
Role of Organic Cation Transporter 3 and Plasma Membrane Monoamine Transporter in the Rewarding Properties and Locomotor Sensitizing Effects of Amphetamine in Male andFemale Mice


Abstract
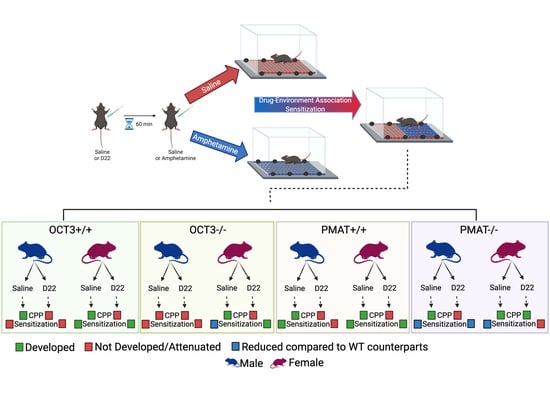
:
1. Introduction
2. Results
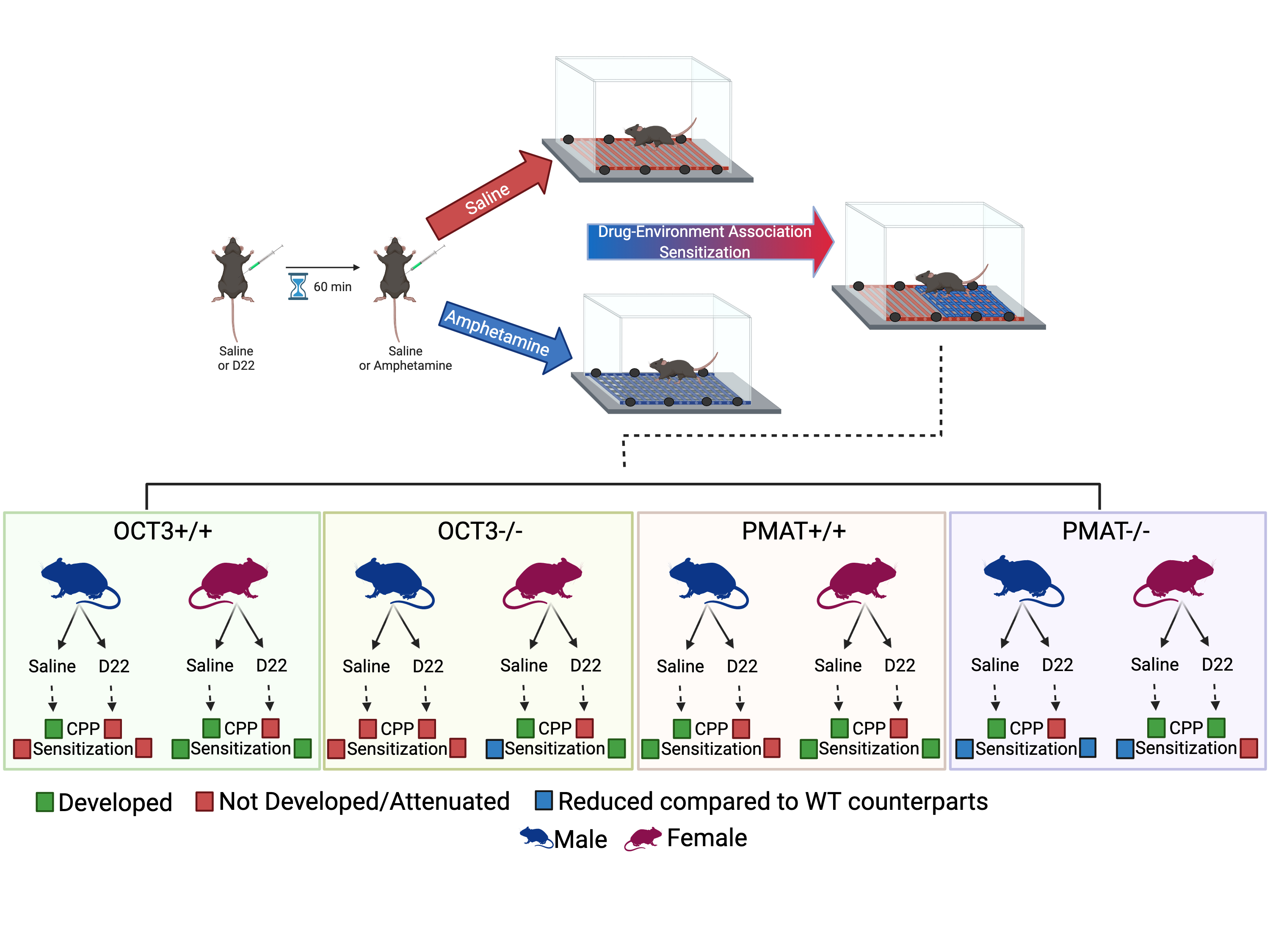
2.1. CPP
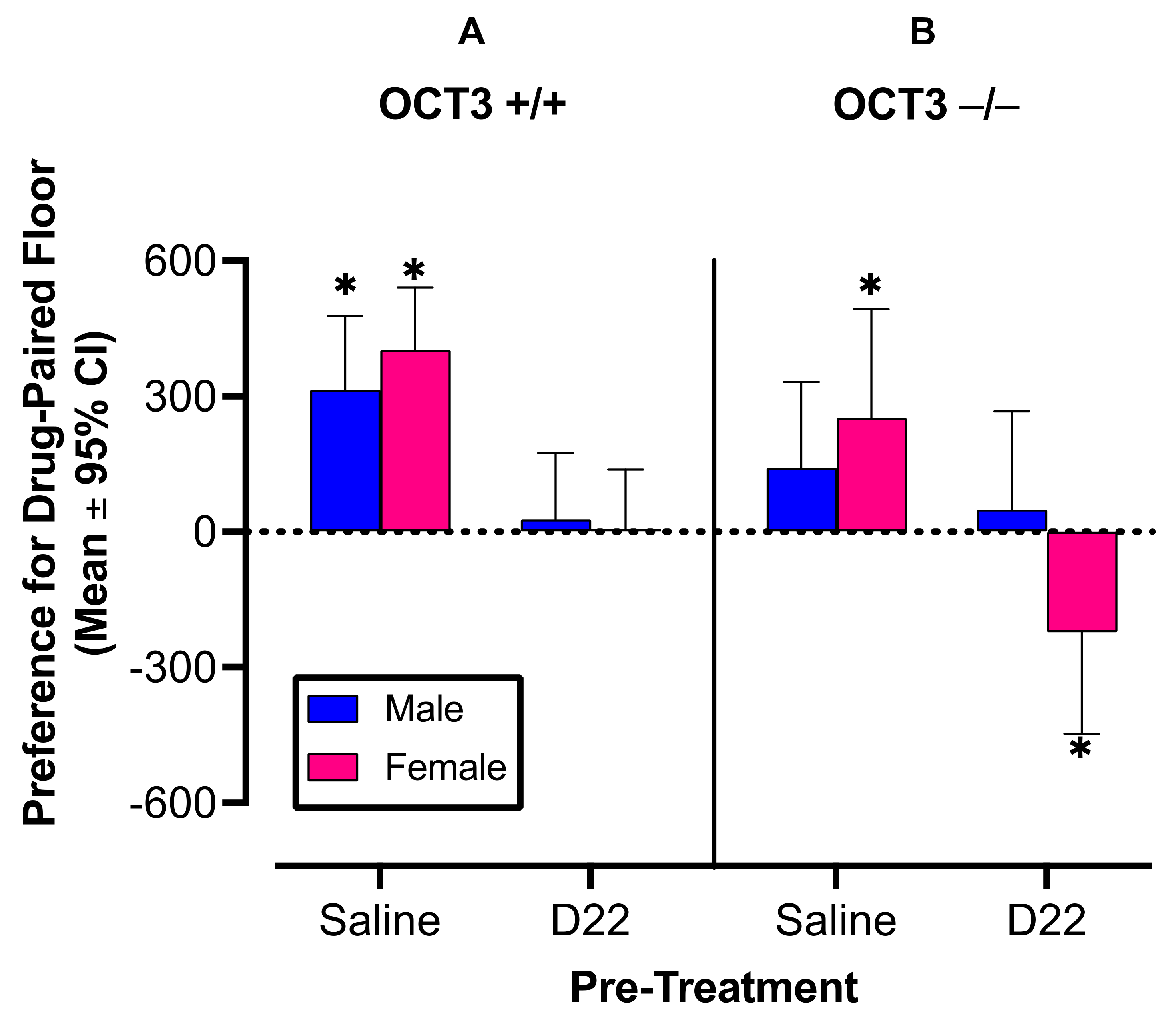
2.1.1. OCT3
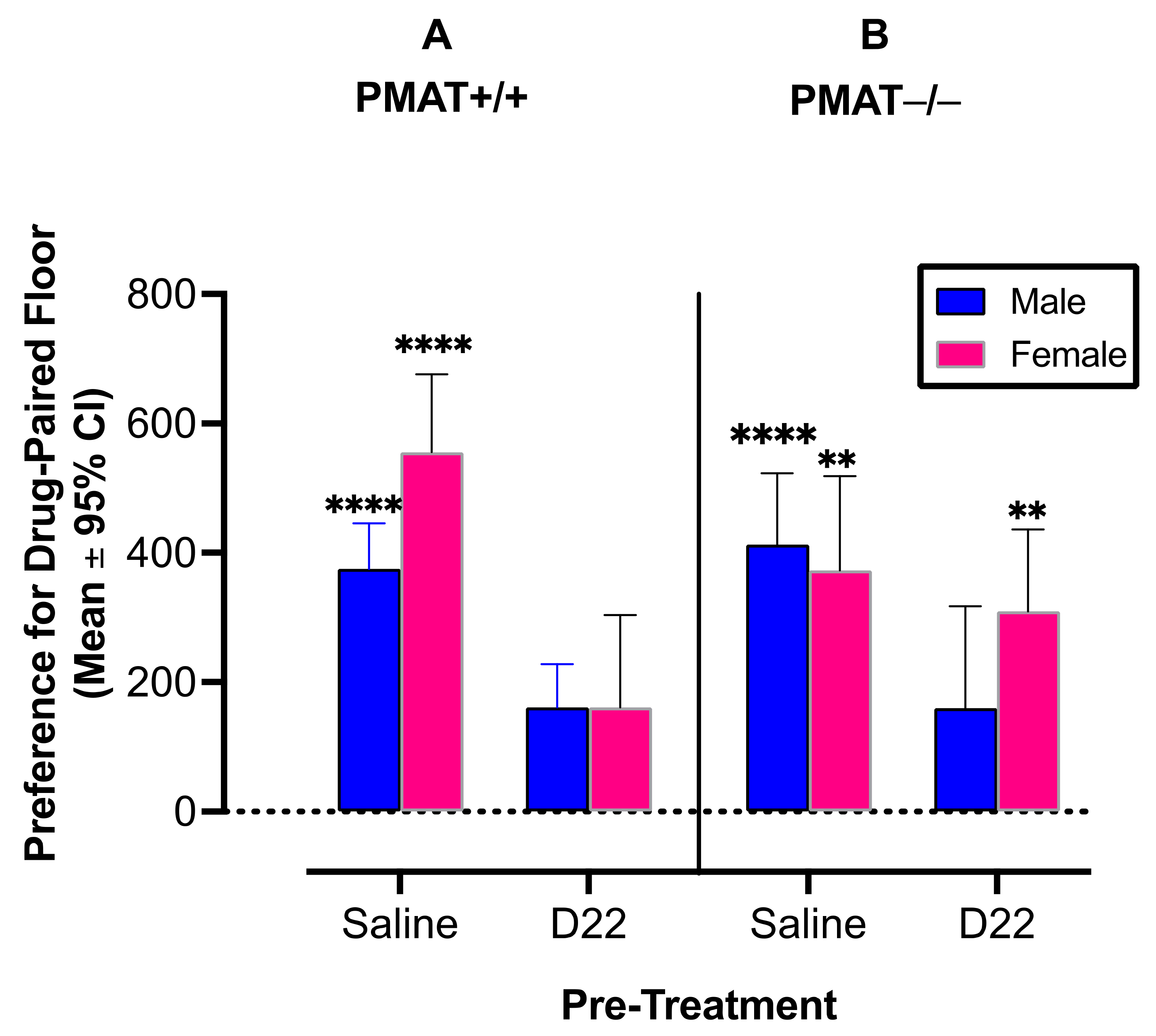
2.1.2. PMAT
2.2. Locomotor Activity and Sensitization
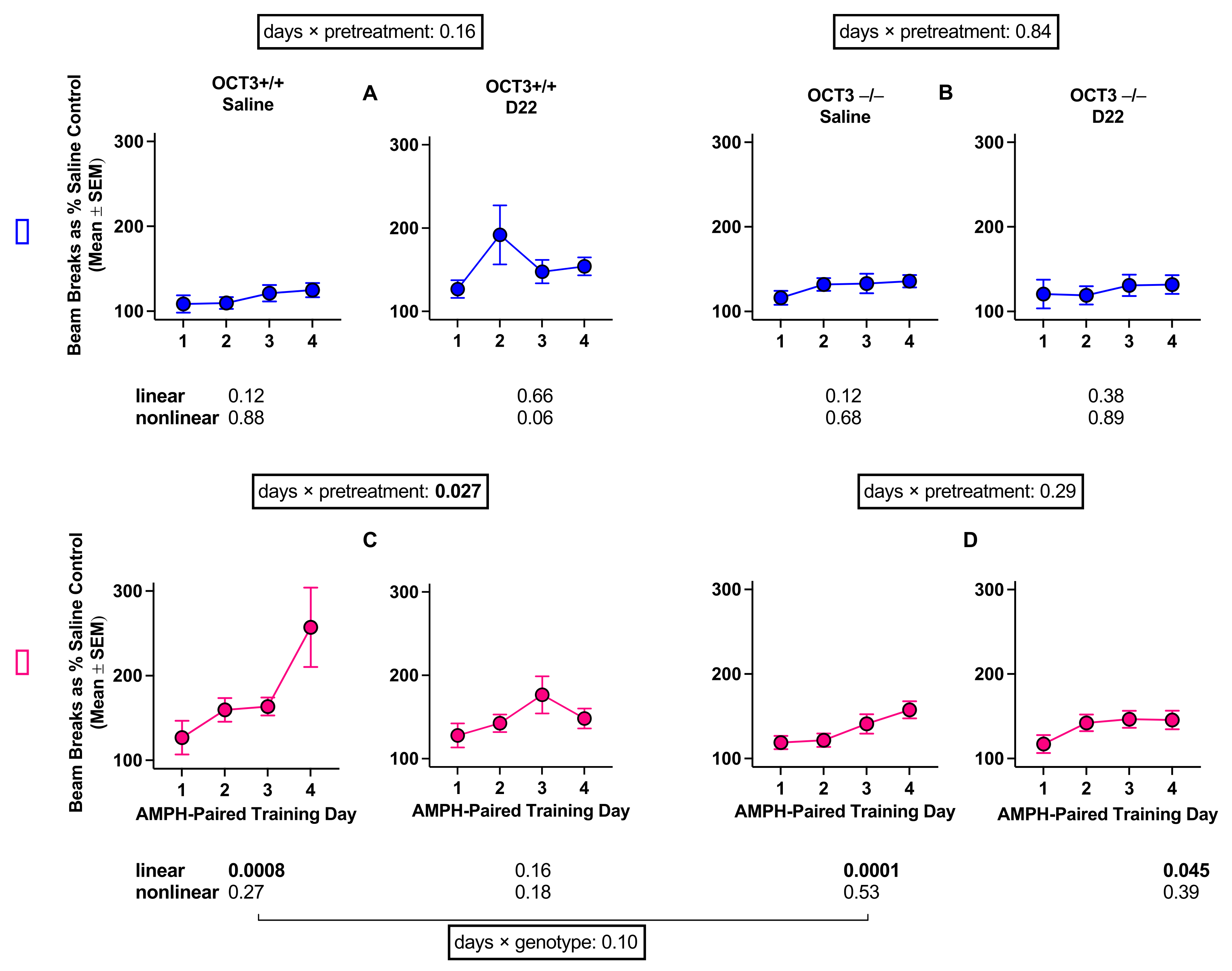
