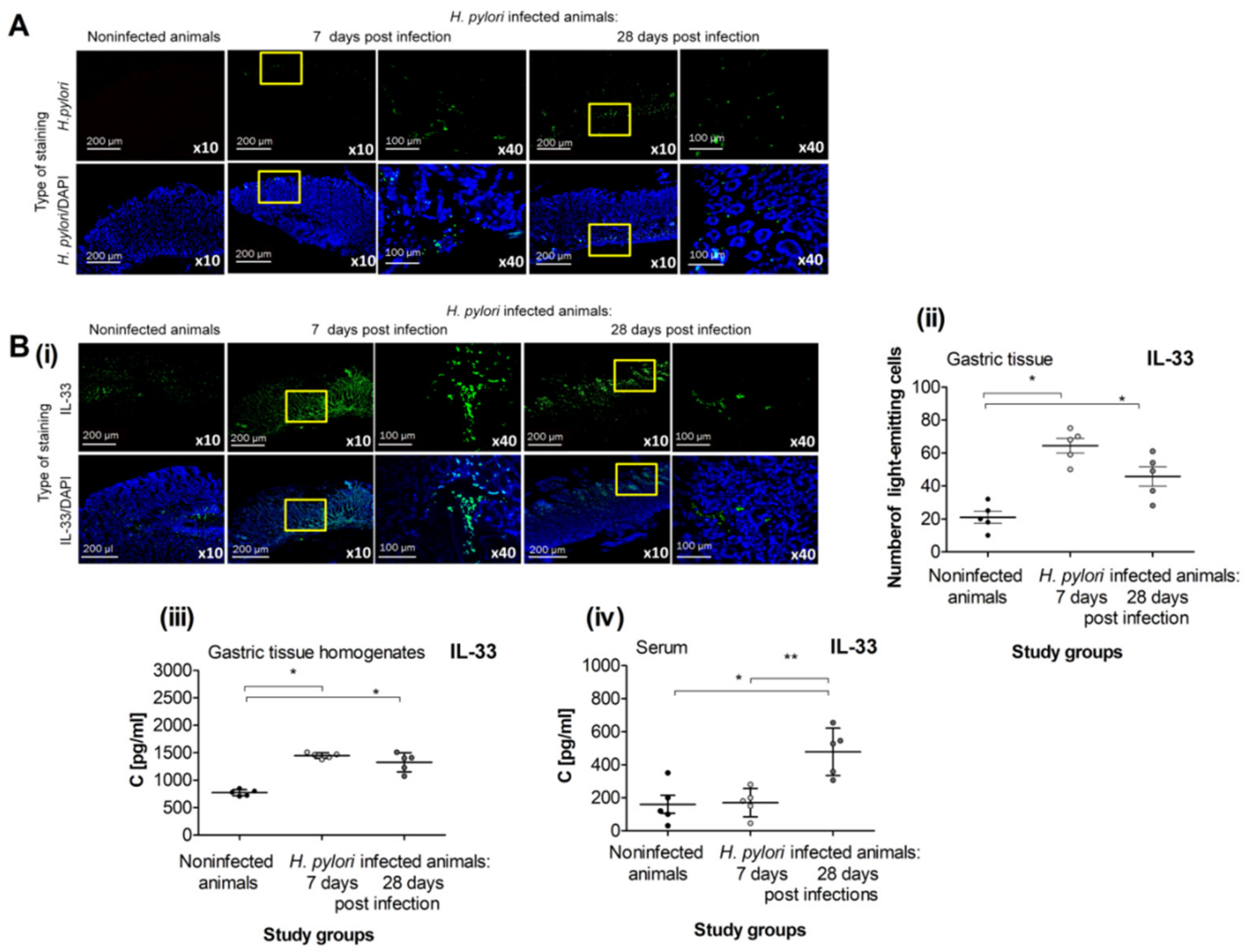
2.1. Production of IL-33 in Caviae porcellus colonized with H. pylori
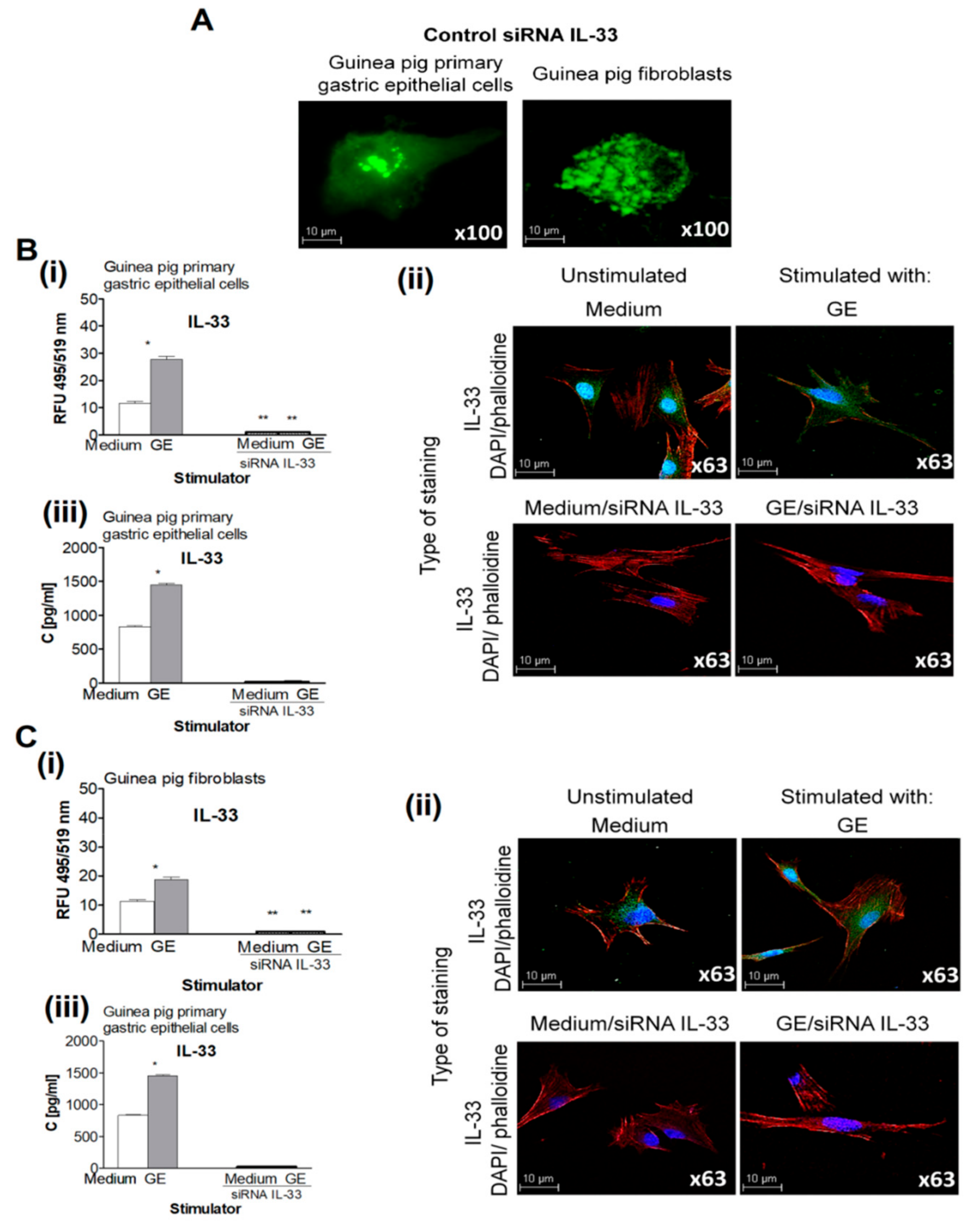
2.2. IL-33 Production in Cell Cultures Exposed to H. pylori Components
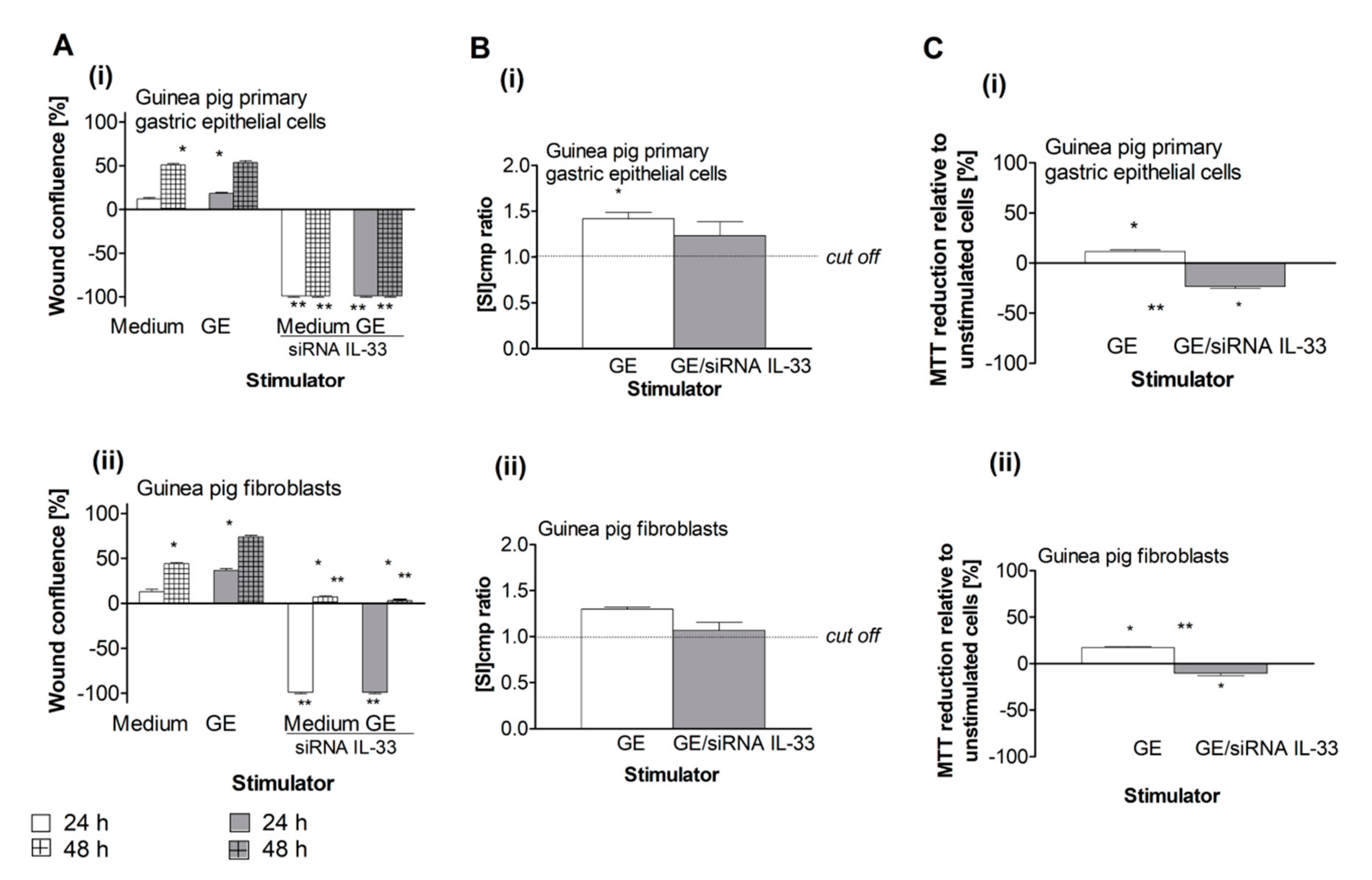
2.3. Pro-Regenerative Activity of IL-33 in the Cell Cultures Exposed to H. pylori Components
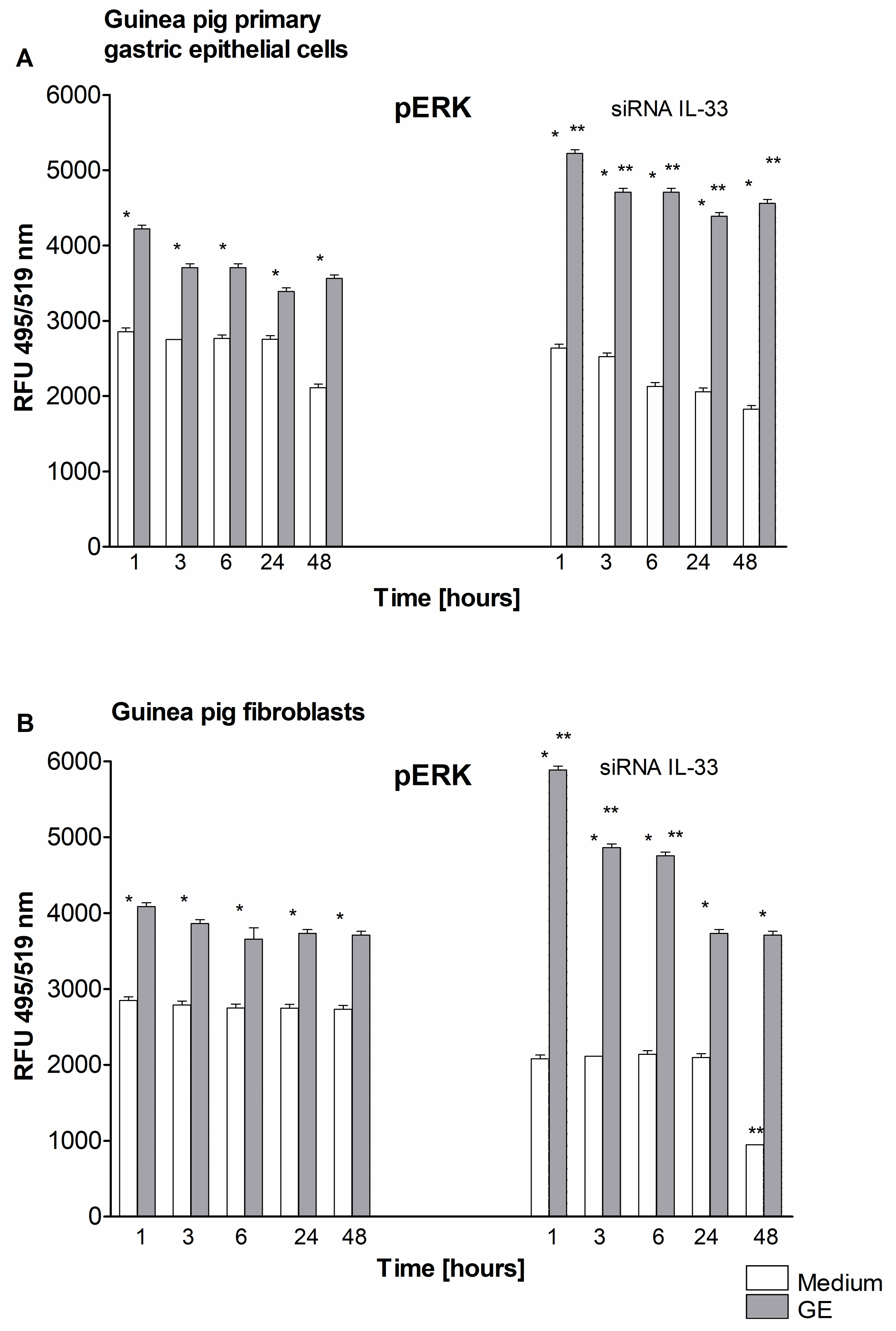
2.4. Control of H. pylori-Induced Erk Activation by IL-33
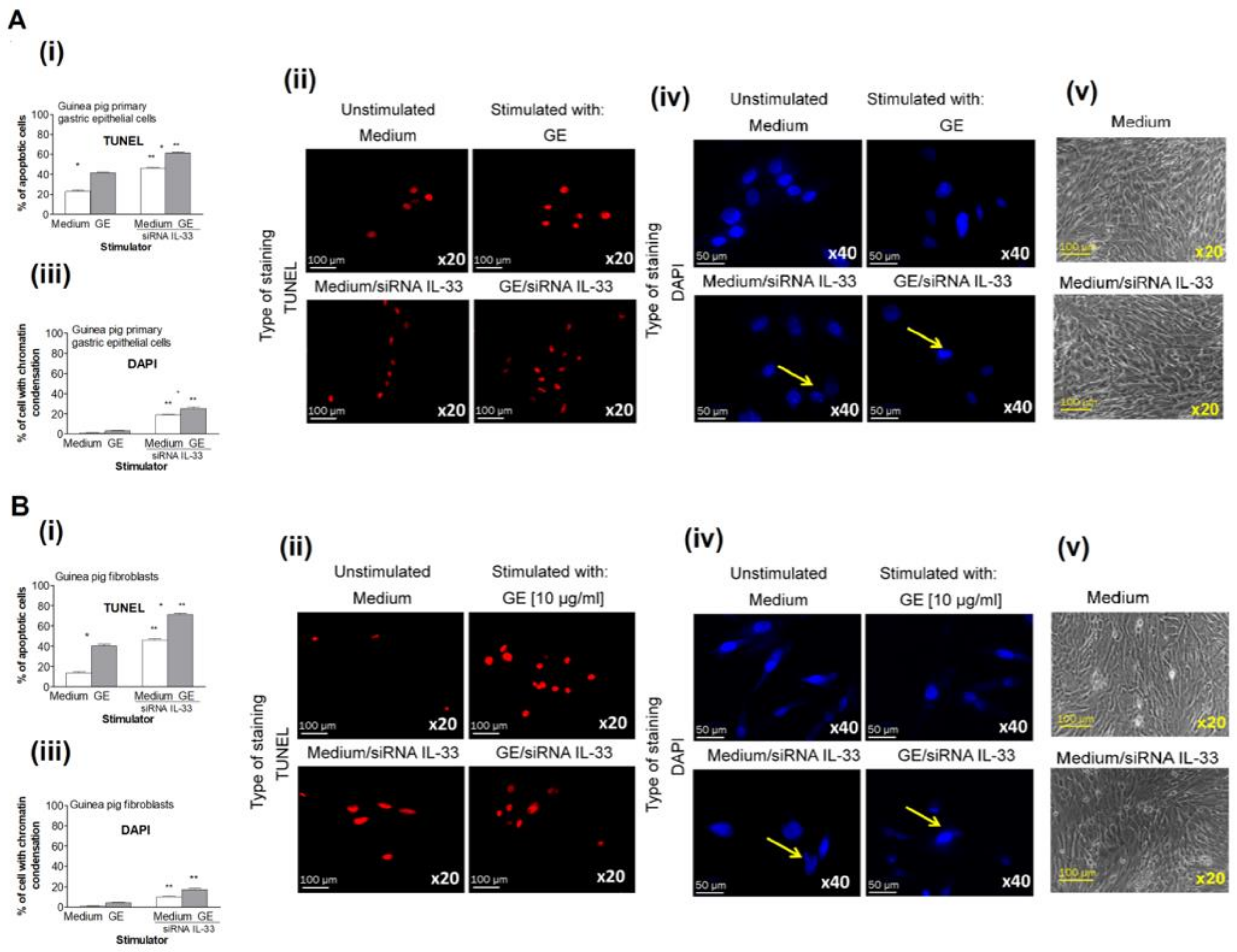
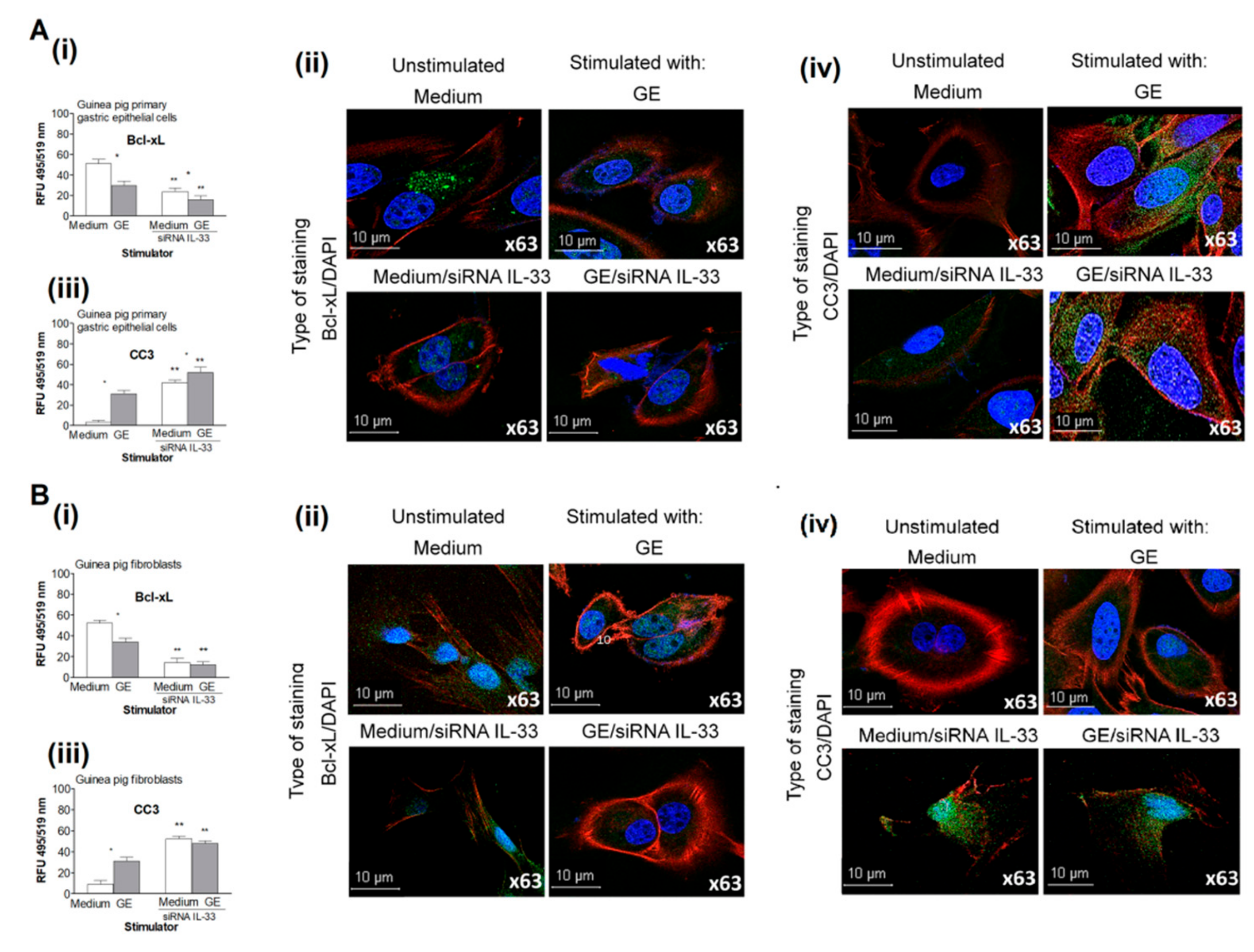
2.5. Regulation of Cell Apoptosis by IL-33
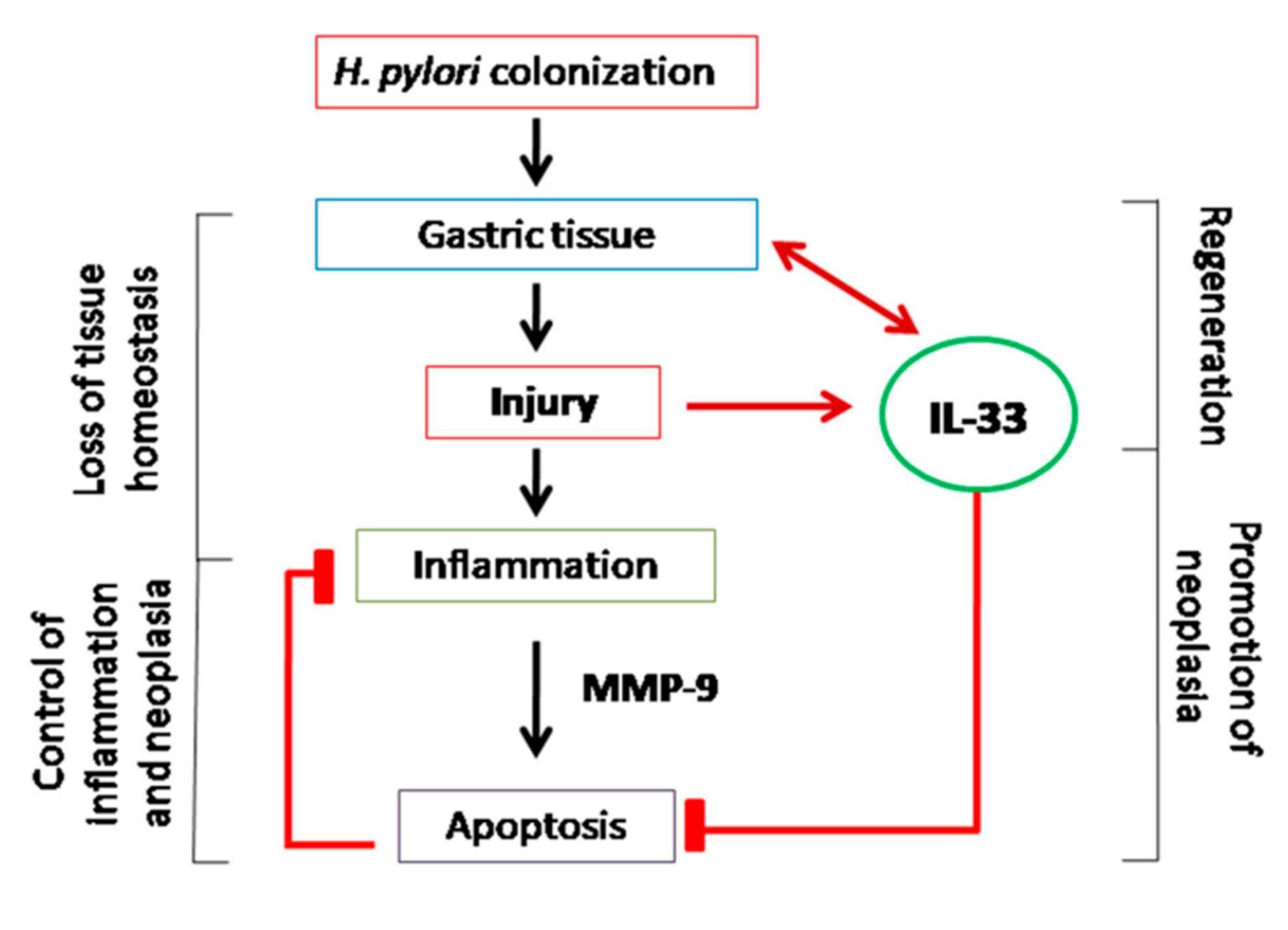
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. H. pylori Infection in Caviae porcellus (Guinea Pigs)
4.3. Cell Cultures
4.4. Bacterial Stimuli
4.5. IL-33 siRNA Silencing
4.6. Evaluation of Cell Migration in a Scratch Wound Healing Assay
4.7. Cell Viability Assays
4.8. Cell Apoptosis
4.9. Erk Activation/Phosphorylation (pErk)
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Bcl-xL | B cell lymphoma-extra large |
| BSA | Bovine serum albumin |
| CagA | Cytotoxin-associated gene A |
| CC3 | Cleaved caspase 3 |
| cRPMI | Complete RPMI |
| dsDNA | Double-stranded DNA |
| DAMPs | Damage-associated molecular patterns |
| DAPI | 4′,6-diamino-2-phenylindole |
| ELISA | Enzyme-linked immunosorbent assay |
| FCS | Featal calf serum |
| FPRs | Formyl peptide receptors |
| GE | Glycine acid extract |
| GGT | Gamma-glutamyl transpeptidase |
| HEPES | N-2-hydroxyethylpiperazine-N-2-ethane sulfonicacid |
| HIF | Hypoxia inducible factor |
| HLO | Helicobacter like organism |
| HMGB-1 | High motility group box-1 protein |
| HP-NAP | Neutrophil-activating factor |
| Hsp | Heat shock protein |
| HtrA | High temperature requirement A protein |
| IL | Interleukin |
| ILC2 | Group 2 innate lymphoid cell |
| Le | Lewis |
| LPS | Lipopolysaccharide |
| MALT | Mucosa-associated lymphoid tissue |
| MMP | Metalloproteinase |
| MUC5AC | Mucin 5 |
| MTT | 3-4,5-dimethylthiazole-2-yl]-2,5-diphenyltetrazolium bromide |
| NFkB | Nuclear factor B |
| NLRs | Nod-like receptors |
| PCR | Polimerase chain reaction |
| pERK | Activation/phosphorylation of extracellular signal-regulated kinase |
| PPRs | Pathogen recognition receptors |
| RFU | Relative fluorescence units |
| SI | Stimulation index |
| siRNA | Small interfering RNA |
| ST2 | Secretory receptor |
| sTNF-Rs | Soluble TNF receptors |
| TH2 | T helper 2 |
| TLRs | Toll-like receptors |
| TNF | Tumor necrosis factor |
| TUNEL | Terminal deoxynucleotidyl transferase (TdT)-mediated dUTP nick end labeling |
| VacA | Vacuolating cytotoxin A |
| VEGF | Vascular endothelial growth factor |
References
- Tarnawski, A. Cellular and molecular mechanisms of mucosal defense and repair. In Bioregulation and Its Disorders in Gastrointestinal, Tract ed.; Blackwell Science: Japan, Tokyo, 1998; Volume 3, p. 17. [Google Scholar]
