
SLC22 Transporters in the Fly Renal System Regulate Response to Oxidative Stress In Vivo

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
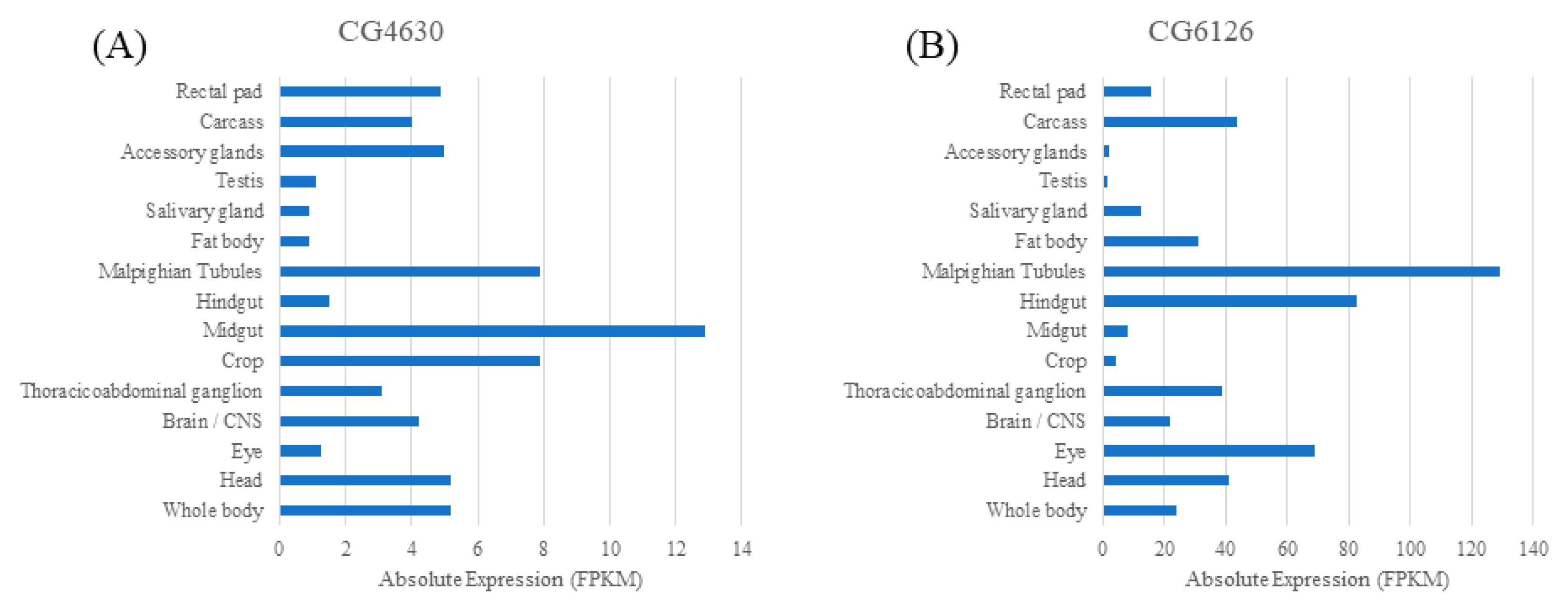
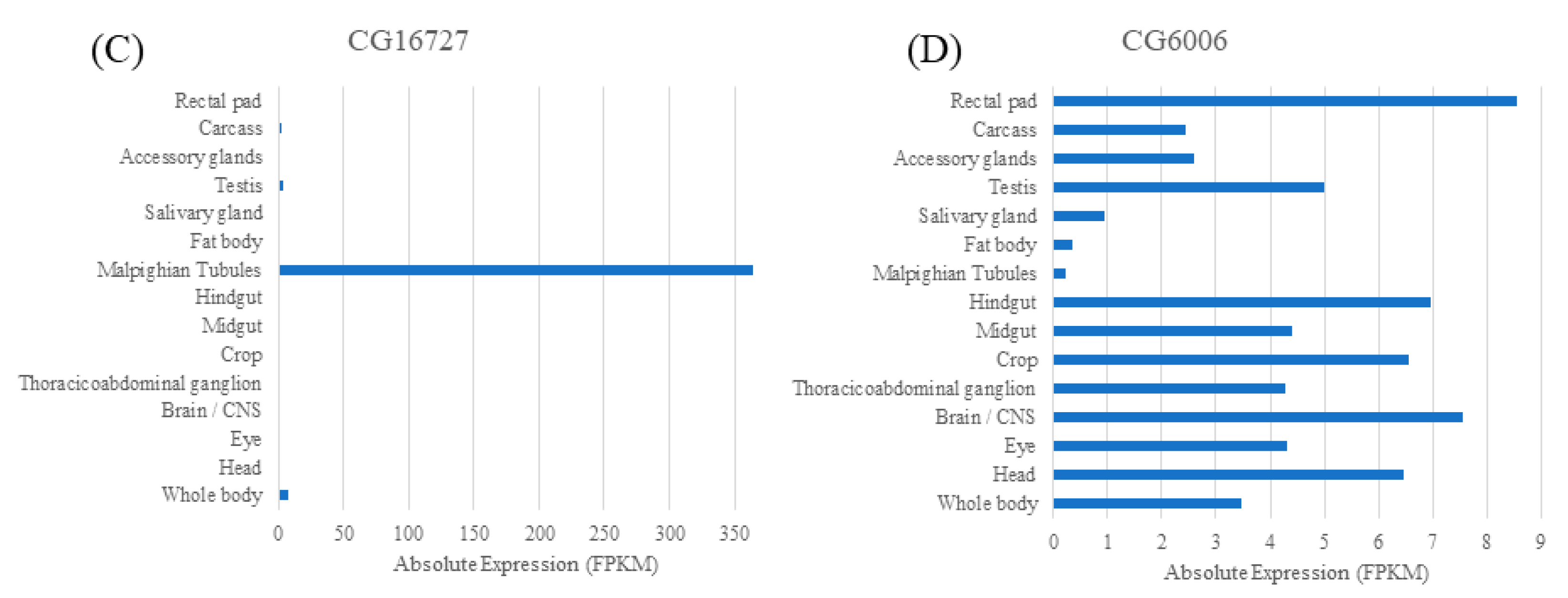
2.1. Human Relatives of the Four Fly SLC22 Transporters and Their Expression in Malpighian Tubule and Other Tissues
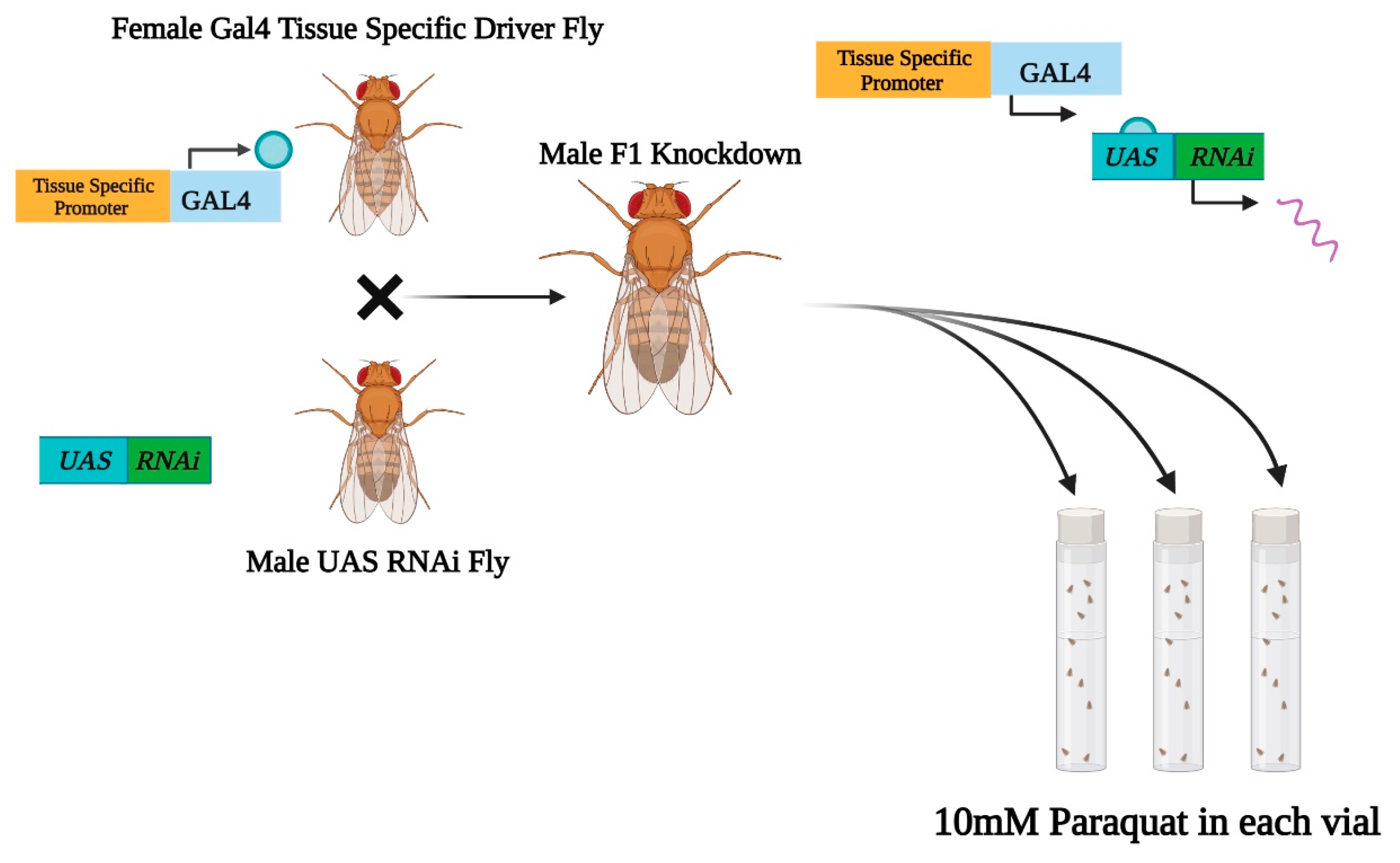
2.2. Tissue-Selective Knockdowns
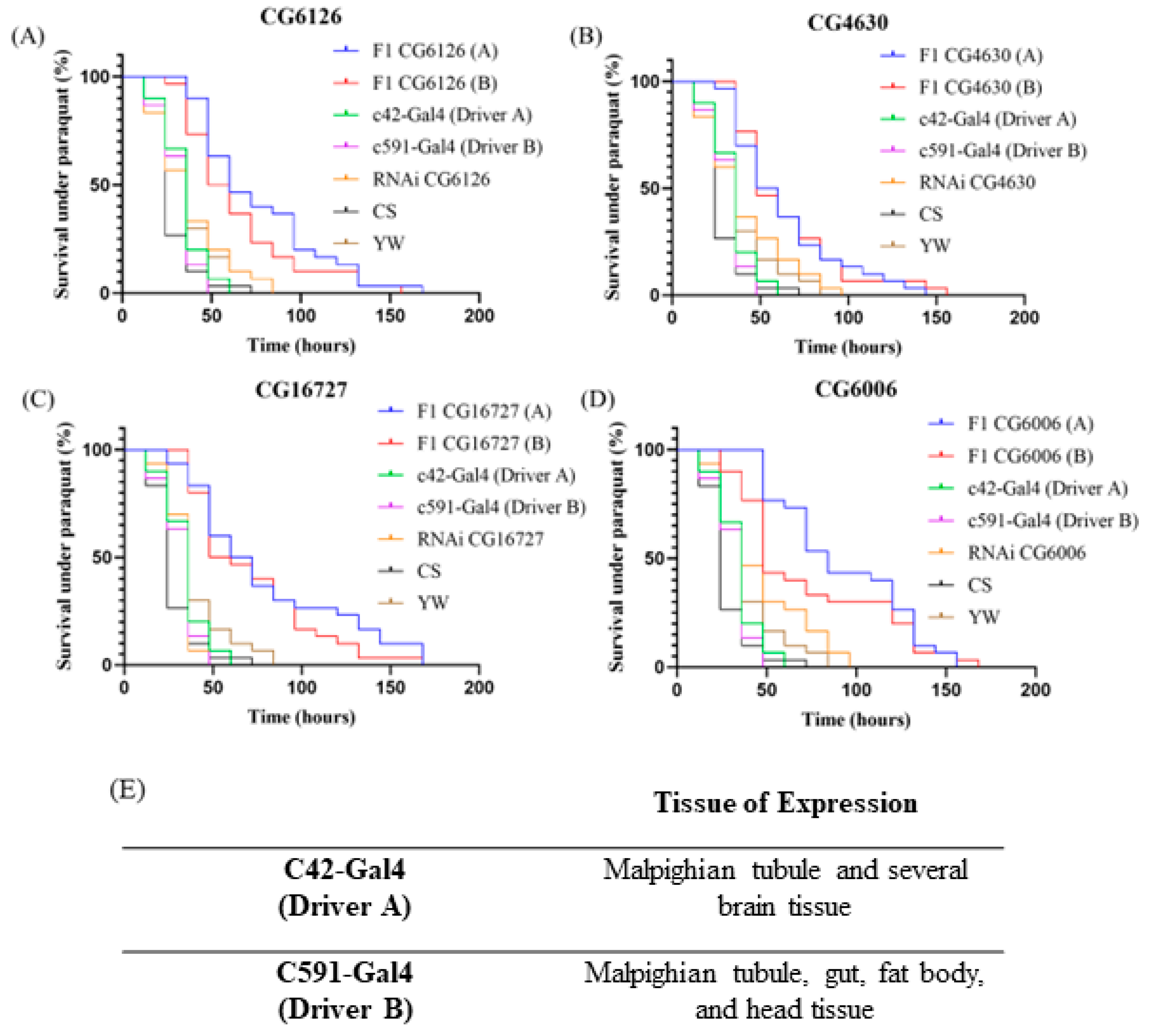
2.3. Long Term Survival of Tissue-Selective SLC22 Transporter Knockdowns under Severe Oxidative Stress
2.4. CG6126 (SLC22A16): Potential Involvement in the Transport of Ergothioneine and Carnitine
2.5. CG16727 (SLC22A7): Potential Regulator of Uric Acid
2.6. CG6006 (SLC22A1, A2, A3, A6, A7, A8, A11, A12, A13, and A14): Possible Role in Uric Acid Regulation
2.7. CG4630 (SLC22A3): Potential Role in Handling Carnitine Derivatives
3. Materials and Methods
3.1. Data Collection
3.2. Drosophila Strains and Genetics:
3.3. Paraquat Exposure of F1 RNAi Flies
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| SLC22 | Solute Carrier Family 22 |
| OAT | Organic Anion Transporter |
| OCTN | Organic Zwitterion (Carnitine) Transporter |
| OCT | Organic Cation transporter |
| FLIPT | Fly-Like Putative Transporter |
| WT | Wild-Type |
| URAT | Urate Transporter |
| CT | Carnitine Transporter |
| DME | Drug Metabolizing Enzyme |
| RSST | Remote Sensing and Signaling Theory |
| ADME | Administration, Distribution, Metabolism, and Excretion |
| GLK | Gut-Liver-Kidney |
| NKT | Novel Kidney Transporter |
| ROS | Reactive Oxygen Species |
| PQ | Paraquat |
| UAS | Upstream Activation Sequence |
| RNAi | Interfering RNA |
| DIOPT | Drosophila RNAi Screening Center Integrative Ortholog Prediction Tool |
| ABC | ATP-Binding Cassette |
| BDSC | Bloomington Drosophila Stock Center |
| YW | Yellow White |
| CS | Canton S |
| EGT | Ergothioneine |
References
- Yin, J.; Sun, W.; Li, F.; Hong, J.; Li, X.; Zhou, Y.; Lu, Y.; Liu, M.; Zhang, X.; Chen, N.; et al. VARIDT 1.0: Variability of Drug Transporter Database. Nucleic Acids Res. 2020, 48, D1042–D1050. [Google Scholar] [CrossRef]
