
Influence of Serratia marcescens and Rhodococcus rhodnii on the Humoral Immunity of Rhodnius prolixus

 ,
, 
Abstract
:1. Introduction
2. Results
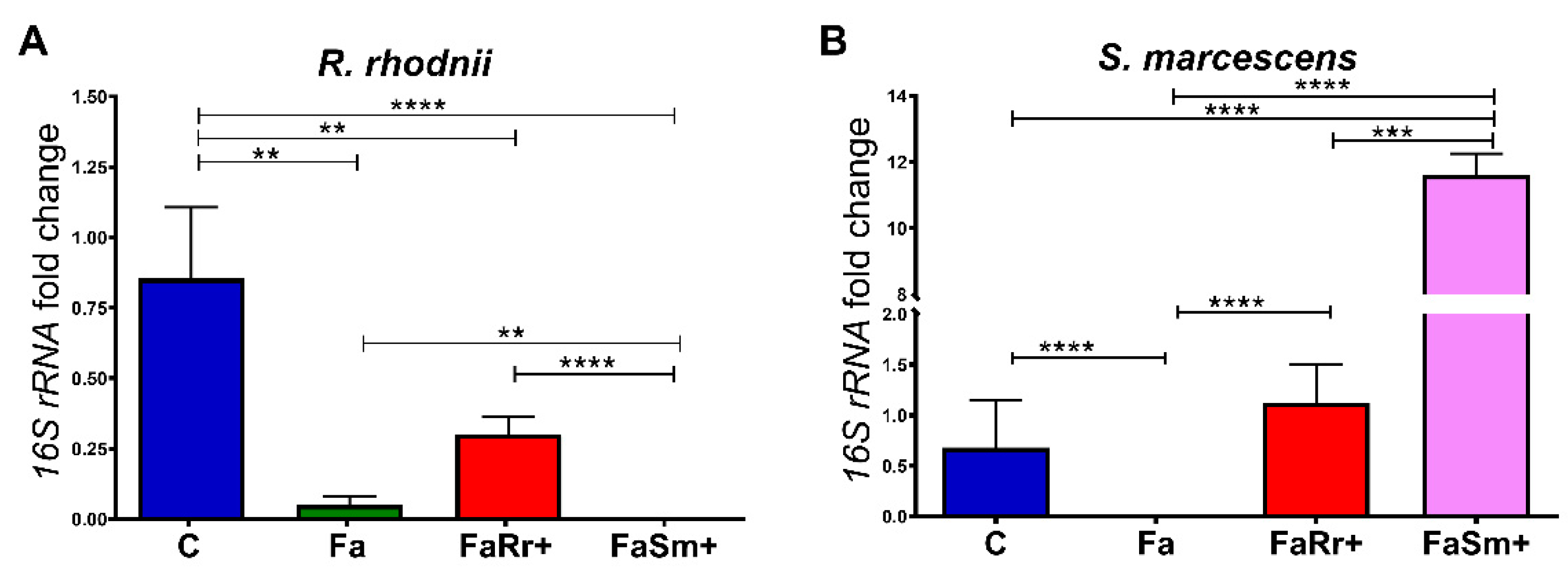
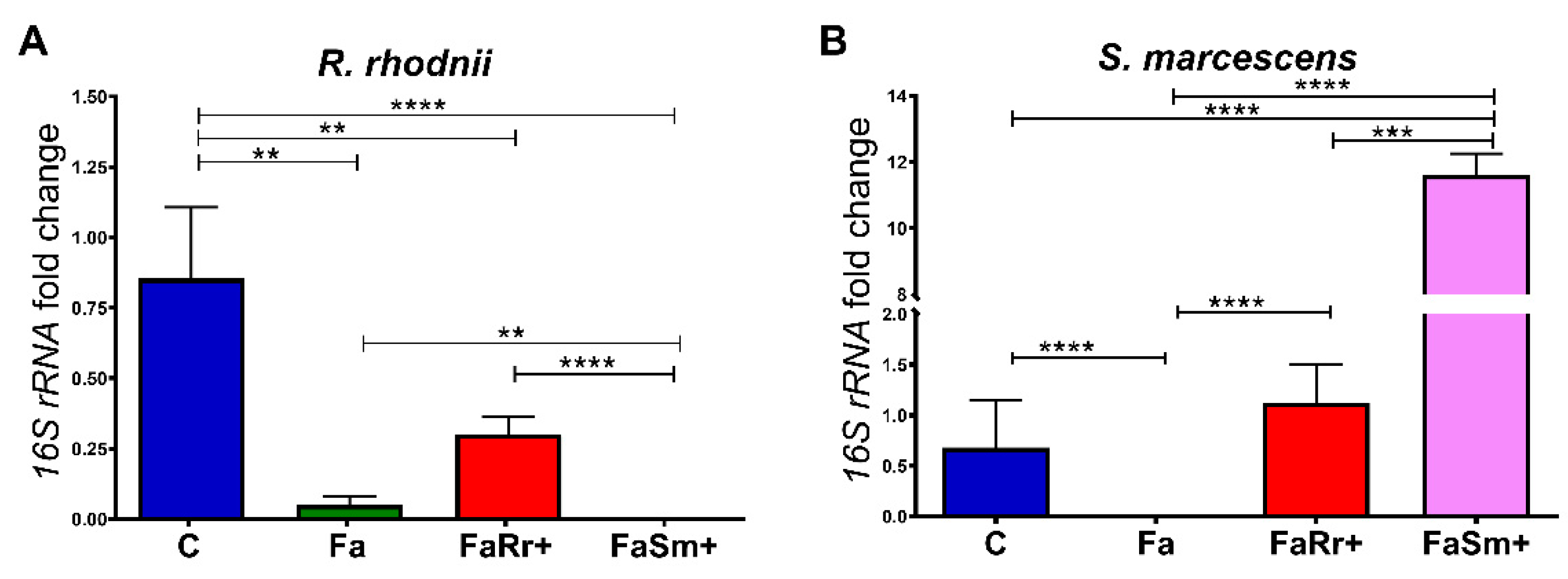
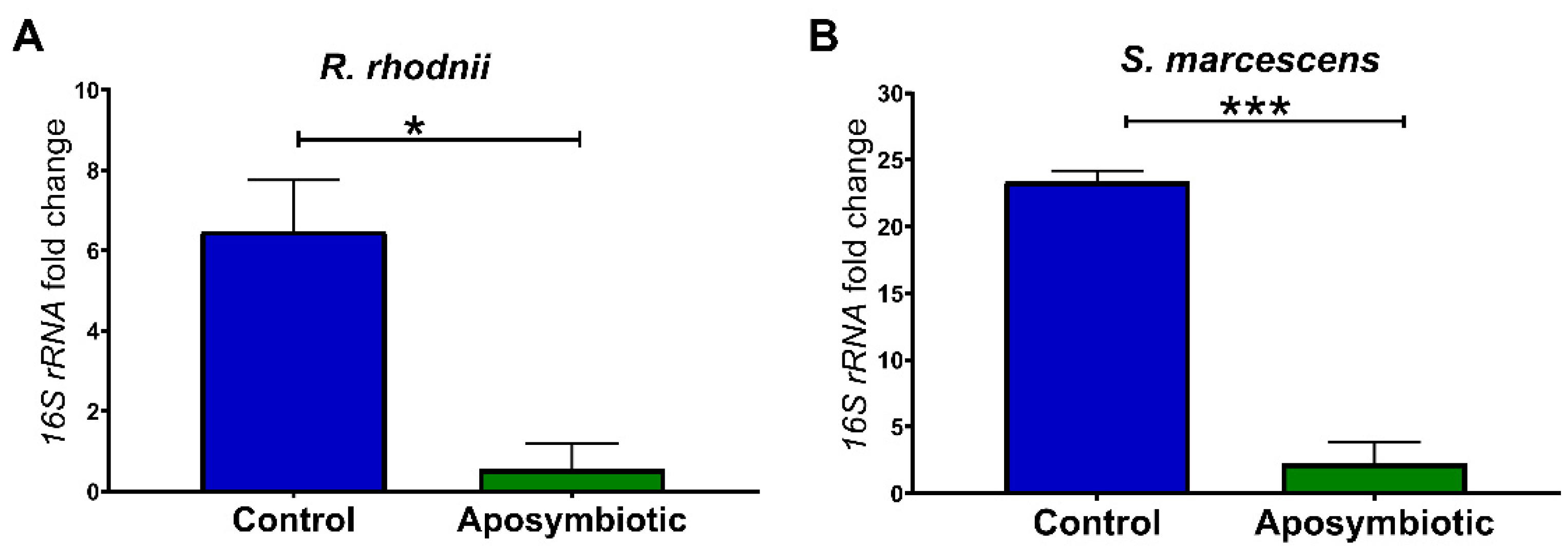
2.1. Quantification of Serratia Marcescens and Rhodococcus Rhodnii by qPCR
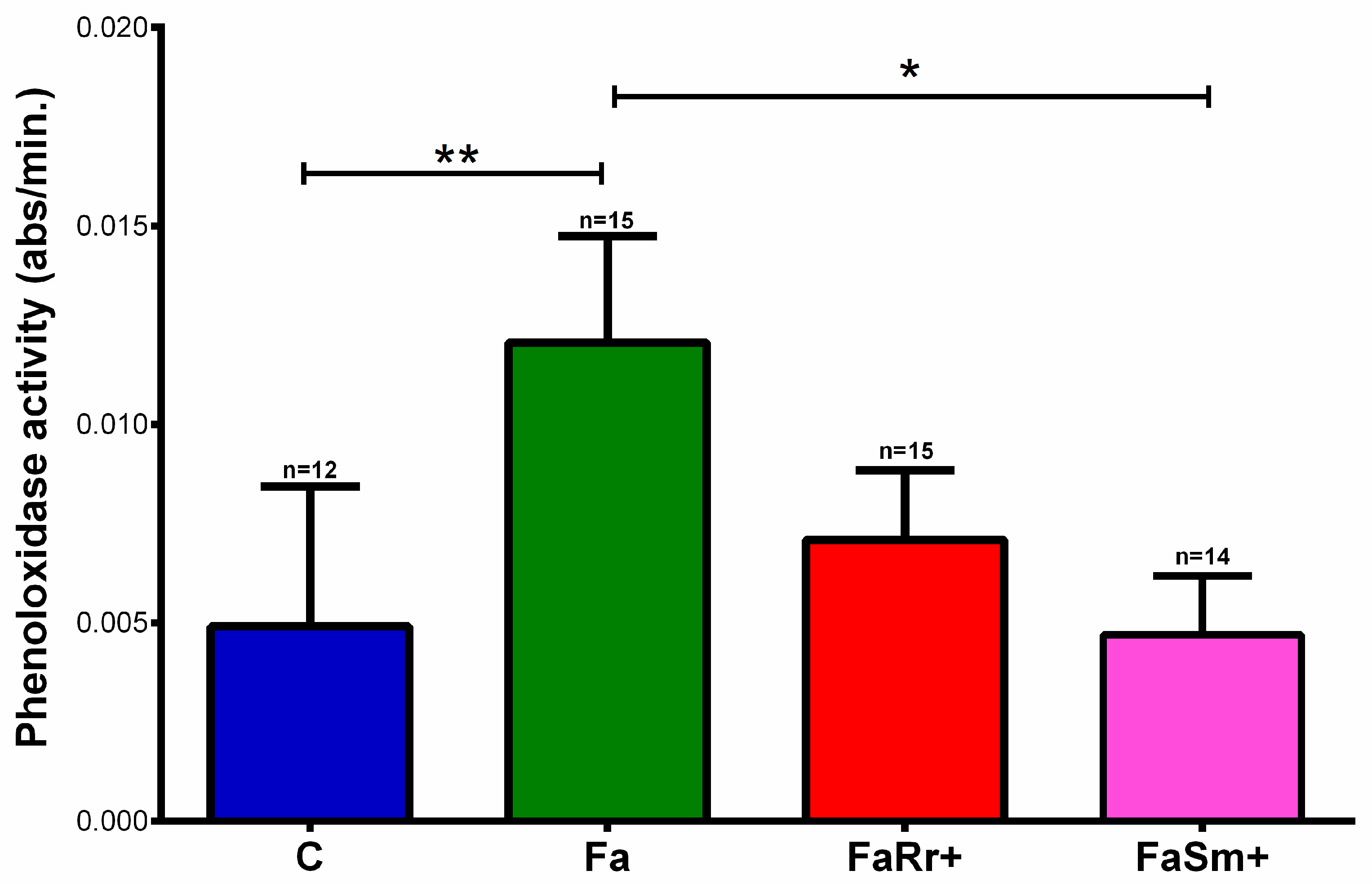
2.2. Phenoloxidase Enzyme Activity
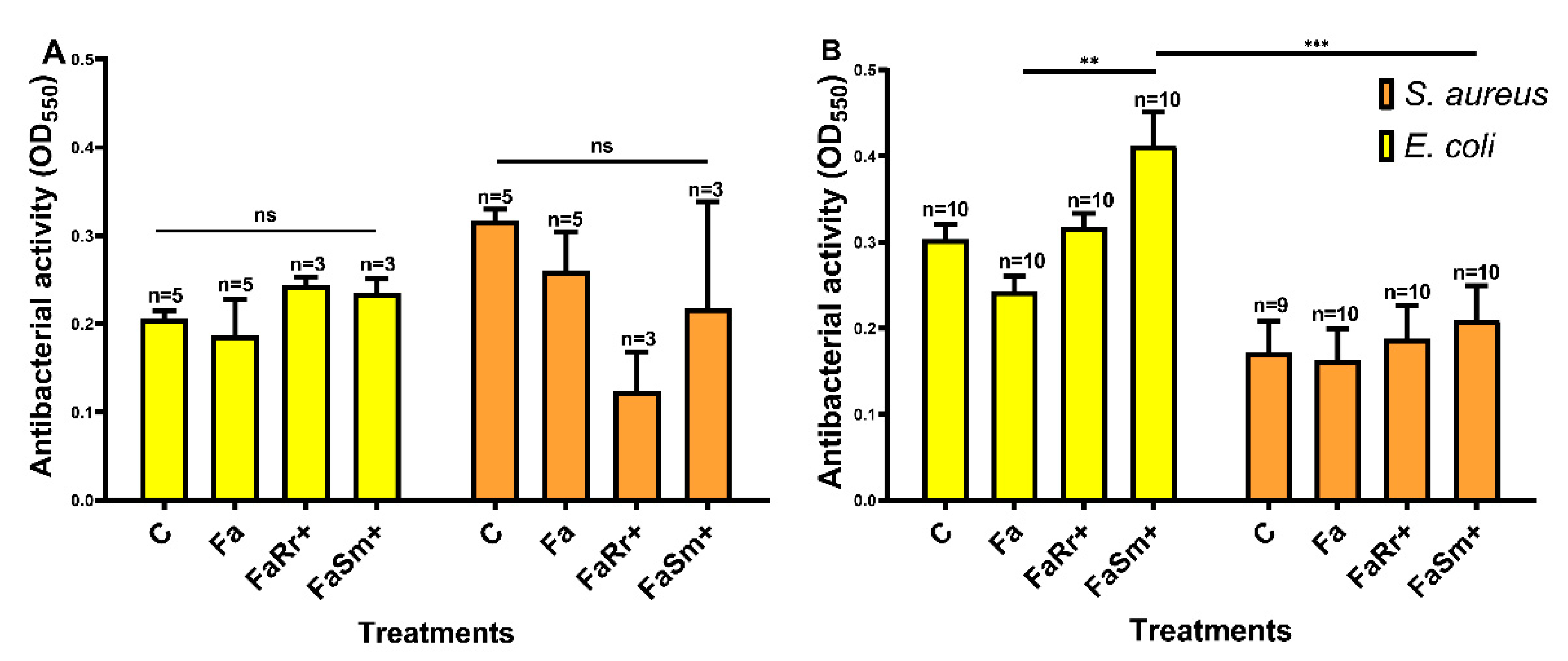
2.3. Antibacterial Activity
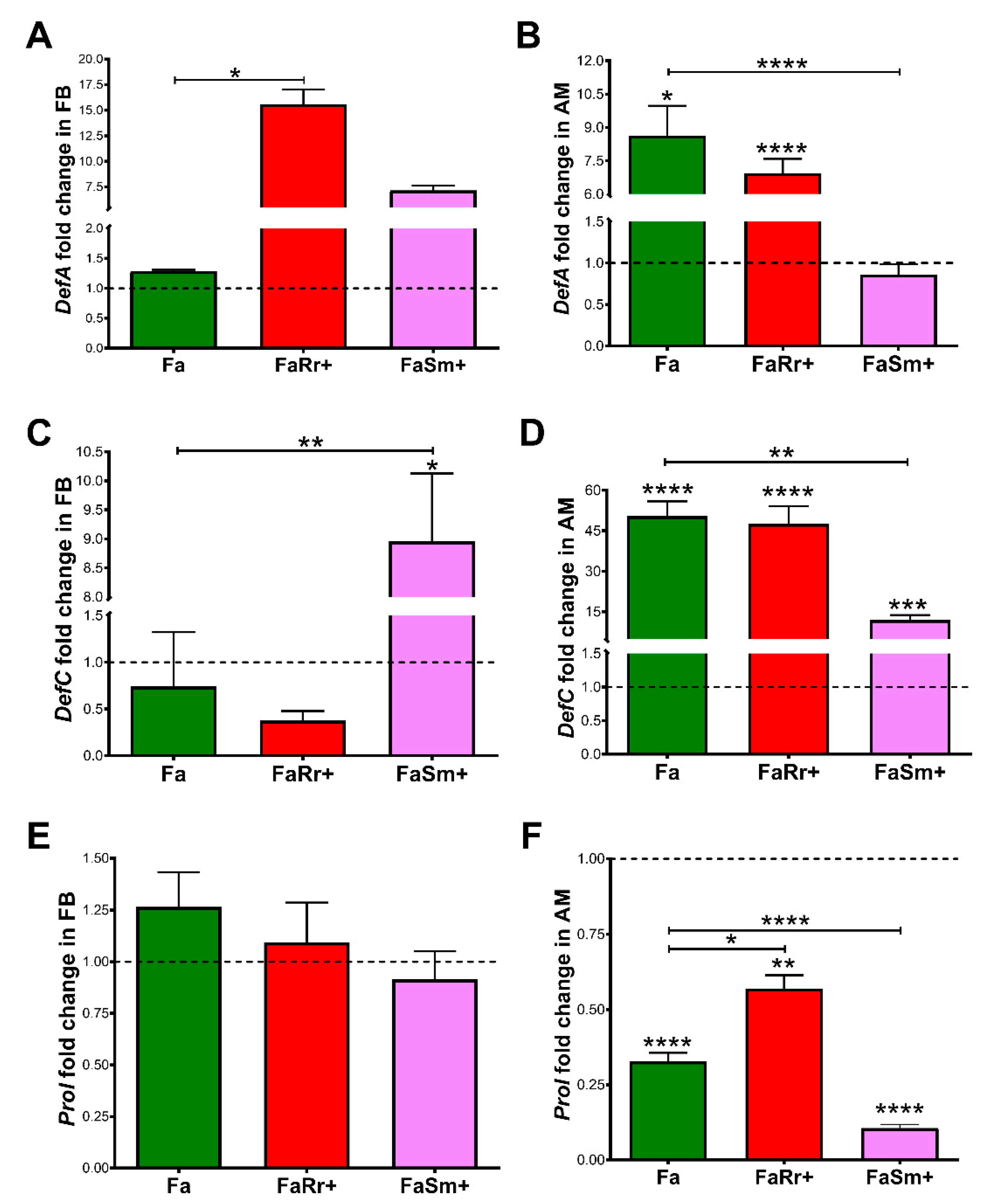
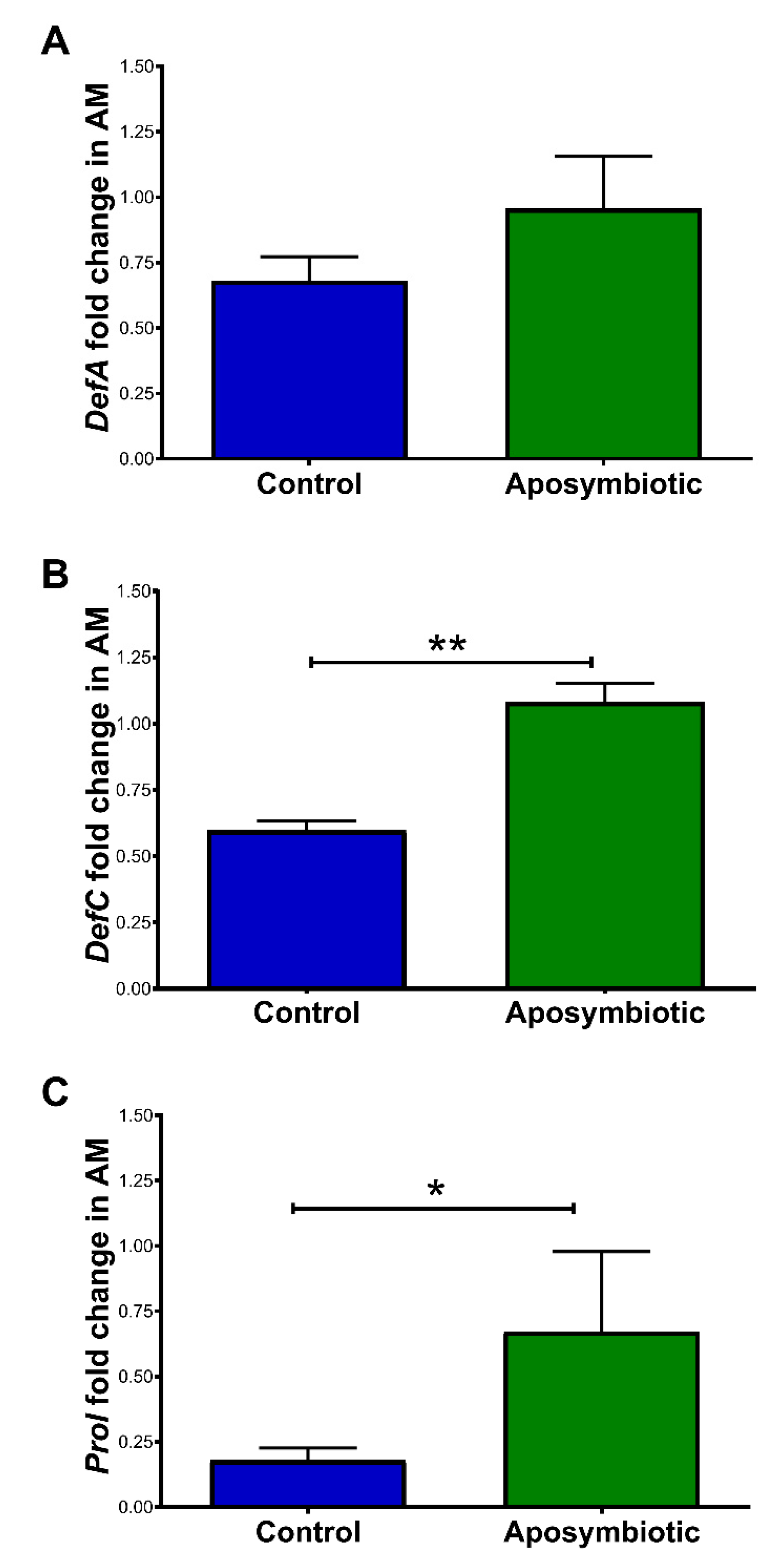
2.4. Antimicrobial Peptides (AMP) Gene Expression
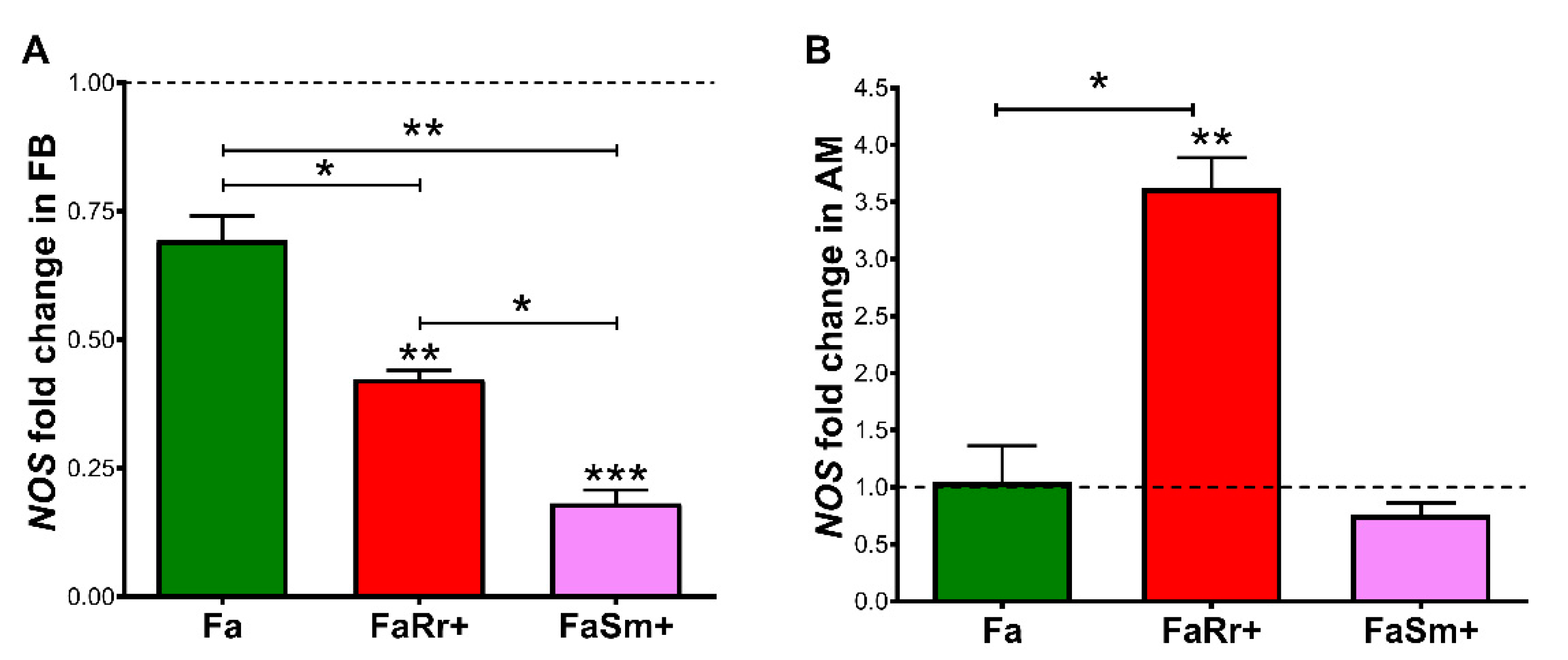
2.5. Nitric Oxide Synthase (NOS) Gene Expression
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Insects Maintenance
4.3. Bacteria Maintenance and Preparation for Recolonization
4.4. Insect Treatments and Recolonization
4.5. Aposymbiotic Nymphs
4.6. Antimicrobial Peptides (AMPs) and NOS Gene Expression, and Quantification of Serratia Marcescens and Rhodococcus Rhodnii by qPCR
4.7. Antibacterial Activity
4.8. Phenoloxidase (PO) Assay
4.9. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Chagas, C. Nova tripanozomiaze humana: Estudos sobre a morfologia e o ciclo evolutivo do Schizotrypanum cruzi n. gen., n. sp., agente etiologico de nova entidade morbida do homem. Memórias Inst. Oswaldo Cruz 1909, 1, 159–218. [Google Scholar] [CrossRef] [Green Version]
- WHO. Chagas Disease (American Trypanosomiasis). Available online: https://www.who.int/chagas/epidemiology/en/ (accessed on 1 June 2021).
- Costa, J.; Peterson, A.T. Ecological niche modeling as a tool for understanding distributions and interactions of vectors, hosts, and etiologic agents of Chagas disease. Adv. Exp. Med. Biol. 2012, 710, 59–70. [Google Scholar] [CrossRef]
- Coura, J.R. The main sceneries of Chagas disease transmission. The vectors, blood and oral transmissions—A comprehensive review. Memórias Inst. Oswaldo Cruz 2015, 110, 277–282. [Google Scholar] [CrossRef] [Green Version]
- Garcia, E.S.; Castro, D.P.; Figueiredo, M.B.; Azambuja, P. Parasite-mediated interactions within the insect vector: Trypanosoma rangeli strategies. Parasites Vectors 2012, 5, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azambuja, P.; Garcia, E.S.; Ratcliffe, N.A. Gut microbiota and parasite transmission by insect vectors. Trends Parasitol. 2005, 21, 568–572. [Google Scholar] [CrossRef] [PubMed]
