
Gibberellin Oxidase Gene Family in L. chinense: Genome-Wide Identification and Gene Expression Analysis

 and
and
Abstract
:1. Introduction
2. Result
2.1. Identification of GA2ox and GA20ox genes in L. chinense
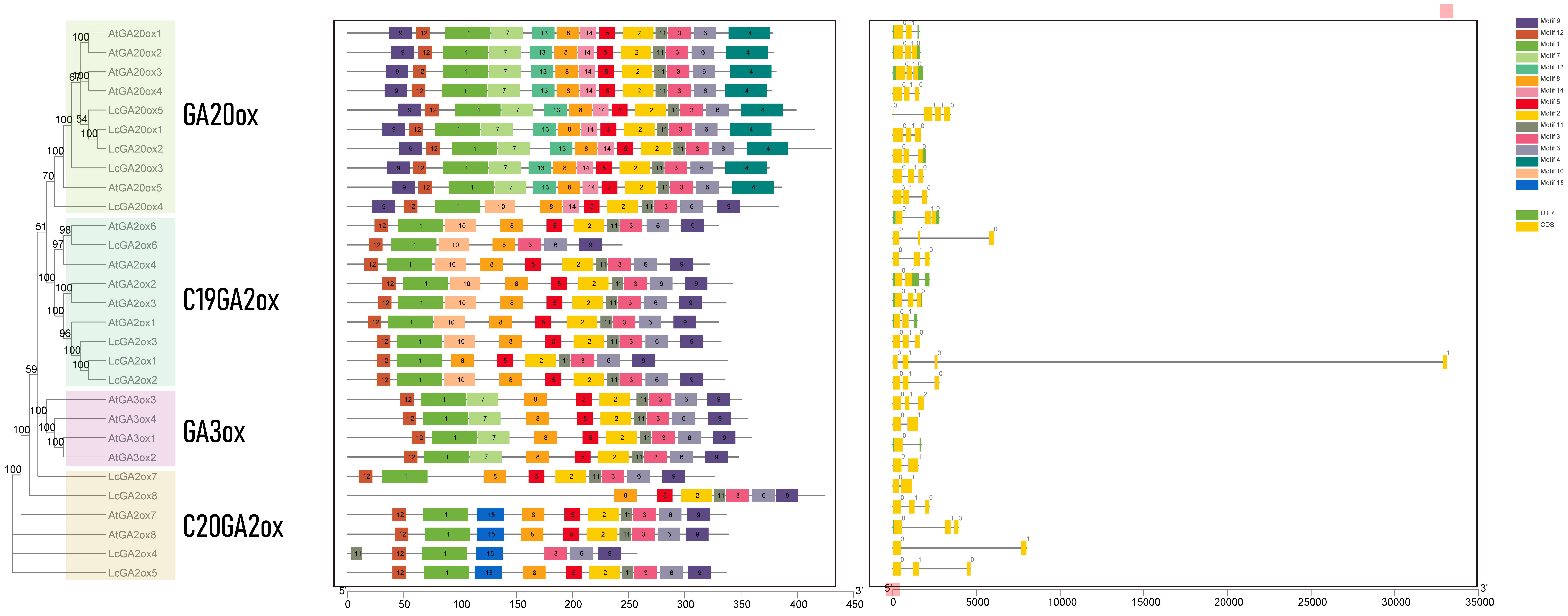
2.2. Evolutionary Analysis of LcGA20ox and LcGA2ox Genes
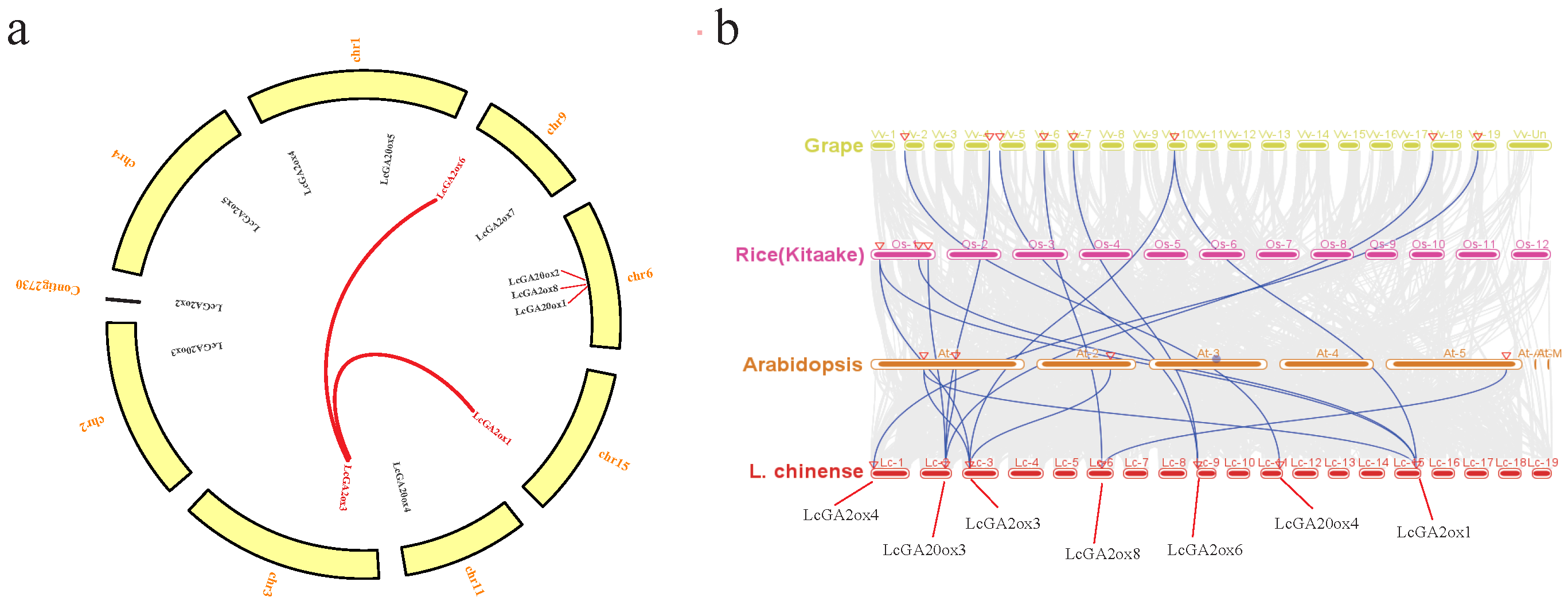
2.3. Synteny Analysis of GAox Genes in L. chinense, Grape, Arabidopsis, and Rice
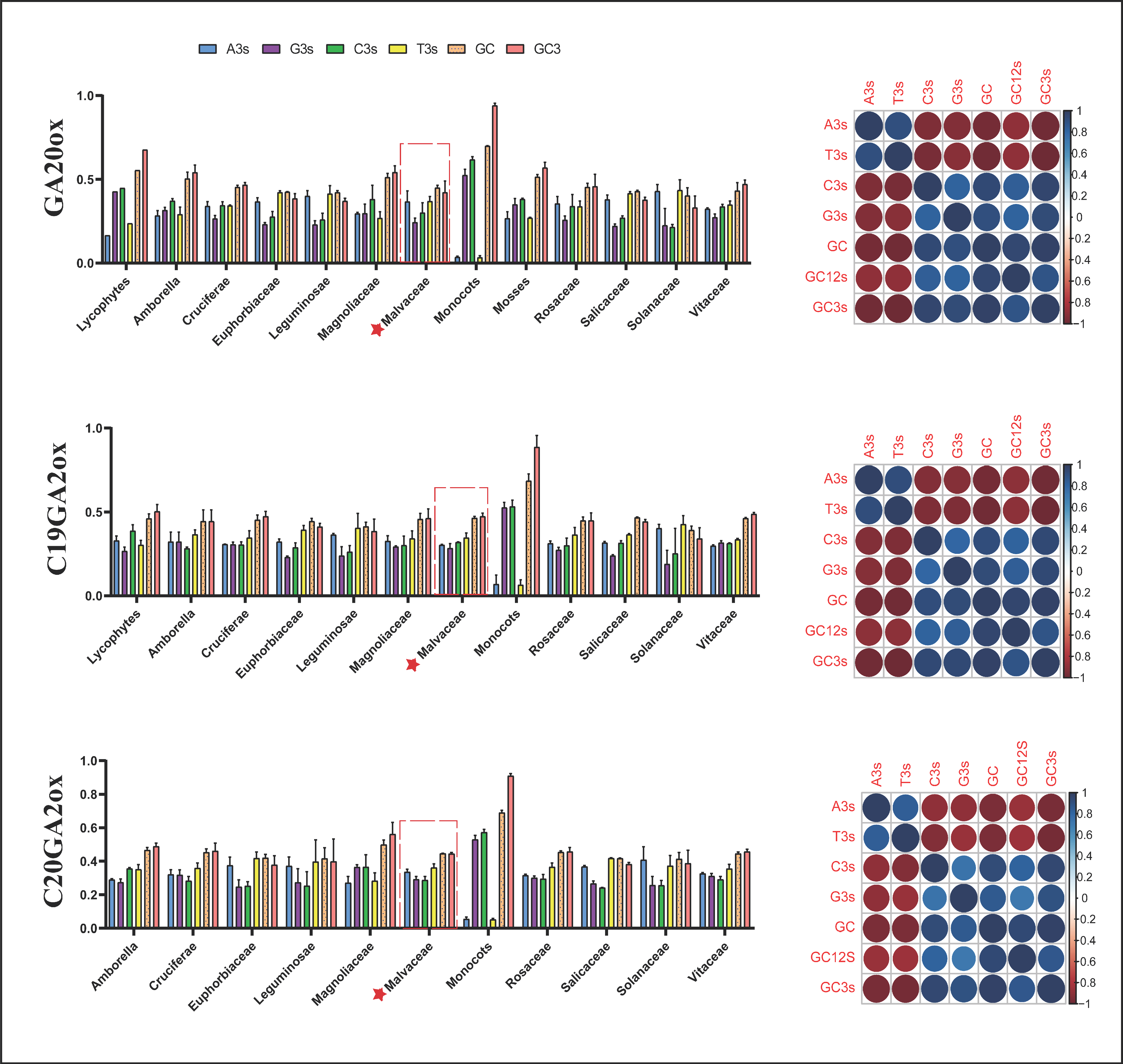
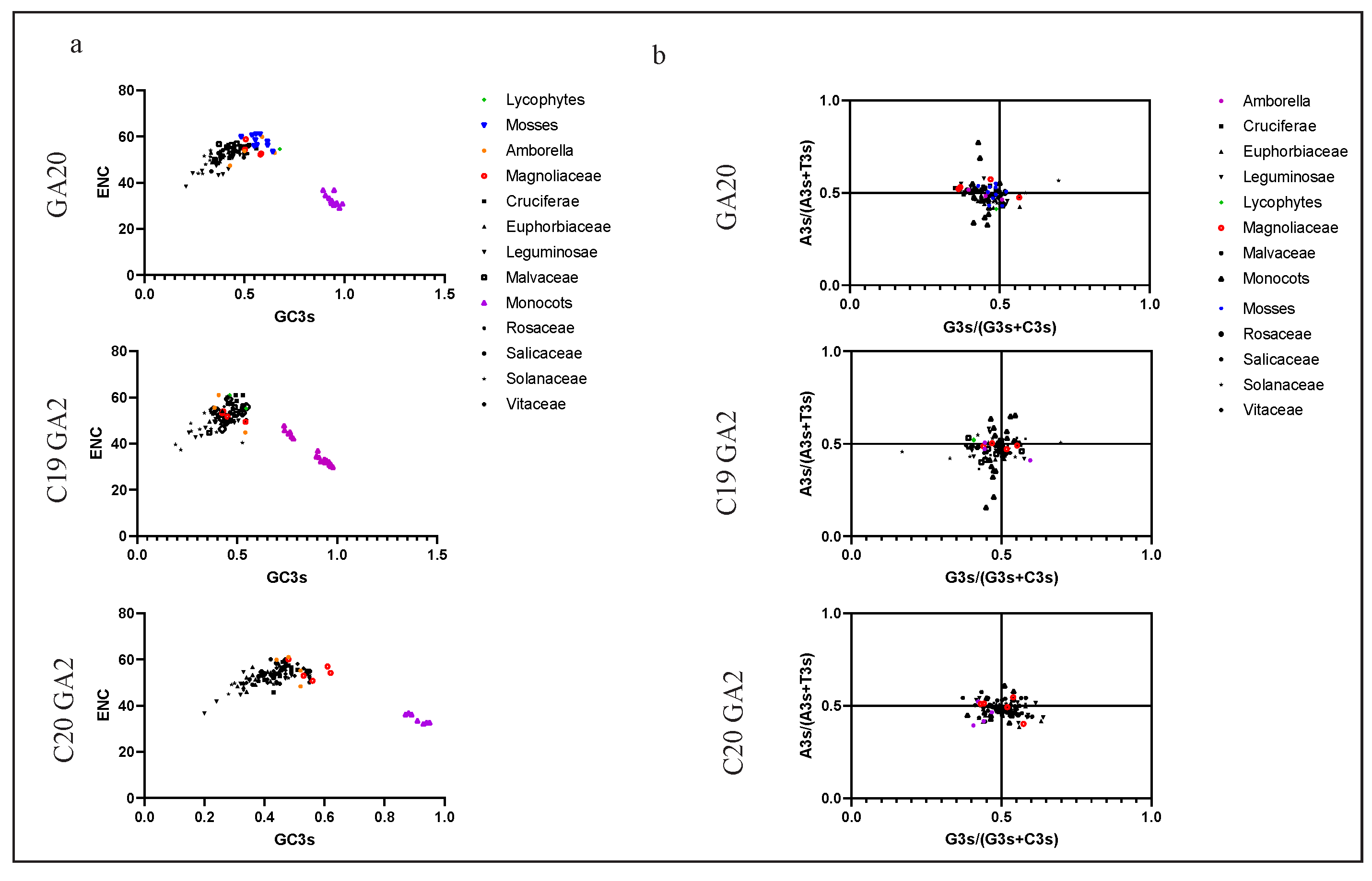
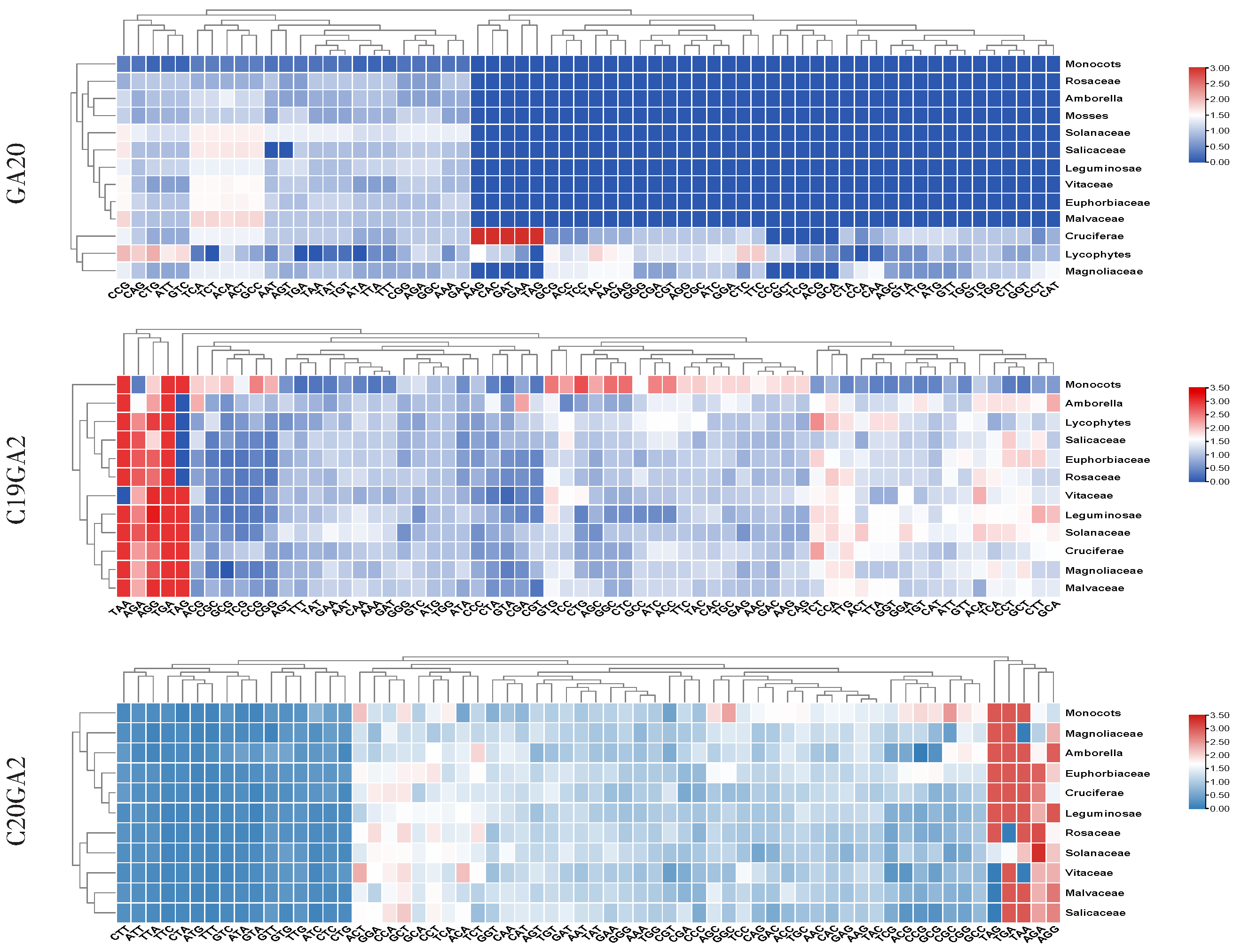
2.4. Codon Usage Pattern Analysis in GAox Genes
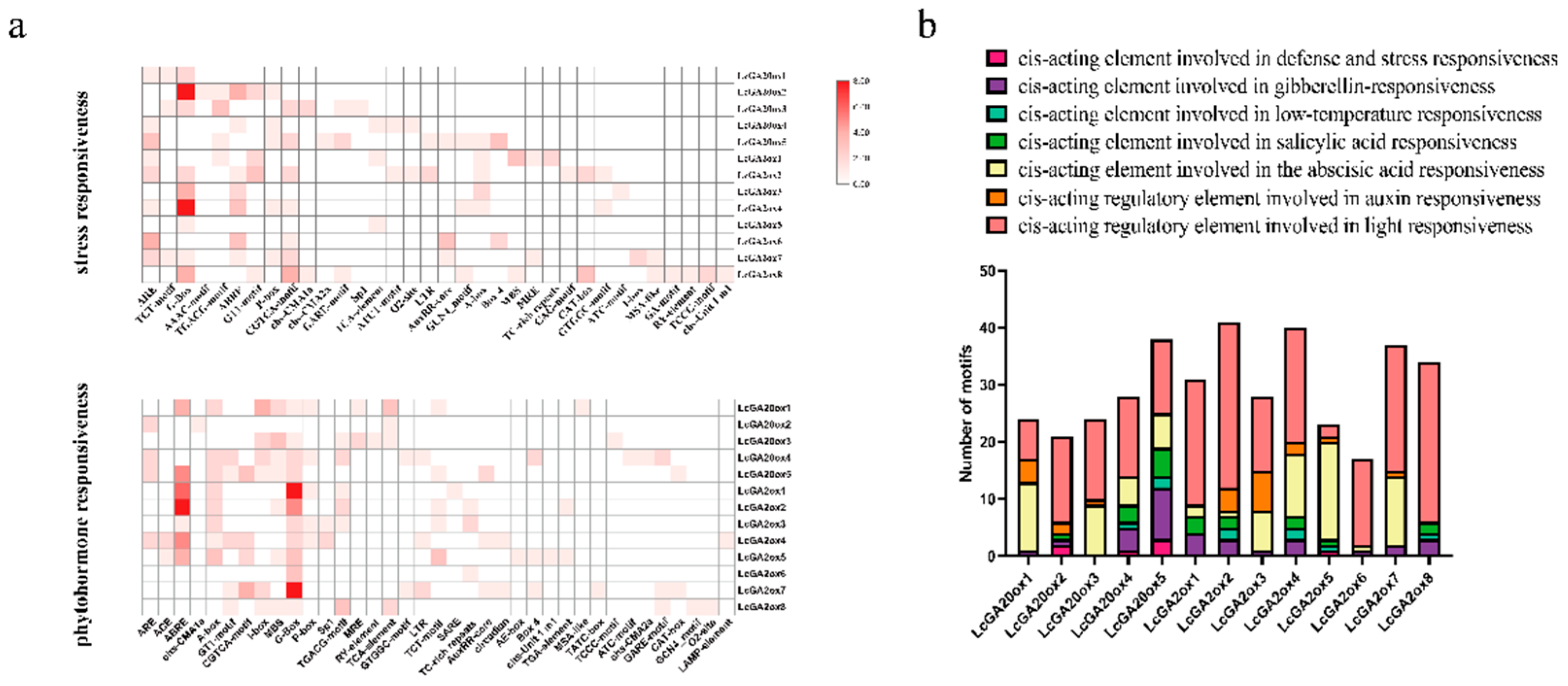
2.5. Cis-Elements in the Promoters of LcGA2ox and LcGA20ox
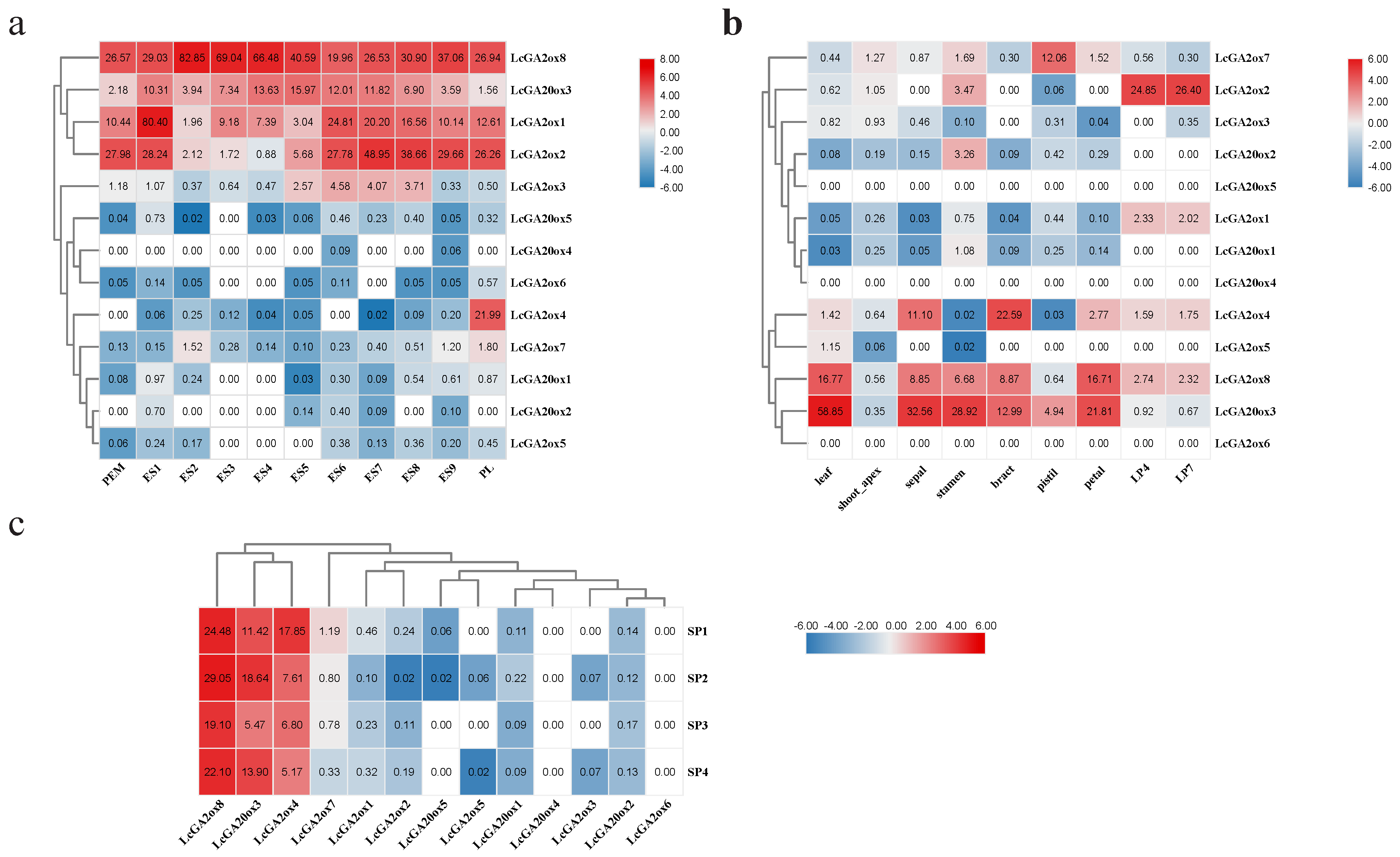
2.6. Analysis of the Expression Patterns of LcGA2ox and LcGA20ox Genes in Different Organs
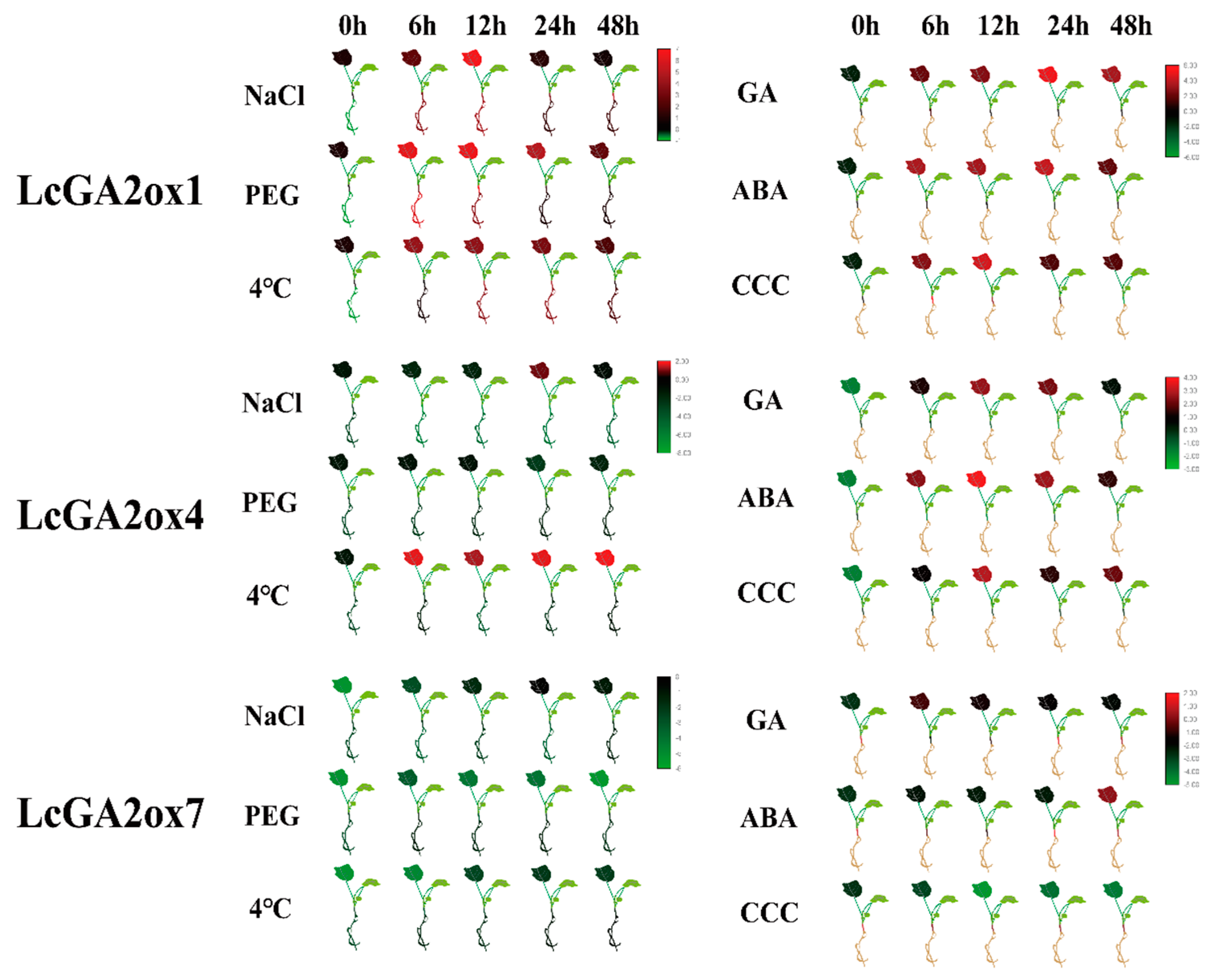
2.7. Effects of Phytohormone and Abiotic Stress Influence the Expression of LcGA2ox and LcGA20ox
3. Discussion
3.1. Gene Identification, Phylogenetic Relationship, Motif Analysis, Gene Structure, and Collinearity Analysis
3.2. Codon Usage Pattern and cis-Element Analysis
3.3. The Spatial Expression Pattern of LcGAox and Their Response to Abiotic and Phytohormone Stress Base on the RNA-seq and qRT-PCR
4. Materials and Methods
4.1. Identification of Gibberellin Oxidase GA2ox, GA20ox Genes in the L. chinense Genome
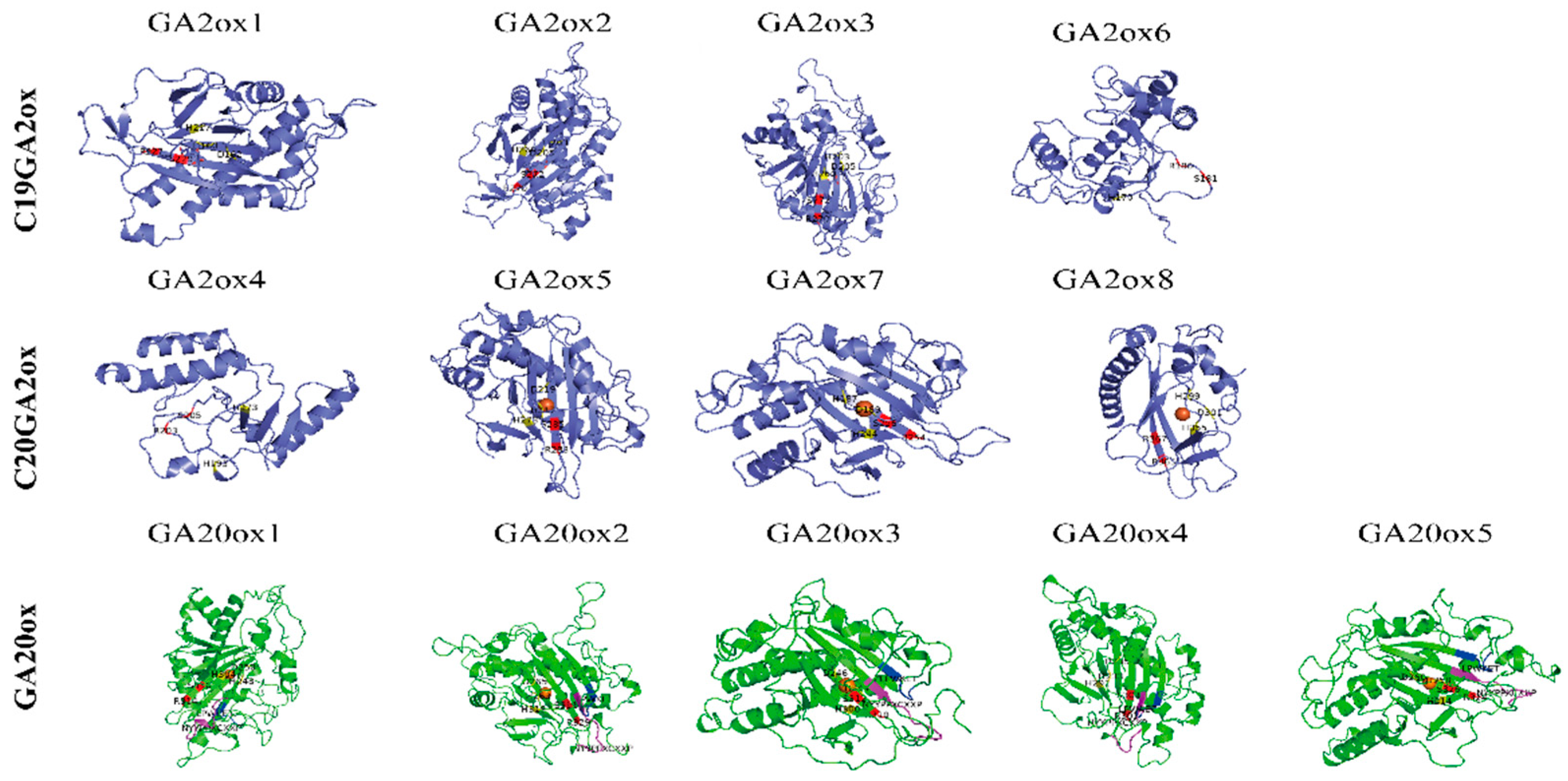
4.2. Phylogenetic Tree Construction and the Analysis of Gene Structure, Conserve Motif in Multiple Species and the 3D Structure Determination
4.3. Codon Usage Pattern Analysis in GAox Genes
4.4. Analysis of the cis-Acting Element in LcGA2ox and LcGA20ox Gene Families
4.5. Analysis of Gene Expression in Organs by the RNA-seq
4.6. Plant Materials Treatment and Expression Analysis by qRT-PCR
