
The Osteogenic Differentiation of Human Dental Pulp Stem Cells through G0/G1 Arrest and the p-ERK/Runx-2 Pathway by Sonic Vibration


Abstract
:1. Introduction
2. Results
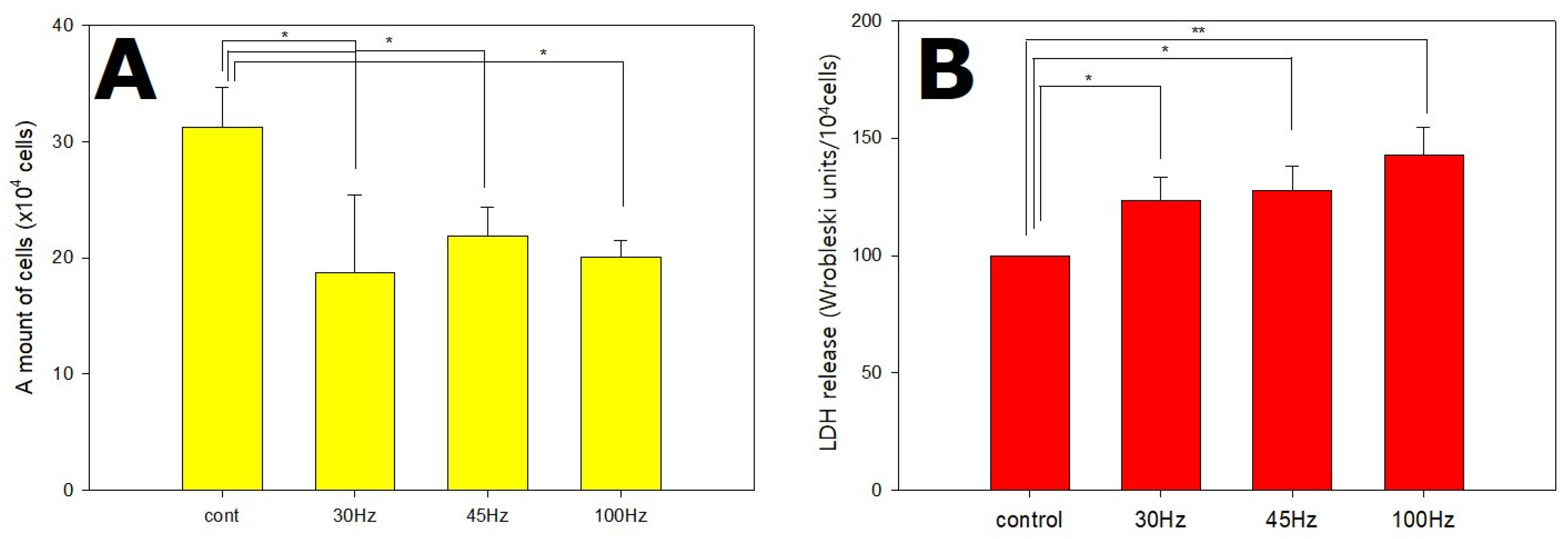
2.1. Effects of Mechanical Vibration Frequency on HDPSC Proliferation and Stress Level
2.2. Effect of Mechanical Vibration Frequency on HDPSC Morphology
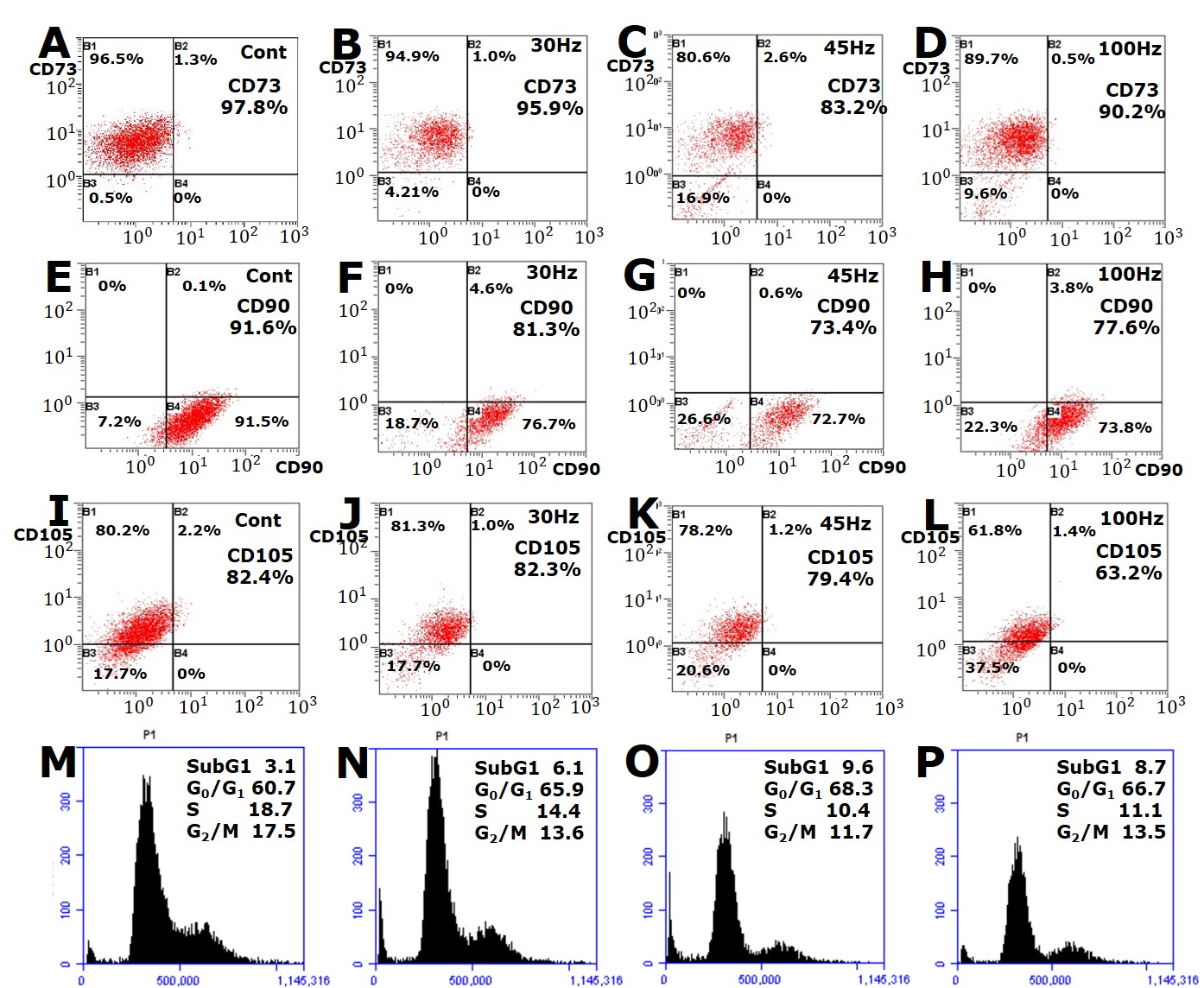
2.3. Effect of Mechanical Vibration Frequency on HDPC Surface Antigen Expression
2.4. Effect of Mechanical Vibration Frequency on the HDPC Cell Cycle
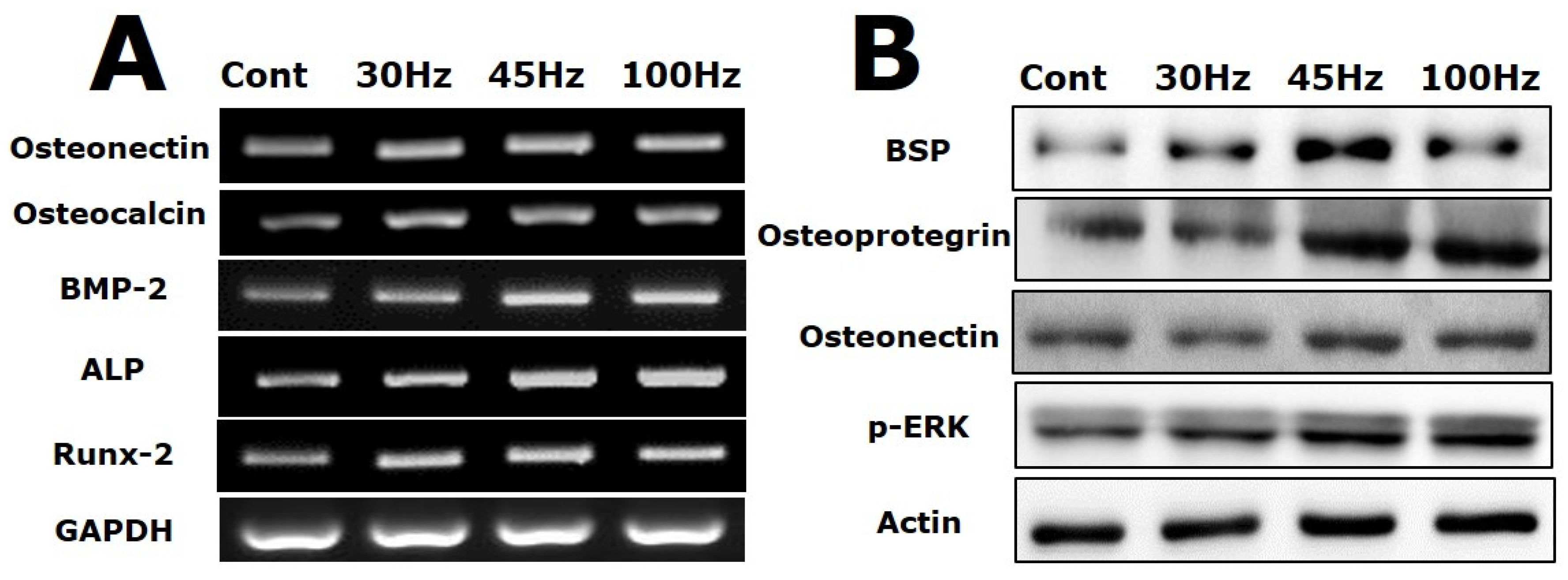
2.5. Effect of Mechanical Vibration Frequency on the Expression of Osteogenic Markers
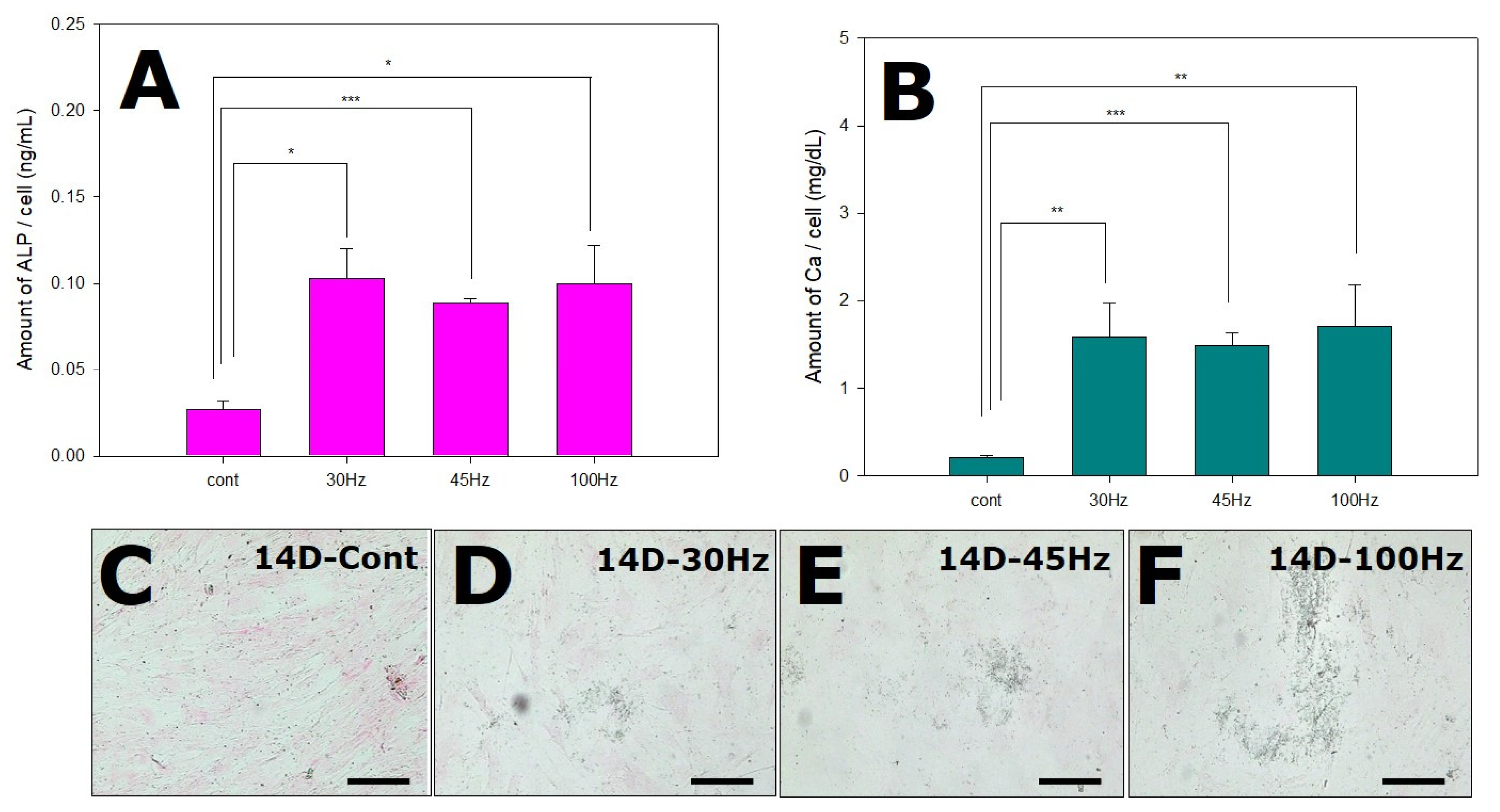
2.6. Effects of Mechanical Vibration Frequency on ALP and Calcium Deposition
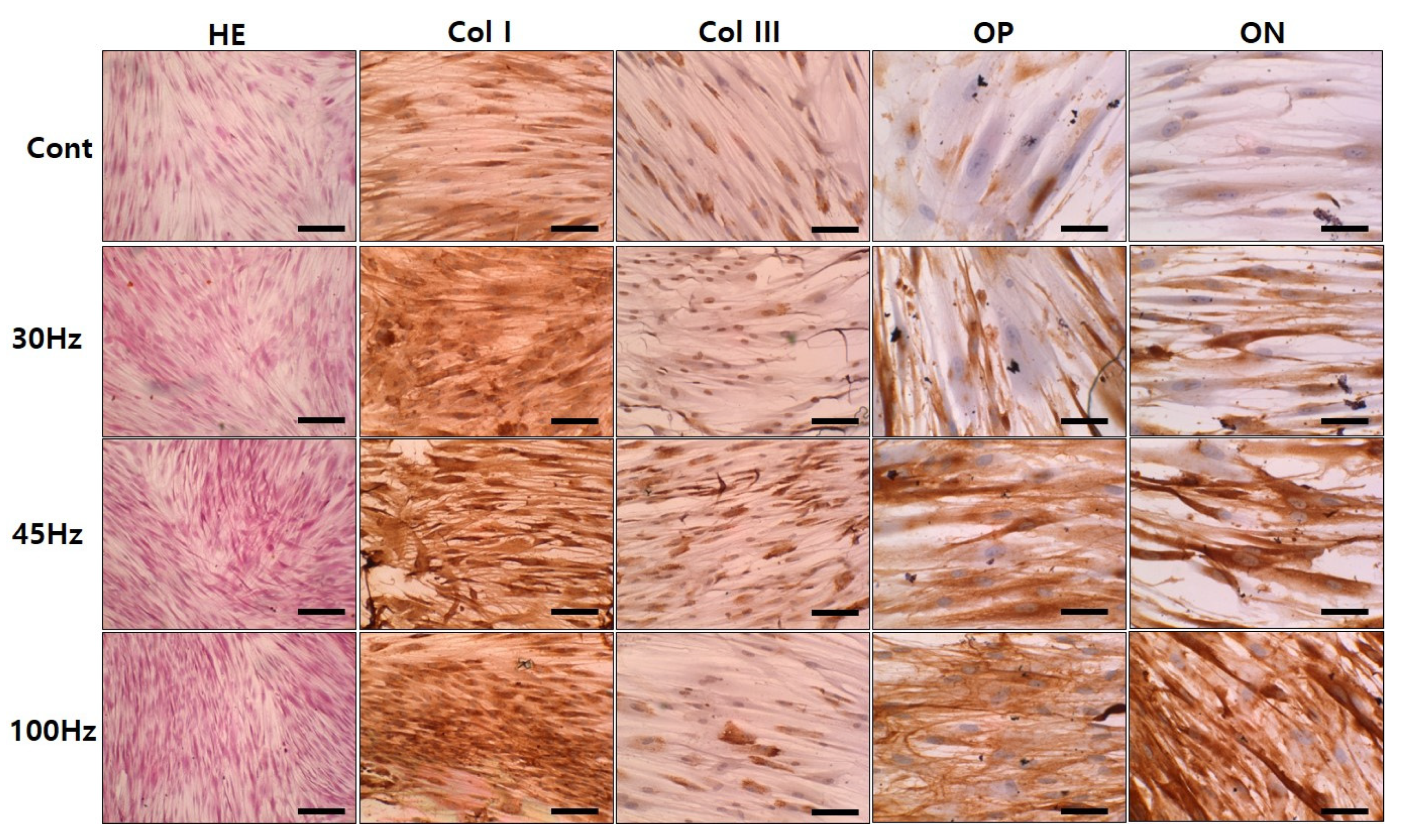
2.7. Immunohistochemical Examination
3. Discussion
4. Materials and Methods
4.1. Culture of HDPSCs
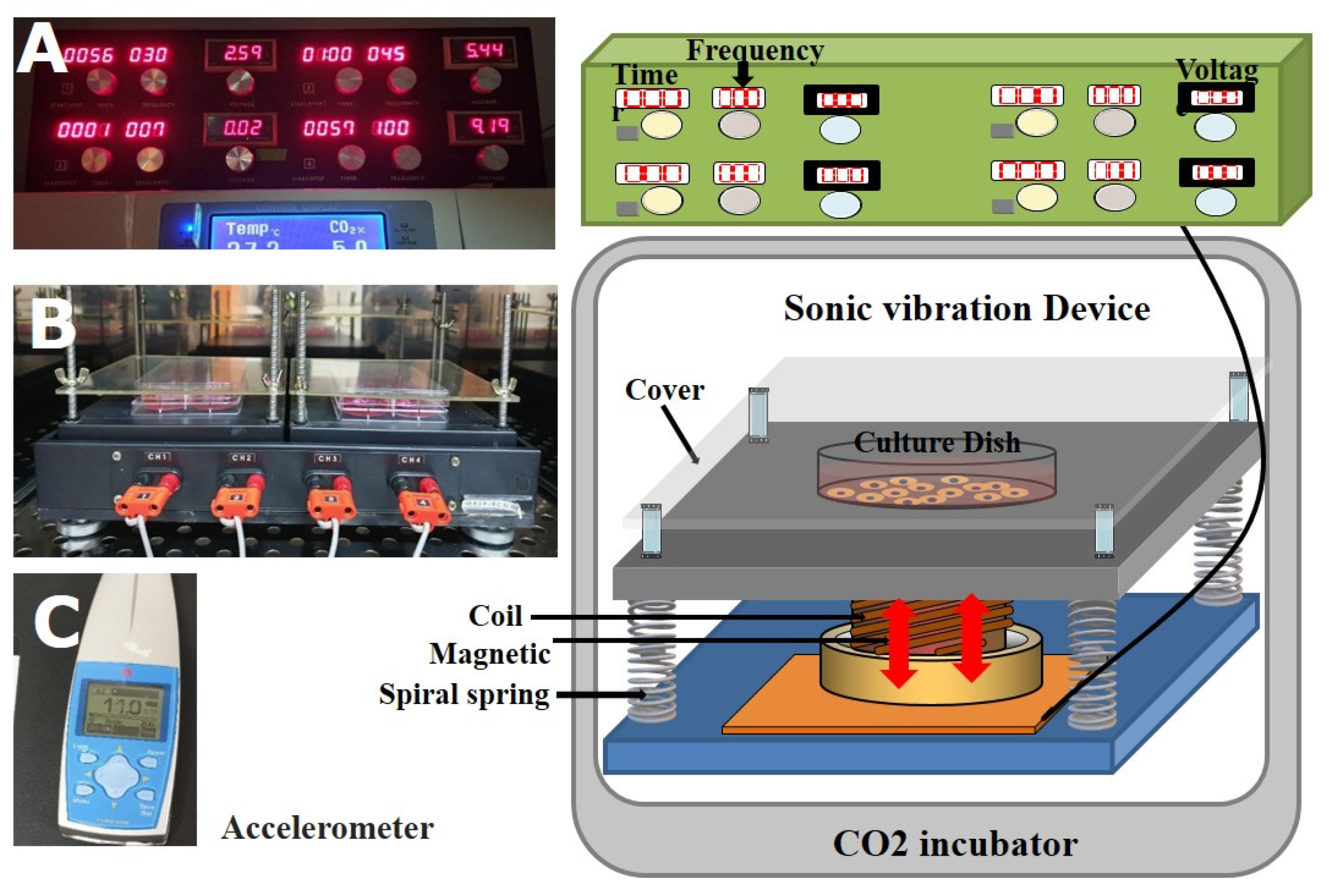
4.2. Stimulation by Mechanical Vibration
4.3. Proliferation and Lactate Dehydrogenase (LDH) Assays
4.4. Cell Surface Antigen Expression Analysis via Fluorescence-Activated Cell Sorting (FACS)
4.5. Cell Cycle Analysis
4.6. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.7. Western Blot Analysis
4.8. ALP and Calcium Deposition Assays
4.9. Immunohistochemical Analysis
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Graziano, A.; d’Aquino, R.; Laino, G.; Papaccio, G. Dental pulp stem cells: A promising tool for bone regeneration. Stem. Cell Rev. 2008, 4, 21–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gronthos, S.; Mankani, M.; Brahim, J.; Robey, P.G.; Shi, S. Postnatal human dental pulp stem cells (DPSCs) in