
Zfhx3 Transcription Factor Represses the Expression of SCN5A Gene and Decreases Sodium Current Density (INa)

 , , , , ,
, , , , ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. Zfhx3 Is Expressed in the Human Myocardium
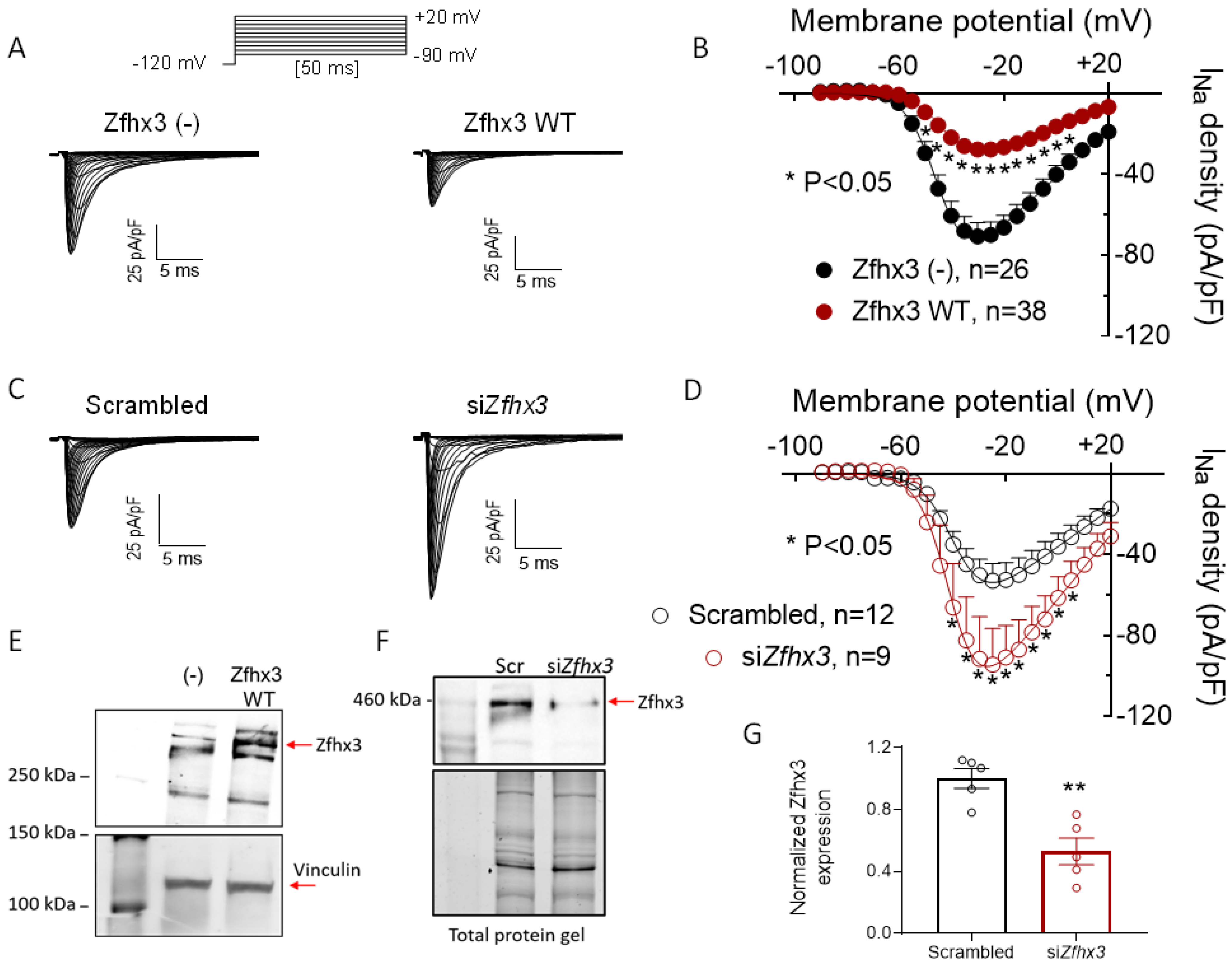
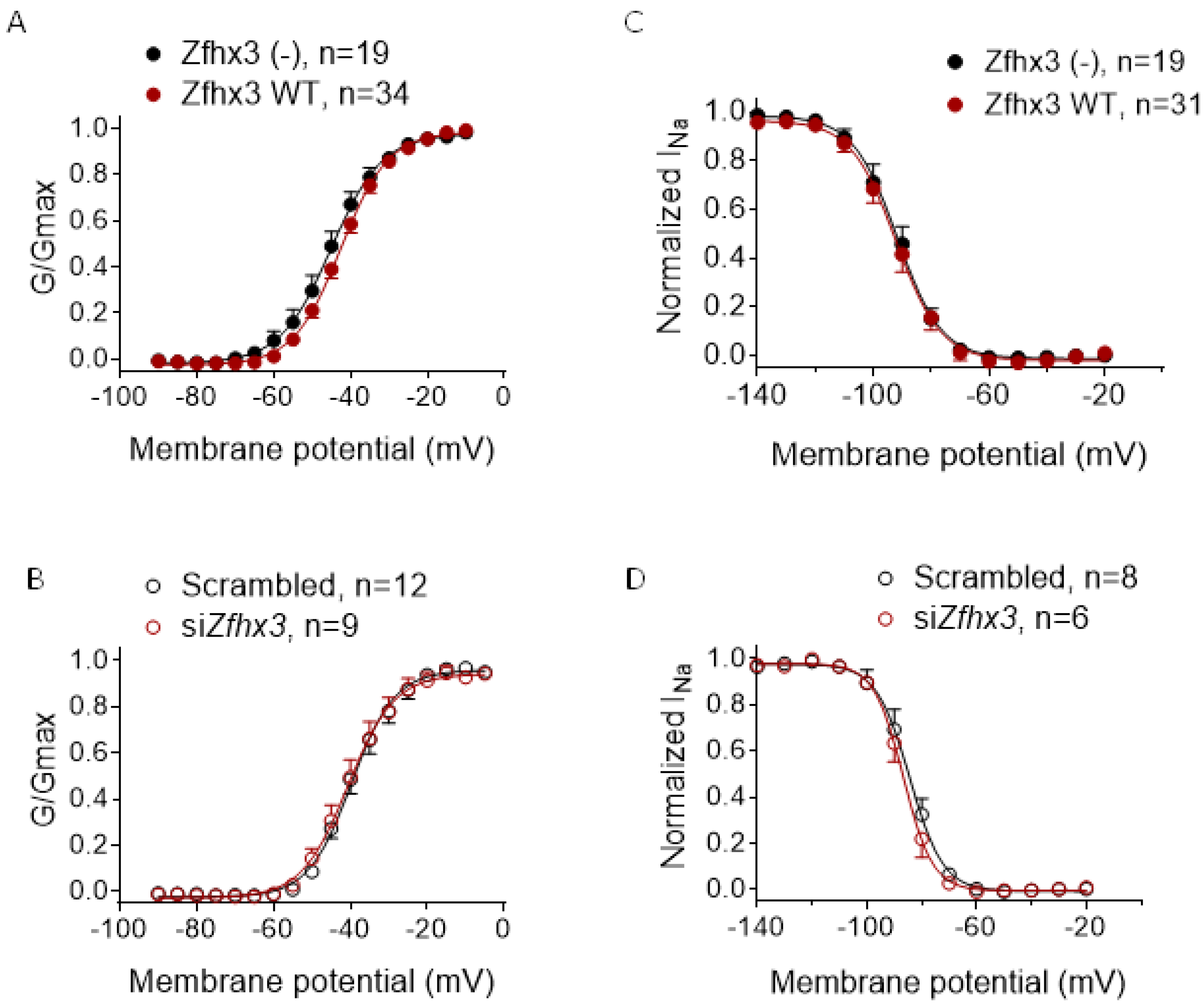
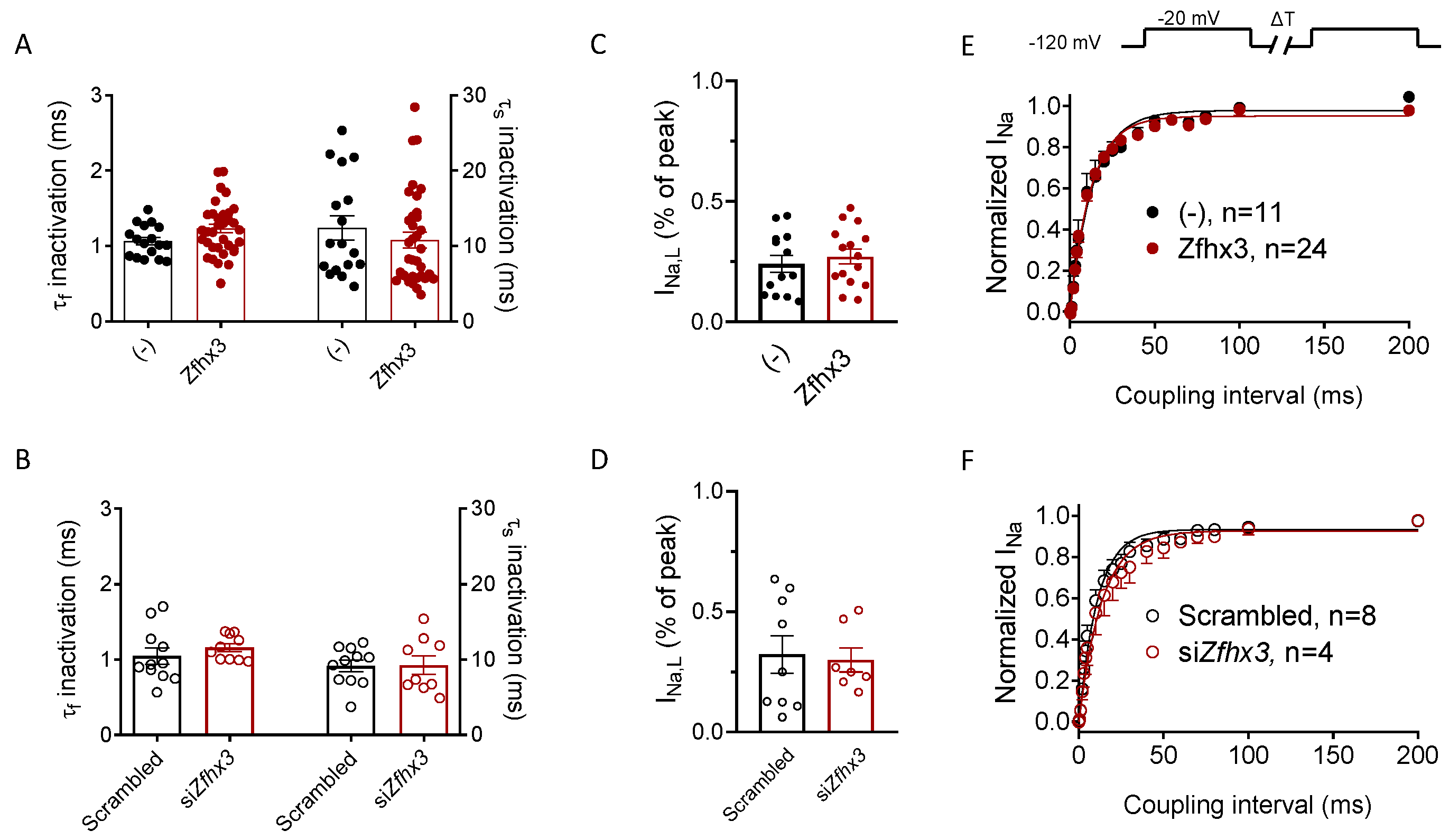
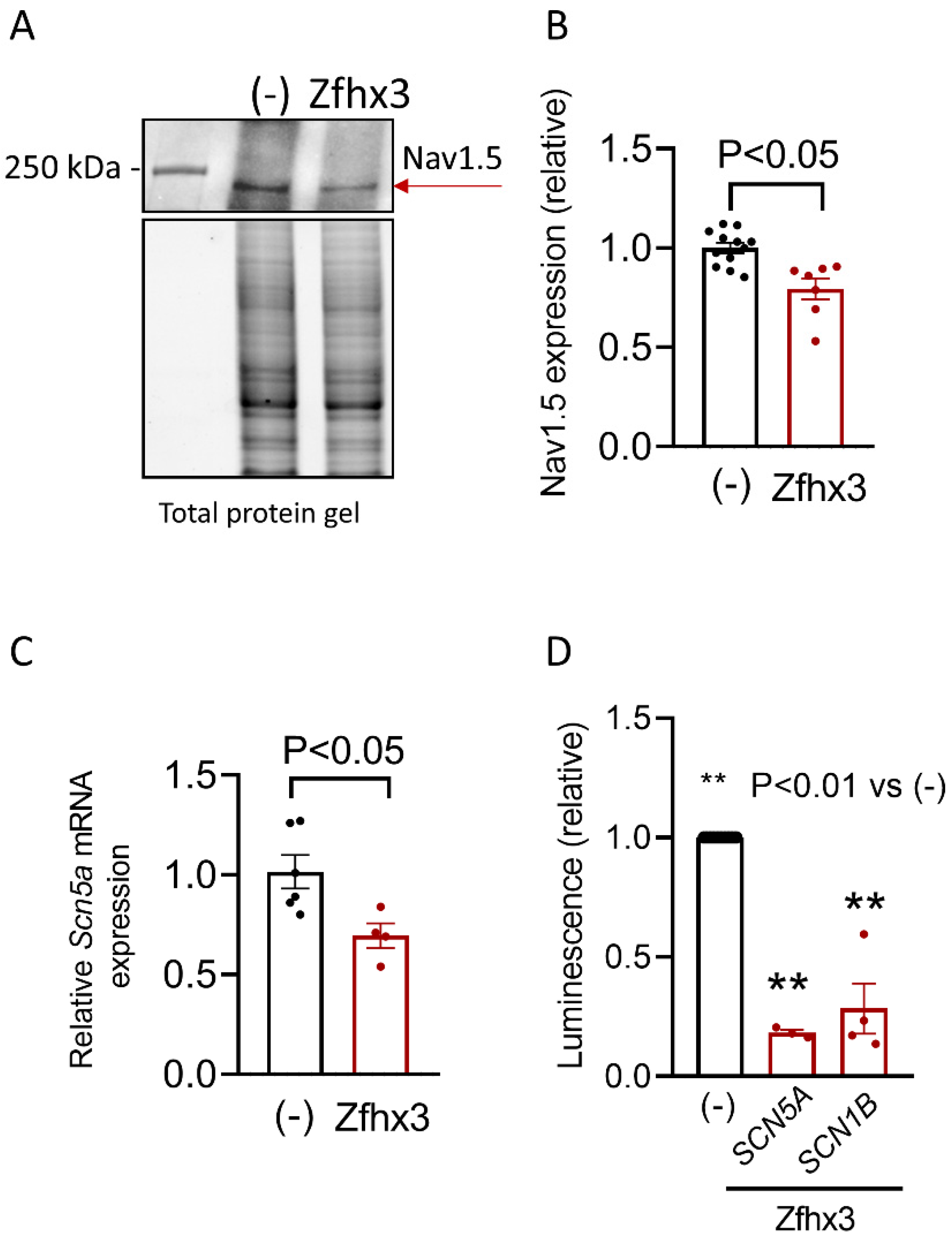
2.2. Zfhx3 Markedly Reduces INa
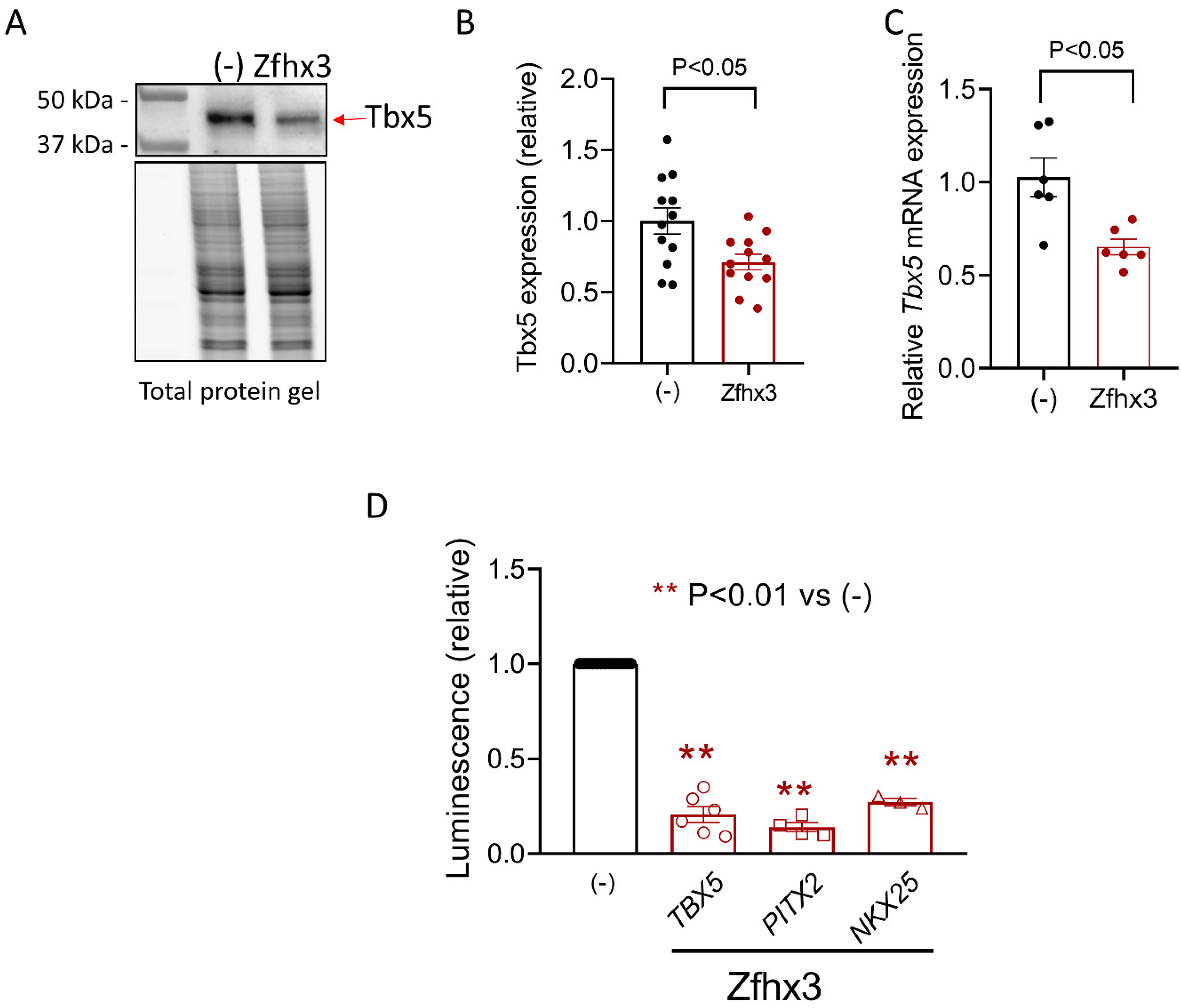
2.3. Zfhx3, Tbx5 and Pitx2c Interplay
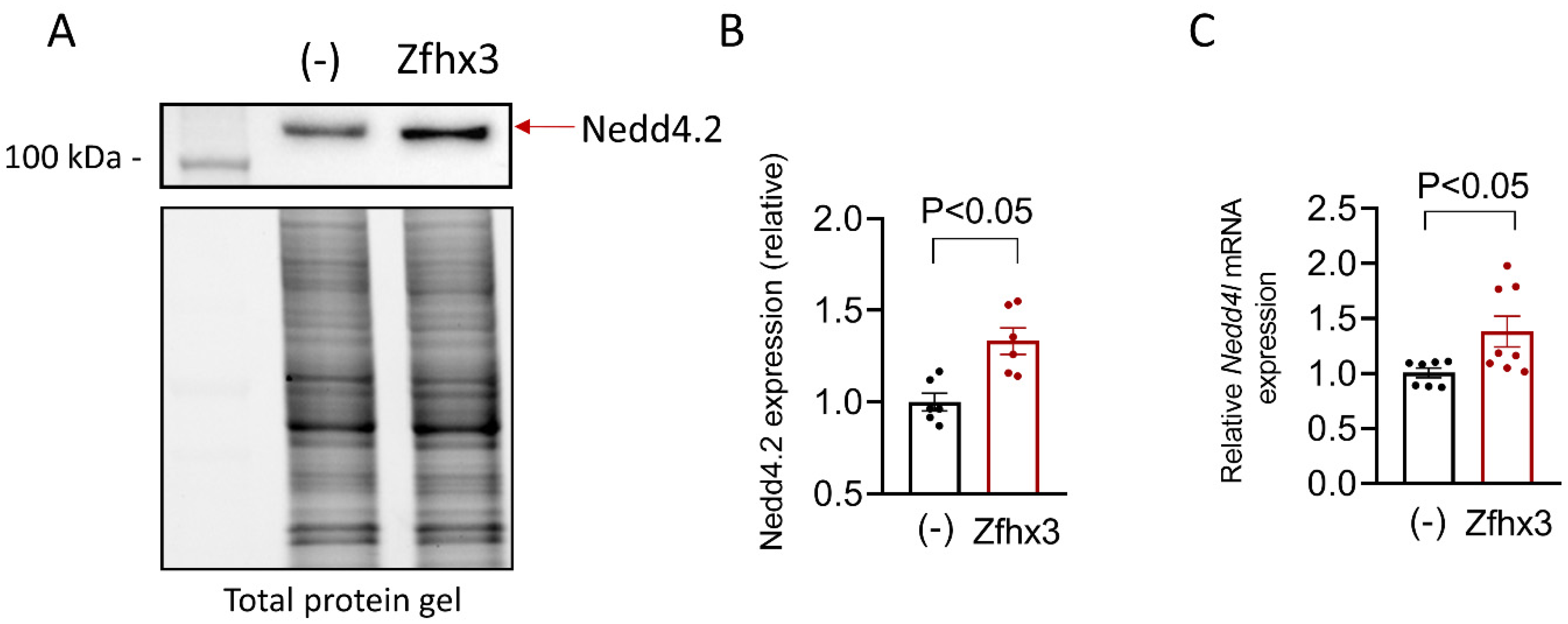
2.4. Zfhx3 and Nedd4.2 Expression
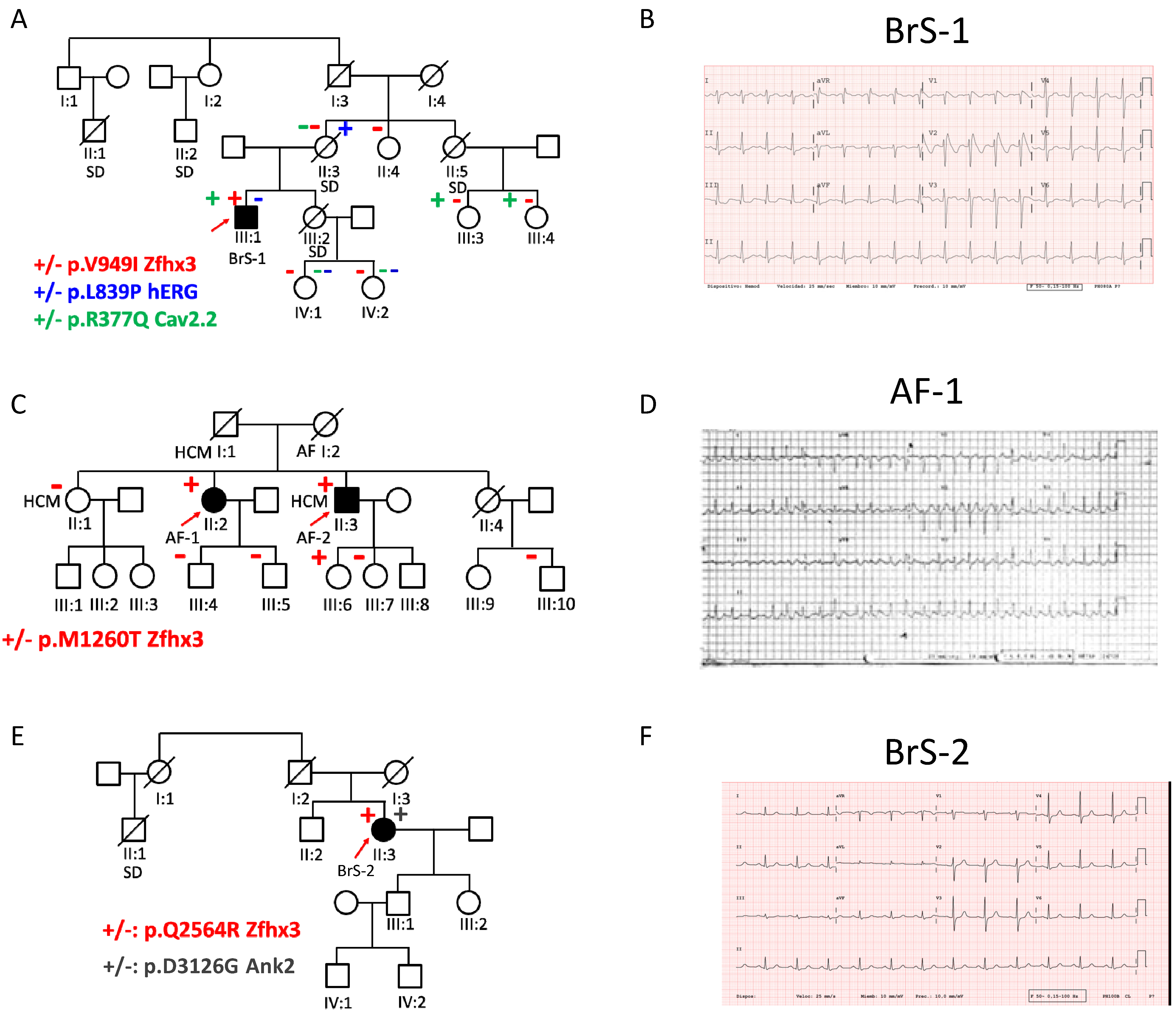
2.5. p.V949I, p.M1260T, and p.Q2564R Zfhx3 Variants
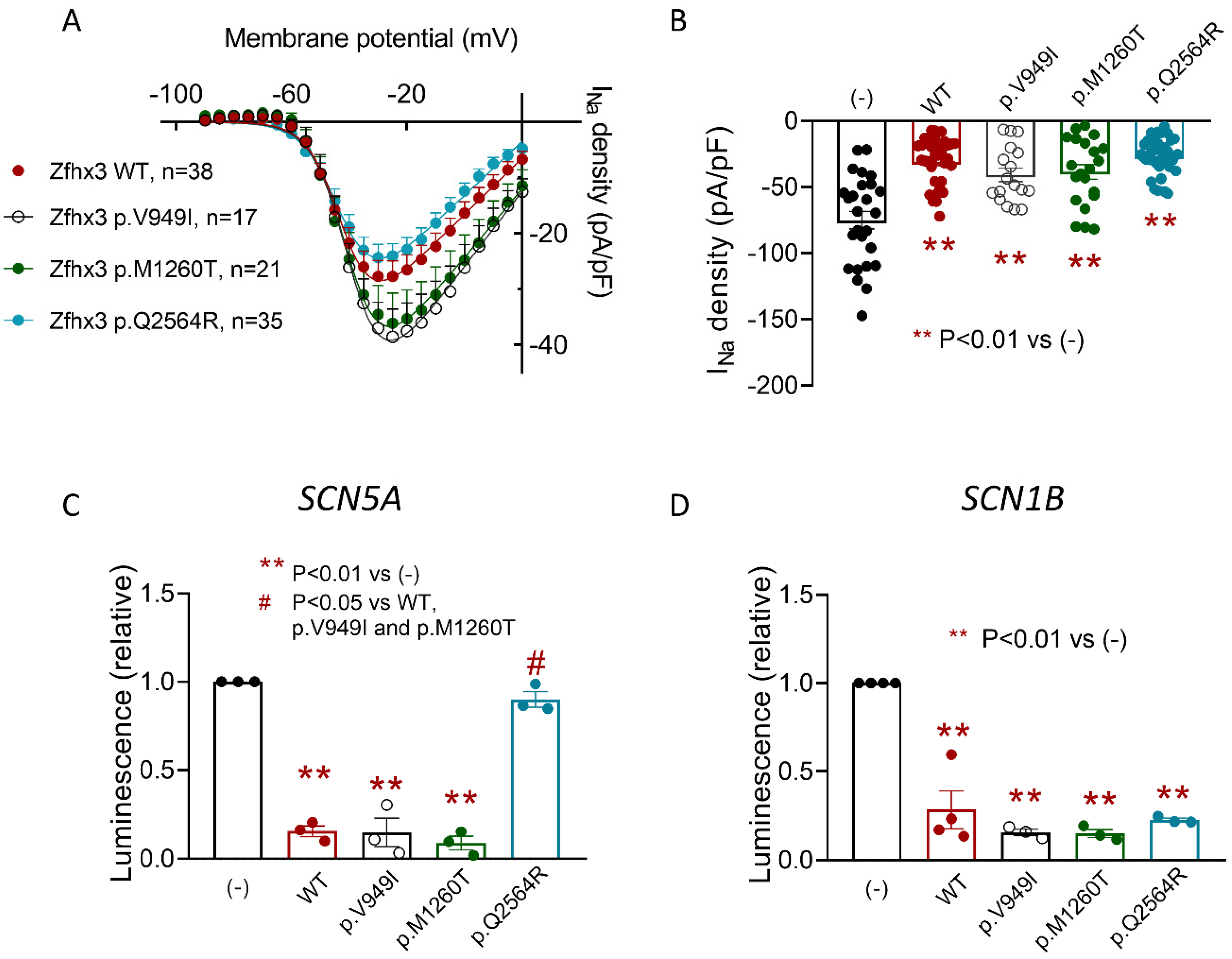
2.6. Effects of the p.V949I, p.M1260T, or p.Q2564R Zfhx3 on the INa
3. Discussion
Limitations of the Study
4. Material and Methods
4.1. Study Approval
4.2. DNA Sequencing
4.3. Access to Public Human Cardiac RNA-Seq Data
4.4. Cell Culture and Transfection
4.5. Patch-Clamp Recordings
4.6. Analysis of the mRNA Expression (RT-qPCR)
4.7. Western Blot Analysis and Zfhx3 Silencing
4.8. Luciferase Assays
4.9. Statistical Analyses
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Flicek, P.; Ahmed, I.; Amode, M.R.; Barrell, D.; Beal, K.; Brent, S.; Carvalho-Silva, D.; Clapham, P.; Coates, G.; Fairley, S.; et al. Ensembl 2013. Nucleic Acids Res. 2013, 41, D48–D55. [Google Scholar] [CrossRef] [PubMed]
- Berry, F.B.; Miura, Y.; Mihara, K.; Kaspar, P.; Sakata, N.; Hashimoto-Tamaoki, T.; Tamaoki, T. Positive and Negative Regulation of Myogenic Differentiation of C2C12 Cells by Isoforms of the Multiple Homeodomain Zinc Finger Transcription Factor ATBF1. J. Biol. Chem. 2001, 276, 25057–25065. [Google Scholar] [CrossRef] [Green Version]