4.7. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Webster, A.D. Vigour mechanisms in dwarfing rootstocks for temperate fruit trees. In Proceedings of the 1st International Symposium on Rootstocks for Deciduous Fruit Tree Species, Zaragoza, Spain, 11–14 June 2002; Sanchez, M.A., Webster, A.D., Eds.; International Society for Horticultural Science (ISHS): Leuven, Belgium, 2004; Volumes 1 and 2, pp. 29–41. [Google Scholar]
- Peng, J.R.; Harberd, N.P. The role of GA-mediated signalling in the control of seed germination. Curr. Opin. Plant Biol. 2002, 5, 376–381. [Google Scholar] [CrossRef]
- Zhang, N.; Xie, Y.-D.; Guo, H.J.; Zhao, L.-S.; Xiong, H.-C.; Gu, J.-Y.; Li, J.-H.; Kong, F.-Q.; Sui, L.; Zhao, Z.-W.; et al. Gibberellins regulate the stem elongation rate without affecting the mature plant height of a quick development mutant of winter wheat (Triticum aestivum L.). Plant Physiol. Biochem. 2016, 107, 228–236. [Google Scholar] [CrossRef]
- Wang, G.-L.; Que, F.; Xu, Z.-S.; Wang, F.; Xiong, A.-S. Exogenous gibberellin altered morphology, anatomic and transcriptional regulatory networks of hormones in carrot root and shoot. Bmc Plant Biol. 2015, 15. [Google Scholar] [CrossRef] [Green Version]
- Appleford, N.E.; Lenton, J.R. Gibberellins and leaf expansion in near-isogenic wheat lines containing Rht1 and Rht3 dwarfing alleles. Planta 1991, 183, 229–236. [Google Scholar] [CrossRef]
- Giacomelli, L.; Rota-Stabelli, O.; Masuero, D.; Acheampong, A.K.; Moretto, M.; Caputi, L.; Vrhovsek, U.; Moser, C. Gibberellin metabolism in Vitis vinifera L. during bloom and fruit-set: Functional characterization and evolution of grapevine gibberellin oxidases. J. Exp. Bot. 2013, 64, 4403–4419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, S.; Wang, X.; Zhang, L.; Lin, S.; Liu, D.; Wang, Q.; Cai, S.; El-Tanbouly, R.; Gan, L.; Wu, H.; et al. Identification and characterization of tomato gibberellin 2-oxidases (GA2oxs) and effects of fruit-specific SlGA2ox1 overexpression on fruit and seed growth and development. Hortic. Res. 2016, 3. [Google Scholar] [CrossRef] [Green Version]
