Transient Receptor Potential Ankyrin 1 (TRPA1) Is Involved in Upregulating Interleukin-6 Expression in Osteoarthritic Chondrocyte Models

 ,
,
Abstract
:1. Introduction
2. Results
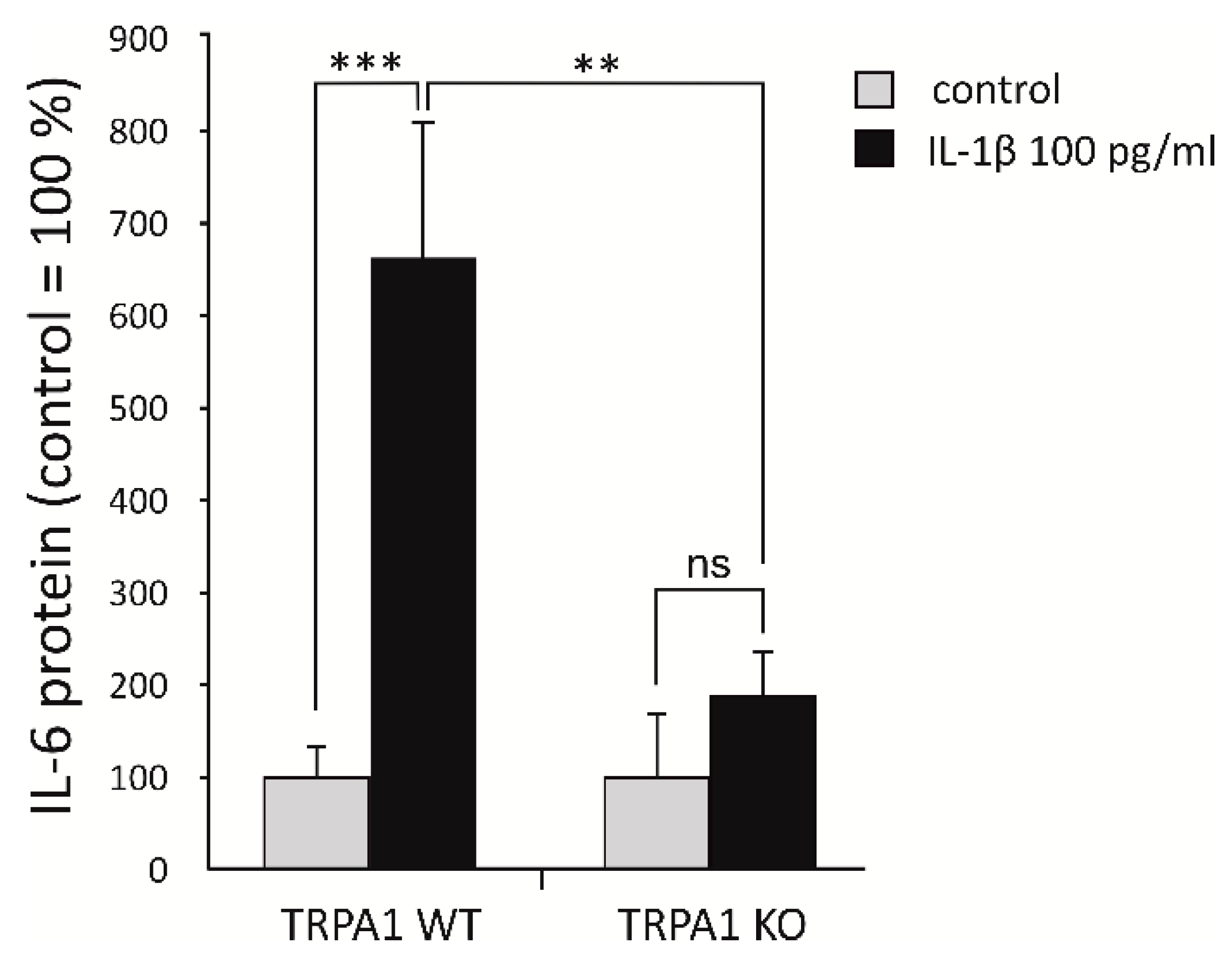
2.1. TRPA1 Regulates IL-6 Expression in Chondrocytes Based on RNA-Seq Data
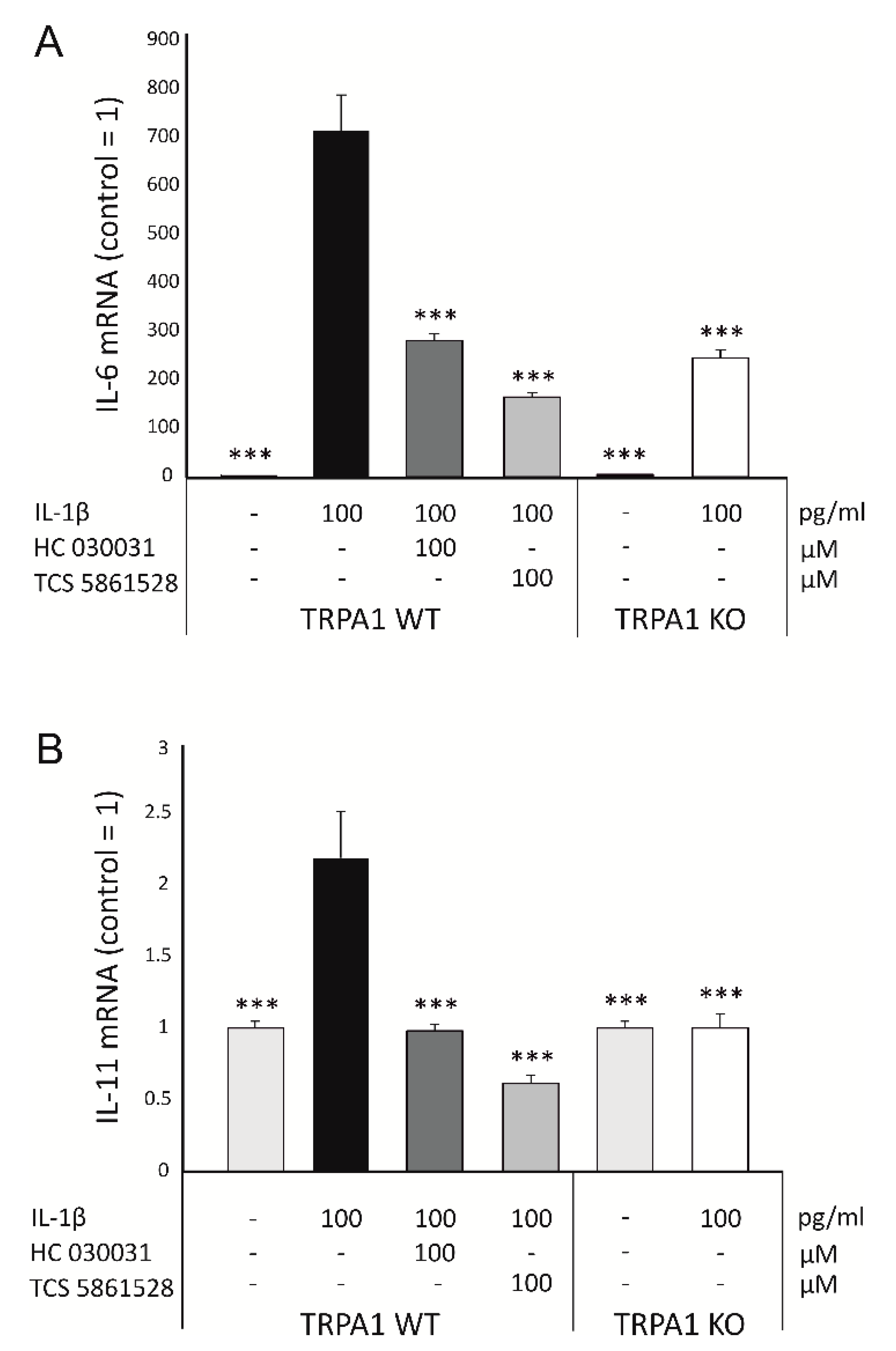
2.2. Expression of IL-6 Family Cytokines Is Downregulated by Genetic Deletion and Pharmacological Inhibition of TRPA1
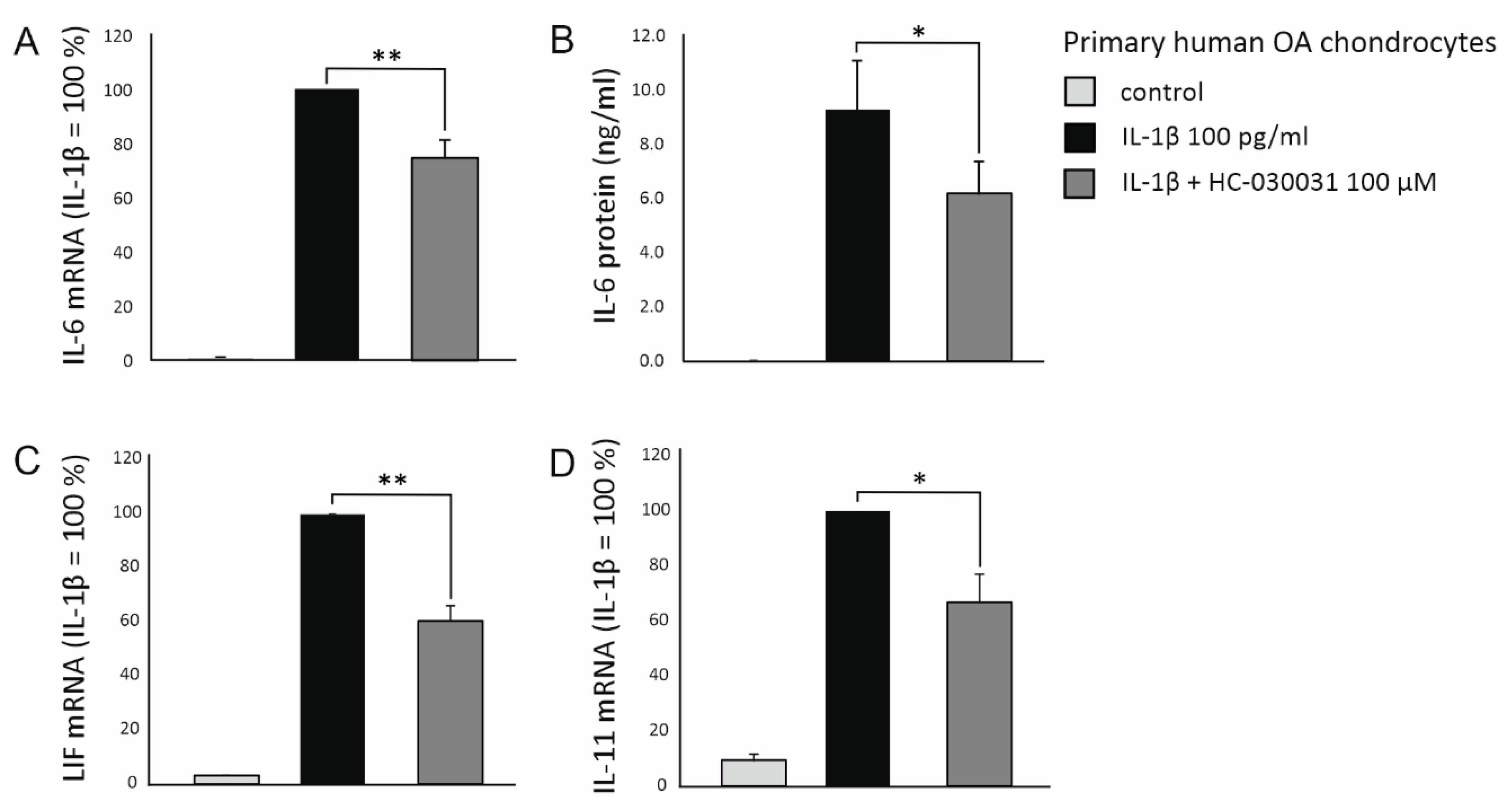
2.3. Possible Mediators Involved in TRPA1-Dependent IL-6 Expression
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Mouse Cartilage and Chondrocyte Culture
4.3. Human Chondrocyte Culture
4.4. RNA Extraction and Sample Preparation
4.5. Next-Generation RNA Sequencing (RNA-Seq) and Data Analysis
4.6. Quantitative RT-PCR
4.7. Immunoassay
4.8. Statistics
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AITC | allyl isothiocyanate |
| ANOVA | analysis of variance |
| ATP | adenosine triphosphate |
| CARD9 | caspase recruitment domain family, member 9 |
| CCR5 | chemokine (C-C motif) receptor 5 |
| CD36 | CD36 antigen |
| ELISA | enzyme-linked immunosorbent assay |
| FC | fold change |
| FDR | false discovery rate |
| GO | Gene Ontology |
| IL | interleukin |
| Lif | leukemia inhibitory factor |
| MIA | monosodium iodoacetate |
| MMP | matrix metalloproteinase |
| MSU | monosodium urate |
| OA | osteoarthritis |
| P2RX7 | purinergic receptor P2X7 |
| qRT-PCR | quantitative reverse transcription polymerase chain reaction |
| RNA-Seq | RNA sequencing |
| RPKM | reads per kilobase per million |
| TLR2 | Toll-like receptor 2 |
| TNF | tumor necrosis factor |
| TRPA1 | transient receptor potential ankyrin 1 |
| WT | wild-type |
References
- Zygmunt, P.M.; Högestätt, E.D. Trpa1. Handb. Exp. Pharmacol. 2014, 222, 583–630. [Google Scholar] [CrossRef] [PubMed]
- Koivisto, A.; Chapman, H.; Jalava, N.; Korjamo, T.; Saarnilehto, M.; Lindstedt, K.; Pertovaara, A. TRPA1: A transducer and amplifier of pain and inflammation. Basic Clin. Pharmacol. Toxicol. 2014, 114, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Bautista, D.M.; Movahed, P.; Hinman, A.; Axelsson, H.E.; Sterner, O.; Hogestatt, E.D.; Julius, D.; Jordt, S.E.; Zygmunt, P.M. Pungent products from garlic activate the sensory ion channel TRPA1. Proc. Natl. Acad. Sci. USA 2005, 102, 12248–12252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jordt, S.E.; Bautista, D.M.; Chuang, H.H.; McKemy, D.D.; Zygmunt, P.M.; Hogestatt, E.D.; Meng, I.D.; Julius, D. Mustard oils and cannabinoids excite sensory nerve fibres through the TRP channel ANKTM1. Nature 2004, 427, 260–265. [Google Scholar] [CrossRef] [PubMed]