- Ahn, S.-Y.; Nigam, S.K. Toward a Systems Level Understanding of Organic Anion and Other Multispecific Drug Transporters: A Remote Sensing and Signaling Hypothesis. Mol. Pharmacol. 2009, 76, 481–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenthal, S.B.; Bush, K.T.; Nigam, S.K. A Network of SLC and ABC Transporter and DME Genes Involved in Remote Sensing and Signaling in the Gut-Liver-Kidney Axis. Sci. Rep. 2019, 9, 11879. [Google Scholar] [CrossRef] [Green Version]
- Nigam, S.K. What Do Drug Transporters Really Do? Nat. Rev. Drug Discov. 2015, 14, 29–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nigam, S.K.; Bush, K.T.; Bhatnagar, V.; Poloyac, S.M.; Momper, J.D. The Systems Biology of Drug Metabolizing Enzymes and Transporters: Relevance to Quantitative Systems Pharmacology. Clin. Pharmacol. Ther. 2020, 108, 40–53. [Google Scholar] [CrossRef]
- Engelhart, D.C.; Granados, J.C.; Shi, D.; Saier, M.H., Jr.; Baker, M.E.; Abagyan, R.; Nigam, S.K. Systems Biology Analysis Reveals Eight SLC22 Transporter Subgroups, Including OATs, OCTs, and OCTNs. Int. J. Mol. Sci. 2020, 21, 1791. [Google Scholar] [CrossRef] [Green Version]
- Nigam, S.K. The SLC22 Transporter Family: A Paradigm for the Impact of Drug Transporters on Metabolic Pathways, Signaling, and Disease. Annu. Rev. Pharmacol. Toxicol. 2018, 58, 663–687. [Google Scholar] [CrossRef]
- Koepsell, H. Organic Cation Transporters in Health and Disease. Pharmacol. Rev. 2020, 72, 253–319. [Google Scholar] [CrossRef] [PubMed]
- Lozano, E.; Briz, O.; Macias, R.I.R.; Serrano, M.A.; Marin, J.J.G.; Herraez, E. Genetic Heterogeneity of SLC22 Family of Transporters in Drug Disposition. J. Pers. Med. 2018, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.C.; Goldenberg, A.; Chen, Y.; Lun, C.; Wu, W.; Bush, K.T.; Balac, N.; Rodriguez, P.; Abagyan, R.; Nigam, S.K. Molecular Properties of Drugs Interacting with SLC22 Transporters OAT1, OAT3, OCT1, and OCT2: A Machine-Learning Approach. J. Pharmacol. Exp. Ther. 2016, 359, 215–229. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Nieto, C.E.; You, G.; Bush, K.T.; Barros, E.J.; Beier, D.R.; Nigam, S.K. Molecular Cloning and Characterization of NKT, a Gene Product Related to the Organic Cation Transporter Family That Is Almost Exclusively Expressed in the Kidney. J. Biol. Chem. 1997, 272, 6471–6478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lepist, E.