
How to Target Spinal Metastasis in Experimental Research: An Overview of Currently Used Experimental Mouse Models and Future Prospects


Abstract
:1. Introduction
2. Technical Aspects of Currently Used In Vivo Metastasis Mouse Models with Special Focus on Spine Metastasis
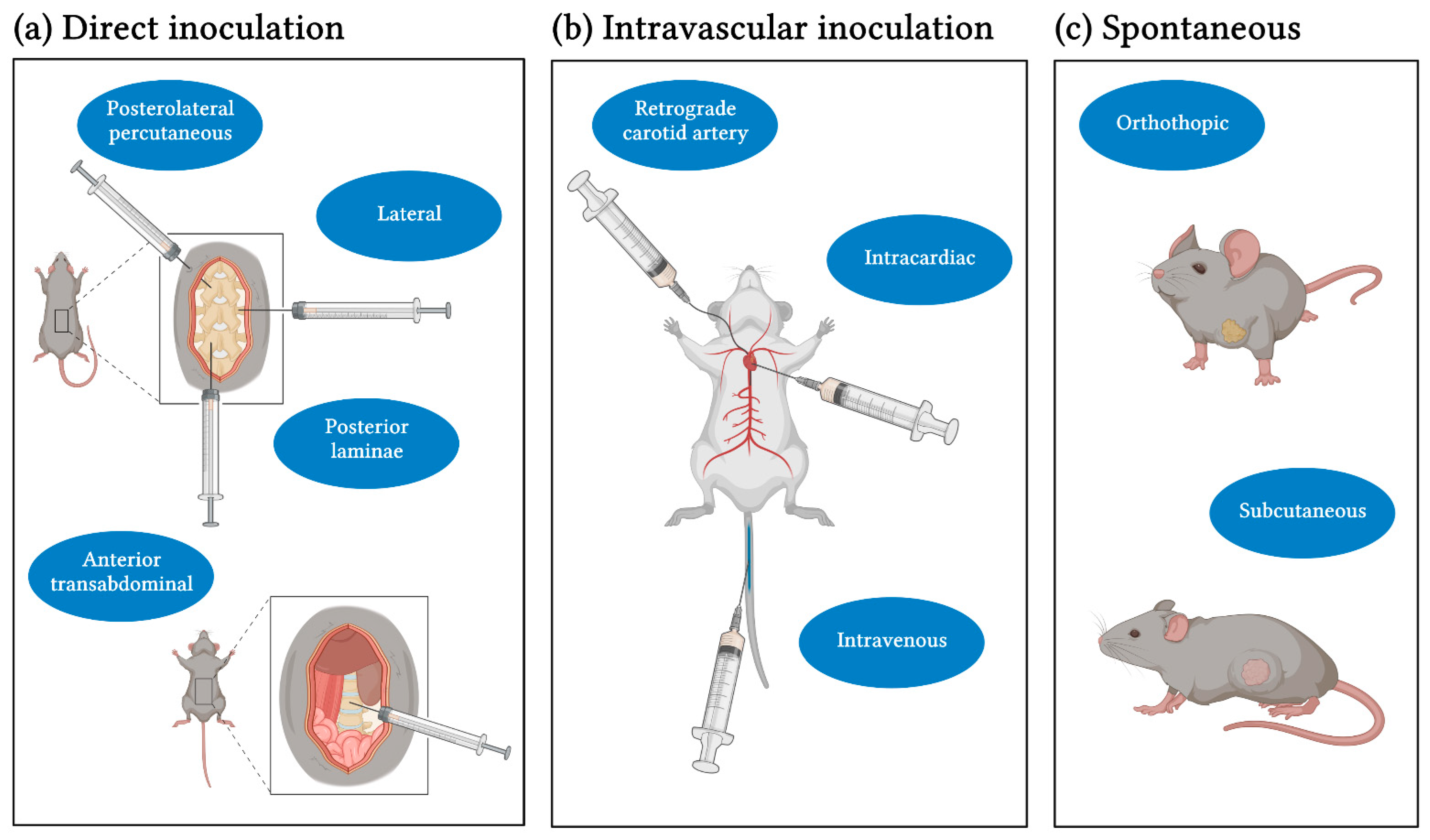
2.1. Inoculation Methods and Cell Lines
2.2. Time to Metastasis and Success Rate of Metastasis Establishment
2.3. Analysis Methods
2.4. Consideration of Clinical Characteristics
2.5. Enhancing Metastatic Potential
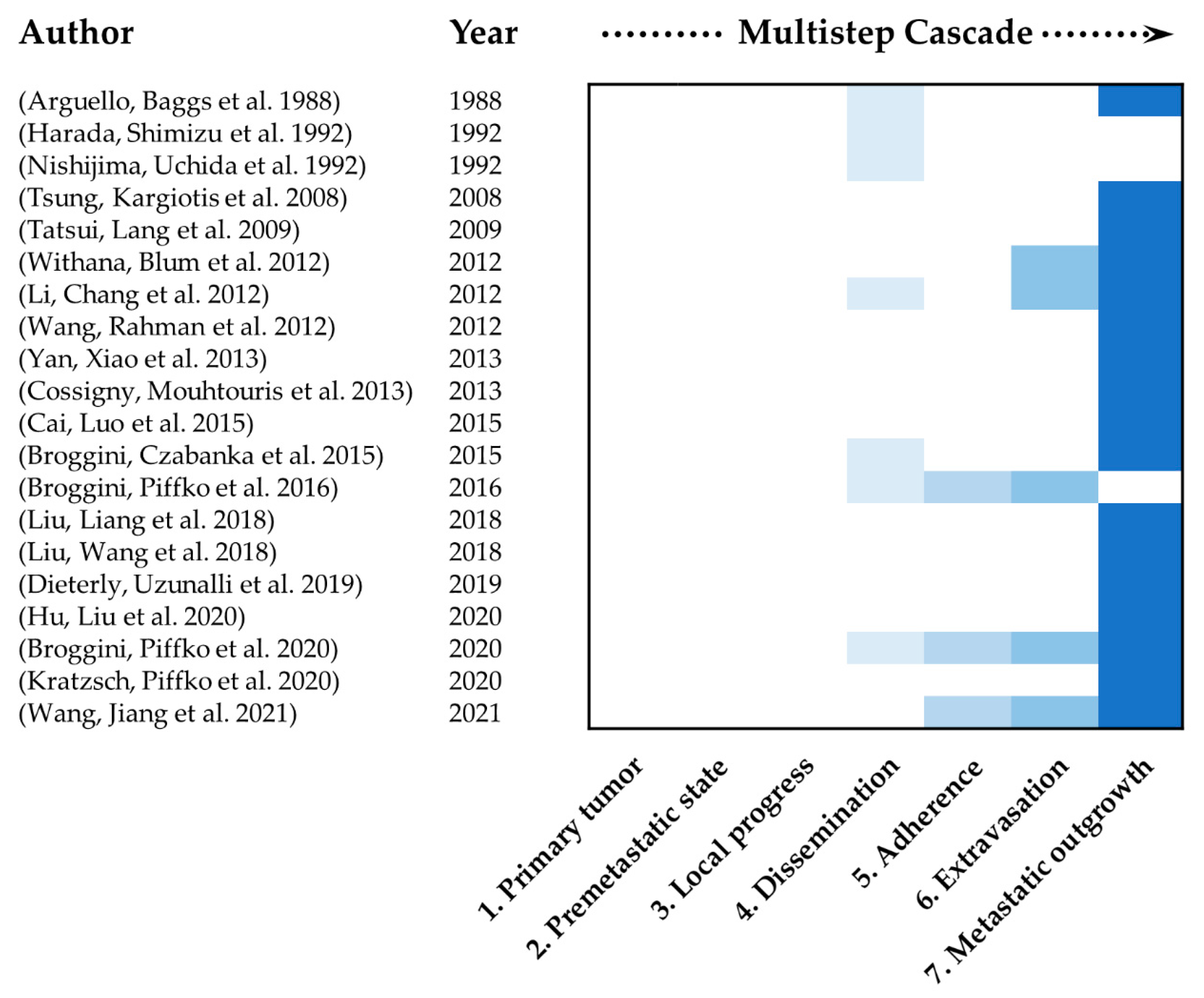
2.6. Mimicking of the Multistep Cascade of Metastasis
3. Spine Organotropism
4. The Complexity of the Metastatic Microenvironment
5. Future Prospects
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Maccauro, G.; Spinelli, M.S.; Mauro, S.; Perisano, C.; Graci, C.; Rosa, M.A. Physiopathology of spine metastasis. Int. J. Surg. Oncol. 2011, 2011, 107969. [Google Scholar] [CrossRef]
- Jacobs, W.B.; Perrin, R.G. Evaluation and treatment of spinal metastases: An overview. Neurosurg. Focus 2001, 11, e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vajkoczy, P. In Zukunft werden Spinale Metastasen nur noch in Tumorzentren versorgt: Pro. Die Wirbelsäule 2020, 04, 16–19. [Google Scholar] [CrossRef]
- Steeg, P.S. Targeting metastasis. Nat. Rev. Cancer 2016, 16, 201–218. [Google Scholar] [CrossRef] [PubMed]
- Lambert, A.W.; Pattabiraman, D.R.; Weinberg, R.A. Emerging Biological Principles of Metastasis. Cell 2017, 168, 670–691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryken, T.C.; Eichholz, K.M.; Gerszten, P.C.; Welch, W.C.; Gokaslan, Z.L.; Resnick, D.K. Evidence-based review of the surgical management of vertebral column metastatic disease. Neurosurg. Focus 2003, 15, 1–10. [Google Scholar] [CrossRef]
- Onken, J.S.; Fekonja, L.S.; Wehowsky, R.; Hubertus, V.; Vajkoczy, P. Metastatic dissemination patterns of different primary tumors to the spine and other bones. Clin. Exp. Metastasis 2019, 36, 493–498. [Google Scholar] [CrossRef]
- Akhtar, M.; Haider, A.; Rashid, S.; Al-Nabet, A. Paget’s “Seed and Soil” Theory of Cancer Metastasis: An Idea Whose Time has Come. Adv. Anat. Pathol. 2019, 26, 69–74. [Google Scholar] [CrossRef]
- Massagué, J.; Obenauf, A.C. Metastatic colonization by circulating tumour cells. Nature 2016, 529, 298–306. [Google Scholar] [CrossRef]
- Fidler, I.J. The pathogenesis of cancer metastasis: The seed and soil hypothesis revisited. Nat. Rev. Cancer 2003, 3, 453–458. [Google Scholar] [CrossRef]
