
Roles of Transcription Factors in the Development and Reprogramming of the Dopaminergic Neurons


Abstract
:1. Introduction
2. TFs in Development of mdDA Neurons
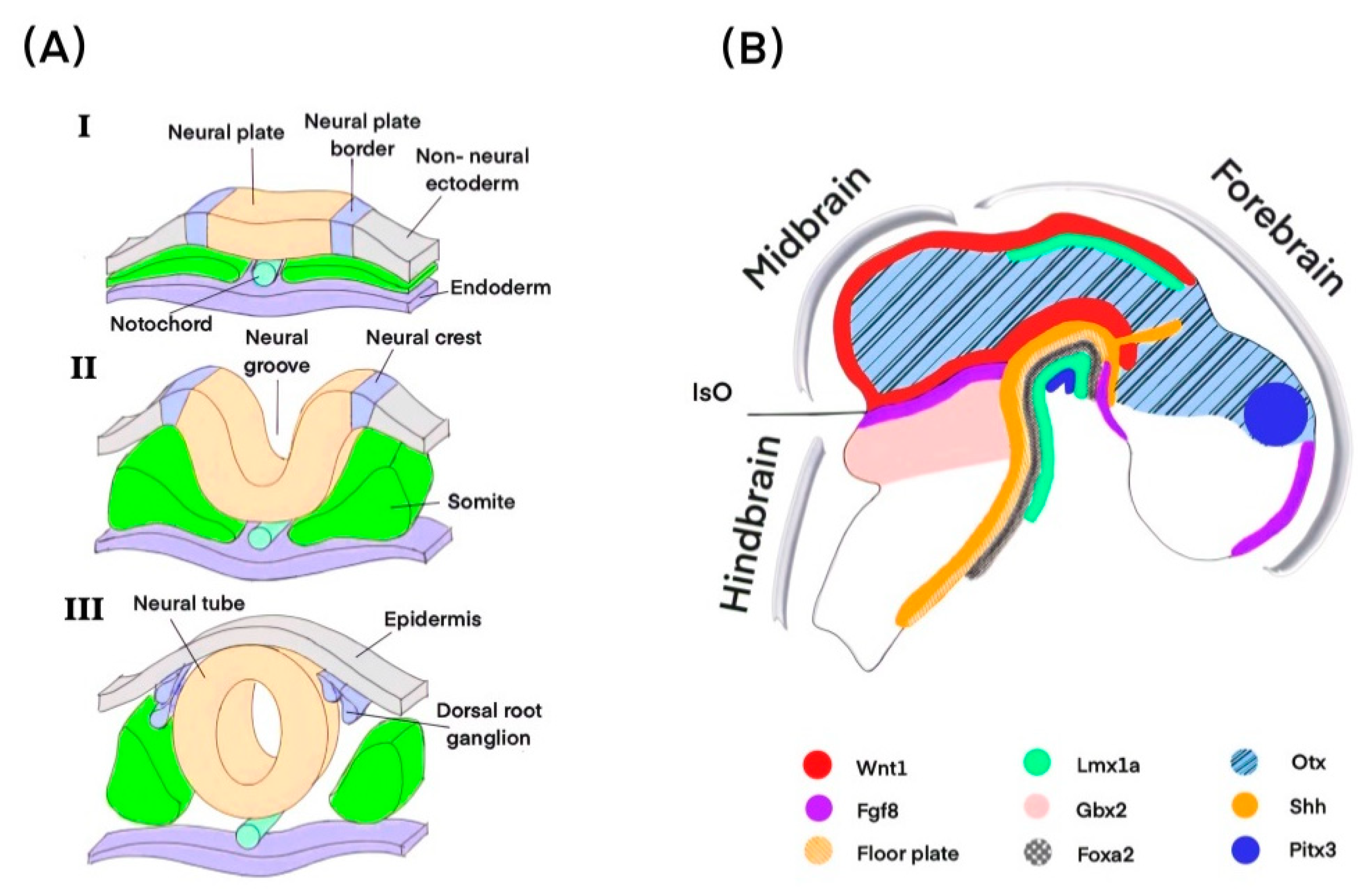
2.1. Induction of Midbrain FP
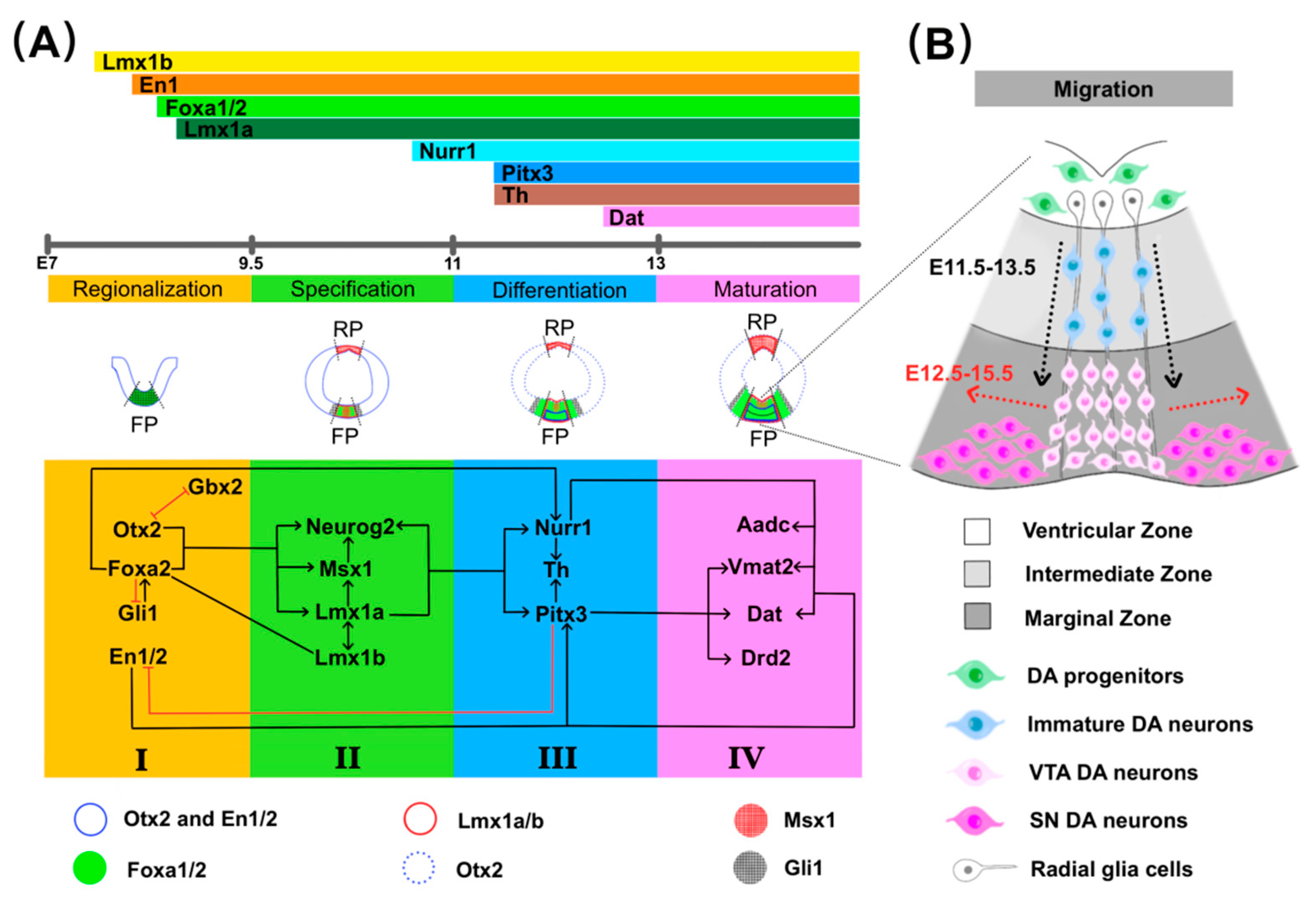
2.2. Specification of mdDA Progenitors
2.3. Differentiation and Maturation
2.4. Migration
3. TFs in Dopamine Cell Therapy
3.1. Dopamine Cell Therapy
3.2. Nurr1
3.3. Pitx3
3.4. Lmx1a/b
3.5. En1
3.6. Foxa1/2
3.7. Combinations of TFs
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Poewe, W.; Seppi, K.; Tanner, C.M.; Halliday, G.M.; Brundin, P.; Volkmann, J.; Schrag, A.-E.; Lang, A.E. Parkinson disease. Nat. Rev. Dis. Primers 2017, 3, 17013. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Mizuno, K.; Sasaki, A.T.; Wada, Y.; Tanaka, M.; Ishii, A.; Tajima, K.; Tsuyuguchi, N.; Watanabe, K.; Zeki, S. Imaging the passionate stage of romantic love by dopamine dynamics. Front. Hum. Neurosci. 2015, 9, 191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gebauer, L.; Kringelbach, M.L.; Vuust, P. Ever-changing cycles of musical pleasure: The role of dopamine and anticipation. Psychomusicol. Music Mind Brain 2012, 22, 152. [Google Scholar] [CrossRef] [Green Version]
- Pascoli, V.; Terrier, J.; Hiver, A.; Lüscher, C. Sufficiency of mesolimbic dopamine neuron stimulation for the progression to addiction. Neuron 2015, 88, 1054–1066. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, J.A.; Tecuapetla, F.; Paixão, V.; Costa, R.M. Dopamine neuron activity before action initiation gates and invigorates future movements. Nature 2018, 554, 244–248. [Google Scholar] [CrossRef]
- Schultz, W. Multiple dopamine functions at different time courses. Annu. Rev. Neurosci. 2007, 30, 259–288. [Google Scholar] [CrossRef] [Green Version]
- Zeng, X.S.; Geng, W.S.; Jia, J.J.; Chen, L.; Zhang, P.P. Cellular and Molecular Basis of Neurodegeneration in Parkinson Disease. Front. Aging Neurosci. 2018, 10, 109. [Google Scholar] [CrossRef] [Green Version]
- Schapira, A.H.V.; Chaudhuri, K.R.; Jenner, P. Non-motor features of Parkinson disease. Nat. Rev. Neurosci. 2017, 18, 435–450. [Google Scholar] [CrossRef]
- Franco, R.; Reyes-Resina, I.; Navarro, G. Dopamine in Health and Disease: Much More Than a Neurotransmitter. Biomedicines 2021, 9, 109. [Google Scholar] [CrossRef]
- Nemade, D.; Subramanian, T.; Shivkumar, V. An Update on Medical and Surgical Treatments of Parkinson’s Disease. Aging Dis. 2021, 12, 1021–1035. [Google Scholar] [CrossRef] [PubMed]
- Le, W.-D. Inauguration of a unique journal Ageing and Neurodegenerative Diseases: A new beginning seeking cures for age-related neurodegenerative diseases. Ageing Neurodegener. Dis. 2021, 1, 1. [Google Scholar] [CrossRef]
- Carmichael, K.; Sullivan, B.; Lopez, E.; Sun, L.; Cai, H. Diverse midbrain dopaminergic neuron subtypes and implications for complex clinical symptoms of Parkinson’s disease. Ageing Neurodegener. Dis. 2021, 1, 4. [Google Scholar] [CrossRef] [PubMed]
- Barbuti, P.A.; Barker, R.A.; Brundin, P.; Przedborski, S.; Papa, S.M.; Kalia, L.V.; Mochizuki, H. Recent Advances in the Development of Stem-Cell-Derived Dopaminergic Neuronal Transplant Therapies for Parkinson’s Disease. Mov. Disord. 2021, 36, 1772–1780. [Google Scholar] [CrossRef]
- Zakrzewski, W.; Dobrzyński, M.; Szymonowicz, M.; Rybak, Z. Stem cells: Past, present, and future. Stem Cell Res. Ther. 2019, 10, 68. [Google Scholar] [CrossRef] [PubMed]
- Azimi, S.M.; Sheridan, S.D.; Ghannad-Rezaie, M.; Eimon, P.M.; Yanik, M.F. Combinatorial programming of human neuronal progenitors using magnetically-guided stoichiometric mRNA delivery. eLife 2018, 7. [Google Scholar] [CrossRef] [PubMed]
- Rivetti di Val Cervo, P.; Romanov, R.A.; Spigolon, G.; Masini, D.; Martín-Montañez, E.; Toledo, E.M.; La Manno, G.; Feyder, M.; Pifl, C.; Ng, Y.H.; et al. Induction of functional dopamine neurons from human astrocytes in vitro and mouse astrocytes in a Parkinson’s disease model. Nat. Biotechnol. 2017, 35, 444–452. [Google Scholar] [CrossRef]
- Ng, Y.H.; Chanda, S.; Janas, J.A.; Yang, N.; Kokubu, Y.; Südhof, T.C.; Wernig, M. Efficient generation of dopaminergic induced neuronal cells with midbrain characteristics. Stem Cell Rep. 2021, 16, 1763–1776. [Google Scholar] [CrossRef] [PubMed]
- Lambert, S.A.; Jolma, A.; Campitelli, L.F.; Das, P.K.; Yin, Y.; Albu, M.; Chen, X.; Taipale, J.; Hughes, T.R.; Weirauch, M.T. The Human Transcription Factors. Cell 2018, 172, 650–665. [Google Scholar] [CrossRef] [Green Version]
- Canals, I.; Quist, E.; Ahlenius, H. Transcription Factor-Based Strategies to Generate Neural Cell Types from Human Pluripotent Stem Cells. Cell Reprogram 2021, 23, 206–220. [Google Scholar] [CrossRef]
- Mesman, S.; Smidt, M.P. Acquisition of the Midbrain Dopaminergic Neuronal Identity. Int. J. Mol. Sci. 2020, 21, 4638. [Google Scholar] [CrossRef]
- Rodríguez-Traver, E.; Solís, O.; Díaz-Guerra, E.; Ortiz, Ó.; Vergaño-Vera, E.; Méndez-Gómez, H.R.; García-Sanz, P.; Moratalla, R.; Vicario-Abejón, C. Role of Nurr1 in the Generation and Differentiation of Dopaminergic Neurons from Stem Cells. Neurotox Res. 2016, 30, 14–31. [Google Scholar] [CrossRef] [Green Version]
- Romanos, M.; Allio, G.; Roussigné, M.; Combres, L.; Escalas, N.; Soula, C.; Médevielle, F.; Steventon, B.; Trescases, A.; Bénazéraf, B. Cell-to-cell heterogeneity in Sox2 and Bra expression guides progenitor motility and destiny. eLife 2021, 10. [Google Scholar] [CrossRef] [PubMed]
- Kadoya, M.; Sasai, N. Negative Regulation of mTOR Signaling Restricts Cell Proliferation in the Floor Plate. Front. Neurosci. 2019, 13, 1022. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Ling, K.H.; Tan, J.J.; Lu, C.B. Development and Differentiation of Midbrain Dopaminergic Neuron: From Bench to Bedside. Cells 2020, 9, 1489. [Google Scholar] [CrossRef]
- Bond, A.M.; Bhalala, O.G.; Kessler, J.A. The dynamic role of bone morphogenetic proteins in neural stem cell fate and maturation. Dev. Neurobiol. 2012, 72, 1068–1084. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arenas, E. Foxa2: The rise and fall of dopamine neurons. Cell Stem Cell 2008, 2, 110–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, Y.; Zhang, S.C. Neural Subtype Specification from Human Pluripotent Stem Cells. Cell Stem Cell 2016, 19, 573–586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maury, Y.; Côme, J.; Piskorowski, R.A.; Salah-Mohellibi, N.; Chevaleyre, V.; Peschanski, M.; Martinat, C.; Nedelec, S. Combinatorial analysis of developmental cues efficiently converts human pluripotent stem cells into multiple neuronal subtypes. Nat. Biotechnol. 2015, 33, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Allodi, I.; Hedlund, E. Directed midbrain and spinal cord neurogenesis from pluripotent stem cells to model development and disease in a dish. Front. Neurosci. 2014, 8, 109. [Google Scholar] [CrossRef] [Green Version]