- Bandell, M.; Story, G.M.; Hwang, S.W.; Viswanath, V.; Eid, S.R.; Petrus, M.J.; Earley, T.J.; Patapoutian, A. Noxious cold ion channel TRPA1 is activated by pungent compounds and bradykinin. Neuron 2004, 41, 849–857. [Google Scholar] [CrossRef] [Green Version]
- Anand, U.; Otto, W.R.; Facer, P.; Zebda, N.; Selmer, I.; Gunthorpe, M.J.; Chessell, I.P.; Sinisi, M.; Birch, R.; Anand, P. TRPA1 receptor localisation in the human peripheral nervous system and functional studies in cultured human and rat sensory neurons. Neurosci. Lett. 2008, 438, 221–227. [Google Scholar] [CrossRef]
- Hatano, N.; Itoh, Y.; Suzuki, H.; Muraki, Y.; Hayashi, H.; Onozaki, K.; Wood, I.C.; Beech, D.J.; Muraki, K. Hypoxia-inducible factor-1alpha (HIF1alpha) switches on transient receptor potential ankyrin repeat 1 (TRPA1) gene expression via a hypoxia response element-like motif to modulate cytokine release. J. Biol. Chem. 2012, 287, 31962–31972. [Google Scholar] [CrossRef] [Green Version]
- Nummenmaa, E.; Hämäläinen, M.; Moilanen, L.J.; Paukkeri, E.L.; Nieminen, R.M.; Moilanen, T.; Vuolteenaho, K.; Moilanen, E. Transient receptor potential ankyrin 1 (TRPA1) is functionally expressed in primary human osteoarthritic chondrocytes. Arthritis Res. Ther. 2016, 18, 185. [Google Scholar] [CrossRef] [Green Version]
- Nassini, R.; Pedretti, P.; Moretto, N.; Fusi, C.; Carnini, C.; Facchinetti, F.; Viscomi, A.R.; Pisano, A.R.; Stokesberry, S.; Brunmark, C.; et al. Transient receptor potential ankyrin 1 channel localized to non-neuronal airway cells promotes non-neurogenic inflammation. PLoS ONE 2012, 7, e42454. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Inoue, R.; Morii, T.; Takahashi, N.; Yamamoto, S.; Hara, Y.; Tominaga, M.; Shimizu, S.; Sato, Y.; Mori, Y. Nitric oxide activates TRP channels by cysteine S-nitrosylation. Nat. Chem. Biol. 2006, 2, 596–607. [Google Scholar] [CrossRef]