- Patel, L.R.; Camacho, D.F.; Shiozawa, Y.; Pienta, K.J.; Taichman, R.S. Mechanisms of cancer cell metastasis to the bone: A multistep process. Future Oncol 2011, 7, 1285–1297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peinado, H.; Zhang, H.; Matei, I.R.; Costa-Silva, B.; Hoshino, A.; Rodrigues, G.; Psaila, B.; Kaplan, R.N.; Bromberg, J.F.; Kang, Y.; et al. Pre-metastatic niches: Organ-specific homes for metastases. Nat. Rev. Cancer 2017, 17, 302–317. [Google Scholar] [CrossRef]
- Neophytou, C.M.; Kyriakou, T.-C.; Papageorgis, P. Mechanisms of Metastatic Tumor Dormancy and Implications for Cancer Therapy. Int. J. Mol. Sci. 2019, 20, 6158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiraga, T. Bone metastasis: Interaction between cancer cells and bone microenvironment. J Oral Biosci. 2019, 61, 95–98. [Google Scholar] [CrossRef] [PubMed]
- Abdul Pari, A.A.; Singhal, M.; Augustin, H.G. Emerging paradigms in metastasis research. J. Exp. Med. 2021, 218. [Google Scholar] [CrossRef]
- Gengenbacher, N.; Singhal, M.; Augustin, H.G. Preclinical mouse solid tumour models: Status quo, challenges and perspectives. Nat. Rev. Cancer 2017, 17, 751–765. [Google Scholar] [CrossRef] [PubMed]
- Yao, A.; Sarkiss, C.A.; Ladner, T.R.; Jenkins, A.L. Contemporary spinal oncology treatment paradigms and outcomes for metastatic tumors to the spine: A systematic review of breast, prostate, renal, and lung metastases. J. Clin. Neurosci. 2017, 41, 11–23. [Google Scholar] [CrossRef]
- Sowder, M.E.; Johnson, R.W. Bone as a Preferential Site for Metastasis. JBMR Plus 2019, 3, e10126. [Google Scholar] [CrossRef]
- Arguello, F.; Baggs, R.B.; Frantz, C.N. A murine model of experimental metastasis to bone and bone marrow. Cancer Res. 1988, 48, 6876–6881. [Google Scholar]
- Harada, M.; Shimizu, A.; Nakamura, Y.; Nemoto, R. Role of the vertebral venous system in metastatic spread of cancer cells to the bone. Adv. Exp. Med. Biol. 1992, 324, 83–92. [Google Scholar] [CrossRef]
- Nishijima, Y.; Uchida, K.; Koiso, K.; Nemoto, R. Clinical significance of the vertebral vein in prostate cancer metastasis. Adv. Exp. Med. Biol. 1992, 324, 93–100. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Xiao, J.; Liu, T.; Huang, W.; Yang, X.; Wu, Z.; Huang, Q.; Qian, M. The effects of Hsp90 expression alteration on spinal metastases of breast carcinoma. Tumor Biol. 2013, 34, 1391–1397. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.; Luo, J.; Yang, X.; Deng, H.; Zhang, J.; Li, S.; Wei, H.; Yang, C.; Xu, L.; Jin, R.; et al. In vivoselection for spine-derived highly metastatic lung cancer cells is associated with increased migration, inflammation and decreased adhesion. Oncotarget 2015, 6, 22905–22917. [Google Scholar] [CrossRef] [Green Version]