- Terrés, A.M.; Pajares, J.M.; Hopkins, A.M.; Murphy, A.; Moran, A.; Baird, A.W.; Kelleher, D. Helicobacter pylori disrupts epithelial barrier function in a process inhibited by protein kinase C activators. Infect. Immun. 1998, 66, 2943–2950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tarnawski, A.S.; Ahluwalia, A.; Jones, M.K. The mechanisms of gastric mucosal injury: Focus on microvascular endothelium as a key target. Curr. Med. Chem. 2012, 19, 4–15. [Google Scholar] [CrossRef] [PubMed]
- Laine, L.; Takeuchi, K.; Tarnawski, A. Gastric mucosal defense and cytoprotection: Bench to bedside. Gastroenterology 2008, 135, 41–60. [Google Scholar] [CrossRef] [PubMed]
- Henriksnäs, J.; Phillipson, M.; Storm, M.; Engstrand, L.; Soleimani, M.; Holm, L. Impaired mucus-bicarbonate barrier in Helicobacter pylori-infected mice. Am. J. Physiol. Gastrointest. Liver Physiol. 2006, 291, G396–G403. [Google Scholar] [CrossRef] [Green Version]
- Goh, K.L.; Chan, W.K.; Shiota, S.; Yamaoka, Y. Epidemiology of Helicobacter pylori infection and public health implications. Helicobacter 2011, 16 (Suppl. S1), 1–9. [Google Scholar] [CrossRef] [Green Version]
- Blaser, M.J.; Atherton, J.C. Helicobacter infection persistence: Biology and disease. J. Clin. Investig. 2004, 113, 321–333. [Google Scholar] [CrossRef] [Green Version]
- Chmiela, M.; Michetti, P. Inflammation, immunity, vaccines for Helicobacter infection. Helicobacter 2006, 11 (Suppl. S1), 21–26. [Google Scholar] [CrossRef]
- . Peek, R.M.; Crabtree, J.M.H. H. pylori infection and gastric neoplasia. J. Pathol. 2006, 208, 233–248. [Google Scholar] [CrossRef]
- Posselt, G.; Backert, S.; Wessler, S. The functional interplay of H. pylori factors with gastric epithelial cells induces a multi-step process in pathogenesis. Cell Commun. Signal 2013, 11, 77. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, N.; Murata-Kamiya, N.; Yanagiya, K.; Suda, W.; Hattori, M.; Kanda, H.; Bingo, A.; Fujii, Y.; Maeda, S.; Koike, K.; et al. Mutual reinforcement of inflammation and carcinogenesis by the H. pylori CagA oncoprotein. Sci. Rep. 2015, 5, 10024. [Google Scholar] [CrossRef] [Green Version]
- Chmiela, M.; Karwowska, Z.; Gonciarz, W.; Allushi, B.; Stączek, P. Host pathogen interactions in Helicobacter pylori related gastric cancer. World J. Gastroenterol. 2017, 23, 1521–1540. [Google Scholar] [CrossRef] [PubMed]
- Rudnicka, K.; Backert, S.; Chmiela, M. Genetic polymorphisms in inflammatory and other regulators in gastric cancer: Risks and clinical consequences. Molecular Mechanisms of Inflammation: Induction, Resolution and Escape by Helicobacter pylori. Curr. Top. Microbiol. Immunol. 2019, 421. [Google Scholar] [CrossRef]
- Kusters, J.G.; van Vliet, A.H.; Kuipers, E.J. Pathogenesis of Helicobacter pylori infection. Clin. Microbiol. Rev. 2006, 19, 449–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chmiela, M.; Kupcinskas, J. Pathogenesis of Helicobacter pylori infection. Helicobacter. 2019, 24 (Suppl. S1), e12638. [Google Scholar] [CrossRef] [Green Version]