-I.; Ray, A.S. Beyond Drug-Drug Interactions: Effects of Transporter Inhibition on Endobiotics, Nutrients and Toxins. Expert Opin. Drug Metab. Toxicol. 2017, 13, 1075–1087. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Yee, S.W.; Chien, H.-C.; Chen, E.C.; Luo, Q.; Zou, L.; Piao, M.; Mifune, A.; Chen, L.; Calvert, M.E.; et al. Organic Cation Transporter 1 (OCT1) Modulates Multiple Cardiometabolic Traits through Effects on Hepatic Thiamine Content. PLoS Biol. 2018, 16, e2002907. [Google Scholar] [CrossRef] [Green Version]
- Jansen, J.; Jansen, K.; Neven, E.; Poesen, R.; Othman, A.; van Mil, A.; Sluijter, J.; Sastre Torano, J.; Zaal, E.A.; Berkers, C.R.; et al. Remote Sensing and Signaling in Kidney Proximal Tubules Stimulates Gut Microbiome-Derived Organic Anion Secretion. Proc. Natl. Acad. Sci. USA 2019, 116, 16105–16110. [Google Scholar] [CrossRef] [Green Version]
- Vallon, V.; Eraly, S.A.; Wikoff, W.R.; Rieg, T.; Kaler, G.; Truong, D.M.; Ahn, S.-Y.; Mahapatra, N.R.; Mahata, S.K.; Gangoiti, J.A.; et al. Organic Anion Transporter 3 Contributes to the Regulation of Blood Pressure. J. Am. Soc. Nephrol. JASN 2008, 19, 1732–1740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granados, J.C.; Nigam, A.K.; Bush, K.T.; Jamshidi, N.; Nigam, S.K. A Key Role for the Transporter OAT1 in Systemic Lipid Metabolism. J. Biol. Chem. 2021, 296, 100603. [Google Scholar] [CrossRef]
- Granados, J.C.; Richelle, A.; Gutierrez, J.M.; Zhang, P.; Zhang, X.; Bhatnagar, V.; Lewis, N.E.; Nigam, S.K. Coordinate Regulation of Systemic and Kidney Tryptophan Metabolism by the Drug Transporters OAT1 and OAT3. J. Biol. Chem. 2021, 296, 100575. [Google Scholar] [CrossRef]
- Wang, Y.; Moussian, B.; Schaeffeler, E.; Schwab, M.; Nies, A.T. The Fruit Fly Drosophila Melanogaster as an Innovative Preclinical ADME Model for Solute Carrier Membrane Transporters, with Consequences for Pharmacology and Drug Therapy. Drug Discov. Today 2018, 23, 1746–1760. [Google Scholar] [CrossRef]