- Tatsui, C.E.; Lang, F.F.; Gumin, J.; Suki, D.; Shinojima, N.; Rhines, L.D. An orthotopic murine model of human spinal metastasis: Histological and functional correlations. J. Neurosurg. Spine 2009, 10, 501–512. [Google Scholar] [CrossRef] [Green Version]
- Broggini, T.; Czabanka, M.; Piffko, A.; Harms, C.; Hoffmann, C.; Mrowka, R.; Wenke, F.; Deutsch, U.; Grötzinger, C.; Vajkoczy, P. ICAM1 depletion reduces spinal metastasis formation in vivo and improves neurological outcome. Eur. Spine J. 2015, 24, 2173–2181. [Google Scholar] [CrossRef]
- Broggini, T.; Piffko, A.; Hoffmann, C.J.; Harms, C.; Vajkoczy, P.; Czabanka, M. Passive Entrapment of Tumor Cells Determines Metastatic Dissemination to Spinal Bone and Other Osseous Tissues. PLoS ONE 2016, 11, e0162540. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Liang, Y.; Jiang, L.; Wang, H.; Wang, S.; Dong, J. CX3CL1/fractalkine enhances prostate cancer spinal metastasis by activating the Src/FAK pathway. Int. J. Oncol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Liu, P.; Wang, H.; Liang, Y.; Hu, A.; Xing, R.; Jiang, L.; Yi, L.; Dong, J. LINC00852 Promotes Lung Adenocarcinoma Spinal Metastasis by Targeting S100A9. J. Cancer 2018, 9, 4139–4149. [Google Scholar] [CrossRef]
- Dieterly, A.M.; Uzunalli, G.; Kemet, C.M.; Soepriatna, A.H.; Goergen, C.J.; Lyle, L.T. Epithelial–mesenchymal Transition Phenotypes in Vertebral Metastases of Lung Cancer. Toxicol. Pathol. 2019, 47, 515–527. [Google Scholar] [CrossRef]
- Hu, W.; Liu, Z.; Xiao, X.; Yang, Y.; Sun, Z.; Wang, X. Comparison of diagnostic efficacy of MRI and PET/CT in lung cancer of mouse with spinal metastasis. Cell. Mol. Biol. 2020, 66, 138. [Google Scholar] [CrossRef]
- Broggini, T.; Piffko, A.; Hoffmann, C.J.; Ghori, A.; Harms, C.; Adams, R.H.; Vajkoczy, P.; Czabanka, M. Ephrin-B2–EphB4 communication mediates tumor–endothelial cell interactions during hematogenous spread to spinal bone in a melanoma metastasis model. Oncogene 2020, 39, 7063–7075. [Google Scholar] [CrossRef] [PubMed]
- Kratzsch, T.; Piffko, A.; Broggini, T.; Czabanka, M.; Vajkoczy, P. Role of mTOR and VEGFR Inhibition in Prevention of Metastatic Tumor Growth in the Spine. Front. Oncol. 2020, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Jiang, L.; Hu, A.; Sun, C.; Zhou, L.; Huang, Y.; Chen, Q.; Dong, J.; Zhou, X.; Zhang, F. Vertebral-specific activation of the CX3CL1/ICAM-1 signaling network mediates non-small-cell lung cancer spinal metastasis by engaging tumor cell-vertebral bone marrow endothelial cell interactions. Theranostics 2021, 11, 4770–4789. [Google Scholar] [CrossRef] [PubMed]