- Lytton, S.D.; Fischer, W.; Nagel, W.; Haas, R.; Beck, F.X. Production of ammonium by Helicobacter pylori mediates occluding processing and disruption of tight junctions in Caco-2 cells. Microbiology 2005, 155, 3267–3276. [Google Scholar] [CrossRef] [Green Version]
- Backert, S.; Kwok, T.; Schmid, M.; Selbach, M.; Moese, S.; Peek, R.M., Jr.; König, W.; Meyer, T.F.; Jungblut, P.R. Subproteomes of soluble and structure-bound H. pylori proteins analyzed by two-dimensional gel electrophoresis and mass spectrometry. Proteomics 2005, 5, 1311–3345. [Google Scholar] [CrossRef]
- Cover, T.L.; Blanke, S.R.H. pylori VacA, a paradigm for toxin multifunctionality. Nat. Rev. Microbiol. 2005, 3, 320–332. [Google Scholar] [CrossRef]
- Palframan, S.L.; Kwok, T.; Gabriel, K. Vacuolating cytotoxin A (VacA), a key toxin for Helicobacter pylori. Front. Cell. Infect. Microbiol. 2012, 2, 92. [Google Scholar] [CrossRef] [Green Version]
- Yahiro, K.; Satoh, M.; Nakano, M.; Hisatsune, J.; Isomoto, H.; Sap, J.; Suzuki, H.; Nomura, F.; Noda, M.; Moss, J.; et al. Low-density lipoprotein receptor-related protein-1 (LRP-1) mediates autophagy and apoptosis caused by H. pylori VacA. J. Biol. Chem. 2012, 287, 31104–31115. [Google Scholar] [CrossRef] [Green Version]
- Hoy, B.; Löwer, M.; Weydig, C.; Carra, G.; Tegtmeyer, N.; Geppert, T.; Schröder, P.; Sewald, N.; Backert, S.; Schneider, G.; et al. H. pylori Htr is a new secreted virulence factor that cleaves E-cadherin to disrupt intercellular adhesion. EMBO Rep. 2010, 11, 798–804. [Google Scholar] [CrossRef] [Green Version]
- Muotiala, A.; Helander, I.M.; Pyhälä, L.; Kosunen, T.U.; Moran, A.P. Low biological activity of Helicobacter pylori lipopolysaccharide. Infect. Immun. 1992, 60, 1714–1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, A.P.; Helander, I.M.; Kosunen, T.U. Compositional analysis of Helicobacter pylori rough-form lipopolysaccharides. J. Bacteriol. 1992, 174, 1370–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moran, A.P.; Aspinall, G.O. Unique structural and biological features of Helicobacter pylori lipopolysaccharides. Prog. Clin. Biol. Res. 1998, 397, 37–49. [Google Scholar] [PubMed]
- Appelmelk, B.J.; Simoons-Smit, I.; Negrini, R.; Moran, A.P.; Aspinall, G.O.; Forte, J.G.; DeVries, T.; Quan, H.; Verboom, T.; Maaskant, J.J.; et al. Potential role of molecular mimicry between Helicobacter pylori lipopolysaccharide and host Lewis blood group antigens in autoimmunity. Infect. Immun. 1996, 64, 2031–2040. [Google Scholar] [CrossRef] [Green Version]
- Paziak-Domańska, B.; Chmiela, M.; Jarosińska, A.; Rudnicka, W. Potential role of CagA in the inhibition of T cell reactivity in Helicobacter pylori infection. Cell. Immunol. 2000, 202, 136–139. [Google Scholar] [CrossRef]
