
Nkx2.9 Contributes to Mid-Hindbrain Patterning by Regulation of mdDA Neuronal Cell-Fate and Repression of a Hindbrain-Specific Cell-Fate

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
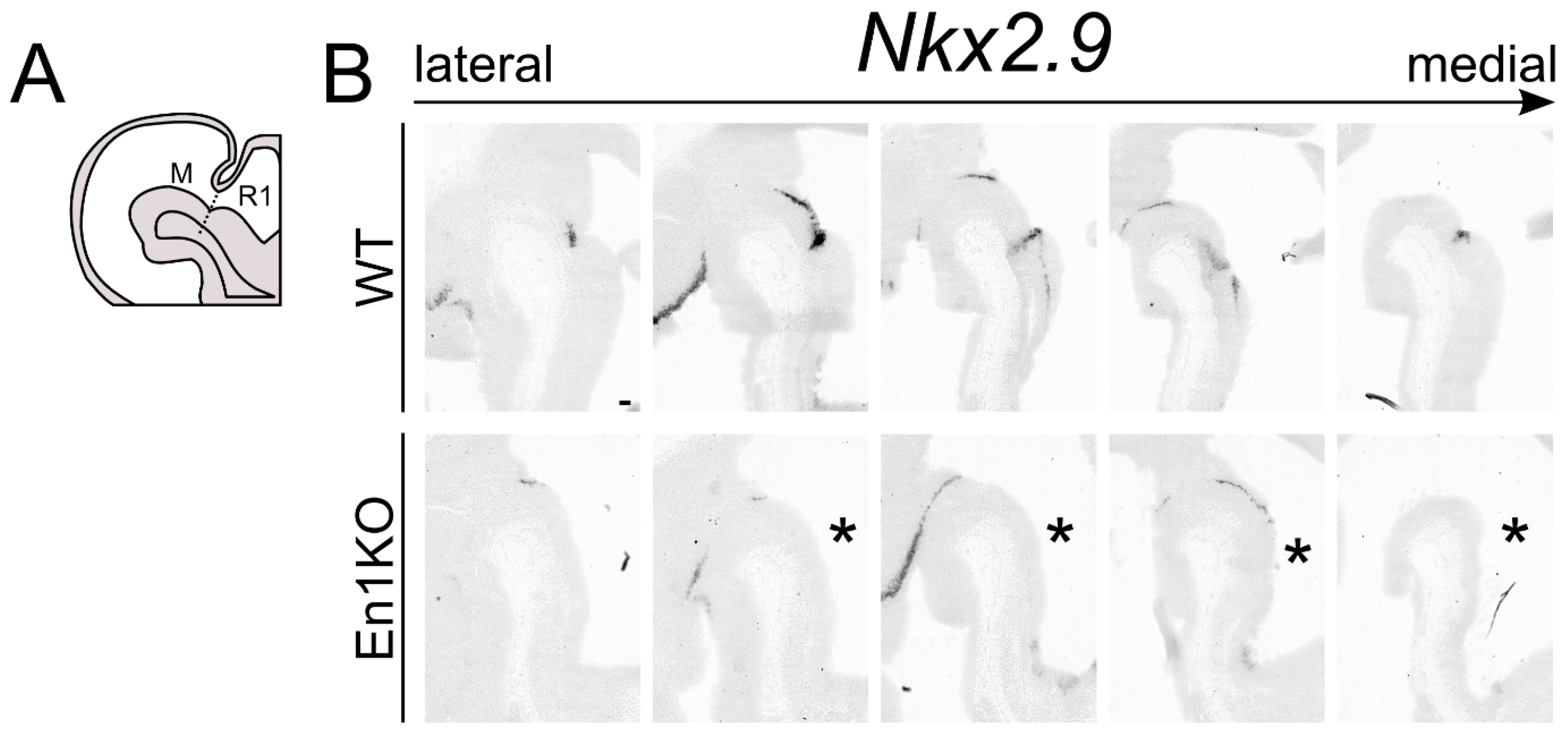
2.1. Nkx2.9 Transcript Expression in Mesencephalon and Metencephalon Is Dependent on En1
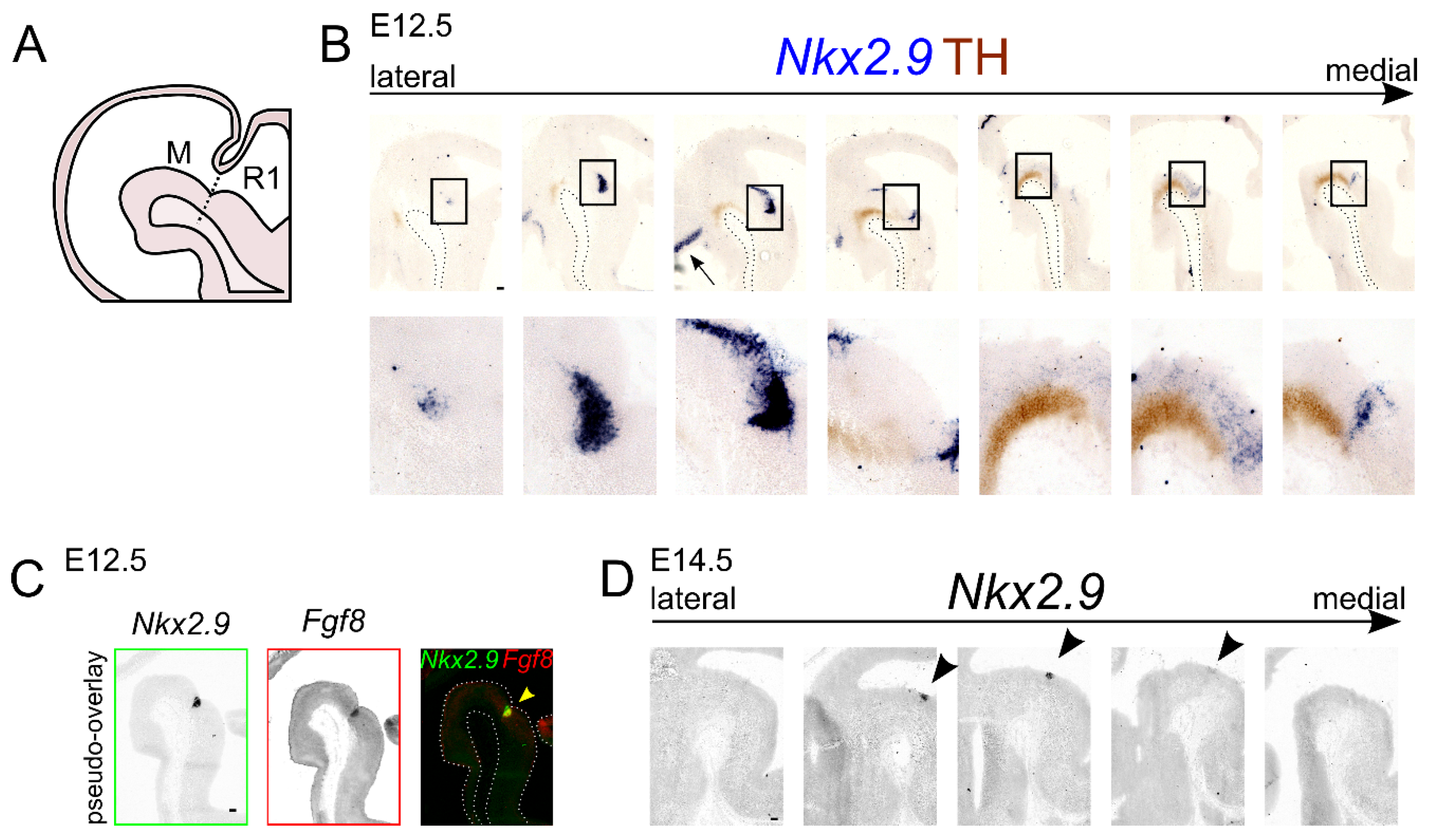
2.2. Nkx2.9 Transcript Expression Is Present in the mdDA Precursor Region
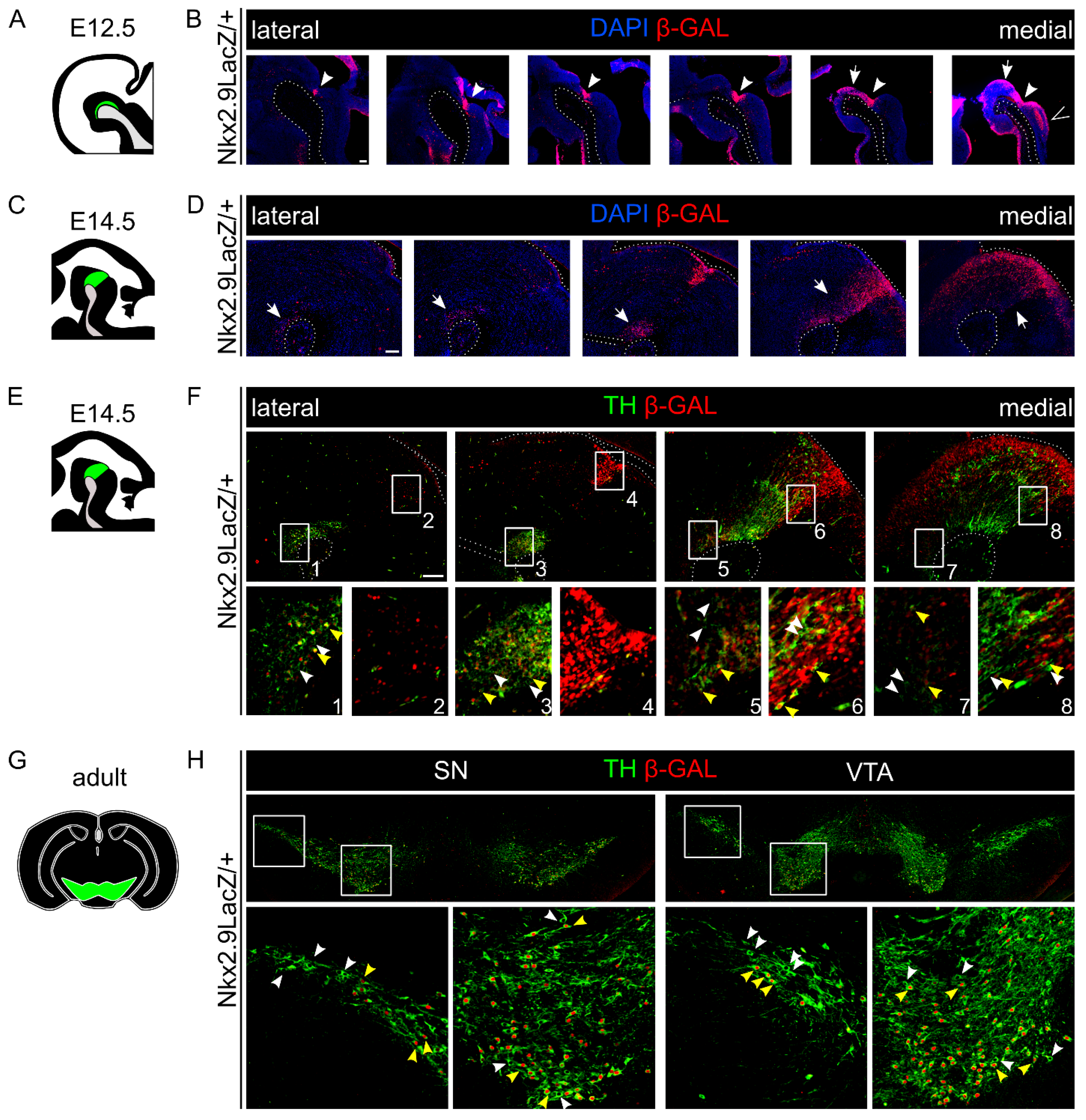
2.3. A Major Subset of mdDA Neurons Originates from a Nkx2.9-Positive Lineage
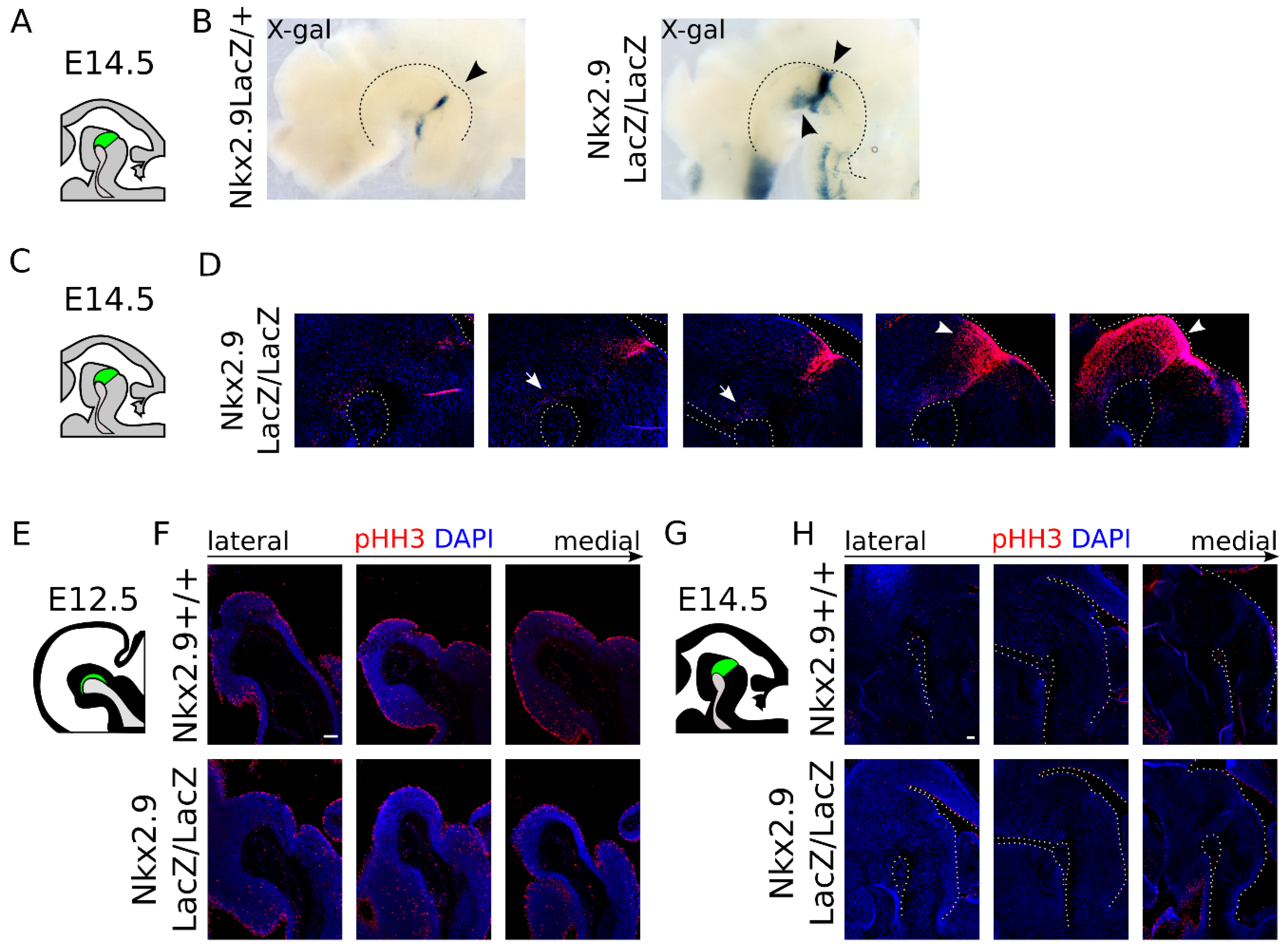
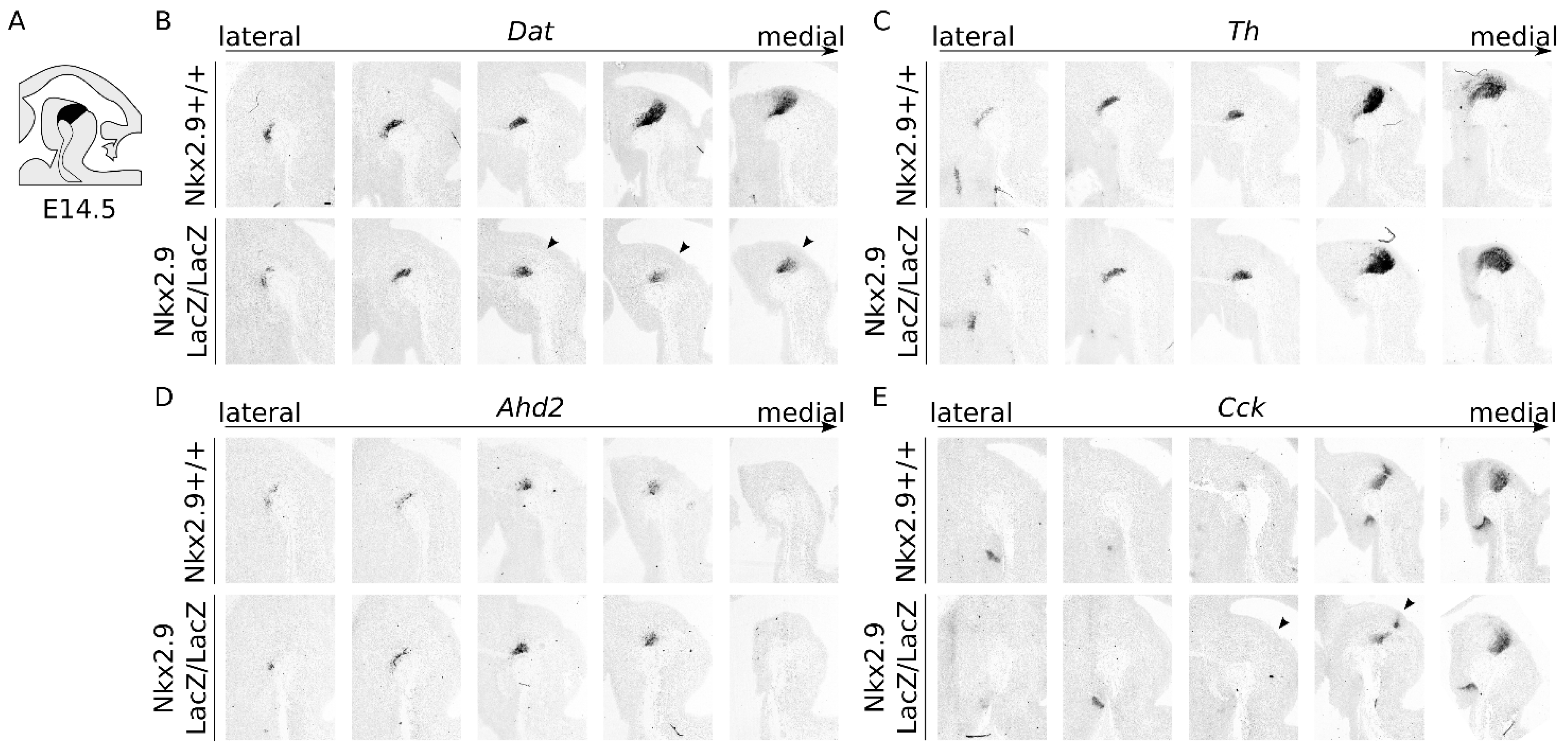
2.4. Nkx2.9-Ablation Does Not Affect the mdDA Precursor Proliferation, but Decreases Dat Expression at E14.5
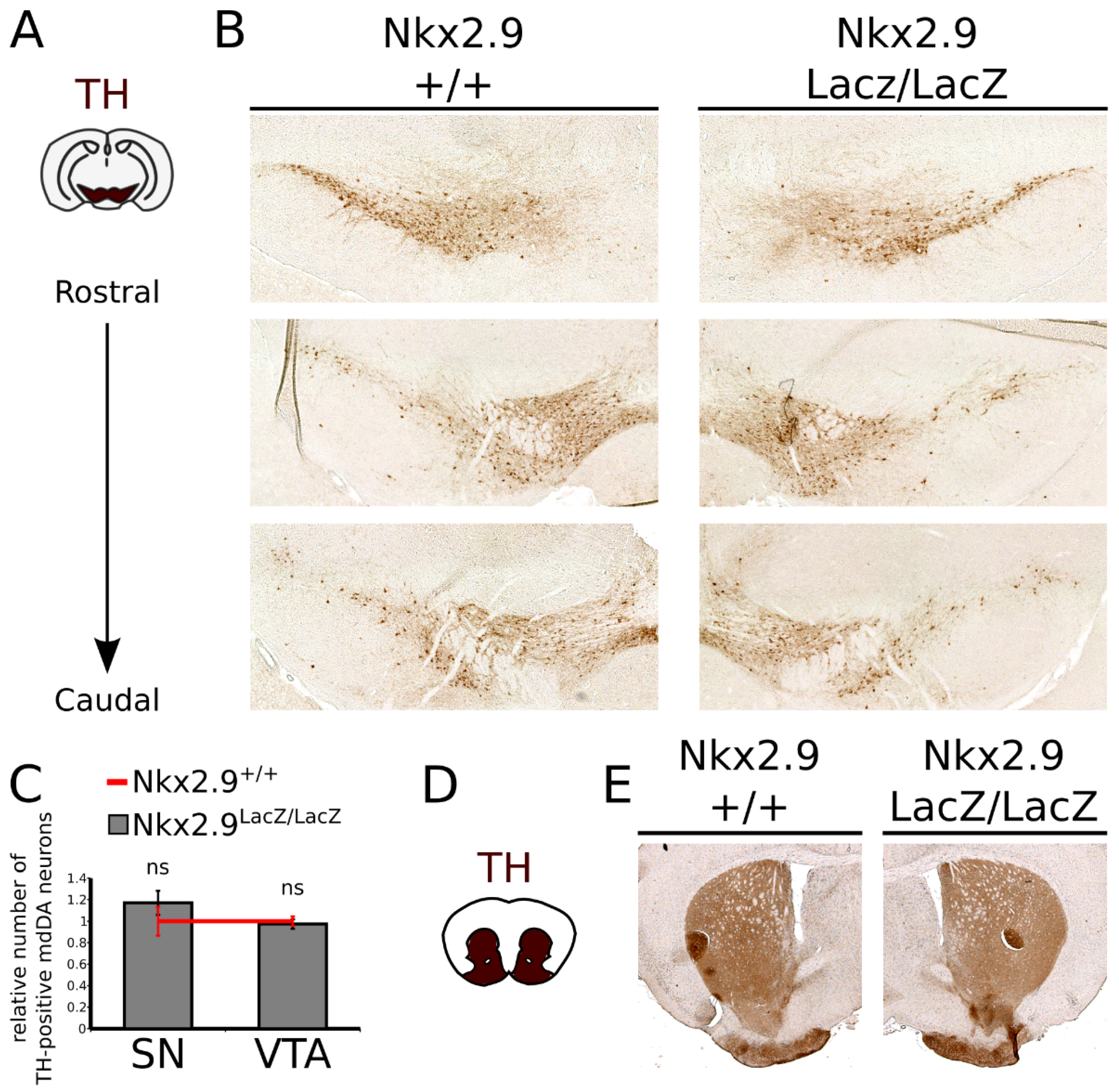
2.5. Loss of Nkx2.9 Does Not Affect the mdDA Neuronal Population in the Adult Midbrain
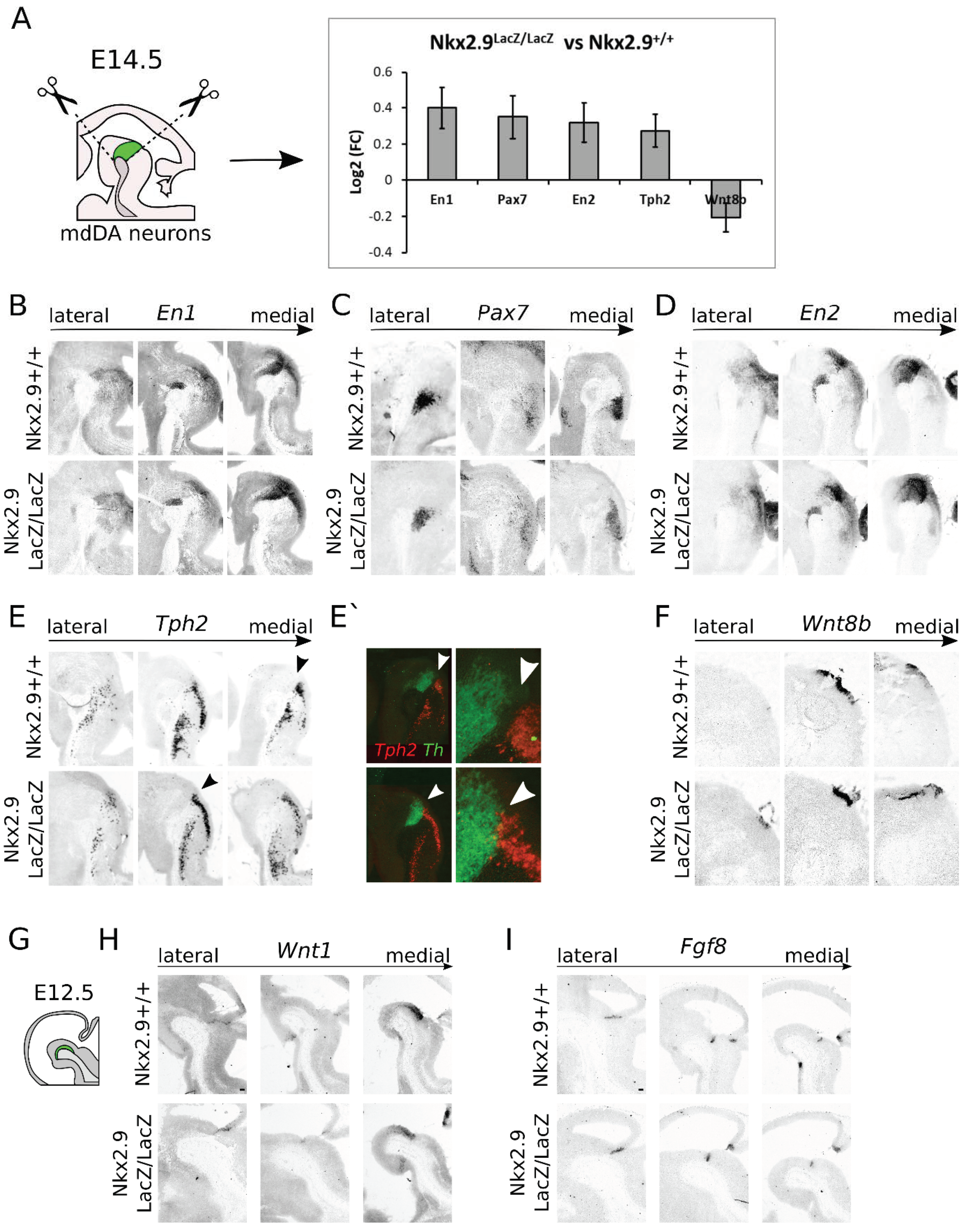
2.6. Loss of Nkx2.9 Results in a Minor Loss of Specification Factors in the Mid- and Hindbrain, Suggesting a Role in Mid- and Hindbrain Fate Determination
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. qPCR
4.3. RNA-seq
4.4. In Situ Hybridization (ISH)
4.5. Histology and Fluorescent Immunohistochemistry
4.6. DAB Immunohistochemistry
4.7. X-galactosidase Staining Protocol
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, Y.; Nirenberg, M. Drosophila NK-Homeobox Genes. Proc. Natl. Acad. Sci. USA 1989, 86, 7716–7720. [Google Scholar] [CrossRef] [Green Version]
- Apergis, G.A.; Crawford, N.; Ghosh, D.; Steppan, C.M.; Vorachek, W.R.; Wen, P.; Locker, J. A Novel Nk-2-Related Transcription Factor Associated with Human Fetal Liver and Hepatocellular Carcinoma. J. Biol. Chem. 1998, 273, 2917–2925. [Google Scholar] [CrossRef] [Green Version]
- Harvey, R.P. NK-2 Homeobox Genes and Heart Development. Dev. Biol. 1996, 178, 203–216. [Google Scholar] [CrossRef] [Green Version]