- Tsung, A.; Kargiotis, O.; Chetty, C.; Lakka, S.; Gujrati, M.; Spomar, D.; Dinh, D.; Rao, J. Downregulation of matrix metalloproteinase-2 (MMP-2) utilizing adenovirus-mediated transfer of small interfering RNA (siRNA) in a novel spinal metastatic melanoma model. Int. J. Oncol. 2008. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Rahman, S.; Lin, C.Y.; Valdivia, J.; Than, K.; La Marca, F.; Park, P. A novel murine model of human renal cell carcinoma spinal metastasis. J. Clin. Neurosci. 2012, 19, 881–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cossigny, D.A.F.; Mouhtouris, E.; Dushyanthen, S.; Gonzalvo, A.; Quan, G.M.Y. An in vivo mouse model of intraosseous spinal cancer causing evolving paraplegia. J. Neuro Oncol. 2013, 115, 189–196. [Google Scholar] [CrossRef]
- Withana, N.P.; Blum, G.; Sameni, M.; Slaney, C.; Anbalagan, A.; Olive, M.B.; Bidwell, B.N.; Edgington, L.; Wang, L.; Moin, K.; et al. Cathepsin B Inhibition Limits Bone Metastasis in Breast Cancer. Cancer Res. 2012, 72, 1199–1209. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.Y.; Chang, J.W.; Hsieh, L.L.; Chen, S.M.; Yeh, K.Y. Low-dose zoledronic acid reduces spinal cord metastasis in pulmonary adenocarcinoma with neuroendocrine differentiation. Anticancer Drugs 2012, 23, 970–978. [Google Scholar] [CrossRef] [PubMed]
- Wright, L.E.; Ottewell, P.D.; Rucci, N.; Peyruchaud, O.; Pagnotti, G.M.; Chiechi, A.; Buijs, J.T.; Sterling, J.A. Murine models of breast cancer bone metastasis. BoneKEy Rep. 2016, 5, 804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anisimov, V.N.; Ukraintseva, S.V.; Yashin, A.I. Cancer in rodents: Does it tell us about cancer in humans? Nat. Rev. Cancer 2005, 5, 807–819. [Google Scholar] [CrossRef]
- Gómez-Cuadrado, L.; Tracey, N.; Ma, R.; Qian, B.; Brunton, V.G. Mouse models of metastasis: Progress and prospects. Dis. Models Mech. 2017, 10, 1061–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiang, L.; Gilkes, D. The Contribution of the Immune System in Bone Metastasis Pathogenesis. Int. J. Mol. Sci. 2019, 20, 999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hibberd, C.; Cossigny, D.A.F.; Quan, G.M.Y. Animal Cancer Models of Skeletal Metastasis. Cancer Growth Metastasis 2013, 6, CGM.S11284. [Google Scholar] [CrossRef] [Green Version]