- Cohen, E.; Sawyer, J.K.; Peterson, N.G.; Dow, J.A.T.; Fox, D.T. Physiology, Development, and Disease Modeling in the Drosophila Excretory System. Genetics 2020, 214, 235–264. [Google Scholar] [CrossRef] [Green Version]
- Weavers, H.; Prieto-Sánchez, S.; Grawe, F.; Garcia-López, A.; Artero, R.; Wilsch-Bräuninger, M.; Ruiz-Gómez, M.; Skaer, H.; Denholm, B. The Insect Nephrocyte Is a Podocyte-like Cell with a Filtration Slit Diaphragm. Nature 2009, 457, 322–326. [Google Scholar] [CrossRef] [Green Version]
- Apidianakis, Y.; Rahme, L.G. Drosophila Melanogaster as a Model for Human Intestinal Infection and Pathology. Dis. Model. Mech. 2011, 4, 21–30. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, D.; Deng, W.-M. Drosophila Model in Cancer: An Introduction. In The Drosophila Model in Cancer; Springer: Cham, Switzerland, 2019; Volume 1167, pp. 1–14. [Google Scholar] [CrossRef]
- Musselman, L.P.; Kühnlein, R.P. Drosophila as a Model to Study Obesity and Metabolic Disease. J. Exp. Biol. 2018, 221 (Suppl. S1), jeb163881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farahani, R.; Haddad, G.G. Understanding the Molecular Responses to Hypoxia Using Drosophila as a Genetic Model. Respir. Physiol. Neurobiol. 2003, 135, 221–229. [Google Scholar] [CrossRef]
- Zhou, D.; Haddad, G.G. Genetic Analysis of Hypoxia Tolerance and Susceptibility in Drosophila and Humans. Annu. Rev. Genomics Hum. Genet. 2013, 14, 25–43. [Google Scholar] [CrossRef] [PubMed]
- Zhao, H.W.; Haddad, G.G. Review: Hypoxic and Oxidative Stress Resistance in Drosophila Melanogaster. Placenta 2011, 32 (Suppl. S2), S104–S108. [Google Scholar] [CrossRef] [Green Version]
- Engelhart, D.C.; Azad, P.; Ali, S.; Granados, J.C.; Haddad, G.G.; Nigam, S.K. Drosophila SLC22 Orthologs Related to OATs, OCTs, and OCTNs Regulate Development and Responsiveness to Oxidative Stress. Int. J. Mol. Sci. 2020, 21, 2002. [Google Scholar] [CrossRef] [Green Version]
- Cohen, G.M.; d’Arcy Doherty, M. Free Radical Mediated Cell Toxicity by Redox Cycling Chemicals. Br. J. Cancer. Suppl. 1987, 8, 46–52. [Google Scholar]
- Pochini, L.; Galluccio, M.; Scalise, M.; Console, L.; Indiveri, C. OCTN: A Small Transporter Subfamily with Great Relevance to Human Pathophysiology, Drug Discovery, and Diagnostics. SLAS Discov. Adv. Life Sci. R&D 2019, 24, 89–110. [Google Scholar] [CrossRef]