- Grebowska, A.; Moran, A.P.; Bielanski, W.; Matusiak, A.; Rechcinski, T.; Rudnicka, K.; Szczęsna, E.; Walencka, M.; Rudnicka, W.; Chmiela, M. Helicobacter pylori lipopolysaccharide activity in human peripheral blood mononuclear leukocyte cultures. J. Physiol. Pharmacol. 2010, 61, 437–442. [Google Scholar]
- Grebowska, A.; Moran, A.P.; Matusiak, A.; Bak-Romaniszyn, L.; Czkwianianc, E.; Rechciński, T.; Walencka, M.; Płaneta-Małecka, I.; Rudnicka, W.; Chmiela, M. Anti-phagocytic activity of Helicobacter pylori lipopolysaccharide (LPS)-possible modulation of the innate immune response to these bacteria. Pol. J. Microbiol. 2008, 57, 185–192. [Google Scholar]
- Chmiela, M.; Miszczyk, E.; Rudnicka, K. Structural modifications of Helicobacter pylori lipopolysaccharide: An idea for how to live in peace. World J. Gastroenterol. 2014, 20, 9882–9897. [Google Scholar] [CrossRef]
- Rudnicka, K.; Miszczyk, E.; Matusiak, A.; Walencka, M.; Moran, A.P.; Rudnicka, W.; Chmiela, M. Helicobacter pylori-driven modulation of NK cell expansion, intracellular cytokine expression and cytotoxic activity. Innate Immun. 2015, 21, 127–139. [Google Scholar] [CrossRef] [Green Version]
- Miszczyk, E.; Walencka, M.; Rudnicka, K.; Matusiak, A.; Rudnicka, W.; Chmiela, M. Antigen-specific lymphocyte proliferation as a marker of immune response in guinea pigs with sustained Helicobacter pylori infection. Acta Biochim. Pol. 2014, 61, 295–303. [Google Scholar] [CrossRef]
- Mnich, E.; Gajewski, A.; Rudnicka, K.; Gonciarz, W.; Stawerski, P.; Hinc, K.; Obuchowski, M.; Chmiela, M. Immunoregulation of antigen presenting and secretory functions of monocytic cells by Helicobacter pylori antigens in relation to impairment of lymphocyte expansion. Acta Biochim. Pol. 2015, 62, 641–650. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Evans, J.E.; Rock, K.L. Molecular identification of a danger signal that alerts the immune system to dying cells. Nature 2003, 425, 516–521. [Google Scholar] [CrossRef] [PubMed]
- Kono, H.; Rock, K.L. How dying cells alert the immune system to danger. Nat. Rev. Immunol. 2008, 8, 279–298. [Google Scholar] [CrossRef] [PubMed]
- Roussel, L.; Erard, M.; Cayrol, C.; Girard, J.P. Molecular mimicry between IL-33 and KSHV for attachment to chromatin through the H2A-H2B acidic pocket. EMBO Rep. 2008, 9, 1006–1012. [Google Scholar] [CrossRef] [Green Version]
- Lűthi, A.U.; Cullen, S.P.; McNeela, E.A.; Duriez, O.P.J.; Afonina, I.S.; Sheridan, C.; Brumatti, G.; Taylor, R.C.; Kersse, K.; Vandenabeele, P.; et al. Suppression of interleukin-33 bioactivity through proteolysis by apoptotic caspases. Immunity 2009, 31, 84–98. [Google Scholar] [CrossRef]
- Ali, S.; Nguyen, D.Q.; Falk, W.; Martin, M.U. Caspase 3 inactivates biologically active full length interleukin-33 as a classical cytokine but does not prohibit nuclear translocation. Biochem. Biophys. Res. Commun. 2010, 91, 512–516. [Google Scholar] [CrossRef]