- Huehnchen, P.; Boehmerle, W.; Endres, M. Assessment of Paclitaxel Induced Sensory Polyneuropathy with “Catwalk” Automated Gait Analysis in Mice. PLoS ONE 2013, 8, e76772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Bian, C.; Liang, Y.; Jiang, L.; Qian, C.; Dong, J. CX3CL1: A potential chemokine widely involved in the process spinal metastases. Oncotarget 2017, 8, 15213–15219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chambers, A.F.; Groom, A.C.; Macdonald, I.C. Dissemination and growth of cancer cells in metastatic sites. Nat. Rev. Cancer 2002, 2, 563–572. [Google Scholar] [CrossRef] [PubMed]
- Ponzetti, M.; Rucci, N. Switching Homes: How Cancer Moves to Bone. Int. J. Mol. Sci. 2020, 21, 4124. [Google Scholar] [CrossRef]
- Yi, L.; Liang, Y.; Zhao, Q.; Wang, H.; Dong, J. CX3CL1 Induces Vertebral Microvascular Barrier Dysfunction via the Src/P115-RhoGEF/ROCK Signaling Pathway. Front. Cell. Neurosci. 2020, 14. [Google Scholar] [CrossRef]
- Szerlip, N.J.; Calinescu, A.; Smith, E.; Tagett, R.; Clines, K.L.; Moon, H.H.; Taichman, R.S.; Van Poznak, C.H.; Clines, G.A. Dural Cells Release Factors Which Promote Cancer Cell Malignancy and Induce Immunosuppressive Markers in Bone Marrow Myeloid Cells. Neurosurgery 2018, 83, 1306–1316. [Google Scholar] [CrossRef]
- Wang, Y.; Chu, Y.; Ren, X.; Xiang, H.; Xi, Y.; Ma, X.; Zhu, K.; Guo, Z.; Zhou, C.; Zhang, G.; et al. Epidural adipose tissue-derived mesenchymal stem cell activation induced by lung cancer cells promotes malignancy and EMT of lung cancer. Stem Cell Res. Ther. 2019, 10, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Batson, O.V. The function of the vertebral veins and their role in the spread of metastases. Ann. Surg. 1940, 112, 138–149. [Google Scholar] [CrossRef] [PubMed]
- Stringer, M.D.; Restieaux, M.; Fisher, A.L.; Crosado, B. The vertebral venous plexuses: The internal veins are muscular and external veins have valves. Clin. Anat. 2012, 25, 609–618. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, K.; Decater, T.; Iwanaga, J.; Maulucci, C.M.; Bui, C.J.; Dumont, A.S.; Tubbs, R.S. Revisiting the Vertebral Venous Plexus-A Comprehensive Review of the Literature. World Neurosurg. 2021, 145, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Yuh, W.T.; Quets, J.P.; Lee, H.J.; Simonson, T.M.; Michalson, L.S.; Nguyen, P.T.; Sato, Y.; Mayr, N.A.; Berbaum, K.S. Anatomic distribution of metastases in the vertebral body and modes of hematogenous spread. Spine 1996, 21, 2243–2250. [Google Scholar] [CrossRef]