- Fasano, C.A.; Chambers, S.M.; Lee, G.; Tomishima, M.J.; Studer, L. Efficient derivation of functional floor plate tissue from human embryonic stem cells. Cell Stem Cell 2010, 6, 336–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joyner, A.L.; Liu, A.; Millet, S. Otx2, Gbx2 and Fgf8 interact to position and maintain a mid-hindbrain organizer. Curr. Opin. Cell Biol. 2000, 12, 736–741. [Google Scholar] [CrossRef]
- Puelles, E.; Annino, A.; Tuorto, F.; Usiello, A.; Acampora, D.; Czerny, T.; Brodski, C.; Ang, S.L.; Wurst, W.; Simeone, A. Otx2 regulates the extent, identity and fate of neuronal progenitor domains in the ventral midbrain. Development 2004, 131, 2037–2048. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Lee, J.S.; Hwang, H.S.; Lee, D.R.; Park, C.Y.; Jung, S.J.; You, Y.R.; Kim, D.S.; Kim, D.W. Wnt signal activation induces midbrain specification through direct binding of the beta-catenin/TCF4 complex to the EN1 promoter in human pluripotent stem cells. Exp. Mol. Med. 2018, 50, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.; Brown, A.; Ellisor, D.; Paul, E.; Hagan, N.; Zervas, M. Dynamic temporal requirement of Wnt1 in midbrain dopamine neuron development. Development 2013, 140, 1342–1352. [Google Scholar] [CrossRef] [Green Version]
- Ásgrímsdóttir, E.S.; Arenas, E. Midbrain Dopaminergic Neuron Development at the Single Cell Level: In vivo and in Stem Cells. Front. Cell Dev. Biol. 2020, 8, 463. [Google Scholar] [CrossRef]
- Gaggi, G.; Di Credico, A.; Izzicupo, P.; Iannetti, G.; Di Baldassarre, A.; Ghinassi, B. Chemical and Biological Molecules Involved in Differentiation, Maturation, and Survival of Dopaminergic Neurons in Health and Parkinson’s Disease: Physiological Aspects and Clinical Implications. Biomedicines 2021, 9, 754. [Google Scholar] [CrossRef] [PubMed]
- Blaess, S.; Ang, S.L. Genetic control of midbrain dopaminergic neuron development. Wiley Interdiscip. Rev. Dev. Biol. 2015, 4, 113–134. [Google Scholar] [CrossRef] [PubMed]
- Nakatani, T.; Kumai, M.; Mizuhara, E.; Minaki, Y.; Ono, Y. Lmx1a and Lmx1b cooperate with Foxa2 to coordinate the specification of dopaminergic neurons and control of floor plate cell differentiation in the developing mesencephalon. Dev. Biol. 2010, 339, 101–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, C.H.; Levesque, M.; Claxton, S.; Johnson, R.L.; Ang, S.L. Lmx1a and lmx1b function cooperatively to regulate proliferation, specification, and differentiation of midbrain dopaminergic progenitors. J. Neurosci. 2011, 31, 12413–12425. [Google Scholar] [CrossRef] [Green Version]
- Deng, Q.; Andersson, E.; Hedlund, E.; Alekseenko, Z.; Coppola, E.; Panman, L.; Millonig, J.H.; Brunet, J.F.; Ericson, J.; Perlmann, T. Specific and integrated roles of Lmx1a, Lmx1b and Phox2a in ventral midbrain development. Development 2011, 138, 3399–3408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veenvliet, J.V.; Dos Santos, M.T.; Kouwenhoven, W.M.; von Oerthel, L.; Lim, J.L.; van der Linden, A.J.; Koerkamp, M.J.; Holstege, F.C.; Smidt, M.P. Specification of dopaminergic subsets involves interplay of En1 and Pitx3. Development 2013, 140, 3373–3384. [Google Scholar] [CrossRef] [Green Version]
- Lin, W.; Metzakopian, E.; Mavromatakis, Y.E.; Gao, N.; Balaskas, N.; Sasaki, H.; Briscoe, J.; Whitsett, J.A.; Goulding, M.; Kaestner, K.H.; et al. Foxa1 and Foxa2 function both upstream of and cooperatively with Lmx1a and Lmx1b in a feedforward loop promoting mesodiencephalic dopaminergic neuron development. Dev. Biol. 2009, 333, 386–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metzakopian, E.; Bouhali, K.; Alvarez-Saavedra, M.; Whitsett, J.A.; Picketts, D.J.; Ang, S.L. Genome-wide characterisation of Foxa1 binding sites reveals several mechanisms for regulating neuronal differentiation in midbrain dopamine cells. Development 2015, 142, 1315–1324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simeone, A.; Puelles, E.; Omodei, D.; Acampora, D.; Di Giovannantonio, L.G.; Di Salvio, M.; Mancuso, P.; Tomasetti, C. Otx genes in neurogenesis of mesencephalic dopaminergic neurons. Dev. Neurobiol. 2011, 71, 665–679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omodei, D.; Acampora, D.; Mancuso, P.; Prakash, N.; Di Giovannantonio, L.G.; Wurst, W.; Simeone, A. Anterior-posterior graded response to Otx2 controls proliferation and differentiation of dopaminergic progenitors in the ventral mesencephalon. Development 2008, 135, 3459–3470. [Google Scholar] [CrossRef] [Green Version]
- Zetterström, R.H.; Solomin, L.; Jansson, L.; Hoffer, B.J.; Olson, L.; Perlmann, T. Dopamine neuron agenesis in Nurr1-deficient mice. Science 1997, 276, 248–250. [Google Scholar] [CrossRef]
- Jankovic, J.; Chen, S.; Le, W.D. The role of Nurr1 in the development of dopaminergic neurons and Parkinson’s disease. Prog. Neurobiol. 2005, 77, 128–138. [Google Scholar] [CrossRef]
- Yi, S.H.; He, X.B.; Rhee, Y.H.; Park, C.H.; Takizawa, T.; Nakashima, K.; Lee, S.H. Foxa2 acts as a co-activator potentiating expression of the Nurr1-induced DA phenotype via epigenetic regulation. Development 2014, 141, 761–772. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.M.; Chang, M.Y.; Song, J.J.; Rhee, Y.H.; Joe, E.H.; Lee, H.S.; Yi, S.H.; Lee, S.H. Combined Nurr1 and Foxa2 roles in the therapy of Parkinson’s disease. EMBO Mol. Med. 2015, 7, 510–525. [Google Scholar] [CrossRef] [Green Version]
- Smidt, M.P.; van Schaick, H.S.; Lanctôt, C.; Tremblay, J.J.; Cox, J.J.; van der Kleij, A.A.; Wolterink, G.; Drouin, J.; Burbach, J.P. A homeodomain gene Ptx3 has highly restricted brain expression in mesencephalic dopaminergic neurons. Proc. Natl. Acad. Sci. USA 1997, 94, 13305–13310. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Chen, X.; Wang, Y.; Li, S.; Cai, H.; Le, W. The essential role of transcription factor Pitx3 in preventing mesodiencephalic dopaminergic neurodegeneration and maintaining neuronal subtype identities during aging. Cell Death Dis. 2021, 12, 1008. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, S.L.; Ho, H.Y.; Kuehner, E.; Zhao, S.; Li, M. Pitx3 regulates tyrosine hydroxylase expression in the substantia nigra and identifies a subgroup of mesencephalic dopaminergic progenitor neurons during mouse development. Dev. Biol. 2005, 282, 467–479. [Google Scholar] [CrossRef] [Green Version]
