
NMR Characterization of Angiogenin Variants and tRNAAla Products Impacting Aberrant Protein Oligomerization


 , and
, and 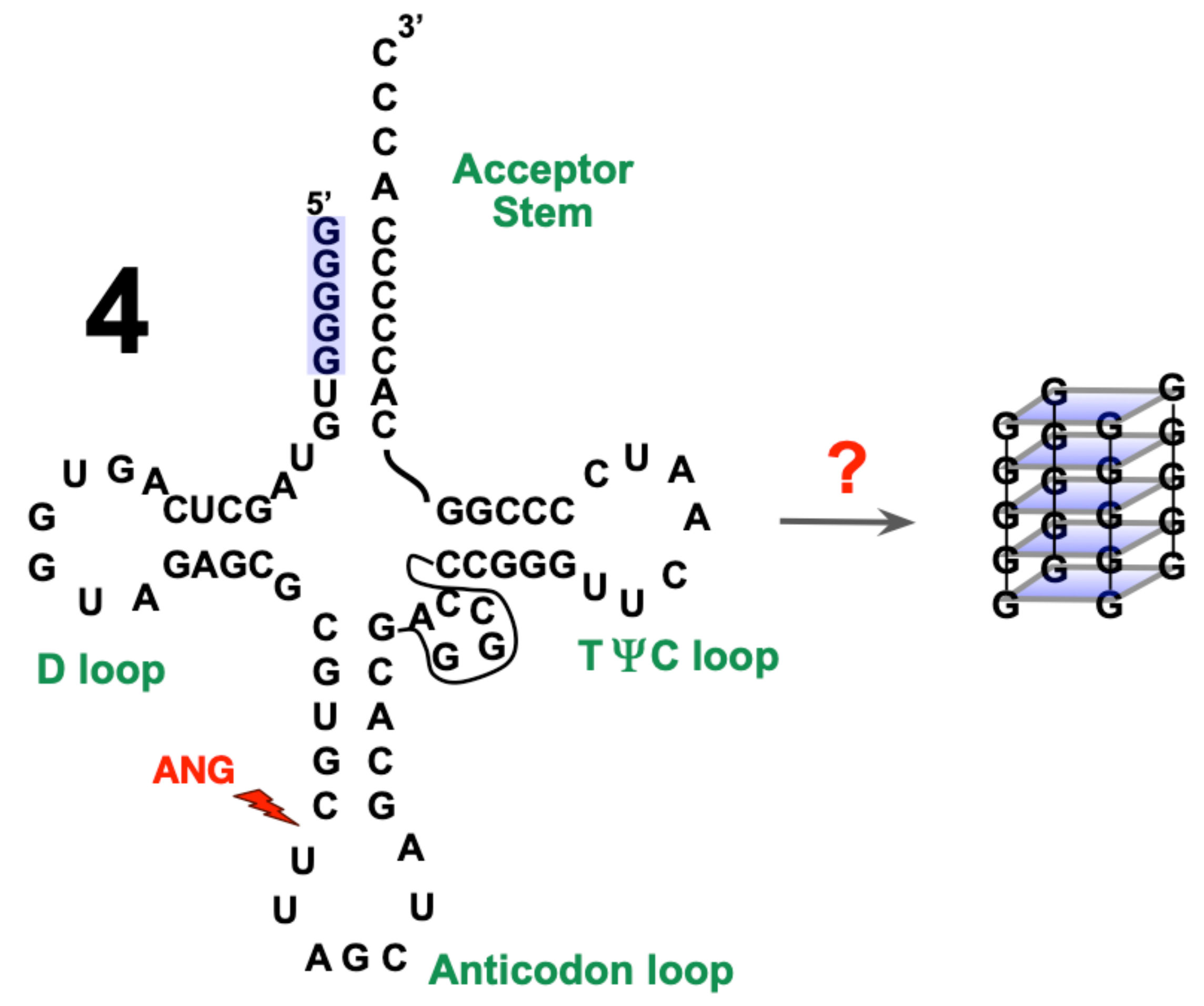{