2.2.1. OCT3
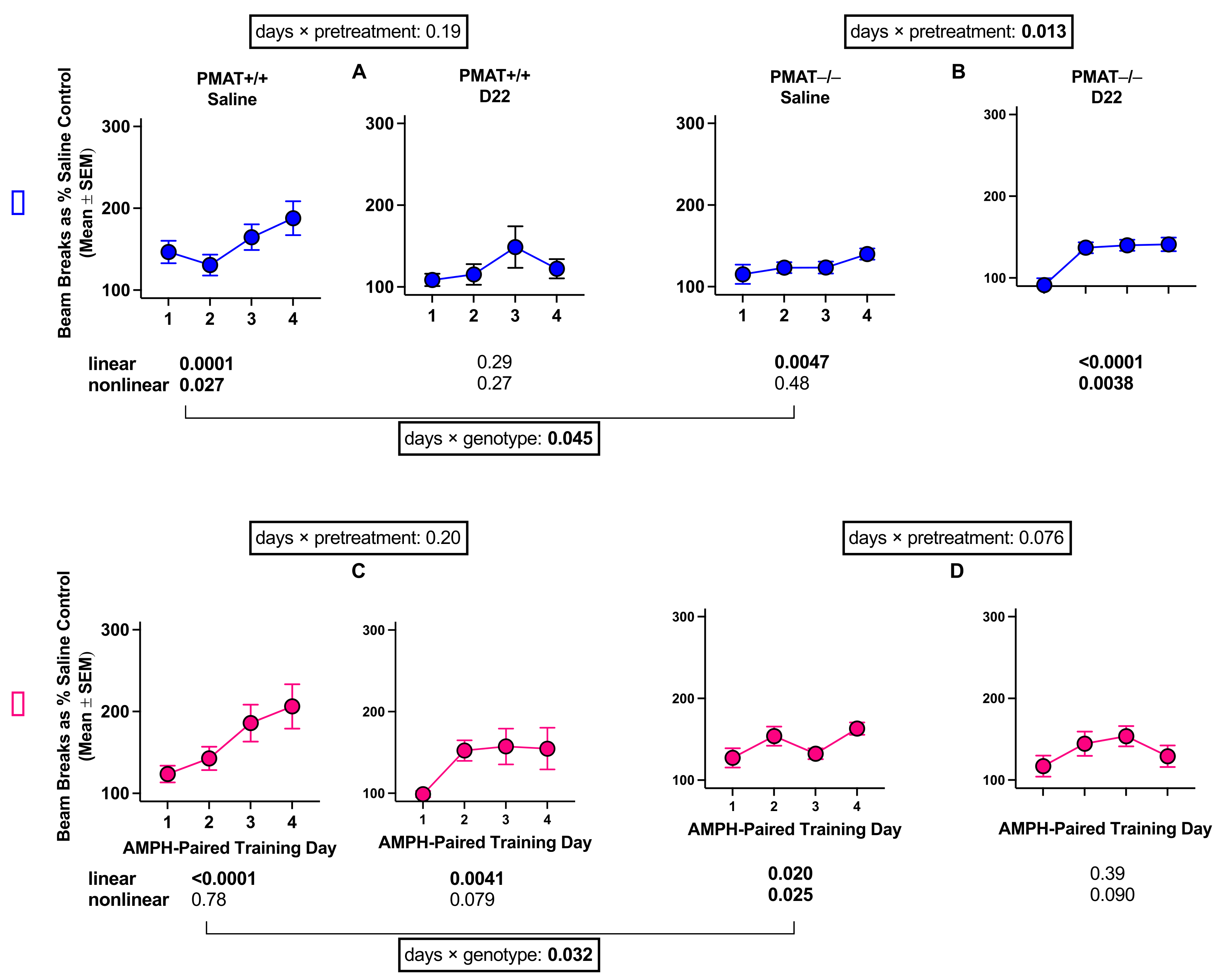
2.2.2. PMAT
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Drugs
4.3. Apparatus
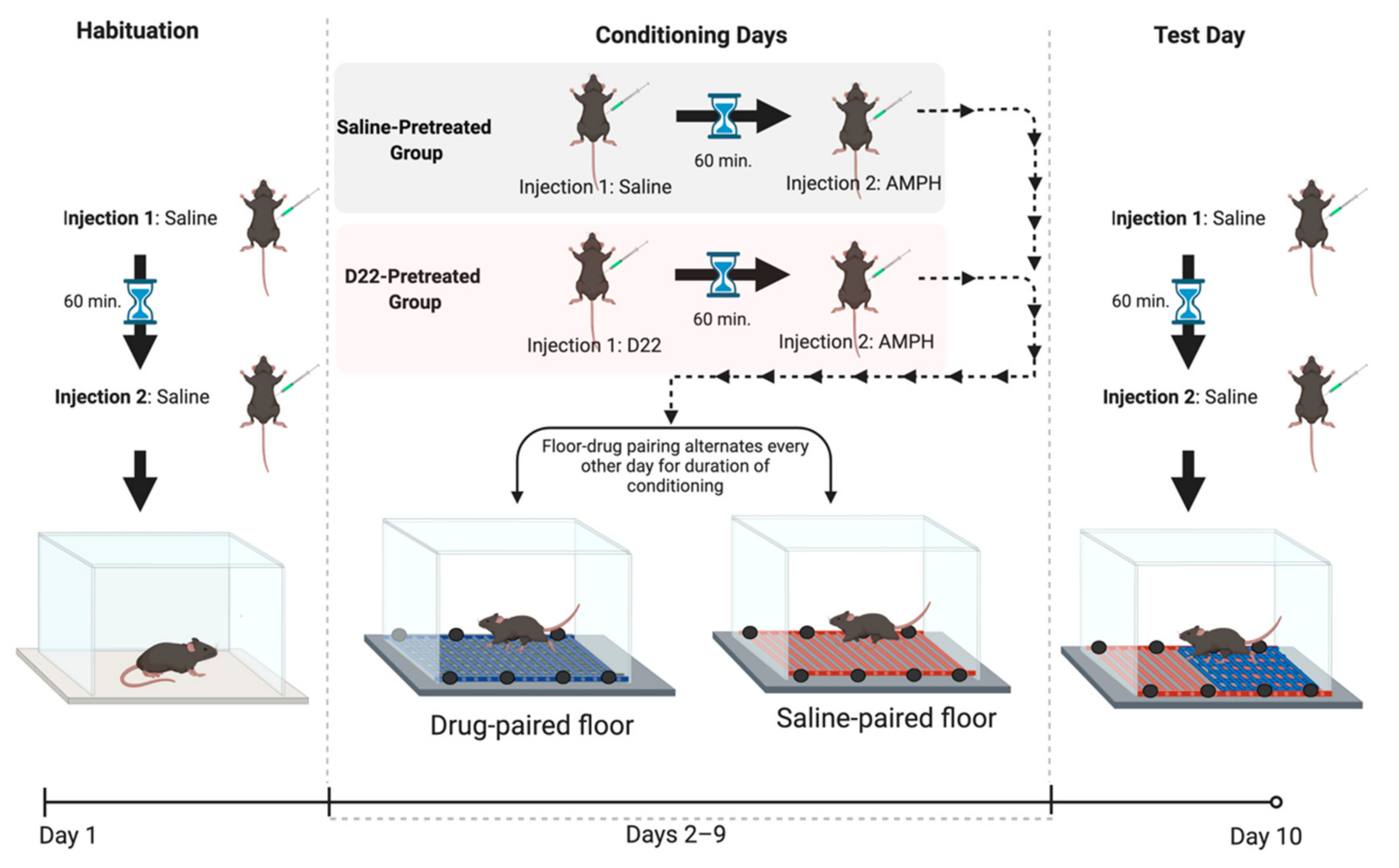
4.4. Conditioning Procedure
4.5. Data Analyses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- United Nations Office on Drugs and Crime (UNODC). World Drug Report. 2017. Available online: http://www.unodc.org/wdr2017/field/Booklet_2_HEALTH.pdf (accessed on 12 September 2021).