- Pabst, O.; Herbrand, H.; Arnold, H.H. Nkx2-9 Is a Novel Homeobox Transcription Factor Which Demarcates Ventral Domains in the Developing Mouse CNS. Mech. Dev. 1998, 73, 85–93. [Google Scholar] [CrossRef]
- Pabst, O.; Herbrand, H.; Takuma, N.; Arnold, H.H. NKX2 Gene Expression in Neuroectoderm but Not in Mesendodermally Derived Structures Depends on Sonic Hedgehog in Mouse Embryos. Dev. Genes Evol. 2000, 210, 47–50. [Google Scholar] [CrossRef]
- Sussel, L.; Marin, O.; Kimura, S.; Rubenstein, J.L. Loss of Nkx2.1 Homeobox Gene Function Results in a Ventral to Dorsal Molecular Respecification within the Basal Telencephalon: Evidence for a Transformation of the Pallidum into the Striatum. Development 1999, 126, 3359–3370. [Google Scholar] [CrossRef] [PubMed]
- Briscoe, J.; Sussel, L.; Serup, P.; Hartigan-O’Connor, D.; Jessell, T.M.; Rubenstein, J.L.; Ericson, J. Homeobox Gene Nkx2.2 and Specification of Neuronal Identity by Graded Sonic Hedgehog Signalling. Nature 1999, 398, 622–627. [Google Scholar] [CrossRef]
- Epstein, D.J. Regulation of Thalamic Development by Sonic Hedgehog. Front Neurosci. 2012, 6, 57. [Google Scholar] [CrossRef] [Green Version]
- Holz, A.; Kollmus, H.; Ryge, J.; Niederkofler, V.; Dias, J.; Ericson, J.; Stoeckli, E.T.; Kiehn, O.; Arnold, H.-H. The Transcription Factors Nkx2.2 and Nkx2.9 Play a Novel Role in Floor Plate Development and Commissural Axon Guidance. Development 2010, 137, 4249–4260. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, K.; Miura, H.; Yanazawa, M.; Miyashita, T.; Kato, K. Expression Patterns of Brx1 (Rieg Gene), Sonic Hedgehog, Nkx2.2, Dlx1 and Arx during Zona Limitans Intrathalamica and Embryonic Ventral Lateral Geniculate Nuclear Formation. Mech. Dev. 1997, 67, 83–96. [Google Scholar] [CrossRef]
- Pabst, O.; Rummelies, J.; Winter, B.; Arnold, H.-H. Targeted Disruption of the Homeobox Gene Nkx2.9 Reveals a Role in Development of the Spinal Accessory Nerve. Development 2003, 130, 1193–1202. [Google Scholar] [CrossRef] [Green Version]