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
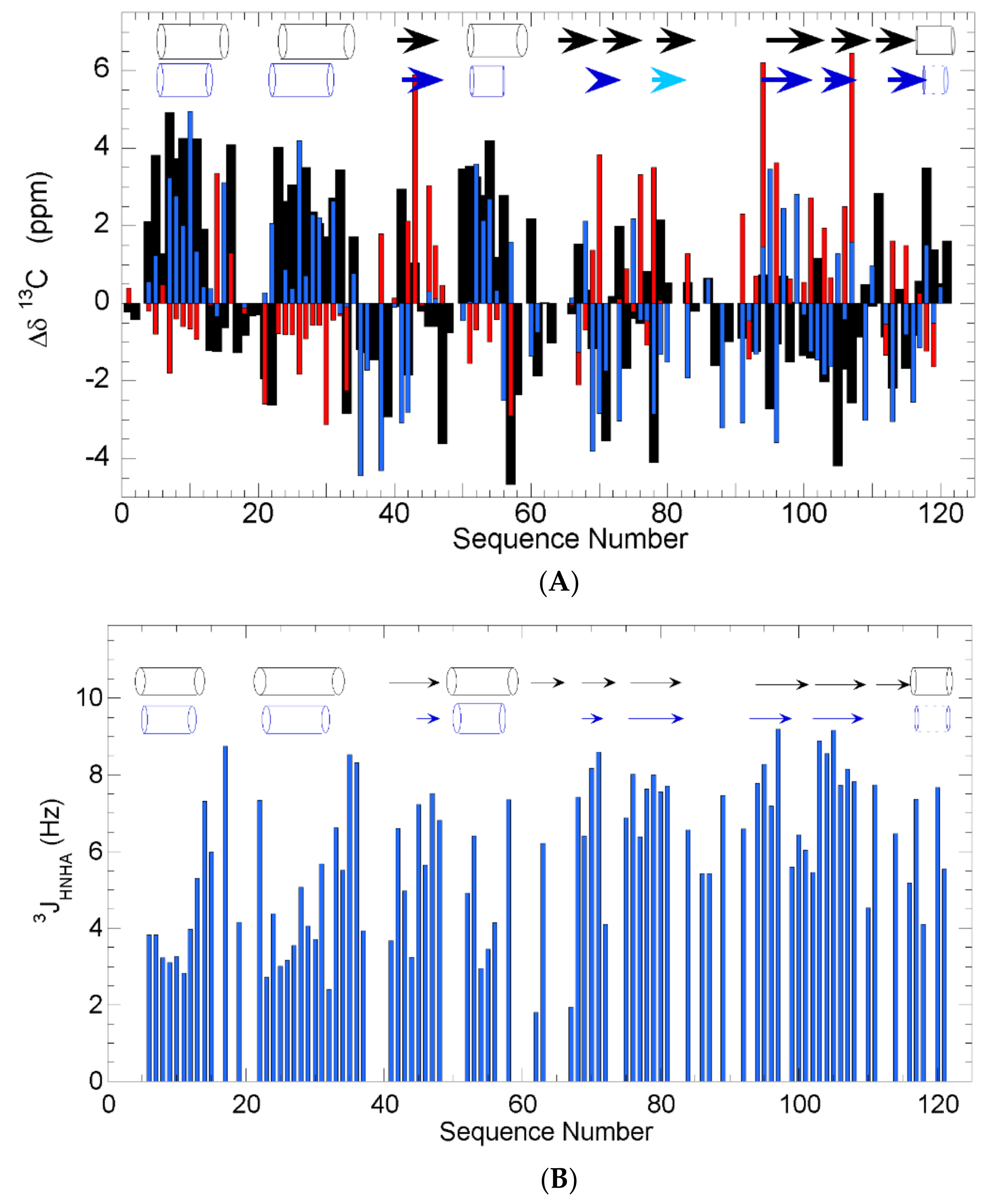