- Kouwenhoven, W.M.; von Oerthel, L.; Smidt, M.P. Pitx3 and En1 determine the size and molecular programming of the dopaminergic neuronal pool. PLoS ONE 2017, 12, e0182421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albéri, L.; Sgadò, P.; Simon, H.H. Engrailed genes are cell-autonomously required to prevent apoptosis in mesencephalic dopaminergic neurons. Development 2004, 131, 3229–3236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blaudin de Thé, F.X.; Rekaik, H.; Prochiantz, A.; Fuchs, J.; Joshi, R.L. Neuroprotective Transcription Factors in Animal Models of Parkinson Disease. Neural Plast. 2016, 2016, 6097107. [Google Scholar] [CrossRef] [Green Version]
- Bodea, G.O.; Spille, J.H.; Abe, P.; Andersson, A.S.; Acker-Palmer, A.; Stumm, R.; Kubitscheck, U.; Blaess, S. Reelin and CXCL12 regulate distinct migratory behaviors during the development of the dopaminergic system. Development 2014, 141, 661–673. [Google Scholar] [CrossRef] [Green Version]
- Brignani, S.; Raj, D.D.A.; Schmidt, E.R.E.; Düdükcü, Ö.; Adolfs, Y.; De Ruiter, A.A.; Rybiczka-Tesulov, M.; Verhagen, M.G.; van der Meer, C.; Broekhoven, M.H.; et al. Remotely Produced and Axon-Derived Netrin-1 Instructs GABAergic Neuron Migration and Dopaminergic Substantia Nigra Development. Neuron 2020, 107, 684–702. [Google Scholar] [CrossRef]
- Yang, S.; Edman, L.C.; Sánchez-Alcañiz, J.A.; Fritz, N.; Bonilla, S.; Hecht, J.; Uhlén, P.; Pleasure, S.J.; Villaescusa, J.C.; Marín, O.; et al. Cxcl12/Cxcr4 signaling controls the migration and process orientation of A9-A10 dopaminergic neurons. Development 2013, 140, 4554–4564. [Google Scholar] [CrossRef] [Green Version]
- Vaswani, A.R.; Weykopf, B.; Hagemann, C.; Fried, H.U.; Brüstle, O.; Blaess, S. Correct setup of the substantia nigra requires Reelin-mediated fast, laterally-directed migration of dopaminergic neurons. eLife 2019, 8, e41623. [Google Scholar] [CrossRef]
- Yin, M.; Liu, S.; Yin, Y.; Li, S.; Li, Z.; Wu, X.; Zhang, B.; Ang, S.L.; Ding, Y.; Zhou, J. Ventral mesencephalon-enriched genes that regulate the development of dopaminergic neurons in vivo. J. Neurosci. 2009, 29, 5170–5182. [Google Scholar] [CrossRef]
- Panman, L.; Papathanou, M.; Laguna, A.; Oosterveen, T.; Volakakis, N.; Acampora, D.; Kurtsdotter, I.; Yoshitake, T.; Kehr, J.; Joodmardi, E.; et al. Sox6 and Otx2 control the specification of substantia nigra and ventral tegmental area dopamine neurons. Cell Rep. 2014, 8, 1018–1025. [Google Scholar] [CrossRef] [Green Version]
- Simeone, A.; Di Salvio, M.; Di Giovannantonio, L.G.; Acampora, D.; Omodei, D.; Tomasetti, C. The role of otx2 in adult mesencephalic-diencephalic dopaminergic neurons. Mol. Neurobiol. 2011, 43, 107–113. [Google Scholar] [CrossRef]
- Zheng, M.; Jiao, L.; Tang, X.; Xiang, X.; Wan, X.; Yan, Y.; Li, X.; Zhang, G.; Li, Y.; Jiang, B.; et al. Tau haploinsufficiency causes prenatal loss of dopaminergic neurons in the ventral tegmental area and reduction of transcription factor orthodenticle homeobox 2 expression. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2017, 31, 3349–3358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallett, P.J.; Cooper, O.; Sadi, D.; Robertson, H.; Mendez, I.; Isacson, O. Long-term health of dopaminergic neuron transplants in Parkinson’s disease patients. Cell Rep. 2014, 7, 1755–1761. [Google Scholar] [CrossRef] [Green Version]
- Brundin, P.; Pogarell, O.; Hagell, P.; Piccini, P.; Widner, H.; Schrag, A.; Kupsch, A.; Crabb, L.; Odin, P.; Gustavii, B.; et al. Bilateral caudate and putamen grafts of embryonic mesencephalic tissue treated with lazaroids in Parkinson’s disease. Brain J. Neurol. 2000, 123, 1380–1390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brundin, P.; Karlsson, J.; Emgård, M.; Schierle, G.S.; Hansson, O.; Petersén, A.; Castilho, R.F. Improving the survival of grafted dopaminergic neurons: A review over current approaches. Cell Transplant 2000, 9, 179–195. [Google Scholar] [CrossRef]
- Haas, S.J.; Beckmann, S.; Petrov, S.; Andressen, C.; Wree, A.; Schmitt, O. Transplantation of immortalized mesencephalic progenitors (CSM14.1 cells) into the neonatal parkinsonian rat caudate putamen. J. Neurosci. Res. 2007, 85, 778–786. [Google Scholar] [CrossRef] [PubMed]
- Jin, G.; Tan, X.; Tian, M.; Qin, J.; Zhu, H.; Huang, Z.; Xu, H. The controlled differentiation of human neural stem cells into TH-immunoreactive (ir) neurons in vitro. Neurosci. Lett. 2005, 386, 105–110. [Google Scholar] [CrossRef]
- Saucedo-Cardenas, O.; Quintana-Hau, J.D.; Le, W.D.; Smidt, M.P.; Cox, J.J.; De Mayo, F.; Burbach, J.P.; Conneely, O.M. Nurr1 is essential for the induction of the dopaminergic phenotype and the survival of ventral mesencephalic late dopaminergic precursor neurons. Proc. Natl. Acad. Sci. USA 1998, 95, 4013–4018. [Google Scholar] [CrossRef] [Green Version]
- Jakaria, M.; Haque, M.E.; Cho, D.Y.; Azam, S.; Kim, I.S.; Choi, D.K. Molecular Insights into NR4A2(Nurr1): An Emerging Target for Neuroprotective Therapy Against Neuroinflammation and Neuronal Cell Death. Mol. Neurobiol. 2019, 56, 5799–5814. [Google Scholar] [CrossRef]
- Wang, Q.; Song, S.; Jiang, L.; Hong, J.-S. Interplay among norepinephrine, NOX2, and neuroinflammation: Key players in Parkinson’s disease and prime targets for therapies. Ageing Neurodegener. Dis. 2021, 1, 6. [Google Scholar] [CrossRef]
- Dong, J.; Liu, X.; Wang, Y.; Cai, H.; Le, W. Nurr1(Cd11bcre) conditional knockout mice display inflammatory injury to nigrostriatal dopaminergic neurons. Glia 2020, 68, 2057–2069. [Google Scholar] [CrossRef]
- Saijo, K.; Winner, B.; Carson, C.T.; Collier, J.G.; Boyer, L.; Rosenfeld, M.G.; Gage, F.H.; Glass, C.K. A Nurr1/CoREST pathway in microglia and astrocytes protects dopaminergic neurons from inflammation-induced death. Cell 2009, 137, 47–59. [Google Scholar] [CrossRef] [Green Version]
- Chu, Y.; Kompoliti, K.; Cochran, E.J.; Mufson, E.J.; Kordower, J.H. Age-related decreases in Nurr1 immunoreactivity in the human substantia nigra. J. Comp. Neurol. 2002, 450, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Chu, Y.; Kordower, J.H. Age-associated increases of α-synuclein in monkeys and humans are associated with nigrostriatal dopamine depletion: Is this the target for Parkinson’s disease? Neurobiol. Dis. 2007, 25, 134–149. [Google Scholar] [CrossRef] [PubMed]
