
Growth Factors, Reactive Oxygen Species, and Metformin—Promoters of the Wound Healing Process in Burns?

 ,
, {kind=link}
{kind=link}
Abstract
:1. Introduction
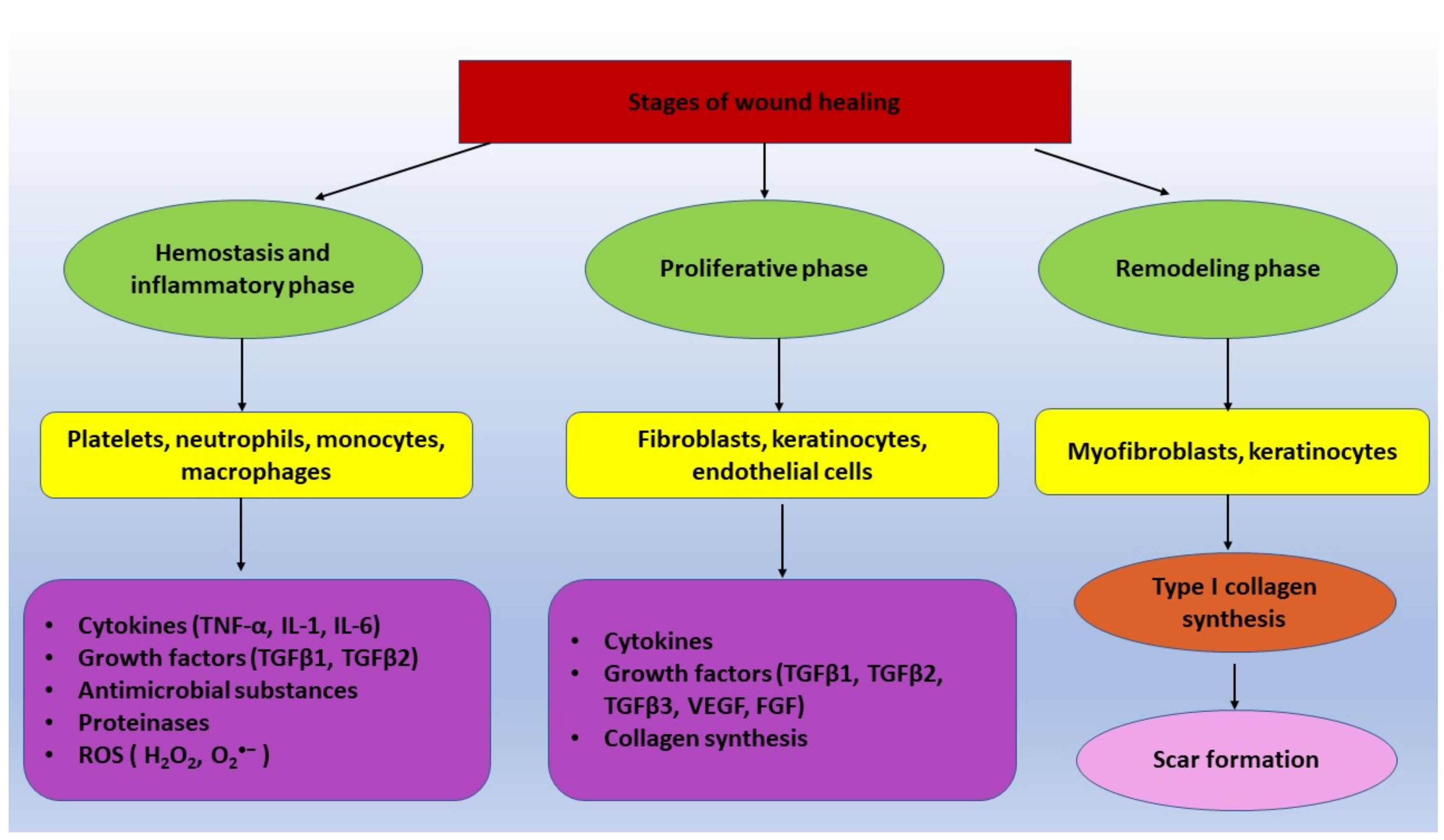
2. Wound Healing Process-General Aspects
3. The Impact of Growth Factors on the Healing Process and Their Clinical Applications
3.1. Fibroblast Growth Factor Family and Wound Healing
3.2. Epidermal Growth Factor and Wound Healing
3.3. Transforming Growth Factor-Beta Family and Wound Healing
3.4. Vascular Endothelial Growth Factor Family and Wound Healing
3.5. Insulin-Like Growth Factor Family and Wound Healing
3.6. Keratinocyte Growth Factor and Wound Healing
3.7. Granulocyte-Macrophage Colony Stimulating Factor and Wound Healing
3.8. Platelet-Derived Growth Factor Family and Wound Healing
4. Oxygen Metabolism and Wound Healing Process
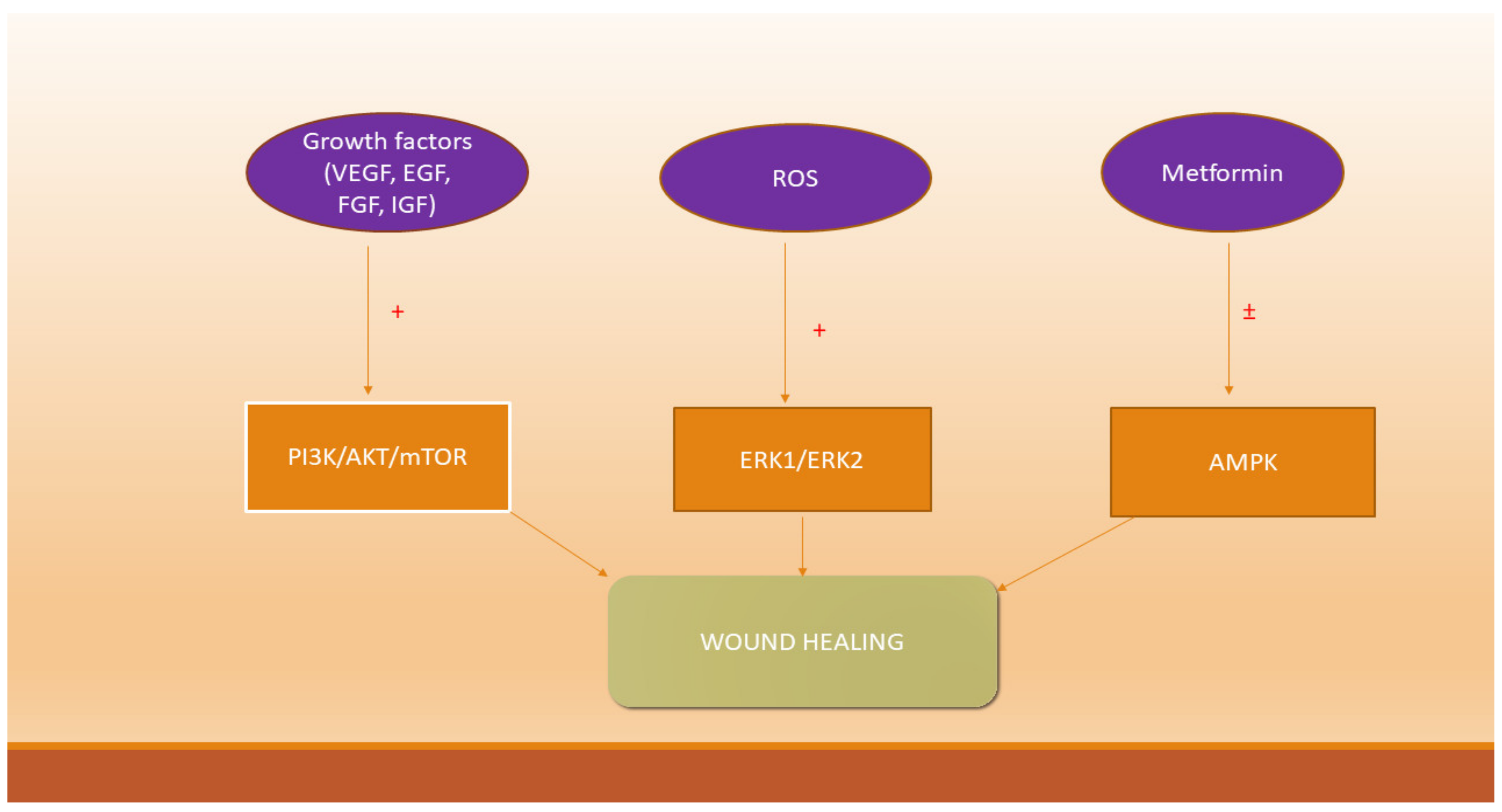
5. Growth Factors, Reactive Oxygen Species and Signaling Pathways Involved in Wound Healing
6. Metformin and Wound Healing
7. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Davies, K.; Johnson, E.L.; Hollén, L.; Jones, H.M.; Lyttle, M.D.; Maguire, S.; Kemp, A.M.P. Incidence of medically attended paediatric burns across the UK. Inj. Prev. 2020, 26, 24–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asena, M.; Aydin Ozturk, P.; Ozturk, U. Sociodemographic and culture results of paediatric burns. Int. Wound J. 2020, 17, 132–136. [Google Scholar] [CrossRef] [PubMed]
- Said, A.; Wahid, F.; Bashir, K.; Rasheed, H.M.; Khan, T.; Hussain, Z.; Siraj, S. Sauromatum guttatum extract promotes wound healing and tissue regeneration in a burn mouse model via up-regulation of growth factors. Pharm. Biol. 2019, 57, 736–743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodarzi, P.; Falahzadeh, K.; Nematizadeh, M.; Farazandeh, P.; Payab, M.; Larijani, B.; Beik, A.T.; Arjmand, B. Tissue engineered skin substitutes. Adv. Exp. Med. Biol. 2018, 1107, 143–188. [Google Scholar]
- Velnar, T.; Bailey, T.; Smrkolj, V. The wound healing process: An overview of the cellular and molecular mechanisms. J. Int. Med. Res. 2009, 37, 1528–1542. [Google Scholar] [CrossRef] [PubMed]
- Jimi, S.; Takagi, S.; De Francesco, F.; Miyazaki, M.; Saparov, A. Acceleration of skin wound-healing reactions by autologous micrograft tissue suspension. Medicina 2020, 56, 321. [Google Scholar] [CrossRef]
- Felician, F.F.; Yu, R.H.; Li, M.Z.; Li, C.J.; Chen, H.Q.; Jiang, Y.; Tang, T.; Qi, W.Y.; Xu, H.M. The wound healing potential of collagen peptides derived from the jellyfish Rhopilema esculentum. Chin. J. Traumatol. 2019, 22, 12–20. [Google Scholar] [CrossRef]
- Zhu, Y.; Wang, Y.; Jia, Y.; Xu, J.; Chai, Y. Roxadustat promotes angiogenesis through HIF-1alpha/VEGF/VEGFR2 signaling and accelerates cutaneous wound healing in diabetic rats. Wound Repair Regen. 2019, 27, 324–334. [Google Scholar] [CrossRef]
- Nesi-Reis, V.; Lera-Nonose, D.S.S.L.; Oyama, J.; Silva-Lalucci, M.P.P.; Demarchi, I.G.; Aristides, S.M.A.; Teixeira, J.J.V.; Silveira, T.G.V.; Lonardoni, M.V.C. Contribution of photodynamic therapy in wound healing: A systematic review. Photodiagnosis Photodyn. Ther. 2018, 21, 294–305. [Google Scholar] [CrossRef] [PubMed]
- Vijayan, A.; James, P.P.; Nanditha, C.K.; Kumar, G.S.V. Multiple cargo deliveries of growth factors and antimicrobial peptide using biodegradable nanopolymer as a potential wound healing system. Int. J. Nanomed. 2019, 14, 2253–2263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, P.H.; Huang, B.S.; Horng, H.C.; Yeh, C.C.; Chen, Y.J. Wound healing. J. Chin. Med. Assoc. 2018, 81, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Kang, X.; Jin, L.; Bai, J.; Liu, W.; Wang, Z. Stimulation of wound healing using bioinspired hydrogels with basic fibroblast growth factor (bFGF). Int. J. Nanomed. 2018, 4, 3897–3906. [Google Scholar] [CrossRef] [Green Version]
- Garraud, O.; Hozzein, W.N.; Badr, G. Wound healing: Time to look for intelligent, natural immunological approaches? BMC Immunol. 2017, 21, 23. [Google Scholar] [CrossRef] [Green Version]
- Elbialy, Z.I.; Atiba, A.; Abdelnaby, A.; Al-Hawary, I.I.; Elsheshtawy, A.; El-Serehy, H.A.; Abdel-Daim, M.M.; Fadl, S.E.; Assar, D.H. Collagen extract obtained from Nile tilapia (Oreochromis niloticus L.) skin accelerates wound healing in rat model via up regulating VEGF, bFGF, and alpha-SMA genes expression. BMC Vet. Res. 2020, 24, 352. [Google Scholar] [CrossRef] [PubMed]
- Dam, D.H.M.; Paller, A.S. Gangliosides in diabetic wound healing. Prog. Mol. Biol. Transl. Sci. 2018, 156, 229–239. [Google Scholar] [PubMed]
- Eroglu, Z.T.; Kurtis, B.; Altug, H.A.; Sahin, S.; Tuter, G.; Baris, E. Effect of topical ozonetherapy on gingival wound healing in pigs: Histological and immuno-histochemical analysis. J. Appl. Oral Sci. 2018, 27, e20180015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, G.; Ceilley, R. Chronic wound healing: A review of current management and treatments. Adv. Ther. 2017, 34, 599–610. [Google Scholar] [CrossRef] [Green Version]
- Reinke, J.M.; Sorg, H. Wound repair and regeneration. Eur. Surg. Res. 2012, 49, 35–43. [Google Scholar] [CrossRef]
- Sorg, H.; Tilkorn, D.J.; Hager, S.; Hauser, J.; Mirastschijski, U. Skin wound healing: An update on the current knowledge and concepts. Eur. Surg. Res. 2017, 58, 81–94. [Google Scholar] [CrossRef]
- Hesketh., M.; Sahin, K.B.; West, Z.E.; Murray, R.Z. Macrophage phenotypes regulate scar formation and chronic wound healing. Int. J. Mol. Sci. 2017, 18, 1545. [Google Scholar] [CrossRef] [Green Version]
- Martin, P.; Nunan, R. Cellular and molecular mechanisms of repair in acute and chronic wound healing. Br. J. Dermatol. 2015, 173, 370–378. [Google Scholar] [CrossRef] [PubMed]
- Berksoy Hayta, S.; Durmuş, K.; Altuntaş, E.E.; Yildiz, E.; Hisarciklıo, M.; Akyol, M. The reduction in inflammation and impairment in wound healing by using strontium chloride hexahydrate. Cutan. Ocul. Toxicol. 2018, 37, 24–28. [Google Scholar] [CrossRef]
- Gonzalez, A.C.D.O.; Costa, T.F.; Andrade, Z.D.A.; Medrado, A.R.A.P. Wound healing—A literature review. An. Bras. Dermatol. 2016, 91, 614–620. [Google Scholar] [CrossRef] [Green Version]
- Zhu, G.; Wang, Q.; Lu, S.; Niu, Y. Hydrogen Peroxide: A potential wound therapeutic target. Med. Princ. Pract. 2017, 26, 301–308. [Google Scholar] [CrossRef]
- Kurek-Górecka, A.; Komosinska-Vassev, K.; Rzepecka-Stojko, A.; Olczyk, P. Bee venom in wound healing. Molecules 2020, 26, 148. [Google Scholar] [CrossRef]