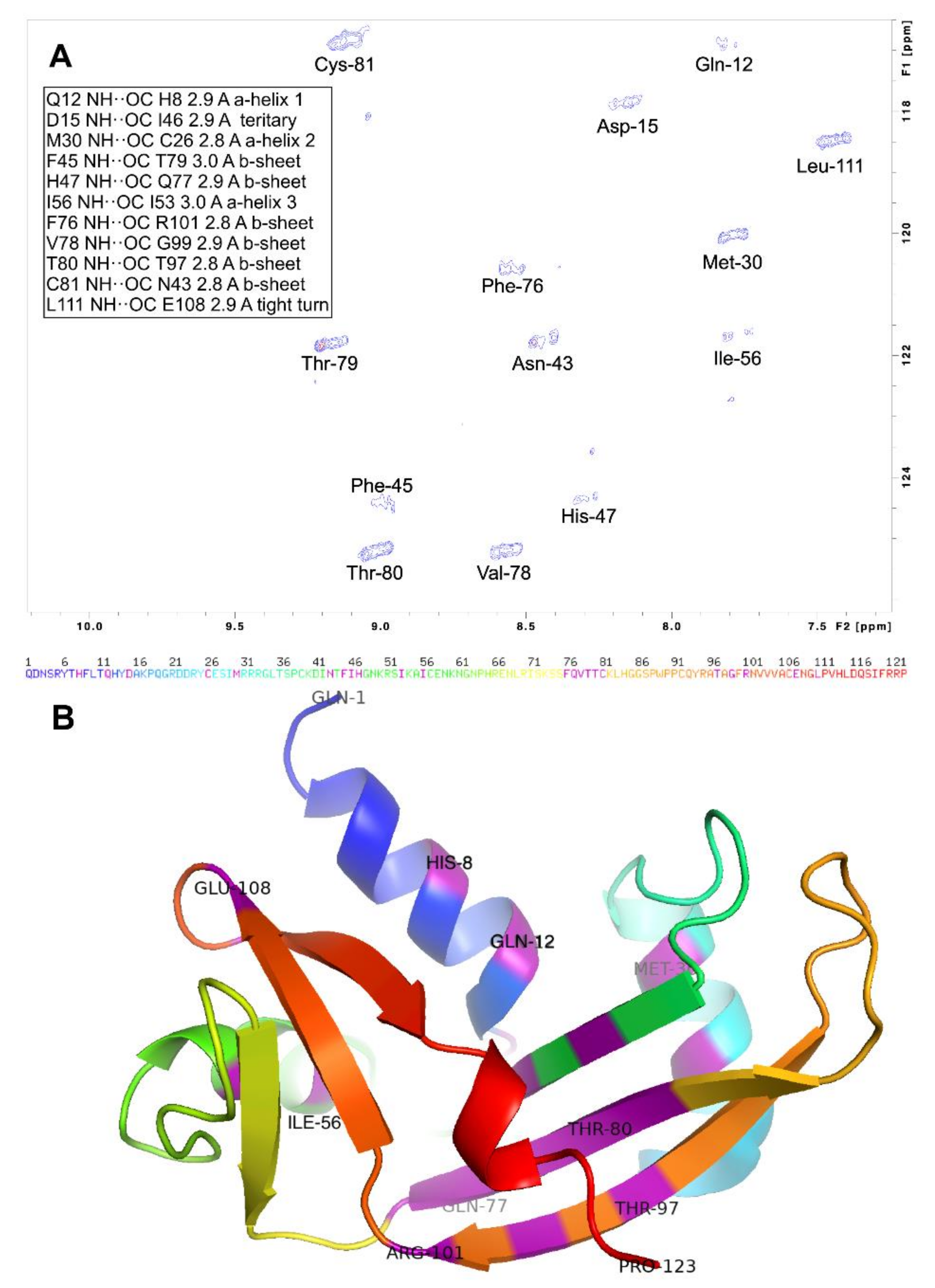
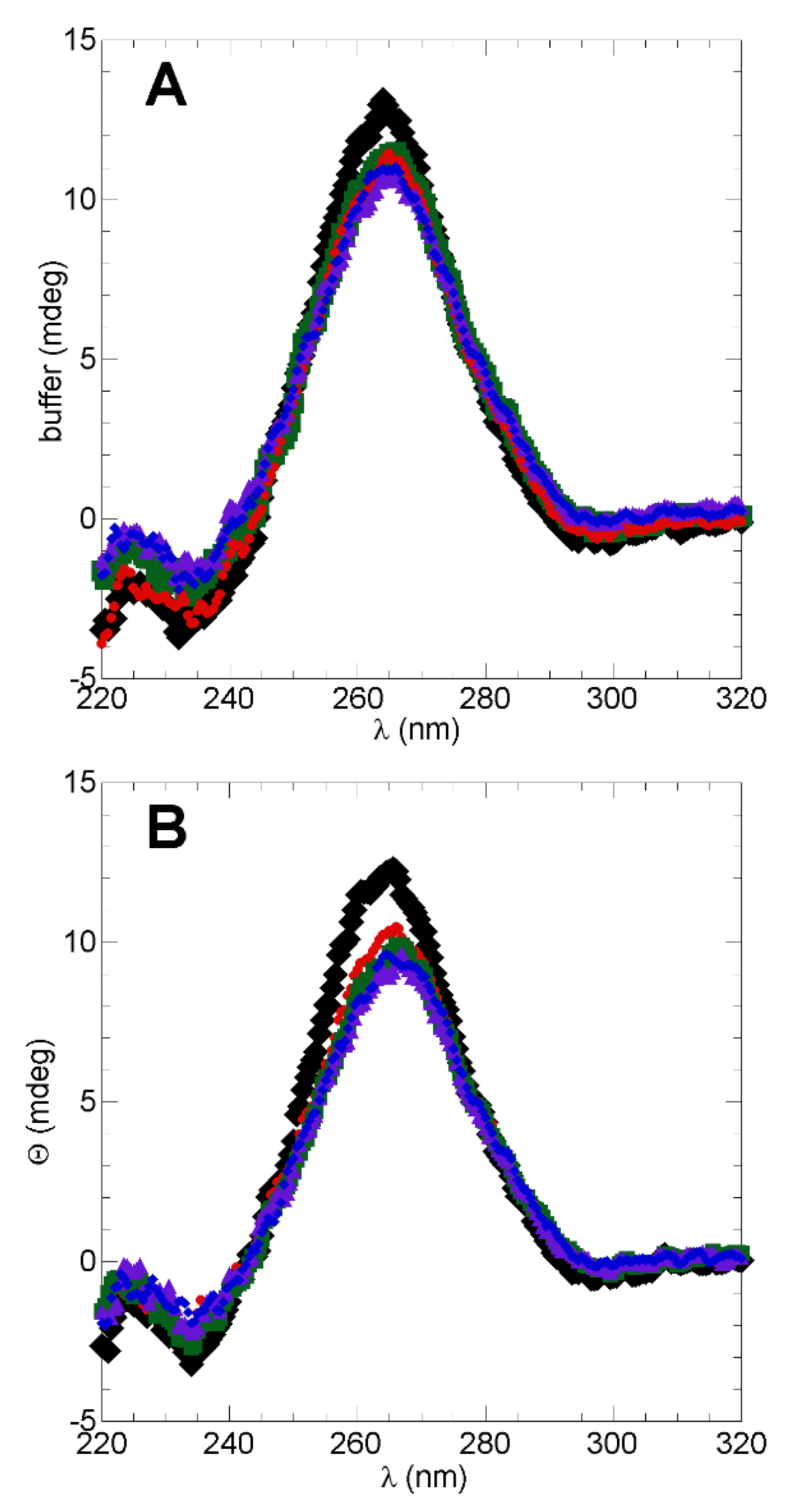
2.1. Secondary