- Heng, X.; Jin, G.; Zhang, X.; Yang, D.; Zhu, M.; Fu, S.; Li, X.; Le, W. Nurr1 regulates Top IIbeta and functions in axon genesis of mesencephalic dopaminergic neurons. Mol. Neurodegener. 2012, 7, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaim, M.; Isik, S. DNA topoisomerase IIβ stimulates neurite outgrowth in neural differentiated human mesenchymal stem cells through regulation of Rho-GTPases (RhoA/Rock2 pathway) and Nurr1 expression. Stem Cell Res. Ther. 2018, 9, 114. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.M.; Cho, S.Y.; Kim, M.W.; Roh, S.R.; Shin, H.S.; Suh, Y.H.; Geum, D.; Lee, M.A. Genome-Wide Analysis Identifies NURR1-Controlled Network of New Synapse Formation and Cell Cycle Arrest in Human Neural Stem Cells. Mol. Cells 2020, 43, 551–571. [Google Scholar] [CrossRef] [PubMed]
- Kadkhodaei, B.; Alvarsson, A.; Schintu, N.; Ramsköld, D.; Volakakis, N.; Joodmardi, E.; Yoshitake, T.; Kehr, J.; Decressac, M.; Björklund, A.; et al. Transcription factor Nurr1 maintains fiber integrity and nuclear-encoded mitochondrial gene expression in dopamine neurons. Proc. Natl. Acad. Sci. USA 2013, 110, 2360–2365. [Google Scholar] [CrossRef] [Green Version]
- Jodeiri Farshbaf, M.; Forouzanfar, M.; Ghaedi, K.; Kiani-Esfahani, A.; Peymani, M.; Shoaraye Nejati, A.; Izadi, T.; Karbalaie, K.; Noorbakhshnia, M.; Rahgozar, S.; et al. Nurr1 and PPARγ protect PC12 cells against MPP(+) toxicity: Involvement of selective genes, anti-inflammatory, ROS generation, and antimitochondrial impairment. Mol. Cell. Biochem. 2016, 420, 29–42. [Google Scholar] [CrossRef]
- Chung, S.; Sonntag, K.C.; Andersson, T.; Bjorklund, L.M.; Park, J.J.; Kim, D.W.; Kang, U.J.; Isacson, O.; Kim, K.S. Genetic engineering of mouse embryonic stem cells by Nurr1 enhances differentiation and maturation into dopaminergic neurons. Eur. J. Neurosci. 2002, 16, 1829–1838. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.H.; Auerbach, J.M.; Rodríguez-Gómez, J.A.; Velasco, I.; Gavin, D.; Lumelsky, N.; Lee, S.H.; Nguyen, J.; Sánchez-Pernaute, R.; Bankiewicz, K.; et al. Dopamine neurons derived from embryonic stem cells function in an animal model of Parkinson’s disease. Nature 2002, 418, 50–56. [Google Scholar] [CrossRef]
- Terraf, P.; Babaloo, H.; Kouhsari, S.M. Directed Differentiation of Dopamine-Secreting Cells from Nurr1/GPX1 Expressing Murine Embryonic Stem Cells Cultured on Matrigel-Coated PCL Scaffolds. Mol. Neurobiol. 2017, 54, 1119–1128. [Google Scholar] [CrossRef]
- Lee, E.H.; Kim, S.M.; Kim, C.H.; Pagire, S.H.; Pagire, H.S.; Chung, H.Y.; Ahn, J.H.; Park, C.H. Dopamine neuron induction and the neuroprotective effects of thyroid hormone derivatives. Sci. Rep. 2019, 9, 13659. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.Y.; Chiu, T.L.; Chang, H.F.; Hsu, H.R.; Pang, C.Y.; Liew, H.K.; Wang, M.J. Epigenetic regulation contributes to urocortin-enhanced midbrain dopaminergic neuron differentiation. Stem Cells 2015, 33, 1601–1617. [Google Scholar] [CrossRef]
- Qian, Y.; Chen, X.X.; Wang, W.; Li, J.J.; Wang, X.P.; Tang, Z.W.; Xu, J.T.; Lin, H.; Yang, Z.Y.; Li, L.Y.; et al. Transplantation of Nurr1-overexpressing neural stem cells and microglia for treating parkinsonian rats. CNS Neurosci. Ther. 2020, 26, 55–65. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.X.; Qian, Y.; Wang, X.P.; Tang, Z.W.; Xu, J.T.; Lin, H.; Yang, Z.Y.; Song, X.B.; Lu, D.; Guo, J.Z.; et al. Nurr1 promotes neurogenesis of dopaminergic neuron and represses inflammatory factors in the transwell coculture system of neural stem cells and microglia. CNS Neurosci. Ther. 2018, 24, 790–800. [Google Scholar] [CrossRef] [Green Version]
- Abumaree, M.; Al Jumah, M.; Pace, R.A.; Kalionis, B. Immunosuppressive properties of mesenchymal stem cells. Stem Cell Rev. Rep. 2012, 8, 375–392. [Google Scholar] [CrossRef]
- Wang, X.; Zhuang, W.; Fu, W.; Wang, X.; Lv, E.; Li, F.; Zhou, S.; Rausch, W.D.; Wang, X. The lentiviral-mediated Nurr1 genetic engineering mesenchymal stem cells protect dopaminergic neurons in a rat model of Parkinson’s disease. Am. J. Transl. Res. 2018, 10, 1583–1599. [Google Scholar] [PubMed]
- Vergaño-Vera, E.; Díaz-Guerra, E.; Rodríguez-Traver, E.; Méndez-Gómez, H.R.; Solís, Ó.; Pignatelli, J.; Pickel, J.; Lee, S.H.; Moratalla, R.; Vicario-Abejón, C. Nurr1 blocks the mitogenic effect of FGF-2 and EGF, inducing olfactory bulb neural stem cells to adopt dopaminergic and dopaminergic-GABAergic neuronal phenotypes. Dev. Neurobiol. 2015, 75, 823–841. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Chung, S.; Leung, K.; Hwang, I.; Moon, J.; Kim, K.S. Functional roles of Nurr1, Pitx3, and Lmx1a in neurogenesis and phenotype specification of dopamine neurons during in vitro differentiation of embryonic stem cells. Stem Cells Dev. 2014, 23, 477–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Luzy, I.R.; Niclis, J.C.; Gantner, C.W.; Kauhausen, J.A.; Hunt, C.P.J.; Ermine, C.; Pouton, C.W.; Thompson, L.H.; Parish, C.L. Isolation of LMX1a Ventral Midbrain Progenitors Improves the Safety and Predictability of Human Pluripotent Stem Cell-Derived Neural Transplants in Parkinsonian Disease. J. Neurosci. 2019, 39, 9521–9531. [Google Scholar] [CrossRef]
- Eskandarian Boroujeni, M.; Aliaghaei, A.; Maghsoudi, N.; Gardaneh, M. Complementation of dopaminergic signaling by Pitx3-GDNF synergy induces dopamine secretion by multipotent Ntera2 cells. J. Cell. Biochem. 2020, 121, 200–212. [Google Scholar] [CrossRef]
- Collazo-Navarrete, O.; Hernández-García, D.; Guerrero-Flores, G.; Drucker-Colín, R.; Guerra-Crespo, M.; Covarrubias, L. The Substantia Nigra Is Permissive and Gains Inductive Signals When Lesioned for Dopaminergic Differentiation of Embryonic Stem Cells. Stem Cells Dev. 2019, 28, 1104–1115. [Google Scholar] [CrossRef] [PubMed]
- Lim, M.S.; Chang, M.Y.; Kim, S.M.; Yi, S.H.; Suh-Kim, H.; Jung, S.J.; Kim, M.J.; Kim, J.H.; Lee, Y.S.; Lee, S.Y.; et al. Generation of Dopamine Neurons from Rodent Fibroblasts through the Expandable Neural Precursor Cell Stage. J. Biol. Chem. 2015, 290, 17401–17414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.; Song, J.J.; Puspita, L.; Valiulahi, P.; Shim, J.W.; Lee, S.H. In Vitro generation of mature midbrain-type dopamine neurons by adjusting exogenous Nurr1 and Foxa2 expressions to their physiologic patterns. Exp. Mol. Med. 2017, 49, e300. [Google Scholar] [CrossRef]