- Vieira, C.S.; Waniek, P.J.; Castro, D.P.; Mattos, D.P.; Moreira, O.C.; Azambuja, P. Impact of Trypanosoma cruzi on antimicrobial peptide gene expression and activity in the fat body and midgut of Rhodnius prolixus. Parasites Vectors 2016, 9, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kollien, A.H.; Schaub, G.A. The development of Trypanosoma cruzi in triatominae. Parasitol. Today 2000, 16, 381–387. [Google Scholar] [CrossRef]
- Nogueira, N.P.; Saraiva, F.M.; Sultano, P.E.; Cunha, P.R.; Laranja, G.A.; Justo, G.A.; Sabino, K.C.; Coelho, M.G.; Rossini, A.; Atella, G.C.; et al. Proliferation and differentiation of Trypanosoma cruzi inside its vector have a new trigger: Redox status. PLoS ONE 2015, 10, e0116712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shikanai-Yasuda, M.A.; Marcondes, C.B.; Guedes, L.A.; Siqueira, G.S.; Barone, A.A.; Dias, J.C.; Amato Neto, V.; Tolezano, J.E.; Peres, B.A.; Arruda Júnior, E.R.; et al. Possible oral transmission of acute Chagas’ disease in Brazil. Rev. Inst. Med. Trop. Sao Paulo 1991, 33, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Futo, M.; Armitage, S.A.; Kurtz, J. Microbiota Plays a Role in Oral Immune Priming in Tribolium castaneum. Front. Microbiol. 2015, 6, 1383. [Google Scholar] [CrossRef]
- Morella, N.M.; Koskella, B. The Value of a Comparative Approach to Understand the Complex Interplay between Microbiota and Host Immunity. Front. Immunol. 2017, 8, 1114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salcedo-Porras, N.; Umaña-Diaz, C.; Bitencourt, R.O.B.; Lowenberger, C. The Role of Bacterial Symbionts in Triatomines: An Evolutionary Perspective. Microorganisms 2020, 8, 1438. [Google Scholar] [CrossRef]
- Douglas, A.E. Multiorganismal insects: Diversity and function of resident microorganisms. Annu. Rev. Entomol. 2015, 60, 17–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habineza, P.; Muhammad, A.; Ji, T.; Xiao, R.; Yin, X.; Hou, Y.; Shi, Z. The Promoting Effect of Gut Microbiota on Growth and Development of Red Palm Weevil, Rhynchophorus ferrugineus (Olivier) (Coleoptera: Dryophthoridae) by Modulating Its Nutritional Metabolism. Front. Microbiol. 2019, 10, 1212. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, A.; Habineza, P.; Ji, T.; Hou, Y.; Shi, Z. Intestinal Microbiota Confer Protection by Priming the Immune System of Red Palm Weevil Rhynchophorus ferrugineus Olivier (Coleoptera: Dryophthoridae). Front. Physiol. 2019, 10, 1303. [Google Scholar] [CrossRef]