- Parfejevs, V.; Debbache, J.; Shakhova, O.; Schaefer, S.M.; Glausch, M.; Wegner, M.; Suter, U.; Riekstina, U.; Werner, S.; Sommer, L. Injury-activated glial cells promote wound healing of the adult skin in mice. Nat. Commun. 2018, 9, 236. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Liu, W.J.; Xue, P.; Ma, Y.; Zhang, L.Q.; Zhu, B.; Qi, M.; Li, L.Y.; Zhang, Y.J.; Wang, Q.T.; et al. Autophagy promotes MSC-mediated vascularization in cutaneous wound healing via regulation of VEGF secretion. Cell Death Dis. 2018, 9, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asai, E.; Yamamoto, M.; Ueda, K.; Waguri, S. Spatiotemporal alterations of autophagy marker LC3 in rat skin fibroblasts during wound healing process. Fukushima J. Med. Sci. 2018, 64, 15–22. [Google Scholar] [CrossRef] [Green Version]
- Wathoni, N.; Rusdiana, T.; Hasanah, A.N.; Muhtadi, A.; Pratiwi, E.D.; Mahmudah, R.; Mohammed, A.F.A.; Okajima, M.; Kaneko, T.; Arima, H. Sacran hydrogel film containing keratinocyte growth factor accelerates wound healing by stimulating fibroblast migration and re-epithelization. Chem. Pharm. Bull. 2019, 67, 849–854. [Google Scholar] [CrossRef] [Green Version]
- Marconi, G.D.; Fonticoli, L.; Rajan, T.S.; Lanuti, P.; Della Rocca, Y.; Pierdomenico, S.D.; Trubiani, O.; Pizzicannella, J.; Diomede, F. Transforming growth factor-beta1 and human gingival fibroblast-to-myofibroblast differentiation: Molecular and morphological modifications. Front. Physiol. 2021, 12, 676512. [Google Scholar] [CrossRef]
- Koike, Y.; Yozaki, M.; Utani, A.; Murota, H. Fibroblast growth factor 2 accelerates the epithelial-mesenchymal transition in keratinocytes during wound healing process. Sci. Rep. 2020, 10, 18545. [Google Scholar] [CrossRef]
- Atiba, A.; Nishimura, M.; Kakinuma, S.; Hiraoka, T.; Goryo, M.; Shimada, Y.; Ueno, H.; Uzuka, Y. Aloe vera oral administration accelerates acute radiation-delayed wound healing by stimulating transforming growth factor-β and fibroblast growth factor production. Am. J. Surg. 2011, 201, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Shabbir, A.; Cox, A.; Rodriguez-Menocal, L.; Salgado, M.; Van Badiavas, E. Mesenchymal stem cell exosomes induce proliferation and migration of normal and chronic wound fibroblasts, and enhance angiogenesis in vitro. Stem Cells Dev. 2015, 24, 1635–1647. [Google Scholar] [CrossRef] [PubMed]
- Ning, J.; Zhao, H.; Chen, B.; Mi, E.Z.; Yang, Z.; Qing, W.; Lam, K.W.J.; Yi, B.; Chen, Q.; Gu, J.; et al. Argon mitigates impaired wound healing process and enhances wound healing in vitro and in vivo. Theranostics 2019, 9, 477–490. [Google Scholar] [CrossRef] [PubMed]
- Casado-Díaz, A.; Quesada-Gómez, J.M.; Dorado, G. Extracellular vesicles derived from mesenchymal stem cells (MSC) in regenerative medicine: Applications in skin wound healing. Front. Bioeng. Biotechnol. 2020, 8, 146. [Google Scholar] [CrossRef] [Green Version]
- Park, J.W.; Hwang, S.R.; Yoon, I.S. Advanced growth factor delivery systems in wound management and skin regeneration. Molecules 2017, 22, 1259. [Google Scholar] [CrossRef] [Green Version]
- Hsu, C.; Chang, J. Clinical implications of growth factors in flexor tendon wound healing. J. Hand Surg. 2004, 29, 551–563. [Google Scholar] [CrossRef]
- Wöltje, M.; Böbel, M.; Bienert, M.; Neuss, S.; Aibibu, D.; Cherif, C. Functionalized silk fibers from transgenic silkworms for wound healing applications: Surface presentation of bioactive epidermal growth factor. J. Biomed. Mater. Res. A 2018, 106, 2643–2652. [Google Scholar] [CrossRef]
- Pikuła, M.; Langa, P.; Kosikowska, P.; Trzonkowski, P. Stem cells and growth factors in wound healing. Postepy Hig. Med. Dosw. 2015, 69, 874–885. [Google Scholar] [CrossRef] [PubMed]
- Zarei, F.; Soleimaninejad, M. Role of growth factors and biomaterials in wound healing. Artif. Cells Nanomed. Biotechnol. 2018, 46, 906–911. [Google Scholar] [CrossRef] [Green Version]
- Titan, A.L.; Foster, D.S.; Chang, J.; Longaker, M.T. Flexor tendon: Development, healing, adhesion formation, and contributing growth factors. Plast. Reconstr. Surg. 2019, 144, 639e–647e. [Google Scholar] [CrossRef] [PubMed]
- Werner, S.; Grose, R. Regulation of wound healing by growth factors and cytokines. Physiol. Rev. 2003, 83, 835–870. [Google Scholar] [CrossRef]
- Cowin, A.J.; Hatzirodos, N.; Holding, C.A.; Dunaiski, V.; Harries, R.H.; Rayner, T.E.; Fitridge, R.; Cooter, R.D.; Schultz, G.S.; Belford, D.A. Effect of healing on the expression of transforming growth factor beta(s) and their receptors in chronic venous leg ulcers. J. Investig. Dermatol. 2001, 117, 1282–1289. [Google Scholar] [CrossRef] [PubMed]
- Ishihara, J.; Ishihara, A.; Starke, R.D.; Peghaire, C.R.; Smith, K.E.; McKinnon, T.A.J.; Tabata, Y.; Sasaki, K.; White, M.J.V.; Fukunaga, K.; et al. The heparin binding domain of von Willebrand factor binds to growth factors and promotes angiogenesis in wound healing. Blood 2019, 13, 2559–2569. [Google Scholar] [CrossRef]
- Berlanga-Acosta, J.; Fernández-Montequín, J.; Valdés-Pérez, C.; Savigne-Gutiérrez, W.; Mendoza-Marí, Y.; García-Ojalvo, A.; Falcón-Cama, V.; García Del Barco-Herrera, D.; Fernández-Mayola, M.; Pérez-Saad, H.; et al. Diabetic foot ulcers and epidermal growth factor: Revisiting the local delivery route for a successful outcome. Biomed. Res. Int. 2017, 2017, 2923759. [Google Scholar] [CrossRef]
- Hui, Q.; Jin, Z.; Li, X.; Liu, C.; Wang, X. FGF Family: From drug development to clinical application. Int. J. Mol. Sci. 2018, 19, 1875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Y.H.; Zhu, Y.T.; Ding, J.; Zhou, F.Y.; Xue, J.X.; Jung, J.H.; Li, Z.J.; Gao, W.Y. Distribution of fibroblast growth factors and their roles in skin fibroblast cell migration. Mol. Med. Rep. 2016, 14, 3336–3342. [Google Scholar] [CrossRef]
- Maddaluno, L.; Urwyler, C.; Werner, S. Fibroblast growth factors: Key players in regeneration and tissue repair. Development 2017, 144, 4047–4060. [Google Scholar] [CrossRef] [Green Version]
- El Agha, E.; Kosanovic, D.; Schermuly, R.T.; Bellusci, S. Role of fibroblast growth factors in organ regeneration and repair. Semin. Cell Dev. Biol. 2016, 53, 76–84. [Google Scholar] [CrossRef]
- Rouabhia, M.; Park, H.; Meng, S.; Derbali, H.; Zhang, Z. Electrical stimulation promotes wound healing by enhancing dermal fibroblast activity and promoting myofibroblast transdifferentiation. PLoS ONE 2013, 8, e71660. [Google Scholar]
- Yang, J.; Qiang, W.; Ren, S.; Yi, S.; Li, J.; Guan, L.; Du, L.; Guo, Y.; Hu, H.; Li, H.; et al. High-efficiency production of bioactive oleosin-basic fibroblast growth factor in A. thaliana and evaluation of wound healing. Gene 2018, 10, 69–76. [Google Scholar] [CrossRef]