- Salazar-Cerezo, S.; Martinez-Montiel, N.; Garcia-Sanchez, J.; Perez-y-Terron, R.; Martinez-Contreras, R.D. Gibberellin biosynthesis and metabolism: A convergent route for plants, fungi and bacteria. Microbiol. Res. 2018, 208, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Kawai, Y.; Ono, E.; Mizutani, M. Evolution and diversity of the 2-oxoglutarate-dependent dioxygenase superfamily in plants. Plant J. 2014, 78, 328–343. [Google Scholar] [CrossRef]
- Sakamoto, T.; Miura, K.; Itoh, H.; Tatsumi, T.; Ueguchi-Tanaka, M.; Ishiyama, K.; Kobayashi, M.; Agrawal, G.K.; Takeda, S.; Abe, K.; et al. An overview of gibberellin metabolism enzyme genes and their related mutants in rice. Plant Physiol. 2004, 134, 1642–1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hedden, P.; Phillips, A.L. Gibberellin metabolism: New insights revealed by the genes. Trends Plant Sci. 2000, 5, 523–530. [Google Scholar] [CrossRef]
- Chen, J.; Hao, Z.; Guang, X.; Zhao, C.; Wang, P.; Xue, L.; Zhu, Q.; Yang, L.; Sheng, Y.; Zhou, Y.; et al. Liriodendron genome sheds light on angiosperm phylogeny and species-pair differentiation. Nat. Plants 2019, 5, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Eshed, Y.; Lippman, Z.B. Revolutions in agriculture chart a course for targeted breeding of old and new crops. Science 2019, 366, 705. [Google Scholar] [CrossRef]
- Hedden, P.; Proebsting, W.M. Genetic analysis of gibberellin biosynthesis. Plant Physiol. 1999, 119, 365–370. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Jiang, H.; Xu, Y.; Wang, Y.; Zhu, L.; Yu, X.; Kong, F.; Zhou, C.; Han, L. Systematic Analysis of Gibberellin Pathway Components in Medicago truncatula Reveals the Potential Application of Gibberellin in Biomass Improvement. Int. J. Mol. Sci. 2020, 21, 7180. [Google Scholar] [CrossRef]