- Azambuja, P.; Feder, D.; Garcia, E.S. Isolation of Serratia marcescens in the midgut of Rhodnius prolixus: Impact on the establishment of the parasite Trypanosoma cruzi in the vector. Exp. Parasitol. 2004, 107, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Castro, D.P.; Moraes, C.S.; Gonzalez, M.S.; Ratcliffe, N.A.; Azambuja, P.; Garcia, E.S. Trypanosoma cruzi immune response modulation decreases microbiota in Rhodnius prolixus gut and is crucial for parasite survival and development. PLoS ONE 2012, 7, e36591. [Google Scholar] [CrossRef] [Green Version]
- Castro, D.P.; Seabra, S.H.; Garcia, E.S.; de Souza, W.; Azambuja, P. Trypanosoma cruzi: Ultrastructural studies of adhesion, lysis and biofilm formation by Serratia marcescens. Exp. Parasitol. 2007, 117, 201–207. [Google Scholar] [CrossRef]
- da Mota, F.F.; Castro, D.P.; Vieira, C.S.; Gumiel, M.; de Albuquerque, J.P.; Carels, N.; Azambuja, P. In vitro Trypanocidal Activity, Genomic Analysis of Isolates, and in vivo Transcription of Type VI Secretion System of Serratia marcescens Belonging to the Microbiota of Rhodnius prolixus Digestive Tract. Front. Microbiol. 2018, 9, 3205. [Google Scholar] [CrossRef] [PubMed]
- da Mota, F.F.; Marinho, L.P.; Moreira, C.J.; Lima, M.M.; Mello, C.B.; Garcia, E.S.; Carels, N.; Azambuja, P. Cultivation-independent methods reveal differences among bacterial gut microbiota in triatomine vectors of Chagas disease. PLoS Negl. Trop. Dis. 2012, 6, e1631. [Google Scholar] [CrossRef] [Green Version]
- Dias, F.A.; Gandara, A.C.; Perdomo, H.D.; Gonçalves, R.S.; Oliveira, C.R.; Oliveira, R.L.; Citelli, M.; Polycarpo, C.R.; Santesmasses, D.; Mariotti, M.; et al. Identification of a selenium-dependent glutathione peroxidase in the blood-sucking insect Rhodnius prolixus. Insect Biochem. Mol. Biol. 2016, 69, 105–114. [Google Scholar] [CrossRef] [PubMed]
- Eichler, S.; Schaub, G.A. Development of symbionts in triatomine bugs and the effects of infections with trypanosomatids. Exp. Parasitol. 2002, 100, 17–27. [Google Scholar] [CrossRef]
- Gumiel, M.; da Mota, F.F.; Rizzo Vde, S.; Sarquis, O.; de Castro, D.P.; Lima, M.M.; Garcia Ede, S.; Carels, N.; Azambuja, P. Characterization of the microbiota in the guts of Triatoma brasiliensis and Triatoma pseudomaculata infected by Trypanosoma cruzi in natural conditions using culture independent methods. Parasit. Vectors 2015, 8, 245. [Google Scholar] [CrossRef] [PubMed]