- Behr, B.; Panetta, N.J.; Longaker, M.T.; Quarto, N. Different endogenous threshold levels of Fibroblast Growth Factor-ligands determine the healing potential of frontal and parietal bones. Bone 2010, 47, 281–294. [Google Scholar] [CrossRef]
- Carvalho, E.D.; Rosa, R.H.; Pereira, F.M.; Anbinder, A.L.; Mello, I.; Habitante, S.M.; Raldi, D.P. Effects of diode laser irradiation and fibroblast growth factor on periodontal healing of replanted teeth after extended extra-oral dry time. Dent. Traumatol. 2017, 33, 91–99. [Google Scholar] [CrossRef]
- Akita, S.; Akino, K.; Tanaka, K.; Anraku, K.; Hirano, A. A basic fibroblast growth factor improves lower extremity wound healing with a porcine-derived skin substitute. J. Trauma Acute Care Surg. 2008, 64, 809–815. [Google Scholar] [CrossRef]
- Zhang, J.B.; Li, H.; Zhang, L.; Wang, J.L. Observation of curative effect of recombinant human basic fibroblast growth factor combined with compound polymyxin B ointment and local application of insulin on wound healing of deep second-degree burn in diabetes mellitus: A randomized study. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 5987–5993. [Google Scholar]
- Hom, D.B.; Unger, G.M.; Pernell, K.J.; Manivel, J.C. Improving surgical wound healing with basic fibroblast growth factor after radiation. Laryngoscope 2005, 115, 412–422. [Google Scholar] [CrossRef]
- Peng, J.; Zhao, H.; Tu, C.; Xu, Z.; Ye, L.; Zhao, L.; Gu, Z.; Zhao, D.; Zhang, J.; Feng, Z. In situ hydrogel dressing loaded with heparin and basic fibroblast growth factor for accelerating wound healing in rat. Mater. Sci. Eng. C 2020, 116, 111169. [Google Scholar] [CrossRef]
- Santa Maria, P.L.; Redmond, S.L.; Atlas, M.D.; Ghassemifar, R. Keratinocyte growth factor 1, fibroblast growth factor 2 and 10 in the healing tympanic membrane following perforation in rats. J. Mol. Histol. 2011, 42, 47–58. [Google Scholar] [CrossRef]
- Ramos-Gonzalez, G.; Wittig, O.; Diaz-Solano, D.; Salazar, L.; Ayala-Grosso, C.; Cardier, J.E. Evaluation of epithelial progenitor cells and growth factors in a preclinical model of wound healing induced by mesenchymal stromal cells. Biosci. Rep. 2020, 40, BSR20200461. [Google Scholar] [CrossRef]
- Xie, J.L.; Bian, H.N.; Qi, S.H.; Chen, H.D.; Li, H.D.; Xu, Y.B.; Li, T.Z.; Liu, X.S.; Liang, H.Z.; Xin, B.R.; et al. Basic fibroblast growth factor (bFGF) alleviates the scar of the rabbit ear model in wound healing. Wound Repair Regen. 2008, 16, 576–581. [Google Scholar] [CrossRef]
- Akita, S.; Akino, K.; Imaizumi, T.; Hirano, A. Basic fibroblast growth factor accelerates and improves second-degree burn wound healing. Wound Repair Regen. 2008, 16, 635–641. [Google Scholar] [CrossRef]
- Numata, Y.; Terui, T.; Okuyama, R.; Hirasawa, N.; Sugiura, Y.; Miyoshi, I.; Watanabe, T.; Kuramasu, A.; Tagami, H.; Ohtsu, H. The accelerating effect of histamine on the cutaneous wound-healing process through the action of basic fibroblast growth factor. J. Investig. Dermatol. 2006, 126, 1403–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, W.; Han, Q.; Lin, H.; Gao, Y.; Sun, W.; Zhao, Y.; Wang, B.; Chen, B.; Xiao, Z.; Dai, J. Improved neovascularization and wound repair by targeting human basic fibroblast growth factor (bFGF) to fibrin. J. Mol. Med. 2008, 86, 1127–1138. [Google Scholar] [CrossRef]
- Guo, Y.; Xu, B.; Wang, Y.; Li, Y.; Si, H.; Zheng, X.; Chen, Z.; Chen, F.; Fan, D. Dramatic promotion of wound healing using a recombinant human-like collagen and bFGF cross-linked hydrogel by transglutaminase. J. Biomater. Sci. Polym. Ed. 2019, 30, 1591–1603. [Google Scholar] [CrossRef]
- Hardwicke, J.; Schmaljohann, D.; Boyce, D.; Thomas, D. Epidermal growth factor therapy and wound healing—Past, present and future perspectives. Surgeon 2008, 6, 172–177. [Google Scholar] [CrossRef]
- Wu, M.; Ruan, J.; Zhong, B. Progress in human epidermal growth factor research. Sheng Wu Gong Cheng Xue Bao 2020, 36, 2813–2823. [Google Scholar]
- Liu, Y.; Liu, Z.G. Role of epidermal growth factor and its receptor family in ocular surface wound healing. Zhonghua Yan Ke Za Zhi 2007, 43, 953–956. [Google Scholar] [PubMed]
- Kim, H.; Kong, W.H.; Seong, K.Y.; Sung, D.K.; Jeong, H.; Kim, J.K.; Yang, S.Y.; Hahn, S.K. Hyaluronate-epidermal growth factor conjugate for skin wound healing and regeneration. Biomacromolecules 2016, 17, 3694–3705. [Google Scholar] [CrossRef]
- Yumusak, N.; Yavuz, U.; Sarikaya, B.; Yucel, G. Effects of epidermal growth factor on reduction of the formation of thrombus and vessel wall healing in an experimental rat model. Bratisl. Lek. Listy 2017, 118, 752–758. [Google Scholar] [CrossRef] [PubMed]
- Adam, A.B.; Özdamar, M.Y.; Esen, H.H.; Günel, E. Local effects of epidermal growth factor on the wound healing in esophageal anastomosis: An experimental study. Int. J. Pediatr. Otorhinolaryngol. 2017, 99, 8–12. [Google Scholar] [CrossRef]
- Techapichetvanich, T.; Wanitphakdeedecha, R.; Iamphonrat, T.; Phothong, W.; Eimpunth, S.; Jane, H.I.; Manuskiatti, W. The effects of recombinant human epidermal growth factor containing ointment on wound healing and post inflammatory hyperpigmentation prevention after fractional ablative skin resurfacing: A split-face randomized controlled study. J. Cosmet. Dermatol. 2018, 17, 756–761. [Google Scholar] [CrossRef] [PubMed]
- Berlanga-Acosta, J.; Camacho-Rodríguez, H.; Mendoza-Marí, Y.; Falcón-Cama, V.; García-Ojalvo, A.; Herrera-Martínez, L.; Guillén-Nieto, G. Epidermal growth factor in healing diabetic foot ulcers: From gene expression to tissue healing and systemic biomarker circulation. Medicc Rev. 2020, 22, 24–31. [Google Scholar]
- Lu, K.J.; Wang, W.; Xu, X.L.; Jin, F.Y.; Qi, J.; Wang, X.J.; Kang, X.Q.; Zhu, M.L.; Huang, Q.L.; Yu, C.H.; et al. A dual deformable liposomal ointment functionalized with retinoic acid and epidermal growth factor for enhanced burn wound healing therapy. Biomater. Sci. 2019, 7, 2372–2382. [Google Scholar] [CrossRef]
- Mohanty, C.; Pradhan, J. A human epidermal growth factor-curcumin bandage bioconjugate loaded with mesenchymal stem cell for in vivo diabetic wound healing. Mater. Sci. Eng. C 2020, 111, 110751. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; Basak, P.; Prasad Sinha, B.; Maurye, P.; Kumar Jaiswal, K.; Das, P.; Kumar Mandal, T. Anti-inflammatory effect of epidermal growth factor conjugated silk fibroin immobilized polyurethane ameliorates diabetic burn wound healing. Int. J. Biol. Macromol. 2020, 143, 1009–1032. [Google Scholar] [CrossRef]