- Denholm, B.; Skaer, H. Bringing Together Components of the Fly Renal System. Curr. Opin. Genet. Dev. 2009, 19, 526–532. [Google Scholar] [CrossRef] [Green Version]
- Fan, Q.-X.; Gong, S.-Q.; Hong, X.-Z.; Feng, X.-M.; Zhang, F.-J. Clinical-Grade Garcinia Cambogia Extract Dissolves Calcium Oxalate Crystals in Drosophila Kidney Stone Models. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 6434–6445. [Google Scholar] [CrossRef]
- Ceder, M.M.; Aggarwal, T.; Hosseini, K.; Maturi, V.; Patil, S.; Perland, E.; Williams, M.J.; Fredriksson, R. CG4928 Is Vital for Renal Function in Fruit Flies and Membrane Potential in Cells: A First In-Depth Characterization of the Putative Solute Carrier UNC93A. Front. Cell Dev. Biol. 2020, 8, 580291. [Google Scholar] [CrossRef]
- Manseau, L.; Baradaran, A.; Brower, D.; Budhu, A.; Elefant, F.; Phan, H.; Philp, A.V.; Yang, M.; Glover, D.; Kaiser, K.; et al. GAL4 Enhancer Traps Expressed in the Embryo, Larval Brain, Imaginal Discs, and Ovary of Drosophila. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 1997, 209, 310–322. [Google Scholar] [CrossRef]
- Amin, A.; Li, Y.; Finkelstein, R. Hedgehog Activates the EGF Receptor Pathway during Drosophila Head Development. Development 1999, 126, 2623–2630. [Google Scholar] [CrossRef]
- Harrison, D.A.; Binari, R.; Nahreini, T.S.; Gilman, M.; Perrimon, N. Activation of a Drosophila Janus Kinase (JAK) Causes Hematopoietic Neoplasia and Developmental Defects. EMBO J. 1995, 14, 2857–2865. [Google Scholar] [CrossRef]
- Jourde-Chiche, N.; Burtey, S. Accumulation of Protein-Bound Uremic Toxins: The Kidney Remains the Leading Culprit in the Gut-Liver-Kidney Axis. Kidney Int. 2020, 97, 1102–1104. [Google Scholar] [CrossRef]
- Eraly, S.A.; Monte, J.C.; Nigam, S.K. Novel SLC22 Transporter Homologs in Fly, Worm, and Human Clarify the Phylogeny of Organic Anion and Cation Transporters. Physiol. Genom. 2004, 18, 12–24. [Google Scholar] [CrossRef]