- Minamiya, Y.; Saito, H.; Ito, M.; Imai, K.; Konno, H.; Takahashi, N.; Motoyama, S.; Ogawa, J.-I. Suppression of Zinc Finger Homeobox 3 Expression in Tumor Cells Decreases the Survival Rate among Non-Small Cell Lung Cancer Patients. Cancer Biomark. Sect. Dis. Markers 2012, 11, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, C.; Yao, Y.; Zuo, X.; Chen, S.; Xu, C.; Zhang, H.; Lu, Q.; Chang, L.; Wang, F.; et al. Molecular Basis of Gene-Gene Interaction: Cyclic Cross-Regulation of Gene Expression and Post-GWAS Gene-Gene Interaction Involved in Atrial Fibrillation. PLoS Genet. 2015, 11, e1005393. [Google Scholar] [CrossRef] [PubMed]
- Gudbjartsson, D.F.; Holm, H.; Gretarsdottir, S.; Thorleifsson, G.; Walters, G.B.; Thorgeirsson, G.; Gulcher, J.; Mathiesen, E.B.; Njølstad, I.; Nyrnes, A.; et al. A Sequence Variant in ZFHX3 on 16q22 Associates with Atrial Fibrillation and Ischemic Stroke. Nat. Genet. 2009, 41, 876–878. [Google Scholar] [CrossRef]
- Benjamin, E.J.; Rice, K.M.; Arking, D.E.; Pfeufer, A.; van Noord, C.; Smith, A.V.; Schnabel, R.B.; Bis, J.C.; Boerwinkle, E.; Sinner, M.F.; et al. Variants in ZFHX3 Are Associated with Atrial Fibrillation in Individuals of European Ancestry. Nat. Genet. 2009, 41, 879–881. [Google Scholar] [CrossRef]
- Traylor, M.; Farrall, M.; Holliday, E.G.; Sudlow, C.; Hopewell, J.C.; Cheng, Y.-C.; Fornage, M.; Ikram, M.A.; Malik, R.; Bevan, S.; et al. Genetic Risk Factors for Ischaemic Stroke and Its Subtypes (the METASTROKE Collaboration): A Meta-Analysis of Genome-Wide Association Studies. Lancet Neurol. 2012, 11, 951–962. [Google Scholar] [CrossRef] [Green Version]
- Lettre, G.; Palmer, C.D.; Young, T.; Ejebe, K.G.; Allayee, H.; Benjamin, E.J.; Bennett, F.; Bowden, D.W.; Chakravarti, A.; Dreisbach, A.; et al. Genome-Wide Association Study of Coronary Heart Disease and Its Risk Factors in 8090 African Americans: The NHLBI CARe Project. PLoS Genet. 2011, 7, e1001300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, C.; Wang, F.; Yang, Y.; Fu, F.; Xu, C.; Shi, L.; Li, S.; Xia, Y.; Wu, G.; Cheng, X.; et al. Significant Association of SNP Rs2106261 in the ZFHX3 Gene with Atrial Fibrillation in a Chinese Han GeneID Population. Hum. Genet. 2011, 129, 239–246. [Google Scholar] [CrossRef] [Green Version]