- Bye, C.R.; Thompson, L.H.; Parish, C.L. Birth Dating of Midbrain Dopamine Neurons Identifies A9 Enriched Tissue for Transplantation into Parkinsonian Mice. Exp. Neurol. 2012, 236, 58–68. [Google Scholar] [CrossRef]
- Hegarty, S.V.; Sullivan, A.M.; O’Keeffe, G.W. Midbrain Dopaminergic Neurons: A Review of the Molecular Circuitry That Regulates Their Development. Dev. Biol. 2013, 379, 123–138. [Google Scholar] [CrossRef] [Green Version]
- Mesman, S.; Smidt, M.P. Acquisition of the Midbrain Dopaminergic Neuronal Identity. Int. J. Mol. Sci. 2020, 21, 4638. [Google Scholar] [CrossRef] [PubMed]
- Veenvliet, J.V.; Smidt, M.P. Molecular Mechanisms of Dopaminergic Subset Specification: Fundamental Aspects and Clinical Perspectives. Cell. Mol. Life Sci. 2014, 71, 4703–4727. [Google Scholar] [CrossRef] [PubMed]
- Veenvliet, J.V.; Alves Dos Santos, M.T.M.; Kouwenhoven, W.M.; von Oerthel, L.; Lim, J.L.; van der Linden, A.J.A.; Groot Koerkamp, M.J.A.; Holstege, F.C.P.; Smidt, M.P. Specification of Dopaminergic Subsets Involves Interplay of En1 and Pitx3. Development 2013, 140, 3373–3384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sgadò, P.; Albéri, L.; Gherbassi, D.; Galasso, S.L.; Ramakers, G.M.J.; Alavian, K.N.; Smidt, M.P.; Dyck, R.H.; Simon, H.H. Slow Progressive Degeneration of Nigral Dopaminergic Neurons in Postnatal Engrailed Mutant Mice. Proc. Natl. Acad. Sci. USA 2006, 103, 15242–15247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sonnier, L.; Le Pen, G.; Hartmann, A.; Bizot, J.-C.; Trovero, F.; Krebs, M.-O.; Prochiantz, A. Progressive Loss of Dopaminergic Neurons in the Ventral Midbrain of Adult Mice Heterozygote for Engrailed1. J. Neurosci. 2007, 27, 1063–1071. [Google Scholar] [CrossRef]
- Kouwenhoven, W.M.; Veenvliet, J.V.; van Hooft, J.A.; van der Heide, L.P.; Smidt, M.P. Engrailed 1 Shapes the Dopaminergic and Serotonergic Landscape through Proper Isthmic Organizer Maintenance and Function. Biol. Open 2016, 5, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Ang, S.-L. Transcriptional Control of Midbrain Dopaminergic Neuron Development. Development 2006, 133, 3499–3506. [Google Scholar] [CrossRef] [Green Version]