- Degenhardt, L.; Hall, W. Extent of illicit drug use and dependence, and their contribution to the global burden of disease. Lancet 2012, 379, 55–70. [Google Scholar] [CrossRef]
- Kariisa, M.; Scholl, L.; Wilson, N.; Seth, P.; Hoots, B. Drug Overdose Deaths Involving Cocaine and Psychostimulants with Abuse Potential—United States, 2003–2017. MMWR Morb. Mortal. Wkly. Rep. 2019, 68, 388–395. [Google Scholar] [CrossRef] [Green Version]
- Cano, M.; Huang, Y. Overdose deaths involving psychostimulants with abuse potential, excluding cocaine: State-level differences and the role of opioids. Drug Alcohol Depend. 2021, 218, 108384. [Google Scholar] [CrossRef]
- Glasner-Edwards, S.; Mooney, L.J. Methamphetamine Psychosis: Epidemiology and Management. CNS Drugs 2014, 28, 1115–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKetin, R.; Dawe, S.; Burns, R.; Hides, L.; Kavanagh, D.J.; Teesson, M.R.; Young, R.; Voce, A.; Saunders, J.B. The profile of psychiatric symptoms exacerbated by methamphetamine use. Drug Alcohol Depend. 2016, 161, 104–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brecht, M.-L.; O’Brien, A.; von Mayrhauser, C.; Anglin, M. Methamphetamine use behaviors and gender differences. Addict. Behav. 2004, 29, 89–106. [Google Scholar] [CrossRef]
- Van Etten, M.L.; Neumark, Y.D.; Anthony, J.C. Male-female differences in the earliest stages of drug involvement. Addiction 1999, 94, 1413–1419. [Google Scholar] [CrossRef]
- Becker, J.B.; Perry, A.N.; Westenbroek, C. Sex differences in the neural mechanisms mediating addiction: A new synthesis and hypothesis. Biol. Sex Differ. 2012, 3, 14. [Google Scholar] [CrossRef] [Green Version]
- Becker, J.B.; Chartoff, E. Sex differences in neural mechanisms mediating reward and addiction. Neuropsychopharmacology 2019, 44, 166–183. [Google Scholar] [CrossRef] [Green Version]
- Seth, P.; Scholl, L.; Rudd, R.A.; Bacon, S. Overdose Deaths Involving Opioids, Cocaine, and Psychostimulants—United States, 2015–2016. MMWR. Morb. Mortal. Wkly. Rep. 2018, 67, 349–358. [Google Scholar] [CrossRef]
- Sulzer, D.; Sonders, M.S.; Poulsen, N.W.; Galli, A. Mechanisms of neurotransmitter release by amphetamines: A review. Prog. Neurobiol. 2005, 75, 406–433. [Google Scholar] [CrossRef] [PubMed]
- Fleckenstein, A.E.; Volz, T.J.; Hanson, G.R. Psychostimulant-induced alterations in vesicular monoamine transporter-2 function: Neurotoxic and therapeutic implications. Neuropharmacology 2009, 56, 133–138. [Google Scholar] [CrossRef] [Green Version]
- Sulzer, D. How Addictive Drugs Disrupt Presynaptic Dopamine Neurotransmission. Neuron 2011, 69, 628–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sitte, H.H.; Freissmuth, M. Amphetamines, new psychoactive drugs and the monoamine transporter cycle. Trends Pharmacol. Sci. 2015, 36, 41–50. [Google Scholar] [CrossRef] [Green Version]
- Fleckenstein, A.E.; Gibb, J.W.; Hanson, G.R. Differential effects of stimulants on monoaminergic transporters: Pharmacological consequences and implications for neurotoxicity. Eur. J. Pharmacol. 2000, 406, 1–13. [Google Scholar] [CrossRef]
- Budygin, E.A.; Brodie, M.S.; Sotnikova, T.D.; Mateo, Y.; John, C.E.; Cyr, M.; Gainetdinov, R.R.; Jones, S.R. Dissociation of rewarding and dopamine transporter-mediated properties of amphetamine. Proc. Natl. Acad. Sci. USA 2004, 101, 7781–7786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, B.; Freeman, M.; Kondo, K.; Ayers, C.; Montgomery, J.; Paynter, R.; Kansagara, D. Pharmacotherapy for methamphetamine/amphetamine use disorder—a systematic review and meta-analysis. Addiction 2019, 114, 2122–2136. [Google Scholar] [CrossRef]
- Howell, L.L.; Negus, S.S. Monoamine Transporter Inhibitors and Substrates as Treatments for Stimulant Abuse. Adv. Pharmacol. 2014, 69, 129–176. [Google Scholar] [CrossRef] [Green Version]
- Gasser, P.J.; Lowry, C.A.; Orchinik, M. Corticosterone-Sensitive Monoamine Transport in the Rat Dorsomedial Hypothalamus: Potential Role for Organic Cation Transporter 3 in Stress-Induced Modulation of Monoaminergic Neurotransmission. J. Neurosci. 2006, 26, 8758–8766. [Google Scholar] [CrossRef]
- Graf, E.N.; Wheeler, R.A.; Baker, D.A.; Ebben, A.L.; Hill, J.E.; McReynolds, J.; Robble, M.A.; Vranjkovic, O.; Wheeler, D.S.; Mantsch, J.R.; et al. Corticosterone acts in the nucleus accumbens to enhance dopamine signaling and potentiate reinstatement of cocaine seeking. J. Neurosci. 2013, 33, 11800–11810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, J.E.; Gasser, P.J. Organic cation transporter 3 is densely expressed in the intercalated cell groups of the amygdala: Anatomical evidence for a stress hormone-sensitive dopamine clearance system. J. Chem. Neuroanat. 2013, 52, 36–43. [Google Scholar] [CrossRef] [PubMed]