- Kao, C.W.; Tseng, Y.Y.; Liu, K.S.; Liu, Y.W.; Chen, J.C.; He, H.L.; Kau, Y.C.; Liu, S.J. Anesthetics and human epidermal growth factor incorporated into anti-adhesive nanofibers provide sustained pain relief and promote healing of surgical wounds. Int. J. Nanomed. 2019, 29, 4007–4016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.S.; Sung, D.K.; Kong, W.H.; Kim, H.; Hahn, S.K. Synergistic effects of hyaluronate epidermal growth factor conjugate patch on chronic wound healing. Biomater. Sci. 2018, 6, 1020–1030. [Google Scholar] [CrossRef]
- Palencia, L.; Das, A.; Palecek, S.P.; Thibeault, S.L.; Leydon, C. Epidermal growth factor mediated healing in stem cell-derived vocal fold mucosa. J. Surg. Res. 2015, 197, 32–38. [Google Scholar] [CrossRef] [Green Version]
- Kaya, O.; Orhan, E.; Sapmaz-Metin, M.; Topçu-Tarladaçalışır, Y.; Gündüz, Ö.; Aydın, B. The effects of epidermal growth factor on early burn-wound progression in rats. Dermatol. Ther. 2020, 33, e13196. [Google Scholar] [CrossRef]
- Lou, Z. The effect of epidermal growth factor on the pseudo-healing of traumatic tympanic membrane perforations. Braz. J. Otorhinolaryngol. 2021, 87, 53–58. [Google Scholar] [CrossRef]
- Bui, T.Q.; Bui, Q.V.P.; Németh, D.; Hegyi, P.; Szakács, Z.; Rumbus, Z.; Tóth, B.; Emri, G.; Párniczky, A.; Sarlós, P.; et al. Epidermal Growth Factor is effective in the treatment of diabetic foot ulcers: Meta-analysis and systematic review. Int J. Environ. Res. Public Health 2019, 16, 2584. [Google Scholar] [CrossRef] [Green Version]
- Dogan, S.; Demirer, S.; Kepenekci, I.; Erkek, B.; Kiziltay, A.; Hasirci, N.; Müftüoglu, S.; Nazikoglu, A.; Renda, N.; Dincer, U.D.; et al. Epidermal growth factor-containing wound closure enhances wound healing in non-diabetic and diabetic rats. Int. Wound J. 2009, 6, 107–115. [Google Scholar] [CrossRef]
- Wei, S.; Wang, W.; Li, L.; Meng, H.Y.; Feng, C.Z.; Dong, Y.Y.; Fang, X.C.; Dong, Q.Q.; Jiang, W.; Xin, H.L.; et al. Recombinant human epidermal growth factor combined with vacuum sealing drainage for wound healing in Bama pigs. Mil. Med. Res. 2021, 8, 18. [Google Scholar]
- Lin, W.C.; Yeh, I.T.; Hsiao, H.Y. Development and evaluation of multistructured and hierarchical epidermal growth factor-poly (epsilon -Caprolactone) scaffolds. IEEE Trans. Nanobioscience 2019, 18, 18–27. [Google Scholar] [CrossRef]
- Kalay, Z.; Cevher, S.C. Oxidant and antioxidant events during epidermal growth factor therapy to cutaneous wound healing in rats. Int. Wound J. 2012, 9, 362–371. [Google Scholar] [CrossRef]
- Poniatowski, Ł.A.; Wojdasiewicz, P.; Gasik, R.; Szukiewicz, D. Transforming growth factor beta family: Insight into the role of growth factors in regulation of fracture healing biology and potential clinical applications. Mediat. Inflamm. 2015, 2015, 137823. [Google Scholar] [CrossRef] [Green Version]
- Khaghani, S.A.B.; Akbarova, G.; Soon, C.F.; Dilbazi, G. Effect of transforming growth factor-beta2 on biological regulation of multilayer primary chondrocyte culture. Cell Tissue Bank. 2018, 19, 763–775. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.F.; Fan, J.; Fedesco, M.; Guan, S.; Li, Y.; Bandyopadhyay, B.; Bright, A.M.; Yerushalmi, D.; Liang, M.; Chen, M.; et al. Transforming growth factor alpha (TGFalpha)-stimulated secretion of HSP90alpha: Using the receptor LRP-1/CD91 to promote human skin cell migration against a TGFbeta-rich environment during wound healing. Mol. Cell Biol. 2008, 28, 3344–3358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagares, D. P311 in scar wars: Myofibroblasts lost without transforming growth factor beta translation. Am. J. Respir Cell Mol. Biol. 2019, 60, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Shu, D.Y.; Lovicu, F.J. Myofibroblast transdifferentiation: The dark force in ocular wound healing and fibrosis. Prog. Retin. Eye Res. 2017, 60, 44–65. [Google Scholar] [CrossRef]
- Abe, Y.; Inagaki, K.; Fujiwara, A.; Kuriyama, K. Wound healing acceleration of a novel transforming growth factor-beta inducer, SEK-1005. Eur. J. Pharmacol. 2000, 17, 213–218. [Google Scholar] [CrossRef]
- Klass, B.R.; Grobbelaar, A.O.; Rolfe, K.J. Transforming growth factor beta1 signalling, wound healing and repair: A multifunctional cytokine with clinical implications for wound repair, a delicate balance. Postgrad. Med. J. 2009, 85, 9–14. [Google Scholar] [CrossRef]
- Farahani, R.M.; DiPietro, L.A. Microgravity and the implications for wound healing. Int. Wound J. 2008, 5, 552–561. [Google Scholar] [CrossRef]
- Koch, R.M.; Roche, N.S.; Parks, W.T.; Ashcroft, G.S.; Letterio, J.J.; Roberts, A.B. Incisional wound healing in transforming growth factor-beta1 null mice. Wound Repair Regen. 2000, 8, 179–191. [Google Scholar] [CrossRef]
- Peplow, P.V.; Baxter, G.D. Gene expression and release of growth factors during delayed wound healing: A review of studies in diabetic animals and possible combined laser phototherapy and growth factor treatment to enhance healing. Photomed. Laser Surg. 2012, 30, 617-36. [Google Scholar] [CrossRef] [PubMed]
- Van den Boom, R.; Wilmink, J.M.; O’Kane, S.; Wood, J.; Ferguson, M.W. Transforming growth factor-beta levels during second- intention healing are related to the different course of wound contraction in horses and ponies. Wound Repair Regen. 2002, 10, 188–194. [Google Scholar] [CrossRef]
- Rorison, P.; Thomlinson, A.; Hassan, Z.; Roberts, S.A.; Ferguson, M.W.; Shah, M. Longitudinal changes in plasma transforming growth factor beta-1 and post-burn scarring in children. Burns 2010, 36, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Olsen, B.R. The roles of vascular endothelial growth factor in bone repair and regeneration. Bone 2016, 91, 30–38. [Google Scholar] [CrossRef] [Green Version]
- Hoeben, A.; Landuyt, B.; Highley, M.S.; Wildiers, H.; Van Oosterom, A.T.; De Bruijn, E.A. Vascular endothelial growth factor and angiogenesis. Pharmacol. Rev. 2004, 56, 549–580. [Google Scholar] [CrossRef]
- Ong, H.T.; Dilley, R.J. Novel non-angiogenic role for mesenchymal stem cell-derived vascular endothelial growth factor on keratinocytes during wound healing. Cytokine Growth Factor Rev. 2018, 44, 69–79. [Google Scholar] [CrossRef]
- Bao, P.; Kodra, A.; Tomic-Canic, M.; Golinko, M.S.; Ehrlich, H.P.; Brem, H. The role of vascular endothelial growth factor in wound healing. J. Surg. Res. 2009, 15, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Hajialyani, M.; Tewari, D.; Sobarzo-Sánchez, E.; Nabavi, S.M.; Farzaei, M.H.; Abdollahi, M. Natural product-based nanomedicines for wound healing purposes: Therapeutic targets and drug delivery systems. Int. J. Nanomed. 2018, 3, 5023–5043. [Google Scholar] [CrossRef] [Green Version]