vitro and in vivo. Proc. Natl. Acad. Sci. USA 2000, 97, 13625–13630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koyama, N.; Okubo, Y.; Nakao, K.; Bessho, K. Evaluation of pluripotency in human dental pulp cells. J. Oral Maxillofac. Surg. 2009, 67, 501–506. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; He, H.; Tang, C.; Zhang, G.; Li, Y.; Wang, R.; Shi, J.; Jin, Y. Differentiation potential of STRO-1+ dental pulp stem cells changes during cell passaging. BMC Cell Biol. 2010, 11, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, W.R.; Yen, S.S.; Rubin, J. Vibration therapy: Clinical applications in bone. Curr. Opin. Endocrinol. Diabetes Obes. 2014, 21, 447–453. [Google Scholar]
- Mehta, S.; McClarren, B.; Aijaz, A.; Chalaby, R.; Cook-Chennault, K.; Olabisi, R.M. The effect of low-magnitude, high-frequency vibration on poly(ethylene glycol)-microencapsulated mesenchymal stem cells. J. Tissue Eng. 2018, 9, 2041731418800101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yong, K.W.; Choi, J.R.; Choi, J.Y.; Cowie, A.C. Recent Advances in Mechanically Loaded Human Mesenchymal Stem Cells for Bone Tissue Engineering. Int. J. Mol. Sci. 2020, 21, 5816. [Google Scholar] [CrossRef] [PubMed]
- Rubin, C.; Recker, R.; Cullen, D.; Ryaby, J.; McCabe, J.; McLeod, K. Prevention of postmenopausal bone loss by a low-magnitude, high-frequency mechanical stimuli: A clinical trial assessing compliance, efficacy, and safety. J. Bone Miner. Res. 2004, 19, 343–351. [Google Scholar] [CrossRef] [PubMed]
- El-Mowafi, H.; Mohsen, M. The effect of low-intensity pulsed ultrasound on callus maturation in tibial distraction osteogenesis. Int. Orthop. 2005, 29, 121–124. [Google Scholar] [CrossRef] [Green Version]