Structure in WT h-ANG
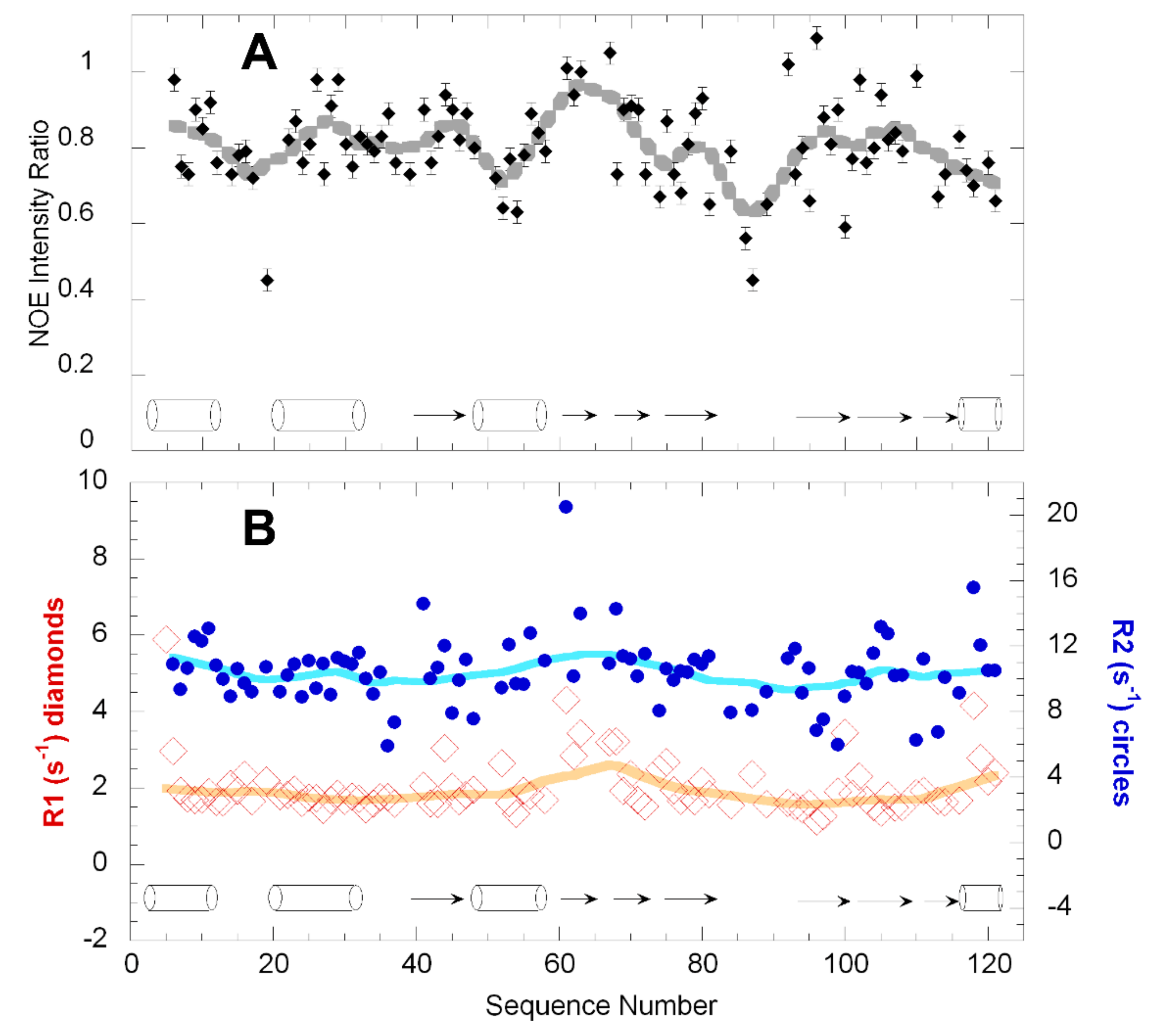
2.2. Dynamics of WT h-ANG