- Mishra, A.; Shiozawa, Y.; Pienta, K.J.; Taichman, R.S. Homing of cancer cells to the bone. Cancer Microenviron. 2011, 4, 221–235. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Xie, X.; Wu, J. Mechanism of lung adenocarcinoma spine metastasis induced by CXCL17. Cell. Oncol. 2020, 43, 311–320. [Google Scholar] [CrossRef]
- Fontanella, C.; Fanotto, V.; Rihawi, K.; Aprile, G.; Puglisi, F. Skeletal metastases from breast cancer: Pathogenesis of bone tropism and treatment strategy. Clin. Exp. Metastasis 2015, 32, 819–833. [Google Scholar] [CrossRef]
- Coleman, R.E.; Croucher, P.I.; Padhani, A.R.; Clézardin, P.; Chow, E.; Fallon, M.; Guise, T.; Colangeli, S.; Capanna, R.; Costa, L. Bone metastases. Nat. Rev. Dis. Primers 2020, 6. [Google Scholar] [CrossRef]
- Wu, M.-Y.; Li, C.-J.; Yiang, G.-T.; Cheng, Y.-L.; Tsai, A.P.-Y.; Hou, Y.-T.; Ho, Y.-C.; Hou, M.-F.; Chu, P.-Y. Molecular Regulation of Bone Metastasis Pathogenesis. Cell. Physiol. Biochem. 2018, 46, 1423–1438. [Google Scholar] [CrossRef]
- Li, R.; Wen, A.; Lin, J. Pro-Inflammatory Cytokines in the Formation of the Pre-Metastatic Niche. Cancers 2020, 12, 3752. [Google Scholar] [CrossRef]
- Monteran, L.; Ershaid, N.; Sabah, I.; Fahoum, I.; Zait, Y.; Shani, O.; Cohen, N.; Eldar-Boock, A.; Satchi-Fainaro, R.; Erez, N. Bone metastasis is associated with acquisition of mesenchymal phenotype and immune suppression in a model of spontaneous breast cancer metastasis. Sci. Rep. 2020, 10. [Google Scholar] [CrossRef]
- Kang, Y.; Pantel, K. Tumor Cell Dissemination: Emerging Biological Insights from Animal Models and Cancer Patients. Cancer Cell 2013, 23, 573–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahashi, M.; Furihata, M.; Akimitsu, N.; Watanabe, M.; Kaul, S.; Yumoto, N.; Okada, T. A highly bone marrow metastatic murine breast cancer model established through in vivo selection exhibits enhanced anchorage-independent growth and cell migration mediated by ICAM-1. Clin. Exp. Metastasis 2008, 25, 517–529. [Google Scholar] [CrossRef] [PubMed]
- Buijs, J.T.; Matula, K.M.; Cheung, H.; Kruithof-De Julio, M.; Van Der Mark, M.H.; Snoeks, T.J.; Cohen, R.; Corver, W.E.; Mohammad, K.S.; Jonkers, J.; et al. Spontaneous bone metastases in a preclinical orthotopic model of invasive lobular carcinoma; the effect of pharmacological targeting TGFβ receptor I kinase. J. Pathol. 2015, 235, 745–759. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, N.; Hoffman, R.M.; Zhao, M. Surgically-Induced Multi-organ Metastasis in an Orthotopic Syngeneic Imageable Model of 4T1 Murine Breast Cancer. Anticancer Res. 2015, 35, 4641–4646. [Google Scholar]