- Alikhani, M.; Lopez, J.A.; Alabdullah, H.; Vongthongleur, T.; Sangsuwon, C.; Alikhani, M.; Alansari, S.; Oliveira, S.M.; Nervina, J.M.; Teixeira, C.C. High-Frequency Acceleration: Therapeutic Tool to Preserve Bone following Tooth Extractions. J. Dent. Res. 2016, 95, 311–318. [Google Scholar] [CrossRef]
- Alikhani, M.; Alikhani, M.; Alansari, S.; Almansour, A.; Hamidaddin, M.A.; Khoo, E.; Lopez, J.A.; Nervina, J.M.; Nho, J.Y.; Oliveira, S.M. Teixeira Therapeutic effect of localized vibration on alveolar bone of osteoporotic rats. PLoS ONE 2019, 14, e0211004. [Google Scholar]
- Prè, D.; Ceccarelli, G.; Visai, L.; Benedetti, L.; Imbriani, M.; De Angelis, M.G.C.; Magenes, G. High-Frequency Vibration Treatment of Human Bone Marrow Stromal Cells Increases Differentiation toward Bone Tissue. Bone Marrow Res. 2013, 2013, 803450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demiray, L.; ÖZÇİVİCİ, E. Bone marrow stem cells adapt to low-magnitude vibrations by altering their cytoskeleton during quiescence and osteogenesis. Turk. J. Biol. 2015, 39, 88–97. [Google Scholar] [CrossRef]
- Baskan, O.; Mese, G.; Ozcivici, E. Low-intensity vibrations normalize adipogenesis-induced morphological and molecular changes of adult mesenchymal stem cells. Proc. Inst. Mech. Eng. H 2017, 231, 160–168. [Google Scholar] [CrossRef]
- Bacabac, R.G.; Smit, T.H.; Van Loon, J.J.; Doulabi, B.Z.; Helder, M.; Klein-Nulend, J. Bone cell responses to high-frequency vibration stress: Does the nucleus oscillate within the cytoplasm? FASEB J. 2006, 20, 858–864. [Google Scholar] [CrossRef]