- Duncan, J.T. On a Bactericidal Principle present in the Alimentary Canal of Insects and Arac. Parasitology 1926, 18, 238–251. [Google Scholar] [CrossRef]
- Wigglesworth, V.B. symbiotic bacteria in a blood-sucking insect, Rhodnius prolixus stal. (hemiptera, triatomidae). Parasitology 1936, 28, 284–289. [Google Scholar] [CrossRef]
- Auden, D.T. Studies on the development of Rhodnius prolixus and the effects of its symbiote Nocardia rhodnii. J. Med. EntoMol. 1974, 11, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Baines, S. The role of the symbiotic bacteria in the nutrition of Rhodnius prolixus (hemiptera). Exp. Biol. 1956, 33, 533–541. [Google Scholar] [CrossRef]
- Ben-Yakir, D. Growth retardation of Rhodnius prolixus symbionts by immunizing host against Nocardia (Rhodococcus) rhodnii. J. Insect Physiol. 1987, 33, 379–383. [Google Scholar] [CrossRef]
- Lake, P.; Friend, W.G. The use of artificial diets to determine some of the effects of Nocardia rhodnii on the development of Rhodnius prolixus. J. Insect Physiol. 1968, 14, 543–562. [Google Scholar] [CrossRef]
- Pachebat, J.A.; van Keulen, G.; Whitten, M.M.; Girdwood, S.; Del Sol, R.; Dyson, P.J.; Facey, P.D. Draft Genome Sequence of Rhodococcus rhodnii Strain LMG5362, a Symbiont of Rhodnius prolixus (Hemiptera, Reduviidae, Triatominae), the Principle Vector of Trypanosoma cruzi. Genome Announc. 2013, 1, e00329-13. [Google Scholar] [CrossRef]
- Tobias, N.J.; Eberhard, F.E.; Guarneri, A.A. Enzymatic biosynthesis of B-complex vitamins is supplied by diverse microbiota in the Rhodnius prolixus anterior midgut following Trypanosoma cruzi infection. Comput. Struct. Biotechnol. J. 2020, 18, 3395–3401. [Google Scholar] [CrossRef]
- Durvasula, R.V.; Gumbs, A.; Panackal, A.; Kruglov, O.; Taneja, J.; Kang, A.S.; Cordon-Rosales, C.; Richards, F.F.; Whitham, R.G.; Beard, C.B. Expression of a functional antibody fragment in the gut of Rhodnius prolixus via transgenic bacterial symbiont Rhodococcus rhodnii. Med. Vet. EntoMol. 1999, 13, 115–119. [Google Scholar] [CrossRef]
- Jose, C.; Klein, N.; Wyss, S.; Fieck, A.; Hurwitz, I.; Durvasula, R. Recombinant Arthrobacter β-1, 3-glucanase as a potential effector molecule for paratransgenic control of Chagas disease. Parasit. Vectors 2013, 6, 65. [Google Scholar] [CrossRef] [Green Version]