- Zubair, M.; Ahmad, J. Role of growth factors and cytokines in diabetic foot ulcer healing: A detailed review. Rev. Endocr. Metab. Disord. 2019, 20, 207–217. [Google Scholar] [CrossRef]
- Seiwerth, S.; Rucman, R.; Turkovic, B.; Sever, M.; Klicek, R.; Radic, B.; Drmic, D.; Stupnisek, M.; Misic, M.; Vuletic, L.B.; et al. BPC 157 and standard angiogenic growth factors. gastrointestinal tract healing, lessons from tendon, ligament, muscle and bone healing. Curr. Pharm. Des. 2018, 24, 1972–1989. [Google Scholar] [CrossRef]
- Lord, M.S.; Ellis, A.L.; Farrugia, B.L.; Whitelock, J.M.; Grenett, H.; Li, C.; O’Grady, R.L.; DeCarlo, A.A. Perlecan and vascular endothelial growth factor-encoding DNA-loaded chitosan scaffolds promote angiogenesis and wound healing. J. Control. Release 2017, 28, 48–61. [Google Scholar] [CrossRef]
- Shi, Y.; Tu, L.X.; Deng, Q.; Zhang, Y.P.; Hu, Y.H.; Liu, D.W. Effects and mechanism of rat epidermal stem cells treated with exogenous vascular endothelial growth factor on healing of deep partial-thickness burn wounds in rats. Zhonghua Shao Shang Za Zhi 2020, 20, 195–203. [Google Scholar]
- Li, X.; Liu, M.; Cheng, H.; Wang, Q.; Miao, C.; Ju, S.; Liu, F. Development of ionic liquid assisted-synthesized nano-silver combined with vascular endothelial growth factor as wound healing in the care of femoral fracture in the children after surgery. J. Photochem. Photobiol. B 2018, 183, 385–390. [Google Scholar] [CrossRef]
- Vijayan, A.A.S.; Kumar, G.S.V. PEG grafted chitosan scaffold for dual growth factor delivery for enhanced wound healing. Sci. Rep. 2019, 16, 19165. [Google Scholar] [CrossRef] [Green Version]
- Saaristo, A.; Tammela, T.; Farkkilā, A.; Kärkkäinen, M.; Suominen, E.; Yla-Herttuala, S.; Alitalo, K. Vascular endothelial growth factor-C accelerates diabetic wound healing. Am. J. Pathol. 2006, 169, 1080–1087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bogachkov, Y.Y.; Chen, L.; Le Master, E.; Fancher, I.S.; Zhao, Y.; Aguilar, V.; Oh, M.J.; Wary, K.K.; DiPietro, L.A.; Levitan, I. LDL induces cholesterol loading and inhibits endothelial proliferation and angiogenesis in matrigels: Correlation with impaired angiogenesis during wound healing. Am. J. Physiol. Cell Physiol. 2020, 1, C762–C776. [Google Scholar] [CrossRef] [PubMed]
- Stuard, W.L.; Titone, R.; Robertson, D.M. The IGF/Insulin-IGFBP axis in corneal development, wound healing, and disease. Front. Endocrinol. 2020, 11, 24. [Google Scholar] [CrossRef] [Green Version]
- Botusan, I.R.; Zheng, X.; Narayanan, S.; Grünler, J.; Sunkari, V.G.; Calissendorff, F.S.; Ansurudeen, I.; Illies, C.; Svensson, J.; Jansson, J.O.; et al. Deficiency of liver-derived insulin-like growth factor-I (IGF-I) does not interfere with the skin wound healing rate. PLoS ONE 2018, 13, e0193084. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.B.; Chen, E.H.; Lynch, S.E. A review of the effects of insulin-like growth factor and platelet derived growth factor on in vivo cartilage healing and repair. Osteoarthr. Cartil. 2006, 14, 403–412. [Google Scholar] [CrossRef] [Green Version]
- Gong, F.; Zhao, F.; Cheng, S.L.; Ding, D.; Zhang, B.W.; Li, X.L.; Huang, Y.L. Effect of insulin-like growth factor-1 on promoting healing of skin ulcers in diabetic rats. J. Biol. Regul. Homeost. Agents 2019, 33, 687–694. [Google Scholar] [PubMed]
- Shinchi, M.; Kushibiki, T.; Mayumi, Y.; Ito, K.; Asano, T.; Ishihara, M.; Horiguchi, A. Insulin-like growth factor 1 sustained-release collagen on urethral catheter prevents stricture after urethral injury in a rabbit model. Int. J. Urol. 2019, 26, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Cieszkowski, J.; Warzecha, Z.; Ceranowicz, P.; Ceranowicz, D.; Kusnierz-Cabala, B.; Pedziwiatr, M.; Dembinski, M.; Ambrozy, T.; Kaczmarzyk, T.; Pihut, M.; et al. Therapeutic effect of exogenous ghrelin in the healing of gingival ulcers is mediated by the release of endogenous growth hormone and insulin-like growth factor-1. J. Physiol. Pharmacol. 2017, 68, 609–617. [Google Scholar]
- Todorović, V.; Pesko, P.; Micev, M.; Bjelović, M.; Budec, M.; Mićić, M.; Brasanac, D.; Ilić-Stojanović, O. Insulin-like growth factor-I in wound healing of rat skin. Regul. Pept. 2008, 9, 7–13. [Google Scholar] [CrossRef]
- Reckenbeil, J.; Kraus, D.; Stark, H.; Rath-Deschner, B.; Jäger, A.; Wenghoefer, M.; Winter, J.; Götz, W. Insulin-like growth factor 1 (IGF1) affects proliferation and differentiation and wound healing processes in an inflammatory environment with p38 controlling early osteoblast differentiation in periodontal ligament cells. Arch. Oral Biol. 2017, 73, 142–150. [Google Scholar] [CrossRef]
- Balaji, S.; LeSaint, M.; Bhattacharya, S.S.; Moles, C.; Dhamija, Y.; Kidd, M.; Le, L.D.; King, A.; Shaaban, A.; Crombleholme, T.M.; et al. Adenoviral-mediated gene transfer of insulin-like growth factor 1 enhances wound healing and induces angiogenesis. J. Surg. Res. 2014, 190, 367–377. [Google Scholar] [CrossRef] [Green Version]
- Achar, R.A.; Silva, T.C.; Achar, E.; Martines, R.B.; Machado, J.L. Use of insulin-like growth factor in the healing of open wounds in diabetic and non-diabetic rats. Acta Cir. Bras. 2014, 29, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Pastar, I.; Stojadinovic, O.; Yin, N.C.; Ramirez, H.; Nusbaum, A.G.; Sawaya, A.; Patel, S.B.; Khalid, L.; Isseroff, R.R.; Tomic-Canic, M. Epithelialization in wound healing: A comprehensive review. Adv. Wound Care 2014, 3, 445–464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pan, A.; Zhong, M.; Wu, H.; Peng, Y.; Xia, H.; Tang, Q.; Huang, Q.; Wei, L.; Xiao, L.; Peng, C. Topical application of keratinocyte growth factor conjugated gold nanoparticles accelerate wound healing. Nanomedicine 2018, 14, 1619–1628. [Google Scholar] [CrossRef] [PubMed]
- Denzinger, M.; Link, A.; Kurz, J.; Krauss, S.; Thoma, R.; Schlensak, C.; Wendel, H.P.; Krajewski, S. Keratinocyte growth factor modified messenger RNA accelerating cell proliferation and migration of keratinocytes. Nucleic Acid Ther. 2018, 28, 335–347. [Google Scholar] [CrossRef] [PubMed]
- Devalliere, J.; Dooley, K.; Hu, Y.; Kelangi, S.S.; Uygun, B.E.; Yarmush, M.L. Co-delivery of a growth factor and a tissue-protective molecule using elastin biopolymers accelerates wound healing in diabetic mice. Biomaterials 2017, 141, 149–160. [Google Scholar] [CrossRef]