- Chen, X.; He, F.; Zhong, D.Y.; Luo, Z.P. Acoustic-frequency vibratory stimulation regulates the balance between osteogenesis and adipogenesis of human bone marrow-derived mesenchymal stem cells. Biomed. Res. Int. 2015, 2015, 540731. [Google Scholar] [CrossRef] [Green Version]
- Lau, E.; Al-Dujaili, S.; Guenther, A.; Liu, D.; Wang, L.; You, L. Effect of low-magnitude, high-frequency vibration on osteocytes in the regulation of osteoclasts. Bone 2010, 46, 1508–1515. [Google Scholar] [CrossRef] [Green Version]
- Tirkkonen, L.; Halonen, H.; Hyttinen, J.; Kuokkanen, H.; Sievänen, H.; Koivisto, A.-M.; Mannerström, B.; Sandor, G.; Suuronen, R.; Miettinen, S.; et al. The effects of vibration loading on adipose stem cell number, viability and differentiation towards bone-forming cells. J. R. Soc. Interface 2011, 8, 1736–1747. [Google Scholar] [CrossRef] [Green Version]
- Jaiswal, N.; Haynesworth, S.E.; Caplan, A.I.; Bruder, S.P. Osteogenic differentiation of purified, culture-expanded human mesenchymal stem cells in vitro. J. Cell. Biochem. 1997, 64, 295–312. [Google Scholar] [CrossRef]
- Zhou, Y.; Guan, X.; Zhu, Z.; Gao, S.; Zhang, C.H.; Li, C.; Zhou, K.; Hou, W.; Yu, H. Osteogenic differentiation of bone marrow-derived mesenchymal stromal cells on bone-derived scaffolds: Effect of microvibration and role of ERK1/2 activation. Eur. Cell. Mater. 2011, 22, 12–25. [Google Scholar] [CrossRef]
- Zhang, C.; Li, J.; Zhang, L.; Zhou, Y.; Hou, W.; Quan, H.; Li, X.; Chen, Y.; Yu, H. Effects of mechanical vibration on proliferation and osteogenic differentiation of human periodontal ligament stem cells. Arch. Oral Biol. 2012, 57, 1395–1407. [Google Scholar] [CrossRef] [PubMed]