- Magnani, J.W.; Yin, X.; McManus, D.D.; Chuang, M.L.; Cheng, S.; Lubitz, S.A.; Arora, G.; Manning, W.J.; Ellinor, P.T.; Benjamin, E.J. Genetic Loci Associated with Atrial Fibrillation: Relation to Left Atrial Structure in the Framingham Heart Study. J. Am. Heart Assoc. 2014, 3, e000616. [Google Scholar] [CrossRef] [Green Version]
- Jiang, T.; Wang, Y.-N.; Qu, Q.; Qi, T.-T.; Chen, Y.-D.; Qu, J. Association between Gene Variants and the Recurrence of Atrial Fibrillation: An Updated Meta-Analysis. Medicine 2019, 98, e15953. [Google Scholar] [CrossRef] [PubMed]
- Thorolfsdottir, R.B.; Sveinbjornsson, G.; Aegisdottir, H.M.; Benonisdottir, S.; Stefansdottir, L.; Ivarsdottir, E.V.; Halldorsson, G.H.; Sigurdsson, J.K.; Torp-Pedersen, C.; Weeke, P.E.; et al. Genetic Insight into Sick Sinus Syndrome. Eur. Heart J. 2021, 42, 1959–1971. [Google Scholar] [CrossRef]
- Nieto-Marín, P.; Tinaquero, D.; Utrilla, R.G.; Cebrián, J.; González-Guerra, A.; Crespo-García, T.; Cámara-Checa, A.; Rubio-Alarcón, M.; Dago, M.; Alfayate, S.; et al. Tbx5 Variants Disrupt Nav1.5 Function Differently in Patients Diagnosed with Brugada or Long QT Syndrome. Cardiovasc. Res. 2021. published ahead of print. [Google Scholar] [CrossRef]
- Chinchilla, A.; Daimi, H.; Lozano-Velasco, E.; Dominguez, J.N.; Caballero, R.; Delpón, E.; Tamargo, J.; Cinca, J.; Hove-Madsen, L.; Aranega, A.E.; et al. PITX2 Insufficiency Leads to Atrial Electrical and Structural Remodeling Linked to Arrhythmogenesis. Circ. Cardiovasc. Genet. 2011, 4, 269–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nadadur, R.D.; Broman, M.T.; Boukens, B.; Mazurek, S.R.; Yang, X.; van den Boogaard, M.; Bekeny, J.; Gadek, M.; Ward, T.; Zhang, M.; et al. Pitx2 Modulates a Tbx5-Dependent Gene Regulatory Network to Maintain Atrial Rhythm. Sci. Transl. Med. 2016, 8, 354ra115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- GTEx Consortium The Genotype-Tissue Expression (GTEx) Project. Nat. Genet. 2013, 45, 580–585. [CrossRef]