- Bienert, M.; Hoss, M.; Bartneck, M.; Weinandy, S.; Böbel, M.; Jockenhövel, S.; Knüchel, R.; Pottbacker, K.; Wöltje, M.; Jahnen-Dechent, W.; et al. Growth factor-functionalized silk membranes support wound healing in vitro. Biomed. Mater. 2017, 16, 045023. [Google Scholar] [CrossRef]
- Chomiski, V.; Gragnani, A.; Bonucci, J.; Correa, S.A.; Noronha, S.M.; Ferreira, L.M. Keratinocyte growth factor and the expression of wound-healing-related genes in primary human keratinocytes from burn patients. Acta Cir. Bras. 2016, 31, 505–512. [Google Scholar] [CrossRef] [Green Version]
- Cai, J.; Zhou, Q.; Wang, Z.; Guo, R.; Yang, R.; Yang, X.; Li, W.; Ahmad, N.; Chen, Q.; Hui, Q.; et al. Comparative analysis of KGF-2 and bFGF in prevention of excessive wound healing and scar formation in a corneal alkali burn model. Cornea 2019, 38, 1430–1437. [Google Scholar] [CrossRef]
- Sen-Britain, S.; Britain, D.M.; Hicks, W.L., Jr.; Gardella, J.A., Jr. ToF-SIMS and TIRF microscopy investigation on the effects of HEMA copolymer surface chemistry on spatial localization, surface intensity, and release of fluorescently labeled keratinocyte growth factor. Biointerphases 2019, 23, 051003. [Google Scholar] [CrossRef]
- Dougan, M.; Dranoff, G.; Dougan, S.K. GM-CSF, IL-3, and IL-5 family of cytokines: Regulators of inflammation. Immunity 2019, 16, 796–811. [Google Scholar] [CrossRef]
- Brem, H.; Howell, R.; Criscitelli, T.; Senderowicz, A.; Siegart, N.; Gorenstein, S.; Gillette, B. Practical application of granulocyte-macrophage colony-stimulating factor (GM-CSF) in patients with wounds. Surg. Technol. Int. 2018, 1, 61–66. [Google Scholar]
- Fang, Y.; Gong, S.J.; Xu, Y.H.; Hambly, B.D.; Bao, S. Impairedcutaneous wound healing in granulocyte/macrophage colonystimulating factor knockout mice. Br. J. Dermatol. 2007, 157, 458-65. [Google Scholar] [CrossRef]
- Rho, C.R.; Park, M.Y.; Kang, S. Effects of granulocyte-macrophage colony-stimulating (GM-CSF) factor on corneal epithelial cells in corneal wound healing model. PLoS ONE 2015, 16, e0138020. [Google Scholar] [CrossRef] [PubMed]
- Yan, D.; Liu, S.; Zhao, X.; Bian, H.; Yao, X.; Xing, J.; Sun, W.; Chen, X. Recombinant human granulocyte macrophage colony stimulating factor in deep second-degree burn wound healing. Medicine 2017, 96, e6881. [Google Scholar] [CrossRef]
- Li, J.; Liu, W.; Zhang, G.; Wang, D.; Lou, H.; Duang, J. Effectiveness of recombinanthuman granulocyte macrophage colony-stimulating factor for treating deep second-degree burns: A systematic review and meta-analysis. BMJ Mil. Health 2020, 166, 352–357. [Google Scholar] [CrossRef] [PubMed]
- Chi, Y.F.; Chai, J.K.; Luo, H.M.; Zhang, Q.X.; Feng, R. Safety of recombinant human granulocyte-macrophage colony-stimulating factor in healing pediatric severe burns. Genet. Mol. Res. 2015, 31, 2735–2741. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.A.; Zhang, X.H.; Xue, Y.; Zhou, X.; Chen, X.X.; Gao, X.X.; Yu, J.A. Effects of recombinanthuman granulocyte macrophage colony stimulating factor gel on treatment of full-thickness frostbite wounds on foot and hand. Zhonghua Shao Shang Za Zhi 2020, 20, 117–121. [Google Scholar]
- Lim, J.Y.; Choi, B.H.; Lee, S.; Jang, Y.H.; Choi, J.S.; Kim, Y.M. Regulation of wound healing by granulocyte-macrophage colony-stimulating factor after vocal fold injury. PLoS ONE 2013, 8, e54256. [Google Scholar]
- Fang, Y.; Shen, J.; Yao, M.; Beagley, K.W.; Hambly, B.D.; Bao, S. Granulocyte-macrophage colony stimulating factor enhances wound healing in diabetes via upregulation of proinflammatory cytokines. Br. J. Dermatol. 2010, 162, 478–486. [Google Scholar] [CrossRef]
- Haisa, M.; Okochi, H.; Grotendorst, G.R. Elevated levels of PDGF alpha receptors in keloid fibroblasts contribute to an enhanced response to PDGF. J. Investig. Dermatol. 1994, 103, 560–563. [Google Scholar] [CrossRef] [Green Version]
- Barrientos, S.; Brem, H.; Stojadinovic, O.; Tomic-Canic, M. Clinical application of growth factors and cytokines in wound healing. Wound Repair Regen. 2014, 22, 569–578. [Google Scholar] [CrossRef] [Green Version]
- Demaria, M.; Ohtani, N.; Youssef, S.A.; Rodier, F.; Toussaint, W.; Mitchell, J.R.; Laberge, R.M.; Vijg, J.; Van Steeg, H.; Dollé, M.E.; et al. An essential role for senescent cells in optimal wound healing through secretion of PDGF-AA. Dev. Cell 2014, 22, 722–733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zehra, M.; Zubairi, W.; Hasan, A.; Butt, H.; Ramzan, A.; Azam, M.; Mehmood, A.; Falahati, M.; Chaudhry, A.A.; Rehman, I.U.; et al. Oxygen generating polymeric nano fibers that stimulate angiogenesis and show efficient wound healing in a diabetic wound model. Int. J. Nanomed. 2020, 18, 3511–3522. [Google Scholar] [CrossRef]
- Tanaka, T.; Minami, A.; Uchida, J.; Nakatani, T. Potential of hyperbaric oxygen in urological diseases. Int. J. Urol. 2019, 26, 860–867. [Google Scholar] [CrossRef] [Green Version]
- Chandan, K.S. Wound healing essentials: Let there be oxygen. Wound Repair Regen. 2009, 17, 1–18. [Google Scholar]
- Lin, W.; Qi, X.; Guo, W.; Liang, D.; Chen, H.; Lin, B.; Deng, X. A barrier against reactive oxygen species: Chitosan/acellular dermal matrix scaffold enhances stem cell retention and improves cutaneous wound healing. Stem Cell Res. Ther. 2020, 7, 383. [Google Scholar] [CrossRef]
- Fosen, K.M.; Thom, S.R. Hyperbaric oxygen, vasculogenic stem cells, and wound healing. Antioxid. Redox Signal. 2014, 21, 1634–1647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nouvong, A.; Ambrus, A.M.; Zhang, E.R.; Hultman, L.; Coller, H.A. Reactive oxygen species and bacterial biofilms in diabetic wound healing. Physiol. Genom. 2016, 48, 889–896. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.; Shi, F.; Zhou, Z.; Sun, F.; Sun, M.H.; Sun, Q.; Chen, L.; Li, D.; Jiang, C.Y.; Zhao, R.Z.; et al. M1 macrophage mediated increased reactive oxygen species (ROS) influence wound healing via the MAPK signaling in vitro and in vivo. Toxicol. Appl. Pharmacol. 2019, 366, 83–95. [Google Scholar] [CrossRef]