- Andersson, D.A.; Gentry, C.; Moss, S.; Bevan, S. Transient receptor potential A1 is a sensory receptor for multiple products of oxidative stress. J. Neurosci. 2008, 28, 2485–2494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gouin, O.; L’Herondelle, K.; Lebonvallet, N.; Le Gall-Ianotto, C.; Sakka, M.; Buhe, V.; Plee-Gautier, E.; Carre, J.L.; Lefeuvre, L.; Misery, L.; et al. TRPV1 and TRPA1 in cutaneous neurogenic and chronic inflammation: Pro-inflammatory response induced by their activation and their sensitization. Protein Cell 2017, 8, 644–661. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, D.J.; Bierma-Zeinstra, S. Osteoarthritis. Lancet 2019, 393, 1745–1759. [Google Scholar] [CrossRef]
- Mobasheri, A.; Batt, M. An update on the pathophysiology of osteoarthritis. Ann. Phys. Rehabil. Med. 2016, 59, 333–339. [Google Scholar] [CrossRef]
- Rahmati, M.; Mobasheri, A.; Mozafari, M. Inflammatory mediators in osteoarthritis: A critical review of the state-of-the-art, current prospects, and future challenges. Bone 2016, 85, 81–90. [Google Scholar] [CrossRef]
- Wojdasiewicz, P.; Poniatowski, L.A.; Szukiewicz, D. The role of inflammatory and anti-inflammatory cytokines in the pathogenesis of osteoarthritis. Mediat. Inflamm. 2014, 2014, 561459. [Google Scholar] [CrossRef] [Green Version]
- Kapoor, M.; Martel-Pelletier, J.; Lajeunesse, D.; Pelletier, J.P.; Fahmi, H. Role of proinflammatory cytokines in the pathophysiology of osteoarthritis. Nat. Rev. Rheumatol. 2011, 7, 33–42. [Google Scholar] [CrossRef]
- Chow, Y.Y.; Chin, K.Y. The Role of Inflammation in the Pathogenesis of Osteoarthritis. Mediat. Inflamm. 2020, 2020, 8293921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moilanen, L.J.; Hämäläinen, M.; Nummenmaa, E.; Ilmarinen, P.; Vuolteenaho, K.; Nieminen, R.M.; Lehtimäki, L.; Moilanen, E. Monosodium Iodoacetate-Induced Inflammation and Joint Pain are Reduced in TRPA1 Deficient mice—Potential Role of TRPA1 in Osteoarthritis. Osteoarthr. Cartil. 2015, 23, 2017–2026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nummenmaa, E.; Hämäläinen, M.; Moilanen, L.J.; Moilanen, T.; Vuolteenaho, K.; Moilanen, E. TRPA1 expression is downregulated by dexamethasone and aurothiomalate in human chondrocytes: TRPA1 as a novel factor and drug target in arthritis. RMD Open 2017, 3, e000556. [Google Scholar] [CrossRef] [PubMed]