- Pérez-Hernández, M.; Matamoros, M.; Barana, A.; Amorós, I.; Gómez, R.; Núñez, M.; Sacristán, S.; Pinto, Á.; Fernández-Avilés, F.; Tamargo, J.; et al. Pitx2c Increases in Atrial Myocytes from Chronic Atrial Fibrillation Patients Enhancing IKs and Decreasing ICa,L. Cardiovasc. Res. 2016, 109, 431–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Bemmelen, M.X.; Rougier, J.-S.; Gavillet, B.; Apothéloz, F.; Daidié, D.; Tateyama, M.; Rivolta, I.; Thomas, M.A.; Kass, R.S.; Staub, O.; et al. Cardiac Voltage-Gated Sodium Channel Nav1.5 Is Regulated by Nedd4-2 Mediated Ubiquitination. Circ. Res. 2004, 95, 284–291. [Google Scholar] [CrossRef] [Green Version]
- Richards, S.; Aziz, N.; Bale, S.; Bick, D.; Das, S.; Gastier-Foster, J.; Grody, W.W.; Hegde, M.; Lyon, E.; Spector, E.; et al. Standards and Guidelines for the Interpretation of Sequence Variants: A Joint Consensus Recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet. Med. Off. J. Am. Coll. Med. Genet. 2015, 17, 405–424. [Google Scholar] [CrossRef]
- Sucharski, H.C.; Dudley, E.K.; Keith, C.B.R.; El Refaey, M.; Koenig, S.N.; Mohler, P.J. Mechanisms and Alterations of Cardiac Ion Channels Leading to Disease: Role of Ankyrin-B in Cardiac Function. Biomolecules 2020, 10, 211. [Google Scholar] [CrossRef] [Green Version]
- Marketou, M.E.; Zareas, I.; Kanoupakis, E.; Patrianakos, A.; Parthenakis, F. A Case Series of Brugada Syndrome with a Novel Mutation in the Ankyrin-B Gene: An Unusual Unmasking in Acute Myocarditis. Eur. Heart J. Case Rep. 2021, 5, ytab225. [Google Scholar] [CrossRef] [PubMed]
- Giudicessi, J.R.; Ackerman, M.J. Established Loss-of-Function Variants in ANK2-Encoded Ankyrin-B Rarely Cause a Concerning Cardiac Phenotype in Humans. Circ. Genom. Precis. Med. 2020, 13, e002851. [Google Scholar] [CrossRef]
- Yasuda, H.; Mizuno, A.; Tamaoki, T.; Morinaga, T. ATBF1, a Multiple-Homeodomain Zinc Finger Protein, Selectively down-Regulates AT-Rich Elements of the Human Alpha-Fetoprotein Gene. Mol. Cell. Biol. 1994, 14, 1395–1401. [Google Scholar] [CrossRef] [PubMed]
- Edokobi, N.; Isom, L.L. Voltage-Gated Sodium Channel Β1/Β1B Subunits Regulate Cardiac Physiology and Pathophysiology. Front. Physiol. 2018, 9, 351. [Google Scholar] [CrossRef] [Green Version]