- Taracena, M.L.; Oliveira, P.L.; Almendares, O.; Umaña, C.; Lowenberger, C.; Dotson, E.M.; Paiva-Silva, G.O.; Pennington, P.M. Genetically modifying the insect gut microbiota to control Chagas disease vectors through systemic RNAi. PLoS Negl. Trop. Dis. 2015, 9, e0003358. [Google Scholar] [CrossRef] [Green Version]
- Petersen, L.M.; Tisa, L.S. Friend or foe? A review of the mechanisms that drive Serratia towards diverse lifestyles. Can. J. Microbiol. 2013, 59, 627–640. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Dos-Santos, A.L.A.; Huang, W.; Liu, K.C.; Oshaghi, M.A.; Wei, G.; Agre, P.; Jacobs-Lorena, M. Driving mosquito refractoriness to Plasmodium falciparum with engineered symbiotic bacteria. Science 2017, 357, 1399–1402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, E.S.; Castro, D.P.; Figueiredo, M.B.; Azambuja, P. Immune homeostasis to microorganisms in the guts of triatomines (Reduviidae)—A review. Mem. Inst. Oswaldo Cruz 2010, 105, 605–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, C.S.; Waniek, P.J.; Mattos, D.P.; Castro, D.P.; Mello, C.B.; Ratcliffe, N.A.; Garcia, E.S.; Azambuja, P. Humoral responses in Rhodnius prolixus: Bacterial feeding induces differential patterns of antibacterial activity and enhances mRNA levels of antimicrobial peptides in the midgut. Parasit. Vectors 2014, 7, 232. [Google Scholar] [CrossRef] [Green Version]
- Beard, C.B.; Durvasula, R.V.; Richards, F.F. Bacterial symbiosis in arthropods and the control of disease transmission. Emerg. Infect Dis. 1998, 4, 581–591. [Google Scholar] [CrossRef]
- Harington, J.S. Studies on Rhodnius prolixus: Growth and development of normal and sterile bugs, and the symbiotic relationship. Parasitology 1960, 50, 279–286. [Google Scholar] [CrossRef] [PubMed]
- Bauer, E.; Salem, H.; Marz, M.; Vogel, H.; Kaltenpoth, M. Transcriptomic immune response of the cotton stainer Dysdercus fasciatus to experimental elimination of vitamin-supplementing intestinal symbionts. PLoS ONE 2014, 9, e114865. [Google Scholar] [CrossRef] [PubMed]
- Becker, T.; Loch, G.; Beyer, M.; Zinke, I.; Aschenbrenner, A.C.; Carrera, P.; Inhester, T.; Schultze, J.L.; Hoch, M. FOXO-dependent regulation of innate immune homeostasis. Nature 2010, 463, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Flatt, T.; Heyland, A.; Rus, F.; Porpiglia, E.; Sherlock, C.; Yamamoto, R.; Garbuzov, A.; Palli, S.R.; Tatar, M.; Silverman, N. Hormonal regulation of the humoral innate