- Moilanen, L.J.; Hämäläinen, M.; Lehtimäki, L.; Nieminen, R.M.; Moilanen, E. Urate crystal induced inflammation and joint pain are reduced in transient receptor potential ankyrin 1 deficient mice—Potential role for transient receptor potential ankyrin 1 in gout. PLoS ONE 2015, 10, e0117770. [Google Scholar] [CrossRef] [PubMed]
- Horvath, A.; Tekus, V.; Boros, M.; Pozsgai, G.; Botz, B.; Borbely, E.; Szolcsanyi, J.; Pinter, E.; Helyes, Z. Transient receptor potential ankyrin 1 (TRPA1) receptor is involved in chronic arthritis: In vivo study using TRPA1-deficient mice. Arthritis Res. Ther. 2016, 18, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandes, E.S.; Russell, F.A.; Alawi, K.M.; Sand, C.; Liang, L.; Salamon, R.; Bodkin, J.V.; Aubdool, A.A.; Arno, M.; Gentry, C.; et al. Environmental cold exposure increases blood flow and affects pain sensitivity in the knee joints of CFA-induced arthritic mice in a TRPA1-dependent manner. Arthritis Res. Ther. 2016, 18, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trevisan, G.; Hoffmeister, C.; Rossato, M.F.; Oliveira, S.M.; Silva, M.A.; Ineu, R.P.; Guerra, G.P.; Materazzi, S.; Fusi, C.; Nassini, R.; et al. Transient receptor potential ankyrin 1 receptor stimulation by hydrogen peroxide is critical to trigger pain during monosodium urate-induced inflammation in rodents. Arthritis. Rheum. 2013, 65, 2984–2995. [Google Scholar] [CrossRef]
- Mäki-Opas, I.; Hämäläinen, M.; Moilanen, L.J.; Haavikko, R.; Ahonen, T.J.; Alakurtti, S.; Moreira, V.M.; Muraki, K.; Yli-Kauhaluoma, J.; Moilanen, E. Pyrazine-Fused Triterpenoids Block the TRPA1 Ion Channel in Vitro and Inhibit TRPA1-Mediated Acute Inflammation in Vivo. ACS. Chem. Neurosci. 2019, 10, 2848–2857. [Google Scholar] [CrossRef]
- Moilanen, L.J.; Laavola, M.; Kukkonen, M.; Korhonen, R.; Leppänen, T.; Hogestatt, E.D.; Zygmunt, P.M.; Nieminen, R.M.; Moilanen, E. TRPA1 contributes to the acute inflammatory response and mediates carrageenan-induced paw edema in the mouse. Sci. Rep. 2012, 2, 380. [Google Scholar] [CrossRef]
- Taylor-Clark, T.E.; Ghatta, S.; Bettner, W.; Undem, B.J. Nitrooleic acid, an endogenous product of nitrative stress, activates nociceptive sensory nerves via the direct activation of TRPA1. Mol. Pharmacol. 2009, 75, 820–829. [Google Scholar] [CrossRef] [Green Version]
- Nummenmaa, E. TRPA1 as a Novel Factor and Drug Target in Osteoarthritis. Ph.D. Thesis, Tampere University, Tampere, Finland, 2020. [Google Scholar]