- Smidt, M.P.; Burbach, J.P.H. How to Make a Mesodiencephalic Dopaminergic Neuron. Nat. Rev. Neurosci. 2007, 8, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Mahmood, R.; Hnasko, R.; Locker, J. Loss of Nkx2.8 Deregulates Progenitor Cells in the Large Airways and Leads to Dysplasia. Cancer Res. 2006, 66, 10399–10407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McManus, K.J.; Hendzel, M.J. The Relationship between Histone H3 Phosphorylation and Acetylation throughout the Mammalian Cell Cycle. Biochem. Cell Biol. 2006, 84, 640–657. [Google Scholar] [CrossRef]
- Smits, S.M.; von Oerthel, L.; Hoekstra, E.J.; Burbach, J.P.H.; Smidt, M.P. Molecular Marker Differences Relate to Developmental Position and Subsets of Mesodiencephalic Dopaminergic Neurons. PLoS ONE 2013, 8, e76037. [Google Scholar] [CrossRef] [Green Version]
- Russek-Blum, N.; Gutnick, A.; Nabel-Rosen, H.; Blechman, J.; Staudt, N.; Dorsky, R.I.; Houart, C.; Levkowitz, G. Dopaminergic Neuronal Cluster Size Is Determined during Early Forebrain Patterning. Development 2008, 135, 3401–3413. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-H.; Shin, J.; Park, H.-C.; Yeo, S.-Y.; Hong, S.-K.; Han, S.; Rhee, M.; Kim, C.-H.; Chitnis, A.B.; Huh, T.-L. Specification of an Anterior Neuroectoderm Patterning by Frizzled8a-Mediated Wnt8b Signalling during Late Gastrulation in Zebrafish. Development 2002, 129, 4443–4455. [Google Scholar] [CrossRef]
- Shima, T.; Znosko, W.; Tsang, M. The Characterization of a Zebrafish Mid-Hindbrain Mutant, Mid-Hindbrain Gone (Mgo). Dev. Dyn. 2009, 238, 899–907. [Google Scholar] [CrossRef]