- Kurisunkal, V.; Gulia, A.; Gupta, S. Principles of Management of Spine Metastasis. Indian J. Orthop. 2020, 54, 181–193. [Google Scholar] [CrossRef]
- Conti, A.; Acker, G.; Kluge, A.; Loebel, F.; Kreimeier, A.; Budach, V.; Vajkoczy, P.; Ghetti, I.; Germano, A.F.; Senger, C. Decision Making in Patients with Metastatic Spine. The Role of Minimally Invasive Treatment Modalities. Front. Oncol. 2019, 9. [Google Scholar] [CrossRef]
- Laufer, I.; Bilsky, M.H. Advances in the treatment of metastatic spine tumors: The future is not what it used to be. J. Neurosurg. Spine 2019, 30, 299–307. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Author, Year | Primary Origin | Cell Line | Model | Inoculation Method | Time to Metastasis and Success Rate | Analysis | Clinical Characteristics |
|---|---|---|---|---|---|---|---|
| Intravascular Inoculation | |||||||
| Arguello, Baggs et al., 1988 [19] | Melanoma | G3.26 (B16) | Syngeneic, C57BL/6 | IV, IC | 14–20d, up to 100% | X-ray, Microscopic evaluation, Histology | No |
| Harada, Shimizu et al., 1992 [20] | Prostate | BrdU labelled PC-3 | Xenograft, ICR athymic mice | IV ± caval occlusion | 5 min, 75–100% | Histology | No |
| Nishijima, Uchida et al., 1992 [21] | Prostate | MBT-2 | Syngeneic, C3H/He | IV | 21d, 80% | Histology | Yes |
| Yan, Xiao et al., 2013 [22] | Breast | TM40D | Syngeneic, BALB/c | IC | 35d, 60% | BLI, Digital X-ray, Histology, RT-PCR | Yes, # |
| Cai, Luo et al., 2015 [23] | Lung | PC-9, A549, NCI-H1299, NCI-H460, H2030 | Xenograft, BALB/c nu/nu | IC | 40d, success rates depending on cell line, A549L6: 80% | X-ray, Micro-CT, MRI, Histology | Yes, milestones [24] |
| Broggini, Czabanka et al., 2015 [25] | Melanoma | B16-luc and mB16-luc cells | Syngeneic, C57BL/6J | RCAI | 19,5–29d | BLI, MRI, Histology | Yes |
| Broggini, Piffko et al., 2016 [26] | Lung, prostate, melanoma | B16-F1, LLC1, TRAMP-C2 | Syngeneic, C57BL/6J | RCAI | - | Luciferase measurement | No |
| Liu, Liang et al., 2018 [27] | Prostate | PC-3 | Xenograft, NOD/SCID mice | IC | 42–56d, 45% (CX3CR1-overexpression) | PET-CT, Histology | No |
| Liu, Wang et al., 2018 [28] | Lung | A549 and SPCA-1 | Xenograft, Balb/c nude mice | IC | 42–56d, 30% | PET-CT, Histology | No |
| Dieterly, Uzunalli et al., 2019 [29] | Lung | A549-Br (Brain seeking variant) | Xenograft, Athymic Nude-Foxn1nu mice | IC | 4–6 weeks, 39% | Histology | Yes |
| Hu, Liu et al., 2020 [30] | Lung | A549 | Xenograft, Balb/c | IC | 28–42d, 82,5% after 35d | MRI, PET-CT | No |