- Millar, N.L.; O’Donnell, C.; McInnes, I.B.; Brint, E. The role of the IL-33/ST2 pathway in tissue repair and tumorigenesis. Semin. Cell Dev. Biol. 2017, 61, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Ohno, T.; Oboki, K.; Morita, H.; Kajiwara, N.; Arae, K.; Tanaka, S.; Ikeda, M.; Likura, M.; Akiyama, T.; Inoue, J.; et al. Paracrine IL-33 stimulation enhances lipopolysaccharide-mediated macrophage activation. PLoS ONE 2011, 6, e18404. [Google Scholar] [CrossRef]
- Ivanov, V.N.; Zhou, H.; Ghandhi, S.A.; Karasic, T.B.; Yaghoubian, B.; Amundson, S.A.; Hei, T.K. Radiation-induced bystander signaling pathways in human fibroblasts: A role for interleukin-33 in the signal transmission. Cell. Signal. 2010, 22, 1076–1087. [Google Scholar] [CrossRef] [Green Version]
- Modofsky, A.B.; Savage, A.; Locksley, R.M. Interleukin-33 in tissue homeostasis, injury and inflammation. Immunity 2015, 42, 1005–1019. [Google Scholar] [CrossRef] [Green Version]
- Espinassous, Q.; Garcia-de-Paco, E.; Garcia-Verdugo, I.; Synguelakis, M.; von Aulock, S.; Sallenave, J.M.; McKenzie, A.N.; Kanellopoulos, J. IL-33 enhances lipopolysaccharide-induced inflammatory cytokine production from mouse macrophages by regulating lipopolysaccharide receptor complex. J. Immunol. 2009, 183, 1446–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caselli, C. Inflammation in cardiac disease: Focus on interleukin-33/ST2 pathway. Inflamm. Cell Signal. 2014, 1, 118–151. [Google Scholar] [CrossRef]
- Miller, A.M. Role of IL-33 in inflammation and disease. J. Inflamm. 2011, 8, 22. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Hammel, H.; He, Y.; Tainer, J.A.; Jeng, U.S.; Zhang, L.; Wang, S.; Wang, S.; Wang, X. Structural insights into the interaction of IL-33 with its receptor. PNAS. 2013, 110, 14918–14923. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pastorelli, L.; De Salvo, C.; Vecchi, M.; Pizarro, T.T. The role of IL-33 in gut mucosal inflammation. Mediators Inflamm. 2013, 608187. [Google Scholar] [CrossRef] [PubMed]
- Lopetuso, H.; Scaldaferri, F.; Pizarro, T.T. Emerging role of the interleukin (IL)-23/ST2 axis in gut mucosal wound healing and fibrosis. Fibrogenes. Tissue Repair. 2012, 5, 18–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jovanovic, I.; Radosavljevic, G.; Mitrovic, M.; Juranic, V.L.; McKenzie, A.N.; Arsenijevic, N.; Jonjic, S.; Lukic, M.L. ST2 deletion enhances innate and acquired immunity to murine mammary carcinoma. Eur. J. Immunol. 2011, 41, 1902–1912. [Google Scholar] [CrossRef] [Green Version]
- Kuroiwa, K.; Arai, T.; Okazaki, H.; Minota, S.; Tominaga, S. Identification of human ST2 protein in the sera of patients with autoimmune diseases. Biochem. Biophys. Res. Commun. 2001, 284, 1104–1108. [Google Scholar] [CrossRef]
- López-Casado, M.A.; Lorite, P.; Palomeque, T.; Torres, M.I. Potential role of the IL-33/ST2 axis in celiac disease. Cell. Mol. Immunol. 2015, 1, 282–285. [Google Scholar] [CrossRef] [Green Version]