- Laavola, M.; Leppänen, T.; Hämäläinen, M.; Vuolteenaho, K.; Moilanen, T.; Nieminen, R.; Moilanen, E. IL-6 in Osteoarthritis: Effects of Pine Stilbenoids. Molecules 2018, 24, 109. [Google Scholar] [CrossRef] [Green Version]
- Väänänen, T.; Koskinen, A.; Paukkeri, E.L.; Hämäläinen, M.; Moilanen, T.; Moilanen, E.; Vuolteenaho, K. YKL-40 as a novel factor associated with inflammation and catabolic mechanisms in osteoarthritic joints. Mediat. Inflamm. 2014, 2014, 215140. [Google Scholar] [CrossRef] [Green Version]
- Guerne, P.A.; Carson, D.A.; Lotz, M. IL-6 production by human articular chondrocytes. Modulation of its synthesis by cytokines, growth factors, and hormones in vitro. J. Immunol. 1990, 144, 499–505. [Google Scholar]
- Kaneko, S.; Satoh, T.; Chiba, J.; Ju, C.; Inoue, K.; Kagawa, J. Interleukin-6 and interleukin-8 levels in serum and synovial fluid of patients with osteoarthritis. Cytokines Cell. Mol. Ther. 2000, 6, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Larsson, S.; Englund, M.; Struglics, A.; Lohmander, L.S. Interleukin-6 and tumor necrosis factor alpha in synovial fluid are associated with progression of radiographic knee osteoarthritis in subjects with previous meniscectomy. Osteoarthr. Cartil. 2015, 23, 1906–1914. [Google Scholar] [CrossRef] [Green Version]
- Livshits, G.; Zhai, G.; Hart, D.J.; Kato, B.S.; Wang, H.; Williams, F.M.; Spector, T.D. Interleukin-6 is a significant predictor of radiographic knee osteoarthritis: The Chingford Study. Arthritis Rheum. 2009, 60, 2037–2045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stannus, O.; Jones, G.; Cicuttini, F.; Parameswaran, V.; Quinn, S.; Burgess, J.; Ding, C. Circulating levels of IL-6 and TNF-alpha are associated with knee radiographic osteoarthritis and knee cartilage loss in older adults. Osteoarthr. Cartil. 2010, 18, 1441–1447. [Google Scholar] [CrossRef] [Green Version]
- Bellucci, F.; Meini, S.; Cucchi, P.; Catalani, C.; Nizzardo, A.; Riva, A.; Guidelli, G.M.; Ferrata, P.; Fioravanti, A.; Maggi, C.A. Synovial fluid levels of bradykinin correlate with biochemical markers for cartilage degradation and inflammation in knee osteoarthritis. Osteoarthr. Cartil. 2013, 21, 1774–1780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wong, P.K.; Campbell, I.K.; Egan, P.J.; Ernst, M.; Wicks, I.P. The role of the interleukin-6 family of cytokines in inflammatory arthritis and bone turnover. Arthritis Rheum. 2003, 48, 1177–1189. [Google Scholar] [CrossRef]