- McReynolds, J.; Taylor, A.; Vranjkovic, O.; Ambrosius, T.; Derricks, O.; Nino, B.; Kurtoglu, B.; A Wheeler, R.; A Baker, D.; Gasser, P.J.; et al. Corticosterone Potentiation of Cocaine-Induced Reinstatement of Conditioned Place Preference in Mice is Mediated by Blockade of the Organic Cation Transporter 3. Neuropsychopharmacology 2016, 42, 757–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayer, F.; Schmid, D.; Owens, W.A.; Gould, G.G.; Apuschkin, M.; Kudlacek, O.; Salzer, I.; Boehm, S.; Chiba, P.; Williams, P.H.; et al. An unsuspected role for organic cation transporter 3 in the actions of amphetamine. Neuropsychopharmacology 2018, 43, 2408–2417. [Google Scholar] [CrossRef] [PubMed]
- Gasser, P.J. Roles for the uptake2 transporter OCT3 in regulation of dopaminergic neurotransmission and behavior. Neurochem. Int. 2019, 123, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Holleran, K.M.; Rose, J.H.; Fordahl, S.C.; Benton, K.C.; Rohr, K.E.; Gasser, P.J.; Jones, S.R. Organic cation transporter 3 and the dopamine transporter differentially regulate catecholamine uptake in the basolateral amygdala and nucleus accumbens. Eur. J. Neurosci. 2020, 52, 4546–4562. [Google Scholar] [CrossRef]
- Gasser, P.J. Organic Cation Transporters in Brain Catecholamine Homeostasis. In Handbook of Experimental Pharmacology; Springer: Cham, Switzerland, 2021; Volume 266, pp. 187–197. [Google Scholar] [CrossRef]
- Maier, J.; Niello, M.; Rudin, D.; Daws, L.C.; Sitte, H.H. The Interaction of Organic Cation Transporters 1-3 and PMAT with Psychoactive Substances. In Handbook of Experimental Pharmacology; Springer: Cham, Switzerland, 2021; Volume 266, pp. 199–214. [Google Scholar] [CrossRef]
- Koepsell, H. Organic Cation Transporters in Health and Disease. Pharmacol. Rev. 2020, 72, 253–319. [Google Scholar] [CrossRef]
- Gasser, P.J.; Orchinik, M.; Raju, I.; Lowry, C.A. Distribution of organic cation transporter 3, a corticosterone-sensitive monoamine transporter, in the rat brain. J. Comp. Neurol. 2009, 512, 529–555. [Google Scholar] [CrossRef] [PubMed]
- Sweet, D.H. Organic Cation Transporter Expression and Function in the CNS. In Handbook of Experimental Pharmacology; Springer: Cham, Switzerland, 2021; Volume 266, pp. 41–80. [Google Scholar] [CrossRef]
- Ikemoto, S.; Bonci, A. Neurocircuitry of drug reward. Neuropharmacology 2014, 76, 329–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoyama, N.; Takahashi, N.; Kitaichi, K.; Ishihara, R.; Saito, S.; Maeno, N.; Ji, X.; Takagi, K.; Sekine, Y.; Iyo, M.; et al. Association between gene polymorphisms of SLC22A3 and metham-phetamine use disorder. Alcohol Clin. Exp. Res. 2006, 30, 1644–1649. [Google Scholar] [CrossRef]
- Duan, H.; Wang, J. Selective Transport of Monoamine Neurotransmitters by Human Plasma Membrane Monoamine Transporter and Organic Cation Transporter 3. J. Pharmacol. Exp. Ther. 2010, 335, 743–753. [Google Scholar] [CrossRef] [Green Version]
- Vanderschuren, L.J.M.J.; Pierce, R.C. Sensitization Processes in Drug Addiction. Curr. Top. Behav. Neurosci. 2010, 3, 179–195. [Google Scholar]
- Vieira, L.S.; Wang, J. Brain Plasma Membrane Monoamine Transporter in Health and Disease. In Handbook of Experimental Pharmacology; Springer: Cham, Switzerland, 2021; Volume 266, pp. 253–280. [Google Scholar] [CrossRef]
- Koek, W. Morphine-induced conditioned place preference and effects of morphine pre-exposure in adolescent and adult male C57BL/6J mice. Psychopharmacology 2016, 233, 2015–2024. [Google Scholar] [CrossRef] [Green Version]
- Fraser-Spears, R.; Krause-Heuer, A.M.; Basiouny, M.; Mayer, F.; Manishimwe, R.; Wyatt, N.A.; Dobrowolski, J.C.; Roberts, M.P.; Greguric, I.; Kumar, N.; et al. Comparative analysis of novel decynium-22 analogs to inhibit transport by the low-affinity, high-capacity monoamine transporters, organic cation transporters 2 and 3, and plasma membrane monoamine transporter. Eur. J. Pharmacol. 2019, 842, 351–364. [Google Scholar] [CrossRef]
- Engel, K.; Zhou, M.; Wang, J. Identification and Characterization of a Novel Monoamine Transporter in the Human Brain. J. Biol. Chem. 2004, 279, 50042–50049. [Google Scholar] [CrossRef] [Green Version]
- Vialou, V.; Balasse, L.; Callebert, J.; Launay, J.M.; Giros, B.; Gautron, S. Altered aminergic neurotransmission in the brain of organ-ic cation transporter 3-deficient mice. J. Neurochem. 2008, 106, 1471–1482. [Google Scholar] [CrossRef] [PubMed]