- Do, P.T.; De Tar, J.R.; Lee, H.; Folta, M.K.; Zhang, Z.J. Expression of ZmGA20ox cDNA alters plant morphology and increases biomass production of switchgrass (Panicum virgatum L.). Plant Biotechnol. J. 2016, 14, 1532–1540. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, A.; Ashikari, M.; Ueguchi-Tanaka, M.; Itoh, H.; Nishimura, A.; Swapan, D.; Ishiyama, K.; Saito, T.; Kobayashi, M.; Khush, G.S.; et al. Green revolution: A mutant gibberellin-synthesis gene in rice—New insight into the rice variant that helped to avert famine over thirty years ago. Nature 2002, 416, 701–702. [Google Scholar] [CrossRef]
- Chiang, H.H.; Hwang, I.; Goodman, H.M. Isolation of the Arabidopsis GA4 locus. Plant Cell 1995, 7, 195–201. [Google Scholar] [PubMed] [Green Version]
- Lester, D.R.; Ross, J.J.; Davies, P.J.; Reid, J.B. Mendel’s stem length gene (Le) encodes a gibberellin 3 beta-hydroxylase. Plant Cell 1997, 9, 1435–1443. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Israelsson, M.; Mellerowicz, E.; Chono, M.; Gullberg, J.; Moritz, T. Cloning and overproduction of gibberellin 3-oxidase in hybrid aspen trees. Eff. Gibberellin Homeost. Dev. Plant Physiol. 2004, 135, 221–230. [Google Scholar]
- Reinecke, D.M.; Wickramarathna, A.D.; Ozga, J.A.; Kurepin, L.V.; Jin, A.L.; Good, A.G.; Pharis, R.P. Gibberellin 3-oxidase Gene Expression Patterns Influence Gibberellin Biosynthesis, Growth, and Development in Pea. Plant Physiol. 2013, 163, 929–945. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Zhang, H.; Fan, M.; He, Y.; Guo, P. A mutation in the intron splice acceptor site of a GA3ox gene confers dwarf architecture in watermelon (Citrullus lanatus L.). Sci. Rep. 2020, 10, 14915. [Google Scholar] [CrossRef]
- Martinez-Bello, L.; Moritz, T.; Lopez-Diaz, I. Silencing C-19-GA 2-oxidases induces parthenocarpic development and inhibits lateral branching in tomato plants. J. Exp. Bot. 2015, 66, 5897–5910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, R.; Sun, S.; Wang, H.; Wang, K.; Yu, H.; Zhou, Z.; Xin, P.; Chu, J.; Zhao, T.; Wang, H.; et al. FIS1 encodes a GA2-oxidase that regulates fruit firmness in tomato. Nat. Commun. 2020, 11, 1–12. [Google Scholar] [CrossRef]