immune response in Drosophila melanogaster. J. Exp. Biol. 2008, 211, 2712–2724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garrett, W.S.; Gordon, J.I.; Glimcher, L.H. Homeostasis and inflammation in the intestine. Cell 2010, 140, 859–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krams, I.A.; Kecko, S.; Jõers, P.; Trakimas, G.; Elferts, D.; Krams, R.; Luoto, S.; Rantala, M.J.; Inashkina, I.; Gudrā, D.; et al. Microbiome symbionts and diet diversity incur costs on the immune system of insect larvae. J. Exp. Biol. 2017, 220, 4204–4212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, K.; Luan, X.; Liu, Q.; Wang, J.; Chang, X.; Snijders, A.M.; Mao, J.H.; Secombe, J.; Dan, Z.; Chen, J.H.; et al. Drosophila Histone Demethylase KDM5 Regulates Social Behavior through Immune Control and Gut Microbiota Maintenance. Cell Host Microbe 2019, 25, 537–552. [Google Scholar] [CrossRef] [Green Version]
- Salcedo-Porras, N.; Guarneri, A.; Oliveira, P.L.; Lowenberger, C. Rhodnius prolixus: Identification of missing components of the IMD immune signaling pathway and functional characterization of its role in eliminating bacteria. PLoS ONE 2019, 14, e0214794. [Google Scholar] [CrossRef] [PubMed]
- Vieira, C.S.; Mattos, D.P.; Waniek, P.J.; Santangelo, J.M.; Figueiredo, M.B.; Gumiel, M.; da Mota, F.F.; Castro, D.P.; Garcia, E.S.; Azambuja, P. Rhodnius prolixus interaction with Trypanosoma rangeli: Modulation of the immune system and microbiota population. Parasit. Vectors 2015, 8, 135. [Google Scholar] [CrossRef] [Green Version]
- Telleria, E.L.; Sant’Anna, M.R.; Alkurbi, M.O.; Pitaluga, A.N.; Dillon, R.J.; Traub-Csekö, Y.M. Bacterial feeding, Leishmania infection and distinct infection routes induce differential defensin expression in Lutzomyia longipalpis. Parasit. Vectors 2013, 6, 12. [Google Scholar] [CrossRef] [Green Version]
- Ursic-Bedoya, R.; Buchhop, J.; Joy, J.B.; Durvasula, R.; Lowenberger, C. Prolixicin: A novel antimicrobial peptide isolated from Rhodnius prolixus with differential activity against bacteria and Trypanosoma cruzi. Insect Mol. Biol. 2011, 20, 775–786. [Google Scholar] [CrossRef]