- Del Angel-Mosqueda, C.; Gutierrez-Puente, Y.; López-Lozano, A.P.; Romero-Zavaleta, R.E.; Mendiola-Jiménez, A.; La Garza, C.E.M.-D.; Marquez-M, M.; De La Garza-Ramos, M.A. Epidermal growth factor enhances osteogenic differentiation of dental pulp stem cells in vitro. Head Face Med. 2015, 11, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Awais, S.; Balouch, S.S.; Riaz, N.; Choudhery, M.S. Human Dental Pulp Stem Cells Exhibit Osteogenic Differentiation Potential. Open Life Sci. 2020, 15, 229–236. [Google Scholar] [CrossRef] [PubMed]
- Choi, K.M.; Seo, Y.K.; Yoon, H.H.; Song, K.Y.; Kwon, S.Y.; Lee, H.S.; Park, J.K. Effects of mechanical stimulation on the proliferation of bone marrow-derived human mesenchymal stem cells. Biotechnol. Bioprocess. Eng. 2007, 12, 601–609. [Google Scholar] [CrossRef]
- Liu, S.; Fang, F.; Song, R.; Gao, X.; Jiang, M.; Cang, J. Sevoflurane affects neurogenesis through cell cycle arrest via inhibiting wnt/β-catenin signaling pathway in mouse neural stem cells. Life Sci. 2018, 209, 34–42. [Google Scholar] [CrossRef]
- Zhang, M.-F.; Cai, X.-L.; Jing, K.-P.; Pi, X.-X.; Liao, P.-Y.; Li, S.-J.; Li, W.; Cai, C.-C.; Quan, J.-H.; Fan, Y.-M.; et al. Differentiation Model Establishment and Differentiation-Related Protein Screening in Primary Cultured Human Sebocytes. Biomed. Res. Int. 2018, 2018, 7174561. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.-W.; Kim, H.J.; Lee, Y.S.; Park, H.-J.; Choi, J.-W.; Ha, J. Acteoside inhibits human promyelocytic HL-60 leukemia cell proliferation via inducing cell cycle arrest at G0/G1 phase and differentiation into monocyte. Carcinogenesis 2007, 28, 1928–1936. [Google Scholar] [CrossRef]