- Schomburg, F.M.; Bizzell, C.M.; Lee, D.J.; Zeevaart, J.A.D.; Amasino, R.M. Overexpression of a novel class of gibberellin 2-oxidases decreases gibberellin levels and creates dwarf plants. Plant Cell 2003, 15, 151–163. [Google Scholar] [CrossRef] [Green Version]
- Lv, S.; Yu, D.; Sun, Q.; Jiang, J. Activation of gibberellin 20-oxidase 2 undermines auxin-dependent root and root hair growth in NaCl-stressed Arabidopsis seedlings. Plant Growth Regul. 2018, 84, 225–236. [Google Scholar] [CrossRef]
- Rieu, I.; Ruiz-Rivero, O.; Fernandez-Garcia, N.; Griffiths, J.; Powers, S.J.; Gong, F.; Linhartova, T.; Eriksson, S.; Nilsson, O.; Thomas, S.G.; et al. The gibberellin biosynthetic genes AtGA20ox1 and AtGA20ox2 act, partially redundantly, to promote growth and development throughout the Arabidopsis life cycle. Plant J. 2008, 53, 488–504. [Google Scholar] [CrossRef] [PubMed]
- Shan, C.; Mei, Z.; Duan, J.; Chen, H.; Feng, H.; Cai, W. OsGA2ox5, a Gibberellin Metabolism Enzyme, Is Involved in Plant Growth, the Root Gravity Response and Salt Stress. PLoS ONE 2014, 9, e87110. [Google Scholar] [CrossRef]
- He, H.; Liang, G.; Lu, S.; Wang, P.; Liu, T.; Ma, Z.; Zuo, C.; Sun, X.; Chen, B.; Mao, J. Genome-Wide Identification and Expression Analysis of GA2ox, GA3ox, and GA20ox Are Related to Gibberellin Oxidase Genes in Grape (Vitis vinifera L.). Genes 2019, 10, 680. [Google Scholar] [CrossRef] [Green Version]
- Huang, Y.; Wang, X.; Ge, S.; Rao, G.-Y. Divergence and adaptive evolution of the gibberellin oxidase genes in plants. BMC Evol. Biol. 2015, 15, 207. [Google Scholar] [CrossRef] [Green Version]
- Roach, P.L.; Clifton, I.J.; Fulop, V.; Harlos, K.; Barton, G.J.; Hajdu, J.; Andersson, I.; Schofield, C.J.; Baldwin, J.E. Crystal structure of isopenicillin N synthase is the first from a new structural family of enzymes. Nature 1995, 375, 700–704. [Google Scholar] [CrossRef]