- Kim, J.K.; Lee, J.B.; Huh, Y.R.; Jang, H.A.; Kim, C.H.; Yoo, J.W.; Lee, B.L. Burkholderia gut symbionts enhance the innate immunity of host Riptortus pedestris. Dev. Comp. Immunol. 2015, 53, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Weiss, B.; Aksoy, S. Microbiome influences on insect host vector competence. Trends Parasitol. 2011, 27, 514–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitten, M.; Sun, F.; Tew, I.; Schaub, G.; Soukou, C.; Nappi, A.; Ratcliffe, N. Differential modulation of Rhodnius prolixus nitric oxide activities following challenge with Trypanosoma rangeli, T. cruzi and bacterial cell wall components. Insect Biochem. Mol. Biol. 2007, 37, 440–452. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Gilbreath, T.M., 3rd; Kukutla, P.; Yan, G.; Xu, J. Dynamic gut microbiome across life history of the malaria mosquito Anopheles gambiae in Kenya. PLoS ONE 2011, 6, e24767. [Google Scholar] [CrossRef] [Green Version]
- Batista, K.; Vieira, C.S.; Florentino, E.B.; Caruso, K.F.B.; Teixeira, P.T.P.; Moraes, C.D.S.; Genta, F.A.; de Azambuja, P.; de Castro, D.P. Nitric oxide effects on Rhodnius prolixus’s immune responses, gut microbiota and Trypanosoma cruzi development. J. Insect Physiol. 2020, 126, 104100. [Google Scholar] [CrossRef]
- Dennison, N.J.; Jupatanakul, N.; Dimopoulos, G. The mosquito microbiota influences vector competence for human pathogens. Curr. Opin. Insect Sci. 2014, 3, 6–13. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.E. The molecular basis of bacterial-insect symbiosis. J. Mol. Biol. 2014, 426, 3830–3837. [Google Scholar] [CrossRef] [Green Version]
- Ricci, I.; Valzano, M.; Ulissi, U.; Epis, S.; Cappelli, A.; Favia, G. Symbiotic control of mosquito borne disease. Pathog. Glob. Health 2012, 106, 380–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Y.; Manfredini, F.; Dimopoulos, G. Implication of the mosquito midgut microbiota in the defense against malaria parasites. PLoS Pathog. 2009, 5, e1000423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bahia, A.C.; Dong, Y.; Blumberg, B.J.; Mlambo, G.; Tripathi, A.; BenMarzouk-Hidalgo, O.J.; Chandra, R.; Dimopoulos, G. Exploring Anopheles gut bacteria for Plasmodium blocking activity. Environ. Microbiol. 2014, 16, 2980–2994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, W.; Wang, S.; Jacobs-Lorena, M. Use of Microbiota to Fight Mosquito-Borne Disease. Front. Genet. 2020, 11, 196. [Google Scholar] [CrossRef] [PubMed]
- Dotson, E.M.; Plikaytis, B.; Shinnick, T.M.; Durvasula, R.V.; Beard, C.B. Transformation of Rhodococcus rhodnii, a symbiont of the Chagas disease vector Rhodnius prolixus, with integrative elements of the L1 mycobacteriophage. Infect Genet. Evol. 2003, 3, 103–109. [Google Scholar] [CrossRef]