- Buzelli, J.N.; Chalinor, H.V.; Pavlic, D.I.; Sutton, P.; Menheniott, T.R.; Giraud, A.S.; Judd, L.M. IL-33 is a stomach alarmin that initiates a skewed Th2 response to injury and infection. Cell. Mol. Gastroenterol. Hepatol. 2015, 1, 203–221. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.F.; Mitchell, A.; Li, G.; Ding, S.; Fitzmaurice, A.M.; Ryan, K.; Crowe, S.; Goldberg, J.B. Toll-like receptor (TLR) 2 and TLR5 but not TLR4, are required for H. pylori-induced NF-kappa B activation and chemokine expression by epithelial cells. J. Biol. Chem 2003, 278, 32552–32560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.X.; Kaiedaa, S.; Ameria, S.; Fishgala, N.; Dwyera, D.; Dellingerc, A.; Kepley, C.L.; Gurish, M.F.; Nigrovic, P.A. IL-33/ST2 axis promotes mast cell survival via BCLXL. Proc. Natl. Acad. Sci. USA. 2014, 11, 10281–10286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonciarz, W.; Walencka, M.; Moran, A.P.; Hinc, K.; Obuchowski, M.; Chmiela, M. Upregulation of MUC5AC production and deposition of Lewis determinants by Helicobacter pylori facilitate gastric tissue colonization and the maintenance of infection. J. Biomed. Sci. 2019, 26, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonciarz, W.; Krupa, A.; Hinc, K.; Obuchowski, M.; Moran, A.P.; Gajewski, A.; Chmiela, M. The effect of Helicobacter pylori infection and different H. pylori components on the proliferation and apoptosis of gastric epithelial cells and fibroblasts. PLoS ONE 2019, 14, e0220636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Paulis, A.; Prevete, N.; Rossi, F.W.; Rivellese, F.; Salerno, F.; Delfino, G.; Liccardo, B.; Avilla, E.; Montuori, N.; Mascolo, M.; et al. Helicobacter pylori Hp(2-20) promotes migration and proliferation of gastric epithelial cells by interacting with formyl peptide receptors in vitro and accelerates gastric mucosal healing in vivo. J. Immunol. 2009, 183, 3761–3769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lott, J.M.; Sumpter, T.L.; Turnquist, H.R. New dog and newtricks: Evolving roles for IL-33 in type 2 immunity. J. Leukoc. Biol. 2015, 97, 1037–1048. [Google Scholar] [CrossRef]
- Hu, W.T.; Li, M.Q.; Liu, W.; Jin, L.P.; Li, D.J.; Zhu, X.Y. IL-33 enhances proliferation and invasiveness of decidual stromal cells by up-regulation of CCL2/CCR2 via NF-kB and ERK 1/2 signalling. Mol. Hum. Reprod. 2014, 20, 358–372. [Google Scholar] [CrossRef] [Green Version]
- Seki, K.; Sanada, S.; Kudinova, A.Y.; Steinhauser, M.L.; Handa, V.; Gannon, J.; Lee, R.T. Interleukin-33 prevents apoptosis and improves survival after experimental myocardial infarction through ST2 signaling. Circ. Heart Fail. 2009, 2, 684–691. [Google Scholar] [CrossRef] [Green Version]
- Oboki, K.; Ohno, T.; Kajiwara, N.; Saito, H.; Nakae, S. IL-33 and IL-33 receptors in host defense and diseases. Allergol. Int. 2010, 59, 143–160. [Google Scholar] [CrossRef] [Green Version]