- Honi, U.; Amin, M.R.; Kabir, S.M.T.; Bashar, K.K.; Moniruzzaman, M.; Jahan, R.; Jahan, S.; Haque, M.S.; Islam, S. Genome-wide identification, characterization and expression profiling of gibberellin metabolism genes in jute. BMC Plant Biol. 2020, 20, 306. [Google Scholar] [CrossRef]
- Angellotti, M.C.; Bhuiyan, S.B.; Chen, G.; Wan, X.-F. CodonO: Codon usage bias analysis within and across genomes. Nucleic Acids Res. 2007, 35, W132–W136. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, S.; Yengkhom, S.; Uddin, A. Analysis of codon usage bias of chloroplast genes in Oryza species: Codon usage of chloroplast genes in Oryza species. Planta 2020, 252, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.-H.; Ding, Y.-Z.; He, Y.; Chu, Y.-F.; Zhao, P.; Ma, L.-Y.; Wang, X.-J.; Li, X.-R.; Liu, Y.-S. The Effect of Multiple Evolutionary Selections on Synonymous Codon Usage of Genes in the Mycoplasma bovis Genome. PLoS ONE 2014, 9, e108949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, F.; Zhu, B. Evolutionary analysis of three gibberellin oxidase genesin rice, Arabidopsis, and soybean. Gene 2011, 473, 23–35. [Google Scholar] [CrossRef] [PubMed]
- Babenko, V.N.; Rogozin, I.B.; Mekhedov, S.L.; Koonin, E.V. Prevalence of intron gain over intron loss in the evolution of paralogous gene families. Nucleic Acids Res. 2004, 32, 3724–3733. [Google Scholar] [CrossRef]
- Leister, D. Tandem and segmental gene duplication and recombination in the evolution of plant disease resistance genes. Trends Genet. 2004, 20, 116–122. [Google Scholar] [CrossRef] [Green Version]
- Albert, V.A.; Barbazuk, W.B.; dePamphilis, C.W.; Der, J.P.; Leebens-Mack, J.; Ma, H.; Palmer, J.D.; Rounsley, S.; Sankoff, D.; Schuster, S.C.; et al. The Amborella Genome and the Evolution of Flowering Plants. Science 2013, 342, 1241089. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lescot, M.; Dehais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouze, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Willems, E.; Leyns, L.; Vandesompele, J. Standardization of real-time PCR gene expression data from independent biological replicates. Anal. Biochem. 2008, 379, 127–129. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | GA20ox | GA2ox | GA3ox | GAox | Total |