- Zhu, C.; Nigam, K.B.; Date, R.C.; Bush, K.T.; Springer, S.A.; Saier, M.H.; Wu, W.; Nigam, S.K. Evolutionary Analysis and Classification of OATs, OCTs, OCTNs, and Other SLC22 Transporters: Structure-Function Implications and Analysis of Sequence Motifs. PLoS ONE 2015, 10, e0140569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Jamshidi, N.; Eraly, S.A.; Liu, H.C.; Bush, K.T.; Palsson, B.O.; Nigam, S.K. Multispecific Drug Transporter Slc22a8 (Oat3) Regulates Multiple Metabolic and Signaling Pathways. Drug Metab. Dispos. Biol. Fate Chem. 2013, 41, 1825–1834. [Google Scholar] [CrossRef] [Green Version]
- Ahn, S.-Y.; Jamshidi, N.; Mo, M.L.; Wu, W.; Eraly, S.A.; Dnyanmote, A.; Bush, K.T.; Gallegos, T.F.; Sweet, D.H.; Palsson, B.Ø.; et al. Linkage of Organic Anion Transporter-1 to Metabolic Pathways through Integrated “Omics”-Driven Network and Functional Analysis. J. Biol. Chem. 2011, 286, 31522–31531. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Flockhart, I.; Vinayagam, A.; Bergwitz, C.; Berger, B.; Perrimon, N.; Mohr, S.E. An Integrative Approach to Ortholog Prediction for Disease-Focused and Other Functional Studies. BMC Bioinform. 2011, 12, 357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enomoto, A.; Wempe, M.F.; Tsuchida, H.; Shin, H.J.; Cha, S.H.; Anzai, N.; Goto, A.; Sakamoto, A.; Niwa, T.; Kanai, Y.; et al. Molecular Identification of a Novel Carnitine Transporter Specific to Human Testis. Insights into the Mechanism of Carnitine Recognition. J. Biol. Chem. 2002, 277, 36262–36271. [Google Scholar] [CrossRef] [Green Version]
- Ohashi, R.; Tamai, I.; Yabuuchi, H.; Nezu, J.I.; Oku, A.; Sai, Y.; Shimane, M.; Tsuji, A. Na(+)-Dependent Carnitine Transport by Organic Cation Transporter (OCTN2): Its Pharmacological and Toxicological Relevance. J. Pharmacol. Exp. Ther. 1999, 291, 778–784. [Google Scholar]
- Köttgen, A.; Albrecht, E.; Teumer, A.; Vitart, V.; Krumsiek, J.; Hundertmark, C.; Pistis, G.; Ruggiero, D.; O’Seaghdha, C.M.; Haller, T.; et al. Genome-Wide Association Analyses Identify 18 New Loci Associated with Serum Urate Concentrations. Nat. Genet. 2013, 45, 145–154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Using FlyBase, a Database of Drosophila Genes & Genomes. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/pmc/articles/PMC5107610/ (accessed on 30 September 2021).