- Qu, Y.; Isom, L.L.; Westenbroek, R.E.; Rogers, J.C.; Tanada, T.N.; McCormick, K.A.; Scheuer, T.; Catterall, W.A. Modulation of Cardiac Na+ Channel Expression in Xenopus Oocytes by Beta 1 Subunits. J. Biol. Chem. 1995, 270, 25696–25701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Li, J.; Dong, F.N.; Dong, J.-T. Characterization of Nuclear Localization and SUMOylation of the ATBF1 Transcription Factor in Epithelial Cells. PLoS ONE 2014, 9, e92746. [Google Scholar] [CrossRef]
- Kao, Y.-H.; Hsu, J.-C.; Chen, Y.-C.; Lin, Y.-K.; Lkhagva, B.; Chen, S.-A.; Chen, Y.-J. ZFHX3 Knockdown Increases Arrhythmogenesis and Dysregulates Calcium Homeostasis in HL-1 Atrial Myocytes. Int. J. Cardiol. 2016, 210, 85–92. [Google Scholar] [CrossRef]
- Lkhagva, B.; Lin, Y.-K.; Chen, Y.-C.; Cheng, W.-L.; Higa, S.; Kao, Y.-H.; Chen, Y.-J. ZFHX3 Knockdown Dysregulates Mitochondrial Adaptations to Tachypacing in Atrial Myocytes through Enhanced Oxidative Stress and Calcium Overload. Acta Physiol. Oxf. Engl. 2021, 231, e13604. [Google Scholar] [CrossRef]
- van Ouwerkerk, A.F.; Hall, A.W.; Kadow, Z.A.; Lazarevic, S.; Reyat, J.S.; Tucker, N.R.; Nadadur, R.D.; Bosada, F.M.; Bianchi, V.; Ellinor, P.T.; et al. Epigenetic and Transcriptional Networks Underlying Atrial Fibrillation. Circ. Res. 2020, 127, 34–50. [Google Scholar] [CrossRef] [PubMed]
- Lozano-Velasco, E.; Hernández-Torres, F.; Daimi, H.; Serra, S.A.; Herraiz, A.; Hove-Madsen, L.; Aránega, A.; Franco, D. Pitx2 Impairs Calcium Handling in a Dose-Dependent Manner by Modulating Wnt Signalling. Cardiovasc. Res. 2016, 109, 55–66. [Google Scholar] [CrossRef] [Green Version]
- Hall, A.W.; Chaffin, M.; Roselli, C.; Lin, H.; Lubitz, S.A.; Bianchi, V.; Geeven, G.; Bedi, K.; Margulies, K.B.; de Laat, W.; et al. Epigenetic Analyses of Human Left Atrial Tissue Identifies Gene Networks Underlying Atrial Fibrillation. Circ. Genom. Precis. Med. 2020, 13, e003085. [Google Scholar] [CrossRef]