- Lotz, M.; Moats, T.; Villiger, P.M. Leukemia inhibitory factor is expressed in cartilage and synovium and can contribute to the pathogenesis of arthritis. J. Clin. Investig. 1992, 90, 888–896. [Google Scholar] [CrossRef]
- Jiang, Y.; Xiao, Q.; Hu, Z.; Pu, B.; Shu, J.; Yang, Q.; Lao, H.; Hao, J. Tissue levels of leukemia inhibitory factor vary by osteoarthritis grade. Orthopedics 2014, 37, 460. [Google Scholar] [CrossRef] [Green Version]
- Dunn, S.L.; Soul, J.; Anand, S.; Schwartz, J.M.; Boot-Handford, R.P.; Hardingham, T.E. Gene expression changes in damaged osteoarthritic cartilage identify a signature of non-chondrogenic and mechanical responses. Osteoarthr. Cartil. 2016, 24, 1431–1440. [Google Scholar] [CrossRef] [Green Version]
- Trontzas, P.; Kamper, E.F.; Potamianou, A.; Kyriazis, N.C.; Kritikos, H.; Stavridis, J. Comparative study of serum and synovial fluid interleukin-11 levels in patients with various arthritides. Clin. Biochem. 1998, 31, 673–679. [Google Scholar] [CrossRef]
- Kwan Tat, S.; Padrines, M.; Theoleyre, S.; Heymann, D.; Fortun, Y. IL-6, RANKL, TNF-alpha/IL-1: Interrelations in bone resorption pathophysiology. Cytokine Growth Factor Rev. 2004, 15, 49–60. [Google Scholar]
- ClinicalTrials.gov. A Clinical Trial to Study the Effect of the Interleukin-6 Receptor Antagonist Tocilizumab in Patients with Hand Osteoarthritis. Identifier: NCT02477059. Available online: https://clinicaltrials.gov/ct2/show/NCT02477059 (accessed on 15 September 2020).
- Nilsson, D.; Jennische, E.; Cavallini, N.; Braide, M. TRPA1 Mechanoreceptors Mediate the IL-6 Response to a Single PD Dwell in the Rat. Perit. Dial. Int. 2017, 37, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Escalera, J.; Balakrishna, S.; Fan, L.; Caceres, A.I.; Robinson, E.; Sui, A.; McKay, M.C.; McAlexander, M.A.; Herrick, C.A.; et al. TRPA1 controls inflammation and pruritogen responses in allergic contact dermatitis. FASEB. J. 2013, 27, 3549–3563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ClinicalTrials.gov. A Clinical Trial to Study the Effects of GRC 17536 in Patients with Painful Diabetic Peripheral Neuropathy. Identifier: NCT01726413. Available online: https://clinicaltrials.gov/ct2/show/NCT01726413 (accessed on 15 September 2020).