- Oboki, K.; Ohno, T.; Kajiwara, N.; Arae, K.; Morita, H.; Ishii, A.; Nambu, A.; Abe, T.; Kiyonari, H.; Matsumoto, K.; et al. IL-33 is a crucial amplifier of innate rather than acquired immunity. Proc. Natl. Acad. Sci. USA 2010, 107, 18581–18586. [Google Scholar] [CrossRef] [Green Version]
- Matter, K.; Aijaz, S.; Tsapara, A.; Balda, M.S. Mammalian tight junctions in the regulation of epithelial differentiation and proliferation. Curr. Opin. Cell Biol. 2005, 17, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Varbanova, M.; Frauenschläger, K.; Malfertheiner, P. Chronic gastritis—An update. Best Pract. Res. Clin. Gastroenterol. 2014, 28, 1031–1042. [Google Scholar] [CrossRef] [PubMed]
- Matzinger, P. Tolerance, danger, and the extended family. Annu. Rev. Immunol. 1994, 12, 991–1045. [Google Scholar] [CrossRef] [PubMed]
- Carriere, V.; Roussel, L.; Ortega, N.; Lacorre, D.A.; Americh, L.; Aguilar, L.; Bouche, G.; Girard, J.P. IL-33, the IL-1-like cytokine ligand for ST2 receptor, is a chromatin-associated nuclear factor in vivo. Proc. Natl. Acad. Sci. USA. 2007, 104, 282–287. [Google Scholar] [CrossRef] [Green Version]
- Ramaro, N.; Meyer, T.F. Helicobacter pylori resists phagocytosis by macrophages: Quantitative assessment by confocal microscopy and fluorescence-activated cell sorting. Infect. Immun. 2001, 69, 2604–2611. [Google Scholar] [CrossRef] [Green Version]
- Mnich, E.; Kowalewicz-Kulbat, M.; Sicińska, P.; Hinc, K.; Obuchowski, M.; Gajewski, A.; Moran, A.P.; Chmiela, M. Impact of Helicobacter pylori on the healing process of the gastric barrier. World J. Gastroenterol. 2016, 22, 7536–7558. [Google Scholar] [CrossRef]
- Viladomiu, M.; Bassaganya-Riera, J.; Tubau-Juni, N.; Kronsteiner, B.; Leber, A.; Philipson, C.W.; Zoccoli-Rodriguez, V.; Hontecillas, R. Cooperation of gastric mononuclear phagocytes with Helicobacter pylori during colonization. J. Immunol. 2017, 198, 3195–3204. [Google Scholar] [CrossRef] [Green Version]
- Krzysiek-Maczka, G.; Targosz, A.; Ptak-Belowska, A.; Korbut, E.; Szczyrk, U.; Strzalka, M.; Brzozowski, T. Molecular alterations in fibroblasts exposed to Helicobacter pylori: A missing link in bacterial inflammation progressing into gastric carcinogenesis? J. Physiol. Pharmacol. 2013, 64, 77–87. [Google Scholar]
- Yi, L.; Huang, X.; Guo, F.; Zhou, Z.; Chang, M.; Tang, J.; Huan, J. Lipopolysaccharide induces human pulmonary micro-vascular endothelial apoptosis via the YAP signaling pathway. Front. Cell. Infect. Microbiol. 2016, 6, 133. [Google Scholar] [CrossRef] [Green Version]
- Gajewski, A.; Gonciarz, W.; Hinc, K.; Obuchowski, M.; Chmiela, M. Dysfunction of gastric barrier in the milieu of 7-ketocholesterol, acetylsalicylic acid and Helicobacter pylori compounds. Potential implications in the development of coronary heart disease. Helicobacter. 2018, 23 (Suppl. S1), 65. [Google Scholar] [CrossRef] [Green Version]