- Caballero, R.; Utrilla, R.G.; Amorós, I.; Matamoros, M.; Pérez-Hernández, M.; Tinaquero, D.; Alfayate, S.; Nieto-Marín, P.; Guerrero-Serna, G.; Liu, Q.-H.; et al. Tbx20 Controls the Expression of the KCNH2 Gene and of HERG Channels. Proc. Natl. Acad. Sci. USA 2017, 114, E416–E425. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.-T.; Hsieh, C.-S.; Chang, S.-N.; Chuang, E.Y.; Juang, J.-M.J.; Lin, L.-Y.; Lai, L.-P.; Hwang, J.-J.; Chiang, F.-T.; Lin, J.-L. Next-Generation Sequencing of Nine Atrial Fibrillation Candidate Genes Identified Novel de Novo Mutations in Patients with Extreme Trait of Atrial Fibrillation. J. Med. Genet. 2015, 52, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Rentzsch, P.; Witten, D.; Cooper, G.M.; Shendure, J.; Kircher, M. CADD: Predicting the Deleteriousness of Variants throughout the Human Genome. Nucleic Acids Res. 2019, 47, D886–D894. [Google Scholar] [CrossRef] [PubMed]
- Claycomb, W.C.; Lanson, N.A.; Stallworth, B.S.; Egeland, D.B.; Delcarpio, J.B.; Bahinski, A.; Izzo, N.J. HL-1 Cells: A Cardiac Muscle Cell Line That Contracts and Retains Phenotypic Characteristics of the Adult Cardiomyocyte. Proc. Natl. Acad. Sci. USA 1998, 95, 2979–2984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matamoros, M.; Pérez-Hernández, M.; Guerrero-Serna, G.; Amorós, I.; Barana, A.; Núñez, M.; Ponce-Balbuena, D.; Sacristán, S.; Gómez, R.; Tamargo, J.; et al. Nav1.5 N-Terminal Domain Binding to α1-Syntrophin Increases Membrane Density of Human Kir2.1, Kir2.2 and Nav1.5 Channels. Cardiovasc. Res. 2016, 110, 279–290. [Google Scholar] [CrossRef] [Green Version]
- Shy, D.; Gillet, L.; Ogrodnik, J.; Albesa, M.; Verkerkm, A.O.; Wolswinkelm, R.; Rougier, J.S.; Barc, J.; Shy, D.; Gillet, L.; et al. Cardiac sodium channel NaV1.5 distribution in myocytes via interacting proteins: The multiple pool model. Biochim. Biophys. Acta 2013, 1833, 886–894. [Google Scholar] [CrossRef] [Green Version]