- Fieck, A.; Hurwitz, I.; Kang, A.S.; Durvasula, R. Trypanosoma cruzi: Synergistic cytotoxicity of multiple amphipathic anti-microbial peptides to T. cruzi and potential bacterial hosts. Exp. Parasitol. 2010, 125, 342–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matthews, S.; Rao, V.S.; Durvasula, R.V. Modeling horizontal gene transfer (HGT) in the gut of the Chagas disease vector Rhodnius prolixus. Parasit. Vectors 2011, 4, 77. [Google Scholar] [CrossRef] [Green Version]
- Azambuja, P.; Garcia, E.S. Care and maintenance of triatomine colonies. In Molecular Biology of Insect Disease Vectors: A Methods Manual; Crampton, J., Beard, C., Louis, C., Eds.; Springer: London, UK, 1997; pp. 55–64. [Google Scholar]
- Genta, F.A.; Souza, R.S.; Garcia, E.S.; Azambuja, P. Phenol oxidases from Rhodnius prolixus: Temporal and tissue expression pattern and regulation by ecdysone. J. Insect Physiol. 2010, 56, 1253–1259. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-ΔΔC(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Insect Groups | 4th Instar | 5th Instar |
|---|---|---|
| Antibiotic Treatment † | Bacteria Recolonization | |
| Control (C) | _ | _ |
| Antibiotics (Fa) | + | _ |
| S. marcescens + antibiotics (FaSm+) †† | + | + |
| R. rhodnii + antibiotics (FaRr+) ††† | + | + |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Batista, K.K.S.; Vieira, C.S.; Figueiredo, M.B.; Costa-Latgé, S.G.; Azambuja, P.; Genta, F.A.; Castro, D.P. Influence of Serratia marcescens and Rhodococcus rhodnii on the Humoral Immunity of Rhodnius prolixus. Int. J. Mol. Sci. 2021, 22, 10901. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222010901
Batista KKS, Vieira CS, Figueiredo MB, Costa-Latgé SG, Azambuja P, Genta FA, Castro DP. Influence of Serratia marcescens and Rhodococcus rhodnii on the Humoral Immunity of Rhodnius prolixus. International Journal of Molecular Sciences. 2021; 22(20):10901. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222010901
Chicago/Turabian StyleBatista, Kate K. S., Cecília S. Vieira, Marcela B. Figueiredo, Samara G. Costa-Latgé, Patrícia Azambuja, Fernando A. Genta, and Daniele P. Castro. 2021. "Influence of Serratia marcescens and Rhodococcus rhodnii on the Humoral Immunity of Rhodnius prolixus" International Journal of Molecular Sciences 22, no. 20: 10901. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222010901