- Song, J.J.; Oh, S.M.; Kwon, O.C.; Wulansari, N.; Lee, H.S.; Chang, M.Y.; Lee, E.; Sun, W.; Lee, S.E.; Chang, S.; et al. Cografting astrocytes improves cell therapeutic outcomes in a Parkinson’s disease model. J. Clin. Investig. 2018, 128, 463–482. [Google Scholar] [CrossRef] [Green Version]
- Mattugini, N.; Bocchi, R.; Scheuss, V.; Russo, G.L.; Torper, O.; Lao, C.L.; Götz, M. Inducing Different Neuronal Subtypes from Astrocytes in the Injured Mouse Cerebral Cortex. Neuron 2019, 103, 1086–1095.e5. [Google Scholar] [CrossRef] [Green Version]
- Salemi, S.; Baktash, P.; Rajaei, B.; Noori, M.; Amini, H.; Shamsara, M.; Massumi, M. Efficient generation of dopaminergic-like neurons by overexpression of Nurr1 and Pitx3 in mouse induced Pluripotent Stem Cells. Neurosci. Lett. 2016, 626, 126–134. [Google Scholar] [CrossRef]
- De Gregorio, R.; Pulcrano, S.; De Sanctis, C.; Volpicelli, F.; Guatteo, E.; von Oerthel, L.; Latagliata, E.C.; Esposito, R.; Piscitelli, R.M.; Perrone-Capano, C.; et al. miR-34b/c Regulates Wnt1 and Enhances Mesencephalic Dopaminergic Neuron Differentiation. Stem Cell Rep. 2018, 10, 1237–1250. [Google Scholar] [CrossRef] [Green Version]
- Lim, M.S.; Kim, S.M.; Lee, E.H.; Park, C.H. Efficient induction of neural precursor cells from fibroblasts using stromal cell-derived inducing activity. Tissue Eng. Regen Med. 2016, 13, 554–559. [Google Scholar] [CrossRef] [PubMed]
- Addis, R.C.; Hsu, F.C.; Wright, R.L.; Dichter, M.A.; Coulter, D.A.; Gearhart, J.D. Efficient conversion of astrocytes to functional midbrain dopaminergic neurons using a single polycistronic vector. PLoS ONE 2011, 6, e28719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caiazzo, M.; Dell’Anno, M.T.; Dvoretskova, E.; Lazarevic, D.; Taverna, S.; Leo, D.; Sotnikova, T.D.; Menegon, A.; Roncaglia, P.; Colciago, G.; et al. Direct generation of functional dopaminergic neurons from mouse and human fibroblasts. Nature 2011, 476, 224–227. [Google Scholar] [CrossRef] [PubMed]
- Nolbrant, S.; Giacomoni, J.; Hoban, D.B.; Bruzelius, A.; Birtele, M.; Chandler-Militello, D.; Pereira, M.; Ottosson, D.R.; Goldman, S.A.; Parmar, M. Direct Reprogramming of Human Fetal- and Stem Cell-Derived Glial Progenitor Cells into Midbrain Dopaminergic Neurons. Stem Cell Rep. 2020, 15, 869–882. [Google Scholar] [CrossRef] [PubMed]
- Raina, A.; Mahajani, S.; Bähr, M.; Kügler, S. Neuronal Trans-differentiation by Transcription Factors Ascl1 and Nurr1: Induction of a Dopaminergic Neurotransmitter Phenotype in Cortical GABAergic Neurons. Mol. Neurobiol. 2020, 57, 249–260. [Google Scholar] [CrossRef]
- Niu, W.; Zang, T.; Wang, L.L.; Zou, Y.; Zhang, C.L. Phenotypic Reprogramming of Striatal Neurons into Dopaminergic Neuron-like Cells in the Adult Mouse Brain. Stem Cell Rep. 2018, 11, 1156–1170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Li, F.; Stubblefield, E.A.; Blanchard, B.; Richards, T.L.; Larson, G.A.; He, Y.; Huang, Q.; Tan, A.C.; Zhang, D.; et al. Direct reprogramming of human fibroblasts into dopaminergic neuron-like cells. Cell Res. 2012, 22, 321–332. [Google Scholar] [CrossRef]
- Kim, J.; Su, S.C.; Wang, H.; Cheng, A.W.; Cassady, J.P.; Lodato, M.A.; Lengner, C.J.; Chung, C.Y.; Dawlaty, M.M.; Tsai, L.H.; et al. Functional integration of dopaminergic neurons directly converted from mouse fibroblasts. Cell Stem Cell 2011, 9, 413–419. [Google Scholar] [CrossRef] [Green Version]
- Song, B.; Cha, Y.; Ko, S.; Jeon, J.; Lee, N.; Seo, H.; Park, K.J.; Lee, I.H.; Lopes, C.; Feitosa, M.; et al. Human autologous iPSC-derived dopaminergic progenitors restore motor function in Parkinson’s disease models. J. Clin. Investig. 2020, 130, 904–920. [Google Scholar] [CrossRef] [Green Version]
- Bäckström, D.; Domellöf, M.E.; Granåsen, G.; Linder, J.; Mayans, S.; Elgh, E.; Mo, S.J.; Forsgren, L. PITX3 genotype and risk of dementia in Parkinson’s disease: A population-based study. J. Neurol. Sci. 2017, 381, 278–284. [Google Scholar] [CrossRef]
- Tiklová, K.; Björklund, Å.K.; Lahti, L.; Fiorenzano, A.; Nolbrant, S.; Gillberg, L.; Volakakis, N.; Yokota, C.; Hilscher, M.M.; Hauling, T.; et al. Single-cell RNA sequencing reveals midbrain dopamine neuron diversity emerging during mouse brain development. Nat. Commun. 2019, 10, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, Z.; Jiang, Y.; Li, T.; Zhu, J.; Zeng, S. Signaling of glial cell line-derived neurotrophic factor and its receptor GFRα1 induce Nurr1 and Pitx3 to promote survival of grafted midbrain-derived neural stem cells in a rat model of Parkinson disease. J. Neuropathol. Exp. Neurol. 2011, 70, 736–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’ANGLEMONT de Tassigny, X.; Pascual, A.; López-Barneo, J. GDNF-based therapies, GDNF-producing interneurons, and trophic support of the dopaminergic nigrostriatal pathway. Implications for Parkinson’s disease. Front. Neuroanat. 2015, 9, 10. [Google Scholar] [CrossRef] [Green Version]
- Peng, C.; Fan, S.; Li, X.; Fan, X.; Ming, M.; Sun, Z.; Le, W. Overexpression of pitx3 upregulates expression of BDNF and GDNF in SH-SY5Y cells and primary ventral mesencephalic cultures. FEBS Lett. 2007, 581, 1357–1361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Peng, C.; Li, X.; Fan, X.; Li, L.; Ming, M.; Chen, S.; Le, W. Pitx3-transfected astrocytes secrete brain-derived neurotrophic factor and glial cell line-derived neurotrophic factor and protect dopamine neurons in mesencephalon cultures. J. Neurosci. Res. 2008, 86, 3393–3400. [Google Scholar] [CrossRef] [PubMed]
- Failli, V.; Bachy, I.; Rétaux, S. Expression of the LIM-homeodomain gene Lmx1a (dreher) during development of the mouse nervous system. Mech. Dev. 2002, 118, 225–228. [Google Scholar] [CrossRef]
- Doucet-Beaupré, H.; Gilbert, C.; Profes, M.S.; Chabrat, A.; Pacelli, C.; Giguère, N.; Rioux, V.; Charest, J.; Deng, Q.; Laguna, A.; et al. Lmx1a and Lmx1b regulate mitochondrial functions and survival of adult midbrain dopaminergic neurons. Proc. Natl. Acad. Sci. USA 2016, 113, E4387–E4396. [Google Scholar] [CrossRef] [Green Version]
- Salesse, C.; Charest, J.; Doucet-Beaupré, H.; Castonguay, A.M.; Labrecque, S.; De Koninck, P.; Lévesque, M. Opposite Control of Excitatory and Inhibitory Synapse Formation by Slitrk2 and Slitrk5 on Dopamine Neurons Modulates Hyperactivity Behavior. Cell Rep. 2020, 30, 2374–2386.e5. [Google Scholar] [CrossRef] [Green Version]
- Davis, C.A.; Joyner, A.L. Expression patterns of the homeo box-containing genes En-1 and En-2 and the proto-oncogene int-1 diverge during mouse development. Genes Dev. 1988, 2, 1736–1744. [Google Scholar] [CrossRef]