- Xiao, K.; Liu, C.; Tu, Z.; Xu, Q.; Chen, S.; Zhang, Y.; Wang, X.; Zhang, J.; Hu, C.A.; Liu, Y. Activation of the NF-kappaB and MAPK Signaling pathways contributes to the inflammatory responses, but not cell injury, in IPEC-1 cells challenged with hydrogen peroxide. Oxid. Med. Cell Longev. 2020, 2020, 5803639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeBert, D.; Squirrell, J.M.; Freisinger, C.; Rindy, J.; Golenberg, N.; Frecentese, G.; Gibson, A.; Eliceiri, K.W.; Huttenlocher, A. Damage-induced reactive oxygen species regulate vimentin and dynamic collagen-based projections to mediate wound repair. Elife 2018, 7, e30703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- André-Lévigne, D.; Modarressi, A.; Pepper, M.S.; Pittet-Cuénod, B. Reactive oxygen species and NOX enzymes are emerging as key players in cutaneous wound repair. Int. J. Mol. Sci. 2017, 18, 2149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Butin-Israeli, V.; Bui, T.M.; Wiesolek, H.L.; Mascarenhas, L.; Lee, J.J.; Mehl, L.C.; Knutson, K.R.; Adam, S.A.; Goldman, R.D.; Beyder, A.; et al. Neutrophil-induced genomic instability impedes resolution of inflammation and wound healing. J. Clin. Investig. 2019, 129, 712–726. [Google Scholar] [CrossRef]
- Sardella, E.; Mola, M.G.; Gristina, R.; Piccione, M.; Veronico, V.; Bellis, M.; Cibelli, A.; Buttiglione, M.; Armenise, V.; Favia, P.; et al. A synergistic effect of reactive oxygen and reactive nitrogen species in plasma activated liquid media triggers astrocyte wound healing. Int. J. Mol. Sci. 2020, 21, 3343. [Google Scholar] [CrossRef] [PubMed]
- Thi, P.L.; Lee, Y.; Tran, D.L.; Thi, T.T.H.; Kang, J.I.; Park, K.M.; Park, K.D. In situ forming and reactive oxygen species-scavenging gelatin hydrogels for enhancing wound healing efficacy. Acta Biomater. 2020, 103, 103,142–152. [Google Scholar] [CrossRef]
- Loo, A.E.; Ho, R.; Halliwell, B. Mechanism of hydrogen peroxide-induced keratinocyte migration in a scratch-wound model. Free Radic. Biol. Med. 2011, 15, 884–892. [Google Scholar] [CrossRef]
- Fan, Z.; Wang, X.; Zhang, M.; Zhao, C.; Mei, C.; Li, P. MAPK pathway inhibitors attenuated hydrogen peroxide induced damage in neural cells. Biomed. Res. Int. 2019, 2019, 5962014. [Google Scholar] [CrossRef]
- Hou, B.; Cai, W.; Chen, T.; Zhang, Z.; Gong, H.; Yang, W.; Qiu, L. Vaccarin hastens wound healing by promoting angiogenesis via activation of MAPK/ERK and PI3K/AKT signaling pathways in vivo. Acta Cir. Bras. 2020, 7, 201901202. [Google Scholar] [CrossRef] [Green Version]
- Wahedi, H.M.; Chae, J.K.; Subedi, L.; Kang, M.C.; Cho, H.; Kim, S.; Kim, S.Y. NED416, a novel synthetic Sirt1 activator, promotes cutaneous wound healing via the MAPK/Rho pathway. Int. J. Mol. Med. 2020, 46, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.C.; Song, J.; Lee, A.; Cho, D.; Kim, T.S. Visfatin promotes wound healing through the activation of ERK1/2 and JNK1/2 Pathway. Int. J. Mol. Sci. 2018, 19, 3642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escuin-Ordinas, H.; Li, S.; Xie, M.W.; Sun, L.; Hugo, W.; Huang, R.R.; Jiao, J.; de-Faria, F.M.; Realegeno, S.; Krystofinski, P.; et al. Cutaneous wound healing through paradoxical MAPK activation by BRAF inhibitors. Nat. Commun. 2016, 7, 2348. [Google Scholar] [CrossRef]
- Chen, J.; Chen, Y.; Chen, Y.; Yang, Z.; You, B.; Ruan, Y.C.; Peng, Y. Epidermal CFTR suppresses MAPK/NF-kappaB to promote cutaneous wound healing. Cell Physiol. Biochem. 2016, 39, 2262–2274. [Google Scholar] [CrossRef]
- Yuan, X.; Han, L.; Fu, P.; Zeng, H.; Lv, C.; Chang, W.; Runyon, R.S.; Ishii, M.; Han, L.; Liu, K.; et al. Cinnamaldehyde accelerates wound healing by promoting angiogenesis via up-regulation of PI3K and MAPK signalling pathways. Lab. Investig. 2018, 98, 783–798. [Google Scholar] [CrossRef]
- Su, L.; Fu, L.; Li, X.; Zhang, Y.; Li, Z.; Wu, X.; Li, Y.; Bai, X.; Hu, D. Loss of CAR promotes migration and proliferation of HaCaT cells, and accelerates wound healing in rats via Src-p38 MAPK pathway. Sci. Rep. 2016, 6, 19735. [Google Scholar] [CrossRef] [Green Version]
- Liu, C.; Huang, J.; Li, H.; Yang, Z.; Zeng, Y.; Liu, J.; Hao, Y.; Li, R. Ghrelin accelerates wound healing through GHS-R1a-mediated MAPK-NF-kappaB/GR signaling pathways in combined radiation and burn injury in rats. Sci. Rep. 2016, 7, 27499. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Xu, P.; Wang, X.; Zhang, M.; Yan, Y.; Chen, Y.; Zhang, L.; Zhang, L. Activin B regulates adipose-derived mesenchymal stem cells to promote skin wound healing via activation of the MAPK signalling pathway. Int. J. Biochem. Cell Biol. 2017, 87, 69–76. [Google Scholar] [CrossRef]
- Cui, D.; Xiao, J.; Zhou, Y.; Zhou, X.; Liu, Y.; Peng, Y.; Yu, Y.; Li, H.; Zhou, X.; Yuan, Q.; et al. Epiregulin enhances odontoblastic differentiation of dental pulp stem cells via activating MAPK signalling pathway. Cell Prolif. 2019, 52, e12680. [Google Scholar] [CrossRef] [Green Version]
- Pereira, B.F.; Xue, M.; Maia, G.L.A.; Leite, R.A.; Helena, P.C.; Jackson, C.J. Lupeol, a pentacyclic triterpene, promotes migration, wound closure, and contractile effect in vitro: Possible involvement of PI3K/Akt and p38/ERK/MAPK pathways. Molecules 2018, 30, 2819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultze, S.M.; Hemmings, B.A.; Niessen, M.; Tschopp, O. PI3K/AKT, MAPK and AMPK signalling: Protein kinases in glucose homeostasis. Expert. Rev. Mol. Med. 2012, 14, E1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moldogazieva, N.T.; Lutsenko, S.V.; Terentiev, A.A. Reactive oxygen and nitrogen species-induced protein modifications: Implication in carcinogenesis and anticancer therapy. Cancer Res. 2018, 78, 6040–6047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez-de-Diego, C.; Valer, J.A.; Pimenta-Lopes, C.; Rosa, J.L.; Ventura, F. Interplay between BMPs and reactive oxygen species in cell signaling and pathology. Biomolecules 2019, 9, 534. [Google Scholar] [CrossRef] [Green Version]
- Ornitz, D.M.; Itoh, N. The fibroblast growth factor signaling pathway. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 215–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scarpello, J.H.; Howlett, H.C. Metformin therapy and clinical uses. Diabetes Vasc. Dis. Res. 2008, 5, 157–167. [Google Scholar] [CrossRef] [PubMed]
- Inzucchi, S.E.; Bergenstal, R.M.; Buse, J.B.; Diamant, M.; Ferrannini, E.; Nauck, M.; Peters, A.L.; Tsapas, A.; Wender, R.; Matthews, D.R. American Diabetes Association (ADA); european association for the study of diabetes (EASD). Management of hyperglycemia in type 2 diabetes: A patient-centered approach: Position statement of the American diabetes association (ADA) and the European association for the study of diabetes (EASD). Diabetes Care 2012, 35, 1364–1379. [Google Scholar]