- Shibata, J.; Goto, H.; Arisawa, T.; Niwa, Y.; Hayakawa, T.; Nakayama, A.; Mori, N. Regulation of tumor necrosis factor (TNF) induced apoptosis by soluble TNF receptors in Helicobacter pylori infection. Gut. 1999, 45, 24–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walencka, M.; Gonciarz, W.; Mnich, E.; Gajewski, A.; Stawerski, P.; Knapik-Dabrowicz, A.; Chmiela, M. The microbiological, histological, immunological and molecular determinants of Helicobacer pylori infection in guinea pigs as a convenient animal model to study pathogenicity of these bacteria and the infection dependent immune response of the host. Acta Biochim. Pol. 2015, 62, 697–706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rechciński, T.; Chmiela, M.; Małecka-Panas, E.; Płaneta-Małecka, I.; Rudnicka, W. Serological indicators of Helicobacter pylori infection in adult dyspeptic patients and healthy blood donors. Microbiol. Immunol. 1997, 41, 387–393. [Google Scholar] [CrossRef] [PubMed]
- Czkwianianc, E.; Chmiela, M.; Lawnik, M.; Płaneta-Małecka, I.; Rudnicka, W. Serodiagnosis of Helicobacter pylori in children with gastroduodenitis. Centr. Eur. J. Immunol. 1997, 22, 240–247. [Google Scholar]
- Jensen, E.C. Quantitative analysis of histological staining and fluorescence using image. Anat. Rec. 2013, 3, 378–381. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Function | Activity | Ref. |
|---|---|---|
| DAMP | signals the innate immune cells in response to stress or cell membrane disruption | [33,34] |
| Transcription factor | regulates the gene expression | [35,36,37,38] |
| Cytokine | signals the immune cells via ST2 receptor to eliminate infection and manage tissue injury (proregenerative activity) | [39,40,41,42] |
| modulates the T helper 2 (Th2) lymphocytes as well as Th1 and regulatory lymphocytes | [43,44,45] | |
| upregulates the inflammatory response to bacterial LPS | [42] | |
| potentially is involved in the development of disease due to maintenance of chronic inflammation | [38,44,45,46,47,48,49,50,51] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gonciarz, W.; Krupa, A.; Chmiela, M. Proregenerative Activity of IL-33 in Gastric Tissue Cells Undergoing Helicobacter Pylori-Induced Apoptosis. Int. J. Mol. Sci. 2020, 21, 1801. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051801
Gonciarz W, Krupa A, Chmiela M. Proregenerative Activity of IL-33 in Gastric Tissue Cells Undergoing Helicobacter Pylori-Induced Apoptosis. International Journal of Molecular Sciences. 2020; 21(5):1801. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051801
Chicago/Turabian StyleGonciarz, Weronika, Agnieszka Krupa, and Magdalena Chmiela. 2020. "Proregenerative Activity of IL-33 in Gastric Tissue Cells Undergoing Helicobacter Pylori-Induced Apoptosis" International Journal of Molecular Sciences 21, no. 5: 1801. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms21051801