|---|---|---|---|---|---|
| O. sativa | 5 | 11 | 2 | 3 | 21 |
| A. thaliana | 5 | 7 | 4 | 0 | 16 |
| A. trichopoda | 4 | 5 | 1 | 1 | 11 |
| V. vinifera | 2 | 6 | 3 | 0 | 11 |
| L. chinense | 5 | 8 | 0 | 0 | 13 |
| Gene ID | Gene Name | Accession Number | Position | Location | CDS (bp) | Peptide (aa) | Mw (kDa) | pI | Subcellular Localization |
|---|---|---|---|---|---|---|---|---|---|
| Lchi18432 | LcGA20ox1 | MZ361712 | Chr6 | 41600656-41604092 | 1245 | 415 | 46.976 | 5.93 | Cytoplasm |
| Lchi25954 | LcGA20ox2 | MZ361709 | Chr6 | 38707502-38709176 | 1290 | 430 | 48.367 | 5.97 | Cytoplasm |
| Lchi01764 | LcGA20ox3 | MZ361716 | Chr2 | 73946076-73948031 | 1125 | 375 | 42.662 | 8.36 | Cytoplasm |
| Lchi01394 | LcGA20ox4 | MZ361713 | Chr11 | 56684039-56686083 | 1149 | 383 | 43.514 | 5.78 | Cytoplasm |
| Lchi10458 | LcGA20ox5 | MZ361707 | Chr1 | 79200174-79213961 | 1197 | 399 | 45.202 | 6.1 | Cytoplasm |
| Lchi04083 | LcGA2ox1 | MZ361710 | Chr15 | 63254044-63287148 | 1014 | 338 | 38.043 | 8.18 | Cytoplasm |
| Lchi35277 | LcGA2ox2 | MZ361718 | Contig2730 | 15814-18566 | 1005 | 335 | 37.624 | 7.59 | Cytoplasm |
| Lchi07023 | LcGA2ox3 | MZ361714 | Chr3 | 21930812-21932403 | 996 | 332 | 37.241 | 6.41 | Cytoplasm |
| Lchi12922 | LcGA2ox4 | MZ361717 | Chr1 | 8453778-8461759 | 771 | 257 | 29.734 | 6.34 | Cytoplasm |
| Lchi29789 | LcGA2ox5 | MZ361719 | Chr4 | 67227159-97231797 | 1011 | 337 | 38.25 | 5.83 | Cytoplasm |
| Lchi00133 | LcGA2ox6 | MZ361715 | Chr9 | 2749738-2755769 | 732 | 244 | 27.34 | 8.6 | Cytoplasm |
| Lchi15175 | LcGA2ox7 | MZ361708 | Chr9 | 52008101-52009229 | 978 | 326 | 36.906 | 5.76 | Cytoplasm |
| Lchi18410 | LcGA2ox8 | MZ361711 | Chr6 | 42633756-42642514 | 1272 | 424 | 47.688 | 5.31 | Cytoplasm |
| Duplicated GAox Gene Pairs | Ka | Ks | Ka/Ks | ||
|---|---|---|---|---|---|
| Lc/Lc | LcGA2ox1 | LcGA2ox3 | 0.1752 | 0.7673 | 0.2283 |