- Lieb, M.W.; Weidner, M.; Arnold, M.R.; Loupy, K.M.; Nguyen, K.T.; Hassell, J.E.; Schnabel, K.S.; Kern, R.; Day, H.E.W.; Lesch, K.-P.; et al. Effects of Maternal Separation on Serotonergic Systems in the Dorsal and Median Raphe Nuclei of Adult Male Tph2-Deficient Mice. Behav. Brain Res. 2019, 373, 112086. [Google Scholar] [CrossRef]
- Mlinar, B.; Montalbano, A.; Waider, J.; Lesch, K.-P.; Corradetti, R. Increased Functional Coupling of 5-HT1A Autoreceptors to GIRK Channels in Tph2−/− Mice. Eur. Neuropsychopharmacol. 2017, 27, 1258–1267. [Google Scholar] [CrossRef]
- Plummer, N.W.; Scappini, E.L.; Smith, K.G.; Tucker, C.J.; Jensen, P. Two Subpopulations of Noradrenergic Neurons in the Locus Coeruleus Complex Distinguished by Expression of the Dorsal Neural Tube Marker Pax7. Front. Neuroanat. 2017, 11, 60. [Google Scholar] [CrossRef] [Green Version]
- Jarrar, W.; Vauti, F.; Arnold, H.-H.; Holz, A. Generation of a Nkx2.2(Cre) Knock-in Mouse Line: Analysis of Cell Lineages in the Central Nervous System. Differentiation 2015, 89, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Nouri, P.; Götz, S.; Rauser, B.; Irmler, M.; Peng, C.; Trümbach, D.; Kempny, C.; Lechermeier, C.G.; Bryniok, A.; Dlugos, A.; et al. Dose-Dependent and Subset-Specific Regulation of Midbrain Dopaminergic Neuron Differentiation by LEF1-Mediated WNT1/b-Catenin Signaling. Front. Cell Dev. Biol. 2020, 8, 587778. [Google Scholar] [CrossRef] [PubMed]
- Wilkinson, D.G.; Bailes, J.A.; McMahon, A.P. Expression of the Proto-Oncogene Int-1 Is Restricted to Specific Neural Cells in the Developing Mouse Embryo. Cell 1987, 50, 79–88. [Google Scholar] [CrossRef]
- Wurst, W.; Prakash, N. Wnt1-Regulated Genetic Networks in Midbrain Dopaminergic Neuron Development. J. Mol. Cell Biol. 2013, 6, 34–41. [Google Scholar] [CrossRef] [Green Version]
- Crossley, P.H.; Martin, G.R. The Mouse Fgf8 Gene Encodes a Family of Polypeptides and Is Expressed in Regions That Direct Outgrowth and Patterning in the Developing Embryo. Development 1995, 121, 439–451. [Google Scholar] [CrossRef]