- Wultsch, T.; Grimberg, G.; Schmitt, A.; Painsipp, E.; Wetzstein, H.; Breitenkamp, A.F.S.; Gründemann, D.; Schömig, E.; Lesch, K.-P.; Gerlach, M.; et al. Decreased anxiety in mice lacking the organic cation transporter 3. J. Neural Transm. 2009, 116, 689–697. [Google Scholar] [CrossRef]
- Gilman, T.L.; George, C.M.; Vitela, M.; Herrera-Rosales, M.; Basiouny, M.S.; Koek, W.; Daws, L.C. Constitutive plasma membrane monoamine transporter (PMAT, Slc29a4) deficiency subtly affects anxiety-like and coping behaviours. Eur. J. Neurosci. 2018, 48, 1706–1716. [Google Scholar] [CrossRef] [PubMed]
- Beatty, W.W.; Fessler, R.G. Ontogeny of sex differences in open-field behavior and sensitivity to electric shock in the rat. Physiol. Behav. 1976, 16, 413–417. [Google Scholar] [CrossRef]
- Blizard, D.A.; Lippman, H.; Chen, J.J. Sex differences in open-field behavior in the rat: The inductive and activational role of gonadal hormones. Physiol. Behav. 1975, 14, 601–608. [Google Scholar] [CrossRef]
- Masur, J.; Schutz, M.T.; Boerngen, R. Gender differences in open-field behavior as a function of age. Dev. Psychobiol. 1980, 13, 107–110. [Google Scholar] [CrossRef]
- Beatty, W.W.; Holzer, G.A. Sex differences in stereotyped behavior in the rat. Pharmacol. Biochem. Behav. 1978, 9, 777–783. [Google Scholar] [CrossRef]
- Brass, C.A.; Glick, S.D. Sex differences in drug-induced rotation in two strains of rats. Brain Res. 1981, 223, 229–234. [Google Scholar] [CrossRef]
- Goodrich, C.; Lange, J. A differential sex effect of amphetamine on exploratory behavior in maturing mice. Physiol. Behav. 1986, 38, 663–666. [Google Scholar] [CrossRef]
- Siuciak, J.; McCarthy, S.; Chapin, D.; Reed, T.; Vorhees, C.; Repaske, D. Behavioral and neurochemical characterization of mice deficient in the phosphodiesterase-1B (PDE1B) enzyme. Neuropharmacology 2007, 53, 113–124. [Google Scholar] [CrossRef]
- Van den Buuse, M.; Halley, P.; Hill, R.; Labots, M.; Martin, S. Altered N-methyl-d-aspartate receptor function in reelin heterozygous mice: Male–female differences and comparison with dopaminergic activity. Prog. Neuro Psychopharmacol. Biol. Psychiatry 2012, 37, 237–246. [Google Scholar] [CrossRef]
- Robinson, T.E.; Berridge, K.C. Addiction. Annu. Rev. Psychol. 2003, 54, 25–53. [Google Scholar] [CrossRef] [PubMed]
- Hemby, S.E.; Jones, G.H.; Justice, J.B., Jr.; Neill, D.B. Conditioned locomotor activity but not conditioned place preference following intra-accumbens infusions of cocaine. Psychopharmacology 1992, 106, 330–336. [Google Scholar] [CrossRef]
- Olmstead, M.C.; Franklin, K.B.J. The development of a conditioned place preference to morphine: Effects of microinjections into various CNS sites. Behav. Neurosci. 1997, 111, 1324–1334. [Google Scholar] [CrossRef]
- Boye, S.M.; Rompré, P.-P. Behavioral Evidence of Depolarization Block of Dopamine Neurons after Chronic Treatment with Haloperidol and Clozapine. J. Neurosci. 2000, 20, 1229–1239. [Google Scholar] [CrossRef] [Green Version]
- Shen, F.; Meredith, G.E.; Napier, T.C. Amphetamine-Induced Place Preference and Conditioned Motor Sensitization Requires Activation of Tyrosine Kinase Receptors in the Hippocampus. J. Neurosci. 2006, 26, 11041–11051. [Google Scholar] [CrossRef]
- Koepsell, H. General Overview of Organic Cation Transporters in Brain. In Handbook of Experimental Pharmacology; Springer: Cham, Switzerland, 2021; Volume 226, pp. 1–39. [Google Scholar] [CrossRef]
- Zwart, R.; Verhaagh, S.; Buitelaar, M.; Popp-Snijders, C.; Barlow, D.P. Impaired activity of the extraneuronal monoamine transporter system known as uptake-2 in Orct3/Slc22a3 deficient mice. Mol. Cell. Biol. 2001, 21, 4188–4196. [Google Scholar] [CrossRef] [Green Version]
- Bardo, M.; Klebaur, J.; Deaton, C. Environmental enrichment decreases intravenous self-administration of amphetamine in female and male rats. Psychopharmacology 2001, 155, 278–284. [Google Scholar] [CrossRef]
- Neugebauer, N.M.; Cunningham, S.T.; Zhu, J.; Bryant, R.I.; Middleton, L.S.; Dwoskin, L.P. Effects of environmental enrichment on behavior and dopamine transporter function in medial prefrontal cortex in adult rats prenatally treated with cocaine. Dev. Brain Res. 2004, 153, 213–223. [Google Scholar] [CrossRef]
- Zhu, J.; Apparsundaram, S.; Bardo, M.T.; Dwoskin, L.P. Environmental enrichment decreases cell surface expression of the dopamine transporter in rat medial prefrontal cortex. J. Neurochem. 2005, 93, 1434–1443. [Google Scholar] [CrossRef]
- Stairs, D.J.; Klein, E.D.; Bardo, M.T. Effects of environmental enrichment on extinction and reinstatement of amphetamine self-administration and sucrose-maintained responding. Behav. Pharmacol. 2006, 17, 597–604. [Google Scholar] [CrossRef]