- Raj, D.; Tomar, B.; Lahiri, A.; Mulay, S.R. The Gut-Liver-Kidney Axis: Novel Regulator of Fatty Liver Associated Chronic Kidney Disease. Pharmacol. Res. 2020, 152, 104617. [Google Scholar] [CrossRef]
- Bush, K.T.; Wu, W.; Lun, C.; Nigam, S.K. The Drug Transporter OAT3 (SLC22A8) and Endogenous Metabolite Communication via the Gut-Liver-Kidney Axis. J. Biol. Chem. 2017, 292, 15789–15803. [Google Scholar] [CrossRef] [Green Version]
- Ali, S.N.; Dayarathna, T.K.; Ali, A.N.; Osumah, T.; Ahmed, M.; Cooper, T.T.; Power, N.E.; Zhang, D.; Kim, D.; Kim, R.; et al. Drosophila Melanogaster as a Function-Based High-Throughput Screening Model for Antinephrolithiasis Agents in Kidney Stone Patients. Dis. Model. Mech. 2018, 11, dmm035873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gamberi, C.; Hipfner, D.R.; Trudel, M.; Lubell, W.D. Bicaudal C Mutation Causes Myc and TOR Pathway Up-Regulation and Polycystic Kidney Disease-like Phenotypes in Drosophila. PLoS Genet. 2017, 13, e1006694. [Google Scholar] [CrossRef] [Green Version]
- Caygill, E.E.; Brand, A.H. The GAL4 System: A Versatile System for the Manipulation and Analysis of Gene Expression. In Methods in Molecular Biology; Humana Press: New York, NY, USA, 2016; Volume 1478, pp. 33–52. [Google Scholar] [CrossRef]
- Bus, J.S.; Gibson, J.E. Paraquat: Model for Oxidant-Initiated Toxicity. Environ. Health Perspect. 1984, 55, 37–46. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Lu-Bo, Y.; Haddad, G.G. A Drosophila ABC Transporter Regulates Lifespan. PLoS Genet. 2014, 10, e1004844. [Google Scholar] [CrossRef] [Green Version]
- Azad, P.; Ryu, J.; Haddad, G.G. Distinct Role of Hsp70 in Drosophila Hemocytes during Severe Hypoxia. Free Radic. Biol. Med. 2011, 51, 530–538. [Google Scholar] [CrossRef] [Green Version]
- Arking, R.; Buck, S.; Berrios, A.; Dwyer, S.; Baker, G.T., III. Elevated Paraquat Resistance Can Be Used as a Bioassay for Longevity in a Genetically Based Long-Lived Strain of Drosophila. Dev. Genet. 1991, 12, 362–370. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Rio, D.C.; Haddad, G.G.; Ma, E. Regulatory Role of DADAR in ROS Metabolism in Drosophila CNS. Mol. Brain Res. 2004, 131, 93–100. [Google Scholar] [CrossRef] [PubMed]
- McCormack, A.L.; Atienza, J.G.; Johnston, L.C.; Andersen, J.K.; Vu, S.; Di Monte, D.A. Role of Oxidative Stress in Paraquat-Induced Dopaminergic Cell Degeneration. J. Neurochem. 2005, 93, 1030–1037. [Google Scholar] [CrossRef] [PubMed]
- Bus, J.; Aust, S.; Gibson, J. Paraquat Toxicity: Proposed Mechanism of Action Involving Lipid Peroxidation. Environ. Health Perspect. 1976, 16, 139–146. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.-Y.; Fauman, E.B.; Petersen, A.-K.; Krumsiek, J.; Santos, R.; Huang, J.; Arnold, M.; Erte, I.; Forgetta, V.; Yang, T.-P.; et al. An Atlas of Genetic Influences on Human Blood Metabolites. Nat. Genet. 2014, 46, 543–550. [Google Scholar] [CrossRef] [Green Version]
- Yee, S.W.; Buitrago, D.; Stecula, A.; Ngo, H.X.; Chien, H.-C.; Zou, L.; Koleske, M.L.; Giacomini, K.M. Deorphaning a Solute Carrier 22 Family Member, SLC22A15, through Functional Genomic Studies. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2020, 34, 15734–15752. [Google Scholar] [CrossRef]
- Carillo, M.R.; Bertapelle, C.; Scialò, F.; Siervo, M.; Spagnuolo, G.; Simeone, M.; Peluso, G.; Digilio, F.A. L-Carnitine in Drosophila: A Review. Antioxidants 2020, 9, 1310. [Google Scholar] [CrossRef]
- Gülçin, I. Antioxidant and Antiradical Activities of L-Carnitine. Life Sci. 2006, 78, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Ribas, G.S.; Vargas, C.R.; Wajner, M. L-Carnitine Supplementation as a Potential Antioxidant Therapy for Inherited Neurometabolic Disorders. Gene 2014, 533, 469–476. [Google Scholar] [CrossRef]