- Utrilla, R.G.; Nieto-Marín, P.; Alfayate, S.; Tinaquero, D.; Matamoros, M.; Pérez-Hernández, M.; Sacristán, S.; Ondo, L.; de Andrés, R.; Díez-Guerra, F.J.; et al. Kir2.1-Nav1.5 Channel Complexes Are Differently Regulated than Kir2.1 and Nav1.5 Channels Alone. Front. Physiol. 2017, 8, 903. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Zfhx3 | τact (ms) | Vhact (mV) | kact | τfinact (ms) Afinact (%) | τsinact (ms) Asinact (%) | Vhinact (mV) | kinact | τreact (ms) |
|---|---|---|---|---|---|---|---|---|
| (–) | 0.22 ± 0.03 | −45.4 ± 1.9 | 5.1 ± 0.3 | 1.1 ± 0.05 85.2 ± 1.8 | 12.4 ± 1.6 14.8 ± 1.8 | −91.8 ± 2.2 | 5.0 ± 0.1 | 14.9 ± 4.3 |
| WT | 0.19 ± 0.01 | −42.6 ± 0.9 | 5.5 ± 0.2 | 1.2 ± 0.06 83.5 ± 1.8 | 10.8 ± 1.1 16.5 ± 1.8 | −91.7 ± 1.8 | 5.2 ± 0.2 | 12.9 ± 1.4 |
| p.V949I | 0.20 ± 0.02 | −40.8 ± 2.2 | 5.1 ± 0.3 | 1.1 ± 0.07 83.3 ± 1.8 | 11.7 ± 1.5 16.7 ± 1.8 | −90.6 ± 2.0 | 5.1 ± 0.2 | 15.0 ± 2.2 |
| p.M1260T | 0.24 ± 0.04 | −43.6 ± 2.3 | 5.3 ± 0.3 | 1.1 ± 0.1 80.9 ± 1.9 | 12.1 ± 1.1 19.1 ± 1.9 | −90.4 ± 3.7 | 5.0 ± 0.2 | 15.7 ± 2.5 |
| p.Q2564R | 0.19 ± 0.02 | −44.9 ± 1.6 | 5.3 ± 0.2 | 1.2 ± 0.08 85.9 ± 2.6 | 11.9 ± 1.3 14.1 ± 2.6 | −93.1 ± 1.8 | 5.2 ± 0.2 | 12.3 ± 1.1 |
| Scrambled | 0.18 ± 0.01 | −40.4 ± 1.5 | 5.2 ± 0.3 | 1.1 ± 0.1 85.0 ± 3.0 | 9.1 ± 0.8 15.0 ± 3.0 | −85.0 ± 2.4 | 5.0 ± 0.1 | 12.7 ± 2.9 |
| siZfhx3 | 0.18 ± 0.02 | −40.0 ± 1.9 | 5.1 ± 0.3 | 1.1 ± 0.05 88.1 ± 0.7 | 9.3 ± 1.2 11.9 ± 0.7 | −86.4 ± 1.9 | 5.1 ± 0.3 | 12.4 ± 3.1 |
| Proband | Gene | Genotype | Ancestral Allele /Variant | dbSNP_ID | MAF | Aminoacid Substitution | Transcript | Provean Prediction | SIFT Prediction | Polyphen Prediction | Mutation Taster | Mutation Assesor | LRT |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| BrS-1 | CACNA1B | HET | G/A | rs774297154 | 0.0000041 | R377Q | NM_000718.3 | Deleterious | Damaging | Benign | Benign | - | - |
| FLNB | HET | G/A | rs201369608 | 0.00007 | R1009Q | NM_001164317.1 | Neutral | Tolerated | Benign | Probably deleterious | Low | Neutral | |
| ZFHX3 | HET | G/A | rs113497421 | 0.002 | V949I | NM_006885.3 | Neutral | Tolerated | Probably damaging | Probably deleterious | Medium | Damaging | |