- Marei, H.E.; Althani, A.; Afifi, N.; Michetti, F.; Pescatori, M.; Pallini, R.; Casalbore, P.; Cenciarelli, C.; Schwartz, P.; Ahmed, A.E. Gene expression profiling of embryonic human neural stem cells and dopaminergic neurons from adult human substantia nigra. PLoS ONE 2011, 6, e28420. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.W.; Piao, J.; Koo, S.Y.; Kriks, S.; Chung, S.Y.; Betel, D.; Socci, N.D.; Choi, S.J.; Zabierowski, S.; Dubose, B.N.; et al. Biphasic Activation of WNT Signaling Facilitates the Derivation of Midbrain Dopamine Neurons from hESCs for Translational Use. Cell Stem Cell 2021, 28, 343–355. [Google Scholar] [CrossRef]
- Kouwenhoven, W.M.; Veenvliet, J.V.; van Hooft, J.A.; van der Heide, L.P.; Smidt, M.P. Engrailed 1 shapes the dopaminergic and serotonergic landscape through proper isthmic organizer maintenance and function. Biol. Open 2016, 5, 279–288. [Google Scholar] [CrossRef] [Green Version]
- Prochiantz, A.; Fuchs, J.; Di Nardo, A.A. Postnatal signalling with homeoprotein transcription factors. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2014, 369. [Google Scholar] [CrossRef] [Green Version]
- Rekaik, H.; Blaudin de Thé, F.X.; Fuchs, J.; Massiani-Beaudoin, O.; Prochiantz, A.; Joshi, R.L. Engrailed Homeoprotein Protects Mesencephalic Dopaminergic Neurons from Oxidative Stress. Cell Rep. 2015, 13, 242–250. [Google Scholar] [CrossRef] [Green Version]
- Nordströma, U.; Beauvais, G.; Ghosh, A.; Pulikkaparambil Sasidharan, B.C.; Lundblad, M.; Fuchs, J.; Joshi, R.L.; Lipton, J.W.; Roholt, A.; Medicetty, S.; et al. Progressive nigrostriatal terminal dysfunction and degeneration in the engrailed1 heterozygous mouse model of Parkinson’s disease. Neurobiol. Dis. 2015, 73, 70–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stott, S.R.; Metzakopian, E.; Lin, W.; Kaestner, K.H.; Hen, R.; Ang, S.L. Foxa1 and foxa2 are required for the maintenance of dopaminergic properties in ventral midbrain neurons at late embryonic stages. J. Neurosci. 2013, 33, 8022–8034. [Google Scholar] [CrossRef] [Green Version]
- Dong, D.; Meng, L.; Yu, Q.; Tan, G.; Ding, M.; Tan, Y. Stable expression of FoxA1 promotes pluripotent P19 embryonal carcinoma cells to be neural stem-like cells. Gene Expr. 2012, 15, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Pristerà, A.; Lin, W.; Kaufmann, A.K.; Brimblecombe, K.R.; Threlfell, S.; Dodson, P.D.; Magill, P.J.; Fernandes, C.; Cragg, S.J.; Ang, S.L. Transcription factors FOXA1 and FOXA2 maintain dopaminergic neuronal properties and control feeding behavior in adult mice. Proc. Natl. Acad. Sci. USA 2015, 112, E4929–E4938. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Domanskyi, A.; Alter, H.; Vogt, M.A.; Gass, P.; Vinnikov, I.A. Transcription factors Foxa1 and Foxa2 are required for adult dopamine neurons maintenance. Front. Cell. Neurosci. 2014, 8, 275. [Google Scholar] [CrossRef] [Green Version]
- Mavromatakis, Y.E.; Lin, W.; Metzakopian, E.; Ferri, A.L.; Yan, C.H.; Sasaki, H.; Whisett, J.; Ang, S.L. Foxa1 and Foxa2 positively and negatively regulate Shh signalling to specify ventral midbrain progenitor identity. Mech. Dev. 2011, 128, 90–103. [Google Scholar] [CrossRef] [PubMed]
- Torper, O.; Ottosson, D.R.; Pereira, M.; Lau, S.; Cardoso, T.; Grealish, S.; Parmar, M. In Vivo Reprogramming of Striatal NG2 Glia into Functional Neurons that Integrate into Local Host Circuitry. Cell Rep. 2015, 12, 474–481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, M.; Birtele, M.; Shrigley, S.; Benitez, J.A.; Hedlund, E.; Parmar, M.; Ottosson, D.R. Direct Reprogramming of Resident NG2 Glia into Neurons with Properties of Fast-Spiking Parvalbumin-Containing Interneurons. Stem Cell Rep. 2017, 9, 742–751. [Google Scholar] [CrossRef] [PubMed]
- Tsai, R.Y. Creating a graft-friendly environment for stem cells in diseased brains. J. Clin. Investig. 2018, 128, 116–119. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Zhang, H.; Li, Y.; Tian, C.; Tang, B.; Huang, Y.; Zheng, J. Direct and selective lineage conversion of human fibroblasts to dopaminergic precursors. Neurosci. Lett. 2019, 699, 16–23. [Google Scholar] [CrossRef]
- Schweitzer, J.S.; Song, B.; Herrington, T.M.; Park, T.Y.; Lee, N.; Ko, S.; Jeon, J.; Cha, Y.; Kim, K.; Li, Q.; et al. Personalized iPSC-Derived Dopamine Progenitor Cells for Parkinson’s Disease. N. Engl. J. Med. 2020, 382, 1926–1932. [Google Scholar] [CrossRef] [PubMed]
- Doi, D.; Magotani, H.; Kikuchi, T.; Ikeda, M.; Hiramatsu, S.; Yoshida, K.; Amano, N.; Nomura, M.; Umekage, M.; Morizane, A.; et al. Pre-clinical study of induced pluripotent stem cell-derived dopaminergic progenitor cells for Parkinson’s disease. Nat. Commun. 2020, 11, 3369. [Google Scholar] [CrossRef]
- Doi, D.; Samata, B.; Katsukawa, M.; Kikuchi, T.; Morizane, A.; Ono, Y.; Sekiguchi, K.; Nakagawa, M.; Parmar, M.; Takahashi, J. Isolation of human induced pluripotent stem cell-derived dopaminergic progenitors by cell sorting for successful transplantation. Stem Cell Rep. 2014, 2, 337–350. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, J. iPS cell-based therapy for Parkinson’s disease: A Kyoto trial. Regen. Ther. 2020, 13, 18–22. [Google Scholar] [CrossRef]
- Takahashi, J. Preclinical evaluation of patient-derived cells shows promise for Parkinson’s disease. J. Clin. Investig. 2020, 130, 601–603. [Google Scholar] [CrossRef]
- Park, C.H.; Kang, J.S.; Kim, J.S.; Chung, S.; Koh, J.Y.; Yoon, E.H.; Jo, A.Y.; Chang, M.Y.; Koh, H.C.; Hwang, S.; et al. Differential actions of the proneural genes encoding Mash1 and neurogenins in Nurr1-induced dopamine neuron differentiation. J. Cell Sci. 2006, 119, 2310–2320. [Google Scholar] [CrossRef] [Green Version]
- Andersson, E.K.; Irvin, D.K.; Ahlsiö, J.; Parmar, M. Ngn2 and Nurr1 act in synergy to induce midbrain dopaminergic neurons from expanded neural stem and progenitor cells. Exp. Cell Res. 2007, 313, 1172–1180. [Google Scholar] [CrossRef] [PubMed]
- Xia, N.; Zhang, P.; Fang, F.; Wang, Z.; Rothstein, M.; Angulo, B.; Chiang, R.; Taylor, J.; Reijo Pera, R.A. Transcriptional comparison of human induced and primary midbrain dopaminergic neurons. Sci. Rep. 2016, 6, 20270. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| TFs | Cell Type | Methods | Major Findings | Ref. | |
|---|---|---|---|---|---|
| Nurr1 | Mouse ESCs D3 cell line | In vitro | Overexpress Nurr1 | DA neurons markers: Dat, Aadc, Th and Pitx3, Increase the number of DA neurons and the expression of DA markers DA production and release | [81] |