| Broggini, Piffko et al., 2020 [31] | Lung and melanoma | B16-F1 or LLC1 lung cancer cells | Syngeneic, Tamoxifen-inducible endothelial-specific ephrin-B2 knockout mice (efnb2iΔEC) and efnb2lox/lox | RCAI | 15d | BLI, MRI, Intravital fluorescens video microscopy, Histology | Yes, catwalk experiments |
| Kratzsch, Piffko et al., 2020 [32] | Melanoma | B16-F1 | Syngeneic, C57BL/6J mice | RCAI | 14–21d, 78% | BLI, MRI, Histology | Yes |
| Wang, Jiang et al., 2021 [33] | Lung | A549 | Xenograft, NOD/SCID mice | IC | Approx. 14d | BLI, CT, Histology, PCR | No |
| Direct Inoculation | |||||||
| Tsung, Kargiotis et al., 2008 [34] | Melanoma | Human melanoma cell line A2058 | Xenograft, Nude mice | Laterally into T10 | 18d, 100% | Histology | Yes |
| Wang, Rahman et al., 2012 [35] | Kidney | Human renal carcinoma cell line, ACHN | Xenograft, NOD/SCID | Spinous process and lamina | After 12 weeks, 33% paraplegia | Histology | Yes |
| Tatsui, Lang et al., 2009 [24] | Lung | PC-14 | Xenograft, Athymic Nude mice | Anterior transabdominal injection into L3 | 28d | Histology | Yes, ## |
| Cossigny, Mouhtouris et al., 2013 [36] | Breast and prostate | PC-3MDA-MB-231 | Xenograft, BALB/Nu-Nu athymic mice | Percutaneously upper lumbar spine | 21–35d, 100% | X-ray, Micro-CT Histology | Yes, ### |
| Spontaneous Mouse Models | |||||||
| Withana, Blum et al., 2012 [37] | Breast | 4T1.2 | Syngeneic, BALB/c | Orthotopic | 30d | Fluorescens Tomography, Histology, qRT-PCR | No |
| Li, Chang et al., 2012 [38] | Lung | NSCL murine Line 1 | Syngeneic, BALB/cByJ mice | SC | 7d, micrometastasis | BLI, Histology | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jelgersma, C.; Vajkoczy, P. How to Target Spinal Metastasis in Experimental Research: An Overview of Currently Used Experimental Mouse Models and Future Prospects. Int. J. Mol. Sci. 2021, 22, 5420. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115420
Jelgersma C, Vajkoczy P. How to Target Spinal Metastasis in Experimental Research: An Overview of Currently Used Experimental Mouse Models and Future Prospects. International Journal of Molecular Sciences. 2021; 22(11):5420. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115420
Chicago/Turabian StyleJelgersma, Claudius, and Peter Vajkoczy. 2021. "How to Target Spinal Metastasis in Experimental Research: An Overview of Currently Used Experimental Mouse Models and Future Prospects" International Journal of Molecular Sciences 22, no. 11: 5420. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22115420