- Berridge, M.J.; Lipp, P.; Bootman, M.D. The versatility and universality of calcium signalling. Nat. Rev. Mol. Cell Biol. 2000, 1, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Papathanasiou, I.; Malizos, K.N.; Poultsides, L.; Karachalios, T.; Oikonomou, P.; Tsezou, A. The catabolic role of toll-like receptor 2 (TLR-2) mediated by the NF-kappaB pathway in septic arthritis. J. Orthop. Res. 2011, 29, 247–251. [Google Scholar] [CrossRef] [PubMed]
- Tang, C.H.; Hsu, C.J.; Fong, Y.C. The CCL5/CCR5 axis promotes interleukin-6 production in human synovial fibroblasts. Arthritis Rheum. 2010, 62, 3615–3624. [Google Scholar] [CrossRef] [PubMed]
- Lim, H.; Kim, Y.U.; Sun, H.; Lee, J.H.; Reynolds, J.M.; Hanabuchi, S.; Wu, H.; Teng, B.B.; Chung, Y. Proatherogenic conditions promote autoimmune T helper 17 cell responses in vivo. Immunity 2014, 40, 153–165. [Google Scholar] [CrossRef] [Green Version]
- Lister, M.F.; Sharkey, J.; Sawatzky, D.A.; Hodgkiss, J.P.; Davidson, D.J.; Rossi, A.G.; Finlayson, K. The role of the purinergic P2X7 receptor in inflammation. J. Inflamm. 2007, 4, 5. [Google Scholar] [CrossRef] [Green Version]
- Saijo, S.; Ikeda, S.; Yamabe, K.; Kakuta, S.; Ishigame, H.; Akitsu, A.; Fujikado, N.; Kusaka, T.; Kubo, S.; Chung, S.H.; et al. Dectin-2 recognition of alpha-mannans and induction of Th17 cell differentiation is essential for host defense against Candida albicans. Immunity 2010, 32, 681–691. [Google Scholar] [CrossRef] [Green Version]
- Moulin, D.; Donze, O.; Talabot-Ayer, D.; Mezin, F.; Palmer, G.; Gabay, C. Interleukin (IL)-33 induces the release of pro-inflammatory mediators by mast cells. Cytokine 2007, 40, 216–225. [Google Scholar] [CrossRef] [Green Version]
- Pfander, D.; Cramer, T.; Deuerling, D.; Weseloh, G.; Swoboda, B. Expression of thrombospondin-1 and its receptor CD36 in human osteoarthritic cartilage. Ann. Rheum. Dis. 2000, 59, 448–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, G.; Talabot-Ayer, D.; Lamacchia, C.; Toy, D.; Seemayer, C.A.; Viatte, S.; Finckh, A.; Smith, D.E.; Gabay, C. Inhibition of interleukin-33 signaling attenuates the severity of experimental arthritis. Arthritis Rheum. 2009, 60, 738–749. [Google Scholar] [CrossRef] [PubMed]
- Dosch, M.; Gerber, J.; Jebbawi, F.; Beldi, G. Mechanisms of ATP Release by Inflammatory Cells. Int. J. Mol. Sci. 2018, 19, 1222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egbuniwe, O.; Grover, S.; Duggal, A.K.; Mavroudis, A.; Yazdi, M.; Renton, T.; Di Silvio, L.; Grant, A.D. TRPA1 and TRPV4 activation in human odontoblasts stimulates ATP release. J. Dent. Res. 2014, 93, 911–917. [Google Scholar] [CrossRef] [Green Version]
- Ide, S.; Nishizawa, D.; Fukuda, K.; Kasai, S.; Hasegawa, J.; Hayashida, M.; Minami, M.; Ikeda, K. Haplotypes of P2RX7 gene polymorphisms are associated with both cold pain sensitivity and analgesic effect of fentanyl. Mol. Pain. 2014, 10, 75. [Google Scholar] [CrossRef]
- Solini, A.; Chiozzi, P.; Morelli, A.; Fellin, R.; Di Virgilio, F. Human primary fibroblasts in vitro express a purinergic P2X7 receptor coupled to ion fluxes, microvesicle formation and IL-6 release. J. Cell Sci. 1999, 112 Pt 3, 297–305. [Google Scholar]
- Kwan, K.Y.; Allchorne, A.J.; Vollrath, M.A.; Christensen, A.P.; Zhang, D.S.; Woolf, C.J.; Corey, D.P. TRPA1 contributes to cold, mechanical, and chemical nociception but is not essential for hair-cell transduction. Neuron 2006, 50, 277–289. [Google Scholar] [CrossRef] [Green Version]
- Tuure, L.; Hämäläinen, M.; Nummenmaa, E.; Moilanen, T.; Moilanen, E. Downregulation of microsomal prostaglandin E synthase-1 (mPGES-1) expression in chondrocytes is regulated by MAP kinase phosphatase-1 (MKP-1). Int. Immunopharmacol. 2019, 71, 139–143. [Google Scholar] [CrossRef]
- Jonason, J.H.; Hoak, D.; O’Keefe, R.J. Primary murine growth plate and articular chondrocyte isolation and cell culture. Methods Mol. Biol. 2015, 1226, 11–18. [Google Scholar] [CrossRef]
- Zhang, W.; Doherty, M.; Peat, G.; Bierma-Zeinstra, M.A.; Arden, N.K.; Bresnihan, B.; Herrero-Beaumont, G.; Kirschner, S.; Leeb, B.F.; Lohmander, L.S.; et al. EULAR evidence-based recommendations for the diagnosis of knee osteoarthritis. Ann. Rheum. Dis. 2010, 69, 483–489. [Google Scholar] [CrossRef] [Green Version]
- Tuure, L.; Pemmari, A.; Hämäläinen, M.; Moilanen, T.; Moilanen, E. Regulation of gene expression by MF63, doi:a selective inhibitor of microsomal PGE synthase 1 (mPGES1) in human osteoarthritic chondrocytes. Br. J. Pharmacol. 2020, 177, 4134–4146. [Google Scholar] [CrossRef] [PubMed]
- Andrews, A. FastQC: A Quality Control Tool for High Throughput Sequence Data. 2010. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc (accessed on 22 November 2020).