| Mouse ESCs R1 cell line | In vitro | Overexpress Nurr1 | DA neurons markers: Th, Aadc, Nurr1 and Pitx3 Increase in the proportion of Th+ neurons | [82] | |
| In vivo | Transplantation Nurr1-overexpressed MSCs into PD animals | DA production and release Amelioration of PD motor symptoms in a rodent PD model for 8 weeks Similar electrophysiological characteristics to mesencephalic neurons | |||
| Mouse ESCs R1 cell line | In vitro | Co-express Nurr1 and GPX-1 | DA-like cells: Nestin, Map2 and Tau; Nurr1, DdC, and Th DA release A proportion of immature DA-like cells | [83] | |
| Rat NPCs derived from cortices (E13.5) | In vitro | Exposure of exogenous Nurr1-expressing NPCs to UCN | Increase the expression of mature DA neurons markers: Dat, Aadc, Th Up-regulate the expression of Nurr1, Foxa2, and Pitx3 Increase Th+ cells rather than Nurr1+ cells | [85] | |
| Rat embryo (E16.5) | In vivo | UCN intraperitoneal administration | Increase the differentiation of Nurr1+ precursors into Th+ DA neurons | ||
| Primary microglia | In vitro | Exposure of exogenous Nurr1-expressing primary microglia to LPS | Downregulate inflammatory factors (IL-1 and TNFα) Up-regulate neurotrophic factors (BDNF and GDNF) | [86,87] | |
| Rat mNSCs (E14.5) | In vitro | NNSC + NMG co-culture | DA neurons markers: Th, Pitx3, Dat | [87] | |
| Rat mNSCs (E12.5–14.5) | In vitro | Overexpress Nurr1 in NSCs | DA neurons markers: Th, Dat | [86] | |
| In vivo | Transplantation of NNSC + NMG into PD rats | DA neurons markers: Th, Dat, and Pitx3 in the grafts Reduce the number of reactive microglia after transplantation Reverse motor behavior deficits Ensure a long-time significant outcome | |||
| Rat bone marrow mesenchymal stem cells (MSCs) (6w) | In vivo | Transplantation Nurr1-overexpressed MSCs into PD rats | DA neurons: Nurr1, Th, and Dat Survive and migrate in the brain, Suppress the activation of neuroglial cells and the expression of pro-inflammatory factors Reverse motor behavior deficits | [89] | |
| Mouse embryonic OBSCs (E13.5) | In vitro | Forced Nurr1 Expression in OBSCs | Mature-like mesencephalic neurons: Th, GIRK2, Vmat2, Dat, calretinin, calbindin DA-GABAergic neurons: Th, GAD, GABA, VGAT DA release A proportion of immature neurons | [90] | |
| Pitx3 | NPs derived from mouse blastocyst-derived ES cell line J1 (ES-NP) | In vitro | Overexpression of Pitx3 in Shh/Fgf8 pretreated NPs | DA neurons Th, Aadc, Vmat2, and Dat Directly bind to the Th, Ngn2, and Tuj1 gene promoter and induce their transcription. | [91] |
| Human ESC line H9 | In vivo | Transplantation PITX3-eGFP+ cell into PD rats | Not restore functional deficits in PD rats Reduced size of the PITX3-GFP+ cell grafts | [92] | |
| In vivo | Transplantation PITX3-eGFP-cell into PD rats | Restore functional deficits | |||
| Human teratocarcinoma cell line Ntera2 (NT2) | In vitro | Culture in a growth medium supplemented with knock-out serum and retinoic acid exposed to GDNF | DA neurons markers: Th, Aadc, Dat DA release Accelerate Th expression and induce DA signaling Promote neuroprotection | [93] | |
| In vivo | Transplantation NT2 cells transduced with Pitx3 into PD rats | Promote neuroprotection Increase in striatal volume. Restore functional deficits | |||
| Lmx1a | H9-derived human neural progenitor cell line (hNP1) | In vitro | Forced Lmx1a expression in hNP1 | Increase Th+ neurons both during NPC and induction stages | [15] |
| Human ESC line H9 | In vivo | Transplantation Lmx1a-eGFP+ VM progenitors from human ESC line into PD rats | Improve the Safety and Predictability Accelerate behavioral recovery Highly enrich for DA neurons Reduce proliferating cell populations Eliminate serotonergic neurons | [92] | |
| EBCs (embryoid body cells) derived from the R1B5 ESC line | In vivo | Transplantation EBCs transduced with Lmx1a into the intact SNpc | DA neurons Th+ cells derived only from EBCs exogenously expressing Lmx1a Additional factors appear to be required to complete differentiation and/or increase the long-term survival of these putative | [94] | |
| In vivo | Transplantation into the lesioned SNpc | DA neurons The emergence of Th+ cells from EBCs Fail to long-term survival | |||
| Nurr1 + Foxa2 | NPCs from mouse VM and cortex (E10–12) | In vitro | Co-express Nurr1 and Foxa2 in NPCs | Increase Th+ cells Synergistic increase of DA genes: Dat, Th, Vmat2, Tuj1, Map2, Pitx3 | [48] |
| NPCs from mouse VM (E10–12) | In vivo | Co-express Nurr1 and Foxa2 in NPCs | Resistance to toxic stimuli Restoration of PD motor symptoms in a rodent PD model for a long time | [49] | |
| iNPCs from rat fibroblasts (E13.5) | In vitro | The combined expression of Nurr1 and Foxa2 in iNPCs | Mature/functional DA neuron neurons: Map2, HuC/D, synapsin l, Dat, Vmat2, Th, Tuj1, Pitx3 Action potentials DA release Restore functional deficits The difference in the maturity and function of DA neurons derived from iNPCs in vivo in transplanted brains | [95] | |
| NPCs from cortices of rat embryos (E14) | In vitro | Forced expression of Nurr1 and Foxa2 using lenti-pUb and retro-pLTR systems | Mature DA neuron generation Mature neurons (Map2, NeuN) Mature DA neurons (Dat, Vmat2) Midbrain-specific DA neurons (Pitx3) | [96] | |
| Ctx-Ast or VM-Ast | In vitro | Co-culture with VM-NPCs (mouse E10.5 or rat E12) Or co-graft with VM-NPCs | DA release increase Further potentiates the neuroprotective actions Downregulate pro-inflammatory cytokines (IL-1b, IL-6, TNF-α, and iNOS) and myelin-associated proteins (Mbp, Mag, and Mog) Up-regulate secretory neurotrophic (Shh, BDNF, GDNF, and NT3) and anti-inflammatory (TNF-α, TNF-ß) factors Increase antioxidant enzymes | [97] | |
| Nurr1 + Mash1 | Rat cortical or VM NPCs (E14) and human NPC derived from human ESC line H9 | In vitro | Exposure of Nurr1-Mash1-overexpressing NPCs to thyroid hormone derivatives | DA release DA neurons markers: Th, Dat,Nurr1,En1,Lmx1b Protect DA neurons from neurotoxic damage Not affect astrocytes or non-DA neurons | [84] |
| Nurr1 + Ngn2 | Mouse embryonic OBSCs E13.5 | In vitro | Co-expressing Nurr1 and Ngna2 in OBSCs | Reduction in Th+ neural proportion SV2 and Synapsin | [90] |
| Astrocytes | In vivo | Viral injection after stab wound injury | NeuN+ cells Induced neurons originate from both proliferating and quiescent astrocytes lamina-specific hallmarks Appropriate long-distance axonal projections White matter astrocytes fail to undergo neuronal reprogramming | [98] | |