- Maxwell, S.R.; Thomason, H.; Sandler, D.; Leguen, C.; Baxter, M.A.; Thorpe, G.H.; Jones, A.F.; Barnett, A.H. Antioxidant Status in Patients with Uncomplicated Insulin-Dependent and Non-Insulin-Dependent Diabetes Mellitus. Eur. J. Clin. Investig. 1997, 27, 484–490. [Google Scholar] [CrossRef]
- Paithankar, J.G.; Kudva, A.K.; Raghu, S.V.; Patil, R.K. Radioprotective Role of Uric Acid: Evidence from Studies in Drosophila and Human Dermal Fibroblast Cells. Mol. Biol. Rep. 2020, 47, 2427–2436. [Google Scholar] [CrossRef]
- Miguel-Aliaga, I.; Jasper, H.; Lemaitre, B. Anatomy and Physiology of the Digestive Tract of Drosophila Melanogaster. Genetics 2018, 210, 357–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perkins, L.A.; Holderbaum, L.; Tao, R.; Hu, Y.; Sopko, R.; McCall, K.; Yang-Zhou, D.; Flockhart, I.; Binari, R.; Shim, H.-S.; et al. The Transgenic RNAi Project at Harvard Medical School: Resources and Validation. Genetics 2015, 201, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Piccin, A.; Salameh, A.; Benna, C.; Sandrelli, F.; Mazzotta, G.; Zordan, M.; Rosato, E.; Kyriacou, C.P.; Costa, R. Efficient and Heritable Functional Knock-out of an Adult Phenotype in Drosophila Using a GAL4-Driven Hairpin RNA Incorporating a Heterologous Spacer. Nucleic Acids Res. 2001, 29, e55. [Google Scholar] [CrossRef] [Green Version]
- Leader, D.P.; Krause, S.A.; Pandit, A.; Davies, S.A.; Dow, J.A.T. FlyAtlas 2: A New Version of the Drosophila Melanogaster Expression Atlas with RNA-Seq, MiRNA-Seq and Sex-Specific Data. Nucleic Acids Res. 2018, 46, D809–D815. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Related SLC22 Human Subgroups | SLC22 Subgroup Members | Drosophila SLC22 | SLC22 Human Ortholog | Likely Transported Antioxidants (Mammals) | Drosophila Tissue Expression | ||
|---|---|---|---|---|---|---|---|
| Gut | Fat Body | Malpighian Tubule | |||||
| OCTN/OCTN-related | A4, A5, A15, A16, A21 | CG6126 | A16 | Carnitine, carnitine derivatives, and EGT | ✓ | ✓ | ✓ |
| OATS2 | A7 | CG16727 | A7 | Uric acid | n/a | n/a | ✓ |
| OCT | A1, A2, A3 | CG6006 | A1, A2, A3 | Carnitine derivatives | ✓ | ✓ | ✓(L) |
| OATS1 | A6, A8, A20 | A6, A8 | Uric acid | ||||
| OATS2 | A7 | A7 | Uric acid | ||||
| OATS3 | A11, A12, A22 | A11, A12 | Uric acid | ||||
| OAT-like | A13, A14 | A13, A14 * | Uric acid | ||||
| OCT | A1, A2, A3 | CG4630 | A3 | Carnitine derivatives | ✓ | ✓(L) | ✓ |
| F1 vs. RNAI | F1 vs. Driver | F1 vs. CS | F1 vs. YW | |
|---|---|---|---|---|
| F1 CG6126 (A) | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| F1 CG4630 (A) | 0.0031 | <0.0001 | <0.0001 | 0.0003 |
| F1 CG16727 (A) | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| F1 CG6006 (A) | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| F1 CG6126 (B) | 0.0003 | <0.0001 | <0.0001 | 0.0002 |
| F1 CG4630 (B) | 0.0023 | <0.0001 | <0.0001 | 0.0001 |
| F1 CG16727 (B) | <0.0001 | <0.0001 | <0.0001 | <0.0001 |
| F1 CG6006 (B) | 0.0022 | <0.0001 | <0.0001 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, P.; Azad, P.; Engelhart, D.C.; Haddad, G.G.; Nigam, S.K. SLC22 Transporters in the Fly Renal System Regulate Response to Oxidative Stress In Vivo. Int. J. Mol. Sci. 2021, 22, 13407. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413407
Zhang P, Azad P, Engelhart DC, Haddad GG, Nigam SK. SLC22 Transporters in the Fly Renal System Regulate Response to Oxidative Stress In Vivo. International Journal of Molecular Sciences. 2021; 22(24):13407. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413407
Chicago/Turabian StyleZhang, Patrick, Priti Azad, Darcy C. Engelhart, Gabriel G. Haddad, and Sanjay K. Nigam. 2021. "SLC22 Transporters in the Fly Renal System Regulate Response to Oxidative Stress In Vivo" International Journal of Molecular Sciences 22, no. 24: 13407. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413407