- Martinez, S.; Crossley, P.H.; Cobos, I.; Rubenstein, J.L.; Martin, G.R. FGF8 Induces Formation of an Ectopic Isthmic Organizer and Isthmocerebellar Development via a Repressive Effect on Otx2 Expression. Development 1999, 126, 1189–1200. [Google Scholar] [CrossRef]
- La Manno, G.; Gyllborg, D.; Codeluppi, S.; Nishimura, K.; Salto, C.; Zeisel, A.; Borm, L.E.; Stott, S.R.W.; Toledo, E.M.; Villaescusa, J.C.; et al. Molecular Diversity of Midbrain Development in Mouse, Human, and Stem Cells. Cell 2016, 167, 566–580.e19. [Google Scholar] [CrossRef] [Green Version]
- Poulin, J.-F.; Zou, J.; Drouin-Ouellet, J.; Kim, K.-Y.A.; Cicchetti, F.; Awatramani, R.B. Defining Midbrain Dopaminergic Neuron Diversity by Single-Cell Gene Expression Profiling. Cell Rep. 2014, 9, 930–943. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, F.M.J.; van Erp, S.; van der Linden, A.J.A.; von Oerthel, L.; Burbach, J.P.H.; Smidt, M.P. Pitx3 Potentiates Nurr1 in Dopamine Neuron Terminal Differentiation through Release of SMRT-Mediated Repression. Development 2009, 136, 531–540. [Google Scholar] [CrossRef] [Green Version]
- Di Salvio, M.; Di Giovannantonio, L.G.; Acampora, D.; Prosperi, R.; Omodei, D.; Prakash, N.; Wurst, W.; Simeone, A. Otx2 Controls Neuron Subtype Identity in Ventral Tegmental Area and Antagonizes Vulnerability to MPTP. Nat. Neurosci. 2010, 13, 1481–1488. [Google Scholar] [CrossRef] [Green Version]
- Kele, J.; Simplicio, N.; Ferri, A.L.M.; Mira, H.; Guillemot, F.; Arenas, E.; Ang, S.-L. Neurogenin 2 Is Required for the Development of Ventral Midbrain Dopaminergic Neurons. Development 2006, 133, 495–505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, D.S.; Acharya, C.R.; Balakumaran, B.S.; Riedel, R.F.; Kim, M.K.; Stevenson, M.; Tuchman, S.; Mukherjee, S.; Barry, W.; Dressman, H.K.; et al. Characterizing the Developmental Pathways TTF-1, NKX2-8, and PAX9 in Lung Cancer. Proc. Natl. Acad. Sci. USA 2009, 106, 5312–5317. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bilovocky, N.A.; Romito-DiGiacomo, R.R.; Murcia, C.L.; Maricich, S.M.; Herrup, K. Factors in the Genetic Background Suppress the Engrailed-1 Cerebellar Phenotype. J. Neurosci. 2003, 23, 5105–5112. [Google Scholar] [CrossRef] [PubMed]