- Franceschi, R.T. The developmental control of osteoblast-specific gene expression: Role of specific transcription factors and the extracellular matrix environment. Crit. Rev. Oral Biol. Med. 1999, 10, 40–57. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.K.; Sowa, H.; Hinoi, E.; Ferron, M.; Ahn, J.D.; Confavreux, C.; Dacquin, R.; Mee, P.J.; McKee, M.D.; Jung, D.Y.; et al. Endocrine regulation of energy metabolism by the skeleton. Cell 2007, 130, 456–469. [Google Scholar] [CrossRef] [Green Version]
- Shu, B.; Zhang, M.; Xie, R.; Wang, M.; Jin, H.; Hou, W.; Tang, D.; Harris, S.E.; Mishina, Y.; O’Keefe, R.J.; et al. BMP2, but not BMP4, is crucial for chondrocyte proliferation and maturation during endochondral bone development. J. Cell Sci. 2011, 124, 3428–3440. [Google Scholar] [CrossRef] [Green Version]
- Ducy, P.; Zhang, R.; Geoffroy, V.; Ridall, A.L.; Karsenty, G. Osf2/Cbfa1: A transcriptional activator of osteoblast differentiation. Cell 1997, 89, 747–754. [Google Scholar] [CrossRef] [Green Version]
- Miyoshi, H.; Shimizu, K.; Kozu, T.; Maseki, N.; Kaneko, Y.; Ohki, M. t(8;21) breakpoints on chromosome 21 in acute myeloid leukemia are clustered within a limited region of a single gene, AML1. Proc. Natl. Acad. Sci. USA 1991, 88, 10431–10434. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, Z.; Lohmann, C.H.; Oefinger, J.; Bonewald, L.F.; Dean, D.D.; Boyan, B.D. Implant surface characteristics modulate differentiation behavior of cells in the osteoblastic lineage. Adv. Dent. Res. 1999, 13, 38–48. [Google Scholar] [CrossRef]
- Azuma, N.; Duzgun, S.A.; Ikeda, M.; Kito, H.; Akasaka, N.; Sasajima, T.; Sumpio, B.E. Endothelial cell response to different mechanical forces. J. Vasc. Surg. 2000, 32, 789–794. [Google Scholar] [CrossRef] [Green Version]