- Jeschke, M.G.; Abdullahi, A.; Burnett, M.; Rehou, S.; Stanojcic, M. Glucose control in severely burned patients using metformin: An interim safety and efficacy analysis of a phase II randomized controlled trial. Ann. Surg. 2016, 264, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Rena, G.; Hardie, D.G.; Pearson, E.R. The mechanisms of action of metformin. Diabetologia 2017, 60, 1577–1585. [Google Scholar] [CrossRef] [Green Version]
- Gore, D.C.; Wolf, S.E.; Sanford, A.; Herndon, D.N.; Wolfe, R.R. Influence of metformin on glucose intolerance and muscle catabolism following severe burn injury. Ann. Surg. 2005, 241, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Bridges, H.R.; Jones, A.J.; Pollak, M.N.; Hirst, J. Effects of metformin and other biguanides on oxidative phosphorylation in mitochondria. Biochem. J. 2014, 462, 475–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saisho, Y. Metformin and inflammation: Its potential beyond glucose-lowering effect. Endocr. Metab. Immune Disord. -Drug Targets 2015, 15, 196–205. [Google Scholar] [CrossRef]
- Xu, T.; Wu, X.; Lu, X.; Liang, Y.; Mao, Y.; Loor, J.J.; Yang, Z. Metformin activated AMPK signaling contributes to the alleviation of LPS-induced inflammatory responses in bovine mammary epithelial cells. BMC Vet. Res. 2021, 17, 97. [Google Scholar] [CrossRef] [PubMed]
- Arbab, A.A.I.; Lu, X.; Abdalla, I.M.; Idris, A.A.; Chen, Z.; Li, M.; Mao, Y.; Xu, T.; Yang, Z. Metformin inhibits lipoteichoic acid-induced oxidative stress and inflammation through AMPK/NRF2/NF-κB signaling pathway in bovine mammary epithelial cells. Front. Vet. Sci. 2021, 8, 661380. [Google Scholar] [CrossRef]
- Song, Y.M.; Lee, W.K.; Lee, Y.H.; Kang, E.S.; Cha, B.S.; Lee, B.W. Metformin restores parkin-mediated mitophagy, suppressed by cytosolic p53. Int. J. Mol. Sci. 2016, 17, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doblado, L.; Lueck, C.; Rey, C.; Samhan-Arias, A.K.; Prieto, I.; Stacchiotti, A.; Monsalve, M. Mitophagy in human diseases. Int. J. Mol. Sci. 2021, 22, 3903. [Google Scholar] [CrossRef]
- Diaz, E.C.; Herndon, D.N.; Porter, C.; Sidossis, L.S.; Suman, O.E.; Børsheim, E. Effects of pharmacological interventions on muscle protein synthesis and breakdown in recovery from burns. Burns 2015, 41, 649–657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, H.M.; Kim, S.J.; Chun, S.W.; Park, K.Y.; Lim, D.M.; Lee, J.M.; Hong, J.H.; Park, K.S. A comparison study on efficacy, insulin sensitivity and safety of glimepiride/metformin fixed dose combination versus glimepiride single therapy on type 2 diabetes mellitus patients with basal insulin therapy. Diabetes Res. Clin. Pract. 2019, 155, 107796. [Google Scholar] [CrossRef]
- Cree-Green, M.; Bergman, B.C.; Cengiz, E.; Fox, L.A.; Hannon, T.S.; Miller, K.; Nathan, B.; Pyle, L.; Kahn, D.; Tansey, M. Metformin improves peripheral insulin sensitivity in youth with type 1 diabetes. J. Clin. Endocrinol. Metab. 2019, 104, 3265–3278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sikorskaya, K.; Zarzecka, I.; Ejikeme, U.; Russell, J. The use of metformin as an add-on therapy to insulin in the treatment of poorly controlled type 1 diabetes mellitus in adolescents. Metab. Open 2021, 9, 100080. [Google Scholar] [CrossRef]
- Jeschke, M.G. Clinical review: Glucose control in severely burned patients—current best practice. Crit. Care 2013, 17, 232. [Google Scholar] [CrossRef] [Green Version]
- Shawky, L.M.; El Bana, E.A.; Morsi, A.A. Stem cells and metformin synergistically promote healing in experimentally induced cutaneous wound injury in diabetic rats. Folia Histochem. Cytobiol. 2019, 57, 127–138. [Google Scholar] [CrossRef]
- Taleb, S.; Moghaddas, P.; Rahimi Balaei, M.; Taleb, S.; Rahimpour, S.; Abbasi, A.; Ejtemaei-Mehr, S.; Dehpour, A.R. Metformin improves skin flap survival through nitric oxide system. J. Surg. Res. 2014, 192, 686–691. [Google Scholar] [CrossRef]
- Wu, H.; Ding, J.; Li, S.; Lin, J.; Jiang, R.; Lin, C.; Dai, L.; Xie, C.; Lin, D.; Xu, H.; et al. Metformin Promotes the Survival of Random-Pattern Skin Flaps by Inducing Autophagy via the AMPK-mTOR-TFEB signaling pathway. Int. J. Biol. Sci. 2019, 15, 325–340. [Google Scholar] [CrossRef] [Green Version]
- Young, N.; Kamireddy, A.; Van Nostrand, J.; Eichner, L.; Shokhirev, M.; Dayn, Y.; Shaw, R.J. AMPK governs lineage specification through Tfeb-dependent regulation of lysosomes. Genes Dev. 2016, 30, 535–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ochoa-Gonzalez, F.; Cervantes-Villagrana, A.R.; Fernandez-Ruiz, J.C.; Nava-Ramirez, H.S.; Hernandez-Correa, A.C.; Enciso-Moreno, J.A.; Castañeda-Delgado, J.E. Metformin induces cell cycle arrest, reduced proliferation, wound healing impairment in vivo and is associated to clinical outcomes in diabetic foot ulcer patients. PLoS ONE 2016, 11, e0150900. [Google Scholar]
- Nair, V.; Sreevalsan, S.; Basha, R.; Abdelrahim, M.; Abudayyeh, A.; Rodrigues, H.A.; Safe, S. Mechanism of metformin-dependent inhibition of mammalian target of rapamycin (mTOR) and Ras activity in pancreatic cancer: Role of specificity protein (Sp) transcription factors. J. Biol. Chem. 2014, 3, 27692–27701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Revuelta, B.I.; Hettich, M.M.; Ciociaro, A.; Rotermund, C.; Kahle, P.J.; Krauss, S.; Di Monte, D.A. Metformin lowers Ser- phosphorylated α-synuclein levels via mTOR- dependent protein phosphatase 2A activation. Cell Death Dis. 2014, 8, 1209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wetzel, M.; Herndon, D.; Finnerty, C. Metformin down-regulates epithelial-mesenchymal transition (EMT) in fibroblasts from burned patients. Cancer Metab. 2014, 2, P84. [Google Scholar] [CrossRef] [Green Version]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miricescu, D.; Badoiu, S.C.; Stanescu-Spinu, I.-I.; Totan, A.R.; Stefani, C.; Greabu, M. Growth Factors, Reactive Oxygen Species, and Metformin—Promoters of the Wound Healing Process in Burns? Int. J. Mol. Sci. 2021, 22, 9512. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179512
Miricescu D, Badoiu SC, Stanescu-Spinu I-I, Totan AR, Stefani C, Greabu M. Growth Factors, Reactive Oxygen Species, and Metformin—Promoters of the Wound Healing Process in Burns? International Journal of Molecular Sciences. 2021; 22(17):9512. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179512
Chicago/Turabian StyleMiricescu, Daniela, Silviu Constantin Badoiu, Iulia-Ioana Stanescu-Spinu, Alexandra Ripszky Totan, Constantin Stefani, and Maria Greabu. 2021. "Growth Factors, Reactive Oxygen Species, and Metformin—Promoters of the Wound Healing Process in Burns?" International Journal of Molecular Sciences 22, no. 17: 9512. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22179512