- Nishikawa, T.; Bell, A.; Brooks, J.M.; Setoyama, K.; Melis, M.; Han, B.; Fukumitsu, K.; Handa, K.; Tian, J.; Kaestner, K.H.; et al. Resetting the Transcription Factor Network Reverses Terminal Chronic Hepatic Failure. J. Clin. Investig. 2015, 125, 1533–1544. [Google Scholar] [CrossRef] [Green Version]
- Afgan, E.; Baker, D.; Batut, B.; van den Beek, M.; Bouvier, D.; Čech, M.; Chilton, J.; Clements, D.; Coraor, N.; Grüning, B.A.; et al. The Galaxy Platform for Accessible, Reproducible and Collaborative Biomedical Analyses: 2018 Update. Nucleic Acids Res. 2018, 46, W537–W544. [Google Scholar] [CrossRef] [Green Version]
- Smits, S.M.; Ponnio, T.; Conneely, O.M.; Burbach, J.P.H.; Smidt, M.P. Involvement of Nurr1 in Specifying the Neurotransmitter Identity of Ventral Midbrain Dopaminergic Neurons. Eur. J. Neurosci. 2003, 18, 1731–1738. [Google Scholar] [CrossRef]
- Grima, B.; Lamouroux, A.; Blanot, F.; Biguet, N.F.; Mallet, J. Complete Coding Sequence of Rat Tyrosine Hydroxylase MRNA. Proc. Natl. Acad. Sci. USA 1985, 82, 617–621. [Google Scholar] [CrossRef] [Green Version]
- Jacobs, F.M.J.; Smits, S.M.; Noorlander, C.W.; von Oerthel, L.; van der Linden, A.J.A.; Burbach, J.P.H.; Smidt, M.P. Retinoic Acid Counteracts Developmental Defects in the Substantia Nigra Caused by Pitx3 Deficiency. Development 2007, 134, 2673–2684. [Google Scholar] [CrossRef] [Green Version]
- Mesman, S.; von Oerthel, L.; Smidt, M.P. Mesodiencephalic Dopaminergic Neuronal Differentiation Does Not Involve GLI2A-Mediated SHH-Signaling and Is under the Direct Influence of Canonical WNT Signaling. PLoS ONE 2014, 9, e97926. [Google Scholar] [CrossRef] [Green Version]