| Nurr1 + Pitx3 | iPSCs from Mouse embryonic fibroblasts (MEFs) | In vitro | Transduce iPSCs by Nurr1- and Pitx3-harboring lentiviruses | DA-like cells: Th, Ddc, Dat, Map2 DA release increase | [99] |
| Ascl1 + Nurr1 | MEFs | In vitro | Infect MEFs with inducible lentiviruses expressing miR-34b/c cluster in combination with Ascl1 and Nurr1 | DA neurons increase Spontaneous electrical activity Up-regulate DA markers: Th, Dat, Vmat2, and Pitx3 | [100] |
| Brn2, Ascl1, and Myt1L (BAM) | Mouse or rat fibroblasts (E13.5) | In vitro | Co-express BAM and Bcl-xL in fibroblasts | iNPCs: Tuj1, nestin, Sox2 Features of neural lineage cells. No regional identity and neuronal subtype differentiation potential Mature DA neurons from iNPCs overexpressed Nurr1 and Foxa2 | [95] |
| Shh overexpressing MS5 stromal (MS5-Shh) cells | In vitro | MS5-Shh cells transduced with BAM and Bcl-xL Co-culture with fibroblasts | DA neurons | [101] | |
| Ascl1 + Lmx1a + Nurr1 (ALN) | Primary postnatal mouse astrocytes (strain CD1) | In vitro | Co-expression ALN in Astrocytes | Functional DA Neurons: Tuj1, Pitx3, Lmx1a, En1, aldehyde dehydrogenase, Foxa2, Vmat2, Msx1, and Dat DA release Spontaneous firing of action potentials | [102] |
| Mouse (E14.5) and human fibroblasts (IMR90) | In vitro | Co-express ALN in fibroblasts | Functional DA neurons: Th, Vmat2, Dat, ALDH1A1, calbindin, Release DA Spontaneous electrical activity Down-regulation of the fibroblast markers: Twist2, Zeb2, Tgfb1i1, and Chd2 Establishment of DA synaptic terminals | [103] | |
| In vivo | Transplant fibroblasts transduced ALN into the ventricle of newborn mouse brains. | iDANs markers: Th, Aadc, Vmat2, Dat Maintain excitability and major currents | |||
| Human Fetal- and Stem Cell-Derived Glial Progenitor Cells | In vitro | Transduced together with short hairpin (sh) RNA against the RE1-silencing transcription factor (REST) complex into cells | Functionally mature iDANs Glial markers down-regulation DA-related genes: Th, Dat, Foxa2, Lmx1a, and Pitx3 Spontaneous firing at resting membrane potential | [104] | |
| Rat embryonal cortex at E18.5 | In vitro | Transduced ALN into Rat embryonal cortex | Mature DA neurons: Th, NeuN, Aadc, Vmat2, Dat Fail to detect secreted DA, DOPAC or HVA DA reprogramming takes place only in GABAergic cortical neurons | [105] | |
| Pitx3 + Foxa2 + Lmx1a | H9-derived human neural progenitor cell line (hNP1) | In vitro | Transduced Pitx3, Foxa2, Lmx1a mRNA vectors into hNP1 | Th+ neurons Midbrain-specific markers: Nurr1, Vmat2 | [15] |
| Neurod1 + Ascl1 + Lmx1a + miR-218 (NeAL218) | Human immature astrocytes | In vitro | Reprogram human astrocytes with NeAL218 | iDANs: Th+/TUBB3+ cells Neuronal morphology Lacked membrane properties of excitable DAs | [16] |
| Mouse astrocytes | In vivo | Inject NeAL218 lentiviruses into PD mouse | iDANs without tumors or died No GAD1/2, somatostatin (SST), parvalbumin (PVALB), or calretinin (CALB2) expression in iDANs Mature DA markers: Dat, RBFOX3, Nurr1, and PBX1 DA release Rescue spontaneous motor behavior | ||
| Foxa2 + En1 + Lmx1a + Pitx3 | Human ESCs (H9) or mouse ESCs | In vitro | Coexpress Foxa2, En1, Lmx1a, and Pitx3 in human ESCs (H9) or mouse ESCs | Functional iDANs cells with midbrain characteristics 70% of the Map2+ Th+ induced neural cells Express Ascl1, Nurr1, Lmx1a, En1, Foxa2, and Pitx3 in Th+ induced neural cells DA neurons markers: Th, Vmat2, Dat, Aadc, and Girk2 DA release Electrical properties typical of mesencephalic DA neurons, | [17] |
| Sox2 + Nurr1 + Lmx1a + Foxa2 | Striatal Neurons in the Adult Mouse Brain | In vivo | Inject virus including Sox2, Nurr1, Lmx1a and Foxa2 into adult mouse striatal; VPA in drinking water was administered | Mouse striatal neurons are reprogrammed into induced dopaminergic neuron-like cells (iDALs) without a proliferative progenitor stage DA neurons markers: Ddc, Vamt2, Dat, Th without other neuronal subtypes such as ChAT and VGLUT1. iDALs originate from local striatal neurons Electrophysiological properties similar to DA neurons Firing patterns stereotypical to DA neuron Form functional connections with other neurons | [106] |
| Ascl1 + Ngn2 + Sox2 + Nurr1 + Pitx3 | IMR90 human fibroblasts | In vitro | Co-express Ascl1, Ngn2, Sox2, Nurr1, Pitx3 in IMR90 human fibroblasts | DA Neurons Markers: Ddc, Vamt2, Dat, Th, En1 Negative for serotonin (a marker for serotogenic neurons) and ChAT (a marker for cholinergic neurons) Lack of cell proliferation DA uptake and production DA neuron-like electrophysiology Relief of PD symptoms | [107] |
| Ascl1 + Pitx3 + Nurr1 + Lmx1a + En1 + Foxa2 | Tail tip fibroblasts (TTFs) from adult mice | In vitro | Co-express Ascl1, Pitx3, Nurr1, Lmx1a, En1and Foxa2 in TTFs | DA neuronal marker: Aadc, Vamt2, Dat, Th Mature neuronal makers: Map2 No 5-HT and motor neurons DA release DA neuron-like electrophysiology | [108] |
| In vivo | Transplant Pitx3-eGFP+ cells isolated from TTFs 12 after transduction with 6 factors into PD models | Neuronal morphology cc DA neuron markers: Th, Aadc Relief of PD symptoms Increase DA production | |||
| Oct4 + Sox2 + Klf4 + c-Myc | Human BJ dermal fibroblasts (hDF) | In vitro | spotting culture and quercetin treatment after forced expression Oct4, Sox2, Klf4, c-Myc, miR302s and miR200c in hDF | DA neuronal marker: Th, Dat, Pitx3, Vmat2 DA neuron-like electrophysiology | [109] |
| In vivo | Transplate these iPSCs-derived NPCs into PD models | Relief of PD symptoms Form functional connections with other neurons DA neuronal markers: Th, Dat, Vmat2, Nurr1 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tian, L.; Al-Nusaif, M.; Chen, X.; Li, S.; Le, W. Roles of Transcription Factors in the Development and Reprogramming of the Dopaminergic Neurons. Int. J. Mol. Sci. 2022, 23, 845. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23020845
Tian L, Al-Nusaif M, Chen X, Li S, Le W. Roles of Transcription Factors in the Development and Reprogramming of the Dopaminergic Neurons. International Journal of Molecular Sciences. 2022; 23(2):845. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23020845
Chicago/Turabian StyleTian, Lulu, Murad Al-Nusaif, Xi Chen, Song Li, and Weidong Le. 2022. "Roles of Transcription Factors in the Development and Reprogramming of the Dopaminergic Neurons" International Journal of Molecular Sciences 23, no. 2: 845. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms23020845