- Zakharova, E.; Miller, J.; Unterwald, E.; Wade, D.; Izenwasser, S. Social and physical environment alter cocaine conditioned place preference and dopaminergic markers in adolescent male rats. Neuroscience 2009, 163, 890–897. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.-S.; Yu, J.H.; Kim, C.H.; Choi, J.Y.; Seo, J.H.; Lee, M.-Y.; Yi, C.H.; Choi, T.H.; Ryu, Y.H.; Lee, J.E.; et al. Environmental enrichment enhances synaptic plasticity by internalization of striatal dopamine transporters. J. Cereb. Blood Flow Metab. 2016, 36, 2122–2133. [Google Scholar] [CrossRef] [Green Version]
- Rae, M.; Zanos, P.; Georgiou, P.; Chivers, P.; Bailey, A.; Camarini, R. Environmental enrichment enhances conditioned place preference to ethanol via an oxytocinergic-dependent mechanism in male mice. Neuropharmacology 2018, 138, 267–274. [Google Scholar] [CrossRef]
- Baganz, N.L.; Horton, R.E.; Calderon, A.S.; Owens, W.A.; Munn, J.L.; Watts, L.T.; Koldzic-Zivanovic, N.; Jeske, N.A.; Koek, W.; Toney, G.M.; et al. Organic cation transporter 3: Keeping the brake on extracellular serotonin in serotonin-transporter-deficient mice. Proc. Natl. Acad. Sci. USA 2008, 105, 18976–18981. [Google Scholar] [CrossRef] [Green Version]
- Horton, R.E.; Apple, D.M.; Owens, W.A.; Baganz, N.L.; Cano, S.; Mitchell, N.C.; Vitela, M.; Gould, G.; Koek, W.; Daws, L.C. Decynium-22 Enhances SSRI-Induced Antidepressant-Like Effects in Mice: Uncovering Novel Targets to Treat Depression. J. Neurosci. 2013, 33, 10534–10543. [Google Scholar] [CrossRef]
- Belzung, C.; Barreau, S. Differences in Drug-Induced Place Conditioning Between BALB/c and C57Bl/6 Mice. Pharmacol. Biochem. Behav. 2000, 65, 419–423. [Google Scholar] [CrossRef]
- Orsini, C.; Buchini, F.; Piazza, P.V.; Puglisi-Allegra, S.; Cabib, S. Susceptibility to amphetamine-induced place preference is predicted by locomotor response to novelty and amphetamine in the mouse. Psychopharmacology 2004, 172, 264–270. [Google Scholar] [CrossRef] [PubMed]
- Thanos, C.G.; Bintz, B.; Emerich, D.F. Microencapsulated Choroid Plexus Epithelial Cell Transplants for Repair of the Brain. Adv. Exp. Med. Biol. 2010, 670, 80–91. [Google Scholar] [CrossRef] [PubMed]
- Sherman, J.E.; Pickman, C.; Rice, A.; Liebeskind, J.C.; Holman, E.W. Rewarding and aversive effects of morphine: Temporal and pharmacological properties. Pharmacol. Biochem. Behav. 1980, 13, 501–505. [Google Scholar] [CrossRef]
- Bardo, M.; Bevins, R. Conditioned place preference: What does it add to our preclinical understanding of drug reward? Psychopharmacology 2000, 153, 31–43. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, C.L.; Gremel, C.M.; Groblewski, P.A. Drug-induced conditioned place preference and aversion in mice. Nat. Protocols 2006, 1, 1662–1670. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.L.; Niehus, D.R.; Malott, D.H.; Prather, L.K. Genetic differences in the rewarding and activating effects of morphine and ethanol. Psychopharmacology 1992, 107, 385–393. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genotype | OCT3+/+ | OCT3−/− | PMAT+/+ | PMAT−/− | ||||
|---|---|---|---|---|---|---|---|---|
| Sex | Male | Female | Male | Female | Male | Female | Male | Female |
| Saline | 768 (42) | 944 (56) | 747 (37) | 973 (52) | 821 (40) | 1279 (65) | 921 (33) | 1119 (59) |
| Saline + Amphetamine | 808 (75) | 1055 (79) | 859 (66) | 1090 (98) | 1015 (30) | 1448 (63) | 1084 (146) | 1415 (97) |
| D22 + Amphetamine | 903 (58) | 1067 (51) | 805 (75) | 1119 (85) | 929 (72) | 1255 (75) | 822 (80) | 1186 (110) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clauss, N.J.; Koek, W.; Daws, L.C. Role of Organic Cation Transporter 3 and Plasma Membrane Monoamine Transporter in the Rewarding Properties and Locomotor Sensitizing Effects of Amphetamine in Male andFemale Mice. Int. J. Mol. Sci. 2021, 22, 13420. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413420
Clauss NJ, Koek W, Daws LC. Role of Organic Cation Transporter 3 and Plasma Membrane Monoamine Transporter in the Rewarding Properties and Locomotor Sensitizing Effects of Amphetamine in Male andFemale Mice. International Journal of Molecular Sciences. 2021; 22(24):13420. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413420
Chicago/Turabian StyleClauss, Nikki J., Wouter Koek, and Lynette C. Daws. 2021. "Role of Organic Cation Transporter 3 and Plasma Membrane Monoamine Transporter in the Rewarding Properties and Locomotor Sensitizing Effects of Amphetamine in Male andFemale Mice" International Journal of Molecular Sciences 22, no. 24: 13420. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms222413420