| BrS-2 | ANK2 | HET | A/G | D3126G | NM_001148.4 | Deleterious | Deleterious | Probably damaging | Deleterious | Medium | Damaging | ||
| ZFHX3 | HET | A/G | rs141564201 | 0.0007 | Q2564R | NM_006885.3 | Neutral | Tolerated | Unknown | Probably deleterious | Neutral | ||
| AF-1 | NEURL1 | HET | T/C | F35L | NM_004210.4 | Neutral | Tolerated | Benign | Probably deleterious | Low | Neutral | ||
| ZFHX3 | HET | A/G | rs777360037 | 0.000004 | M1260T | NM_006885.3 | Neutral | Tolerated | Benign | Probably deleterious | Low | Damaging | |
| AF-2 | ANK2 | HET | G/A | rs149963885 | 0.0006 | E3016K | NM_001148.4 | Neutral | Tolerated | Benign | Neutral | Low | Neutral |
| ZFHX3 | HET | A/G | rs777360037 | 0.000004 | M1260T | NM_006885.3 | Neutral | Tolerated | Benign | Probably deleterious | Low | Damaging |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rubio-Alarcón, M.; Cámara-Checa, A.; Dago, M.; Crespo-García, T.; Nieto-Marín, P.; Marín, M.; Merino, J.L.; Toquero, J.; Salguero-Bodes, R.; Tamargo, J.; et al. Zfhx3 Transcription Factor Represses the Expression of SCN5A Gene and Decreases Sodium Current Density (INa). Int. J. Mol. Sci. 2021, 22, 13031. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313031
Rubio-Alarcón M, Cámara-Checa A, Dago M, Crespo-García T, Nieto-Marín P, Marín M, Merino JL, Toquero J, Salguero-Bodes R, Tamargo J, et al. Zfhx3 Transcription Factor Represses the Expression of SCN5A Gene and Decreases Sodium Current Density (INa). International Journal of Molecular Sciences. 2021; 22(23):13031. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313031
Chicago/Turabian StyleRubio-Alarcón, Marcos, Anabel Cámara-Checa, María Dago, Teresa Crespo-García, Paloma Nieto-Marín, María Marín, José Luis Merino, Jorge Toquero, Rafael Salguero-Bodes, Juan Tamargo, and et al. 2021. "Zfhx3 Transcription Factor Represses the Expression of SCN5A Gene and Decreases Sodium Current Density (INa)" International Journal of Molecular Sciences 22, no. 23: 13031. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222313031