- Bolger, A.; Giorgi, F. Trimmomatic: A Flexible Read Trimming Tool for Illumina NGS Data. 2014. Available online: http://www.usadellab.org/cms/index.php (accessed on 22 November 2020).
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Nummenmaa, E.; Hämäläinen, M.; Moilanen, T.; Vuolteenaho, K.; Moilanen, E. Effects of FGF-2 and FGF receptor antagonists on MMP enzymes, aggrecan, and type II collagen in primary human OA chondrocytes. Scand. J. Rheumatol. 2015, 44, 321–330. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Gene | RNA-Seq | qRT-PCR | |||||
|---|---|---|---|---|---|---|---|
| RPKM (WT) | RPKM (TRPA1 KO) | FC | Adj. p-Value | FC | Adj. p-Value | ||
| IL-6 | Interleukin-6 | 202.37 | 48.26 | −3.86 | <0.0001 | −8.93 | <0.001 |
| LIF | Leukemia inhibitory factor | 12.03 | 5.19 | −2.16 | <0.0001 | −3.95 | <0.001 |
| IL-11 | Interleukin-11 | 1.11 | 0.21 | −4.14 | <0.0001 | −4.56 | <0.001 |
| Gene | RNA-Seq | qRT-PCR | |||||
|---|---|---|---|---|---|---|---|
| RPKM (WT) | RPKM (TRPA1 KO) | FC | Adj. p-Value | FC | Adj. p-Value | ||
| TLR2 | toll-like receptor 2 | 26.67 | 12.48 | −2.01 | <0.0001 | −3.92 | <0.001 |
| CD36 | CD36 antigen | 22.39 | 8.59 | −2.41 | <0.0001 | −3.81 | <0.001 |
| CCR5 | chemokine (C-C motif) receptor 5 | 13.15 | 1.70 | −6.87 | <0.0001 | −15.68 | <0.001 |
| P2RX7 | purinergic receptor P2X, ligand-gated ion channel, 7 | 3.73 | 1.35 | −2.57 | <0.0001 | −2.9 | <0.001 |
| CARD9 | caspase recruitment domain family, member 9 | 2.10 | 0.39 | −4.41 | <0.0001 | −9.04 | <0.001 |
| IL-33 | interleukin-33 | 0.34 | 0.06 | −2.81 | 0.0022 | −11.6 | <0.001 |
| Primer/Probe | Sequence | |
|---|---|---|
| hGAPDH | forward | 5′-AAGGTCGGAGTCAACGGATTT-3′ |
| reverse | 5′-GCAACAATATCCACTTTACCAGAGTTAA-3′ | |
| probe | 5′-CGCCTGGTCACCAGGGCTGC-3′ | |
| hIL-6 | forward | 5′-TACCCCCAGGAGAAGATTCCA-3′ |
| reverse | 5′-CCGTCGAGGATGTACCGAATT-3′ | |
| probe | 5′-CGCCCCACACAGACAGCCACTC-3′ | |
| mGAPDH | forward | 5′-GCATGGCCTTCCGTGTTC-3′ |
| reverse | 5′-GATGTCATCATACTTGGCAGGTTT-3′ | |
| probe | 5′-TCGTGGATCTGACGTGCCGCC-3′ | |
| mIL-6 | forward | 5′-TCGGAGGCTTAATTACACATGTTC-3′ |
| reverse | 5′-CAAGTGCATCATCGTTGTTCATAC-3′ | |
| probe | 5′-CAGAATTGCCATTGCACAACTCTTTTCTCA-3′ | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nummenmaa, E.; Hämäläinen, M.; Pemmari, A.; Moilanen, L.J.; Tuure, L.; Nieminen, R.M.; Moilanen, T.; Vuolteenaho, K.; Moilanen, E. Transient Receptor Potential Ankyrin 1 (TRPA1) Is Involved in Upregulating Interleukin-6 Expression in Osteoarthritic Chondrocyte Models. Int. J. Mol. Sci. 2021, 22, 87. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010087
Nummenmaa E, Hämäläinen M, Pemmari A, Moilanen LJ, Tuure L, Nieminen RM, Moilanen T, Vuolteenaho K, Moilanen E. Transient Receptor Potential Ankyrin 1 (TRPA1) Is Involved in Upregulating Interleukin-6 Expression in Osteoarthritic Chondrocyte Models. International Journal of Molecular Sciences. 2021; 22(1):87. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010087
Chicago/Turabian StyleNummenmaa, Elina, Mari Hämäläinen, Antti Pemmari, Lauri J. Moilanen, Lauri Tuure, Riina M. Nieminen, Teemu Moilanen, Katriina Vuolteenaho, and Eeva Moilanen. 2021. "Transient Receptor Potential Ankyrin 1 (TRPA1) Is Involved in Upregulating Interleukin-6 Expression in Osteoarthritic Chondrocyte Models" International Journal of Molecular Sciences 22, no. 1: 87. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22010087