- Ferraro, J.T.; Daneshmand, M.; Bizios, R.; Rizzo, V. Depletion of plasma membrane cholesterol dampens hydrostatic pressure and shear stress-induced mechanotransduction pathways in osteoblast cultures. Am. J. Physiol. Cell Physiol. 2004, 286, C831–C839. [Google Scholar] [CrossRef]
- Lai, C.-F.; Chaudhary, L.; Fausto, A.; Halstead, L.R.; Ory, D.S.; Avioli, L.V.; Cheng, S.-L. Erk is essential for growth, differentiation, integrin expression, and cell function in human osteoblastic cells. J. Biol. Chem. 2001, 276, 14443–14450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oldenhof, A.D.; Shynlova, O.P.; Liu, M.; Langille, B.L.; Lye, S.J. Mitogen-activated protein kinases mediate stretch-induced c-fos mRNA expression in myometrial smooth muscle cells. Am. J. Physiol. Cell Physiol. 2002, 283, C1530–C1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uzer, G.; Pongkitwitoon, S.; Ete Chan, M.; Judex, S. Vibration induced osteogenic commitment of mesenchymal stem cells is enhanced by cytoskeletal remodeling but not fluid shear. J. Biomech. 2013, 46, 2296–2302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, M.; Wu, W.; Tan, L.; Mu, D.; Zhu, D.; Wang, J.; Zhao, B. Low-magnitude mechanical vibration regulates expression of osteogenic proteins in ovariectomized rats. Biochem. Biophys. Res. Commun. 2015, 465, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Baldión, P.A.; Velandia-Romero, M.L.; Castellanos, J.E. Odontoblast-Like Cells Differentiated from Dental Pulp Stem Cells Retain Their Phenotype after Subcultivation. Int. J. Cell Biol. 2018, 2018, 6853189. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Marker | Coll I | Coll III | Osteopontin | Osteonectin |
|---|---|---|---|---|
| Cont | + | + | + | - |
| 30 Hz | ++ | + | ++ | + |
| 45 Hz | ++ | ++ | +++ | ++ |