| LcGA2ox3 | LcGA2ox6 | 0.5347 | 1.5333 | 0.3487 | |
| Lc/At | LcGA2ox1 | AT1G30040 (AtGA2ox2) | 0.7197 | 0.9484 | 0.7589 |
| LcGA20ox3 | AT1G44090 (AtGA20ox5) | 0.5312 | 1.0585 | 0.5018 | |
| LcGA2ox8 | AT5G58660 | 0.6419 | 0.8638 | 0.7431 | |
| Lc/Vv | LcGA2ox4 | VIT_219s0177g00020 | 1.4737 | 1.5654 | 0.9414 |
| LcGA20ox4 | VIT_202s0234g00010 | 0.2196 | 0.686 | 0.3201 | |
| LcGA2ox1 | VIT_210s0003g03490 | 0.5232 | 0.6875 | 0.761 | |
| LcGA20ox3 | VIT_218s0001g01390 | 0.2933 | 0.525 | 0.5587 | |
| LcGA20ox3 | VIT_204s0044g01650 | 0.3052 | 0.7457 | 0.4093 | |
| LcGA2ox8 | VIT_206s0004g06790 | 0.5502 | 0.8525 | 0.6454 | |
| LcGA2ox6 | VIT_205s0077g00520 | 0.5477 | 1.3039 | 0.42 | |
| LcGA2ox6 | VIT_207s0005g01920 | 0.5626 | 1.0286 | 0.547 | |
| Lc/Os | LcGA2ox1 | OsKitaake01g330800 | 0.775 | 0.9395 | 0.8249 |
| LcGA2ox1 | OsKitaake01g077300 | 0.77 | 1.2028 | 0.6402 | |
| LcGA20ox3 | OsKitaake01g424900 | 0.351 | 0.9514 | 0.3689 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, L.; Wang, P.; Hao, Z.; Lu, Y.; Xue, G.; Cao, Z.; Qu, H.; Cheng, T.; Shi, J.; Chen, J. Gibberellin Oxidase Gene Family in L. chinense: Genome-Wide Identification and Gene Expression Analysis. Int. J. Mol. Sci. 2021, 22, 7167. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137167
Hu L, Wang P, Hao Z, Lu Y, Xue G, Cao Z, Qu H, Cheng T, Shi J, Chen J. Gibberellin Oxidase Gene Family in L. chinense: Genome-Wide Identification and Gene Expression Analysis. International Journal of Molecular Sciences. 2021; 22(13):7167. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137167
Chicago/Turabian StyleHu, Lingfeng, Pengkai Wang, Zhaodong Hao, Ye Lu, Guoxia Xue, Zijian Cao, Haoxian Qu, Tielong Cheng, Jisen Shi, and Jinhui Chen. 2021. "Gibberellin Oxidase Gene Family in L. chinense: Genome-Wide Identification and Gene Expression Analysis" International Journal of Molecular Sciences 22, no. 13: 7167. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22137167