- Smidt, M.P.; Asbreuk, C.H.; Cox, J.J.; Chen, H.; Johnson, R.L.; Burbach, J.P. A Second Independent Pathway for Development of Mesencephalic Dopaminergic Neurons Requires Lmx1b. Nat. Neurosci. 2000, 3, 337–341. [Google Scholar] [CrossRef]
- Smidt, M.P.; Cox, J.J.; van Schaick, H.S.; Coolen, M.; Schepers, J.; van der Kleij, A.M.; Burbach, J.P. Analysis of Three Ptx2 Splice Variants on Transcriptional Activity and Differential Expression Pattern in the Brain. J. Neurochem. 2000, 75, 1818–1825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smidt, M.P.; Smits, S.M.; Bouwmeester, H.; Hamers, F.P.T.; van der Linden, A.J.A.; Hellemons, A.J.C.G.M.; Graw, J.; Burbach, J.P.H. Early Developmental Failure of Substantia Nigra Dopamine Neurons in Mice Lacking the Homeodomain Gene Pitx3. Development 2004, 131, 1145–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kouwenhoven, W.M.; von Oerthel, L.; Gruppilo, M.; Tian, J.; Wagemans, C.M.R.J.; Houwers, I.G.J.; Locker, J.; Mesman, S.; Smidt, M.P. Nkx2.9 Contributes to Mid-Hindbrain Patterning by Regulation of mdDA Neuronal Cell-Fate and Repression of a Hindbrain-Specific Cell-Fate. Int. J. Mol. Sci. 2021, 22, 12663. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222312663
Kouwenhoven WM, von Oerthel L, Gruppilo M, Tian J, Wagemans CMRJ, Houwers IGJ, Locker J, Mesman S, Smidt MP. Nkx2.9 Contributes to Mid-Hindbrain Patterning by Regulation of mdDA Neuronal Cell-Fate and Repression of a Hindbrain-Specific Cell-Fate. International Journal of Molecular Sciences. 2021; 22(23):12663. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222312663
Chicago/Turabian StyleKouwenhoven, Willemieke M., Lars von Oerthel, Maria Gruppilo, Jianmin Tian, Cindy M. R. J. Wagemans, Imke G. J. Houwers, Joseph Locker, Simone Mesman, and Marten P. Smidt. 2021. "Nkx2.9 Contributes to Mid-Hindbrain Patterning by Regulation of mdDA Neuronal Cell-Fate and Repression of a Hindbrain-Specific Cell-Fate" International Journal of Molecular Sciences 22, no. 23: 12663. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222312663