| 100 Hz | ++ | + | +++ | +++ |
| Gene | Forward (5′-3′) | Reverse (5′-3′) | Product Size | NCBI Accession Number |
|---|---|---|---|---|
| GAPDH | ACC ACA GTC CAT GCC ATC AC | TCC ACC ACC CTG TTG CTG TA | 452 | NM_001357943.2 |
| Osteonectin | CCA GAA CCA CCA CTG CAA AC | GGC AGG AAG AGT CGA AGG TC | 155 | NM_001309444.2 |
| Osteocalcin | CCA GGC GCT ACC TGT ATC AA | AGG GGA AGA GGA AAG AAG GG | 231 | NM_199173.6 |
| BMP2 | GTA CTA GCG ACA CCC ACA AC | GTC CAG CTG TAA GAG ACA CC | 316 | NM_001200.4 |
| ALP | ATCTCGTTGTCTGAGTACCAGTCC | TGGAGCTTCAGAAGCTCAACACCA | 454 | NM_001127501.4 |
| Runx2 | ACA GTA GAT GGA CCT CGG GA | ATA CTG GGA TGA GGA ATG CG | 113 | NM_001015051.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, W.; Eo, S.-R.; Choi, J.-H.; Kim, Y.-M.; Nam, M.-H.; Seo, Y.-K. The Osteogenic Differentiation of Human Dental Pulp Stem Cells through G0/G1 Arrest and the p-ERK/Runx-2 Pathway by Sonic Vibration. Int. J. Mol. Sci. 2021, 22, 10167. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810167
Lee W, Eo S-R, Choi J-H, Kim Y-M, Nam M-H, Seo Y-K. The Osteogenic Differentiation of Human Dental Pulp Stem Cells through G0/G1 Arrest and the p-ERK/Runx-2 Pathway by Sonic Vibration. International Journal of Molecular Sciences. 2021; 22(18):10167. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810167
Chicago/Turabian StyleLee, Won, Su-Rak Eo, Ju-Hye Choi, Yu-Mi Kim, Myeong-Hyun Nam, and Young-Kwon Seo. 2021. "The Osteogenic Differentiation of Human Dental Pulp Stem Cells through G0/G1 Arrest and the p-ERK/Runx-2 Pathway by Sonic Vibration" International Journal of Molecular Sciences 22, no. 18: 10167. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms221810167