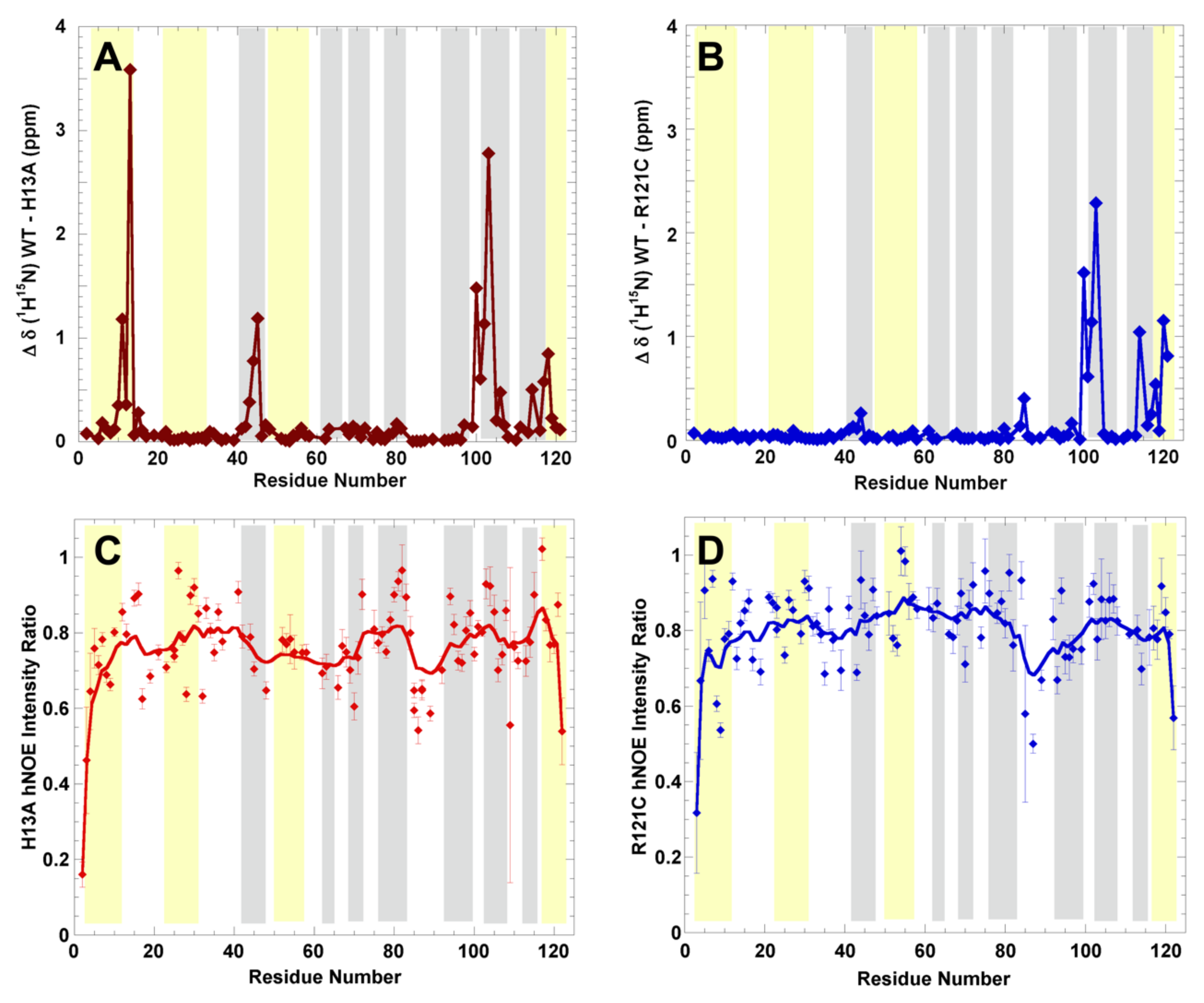
2.3. Structural Changes and Dynamics of h-Ang Variants
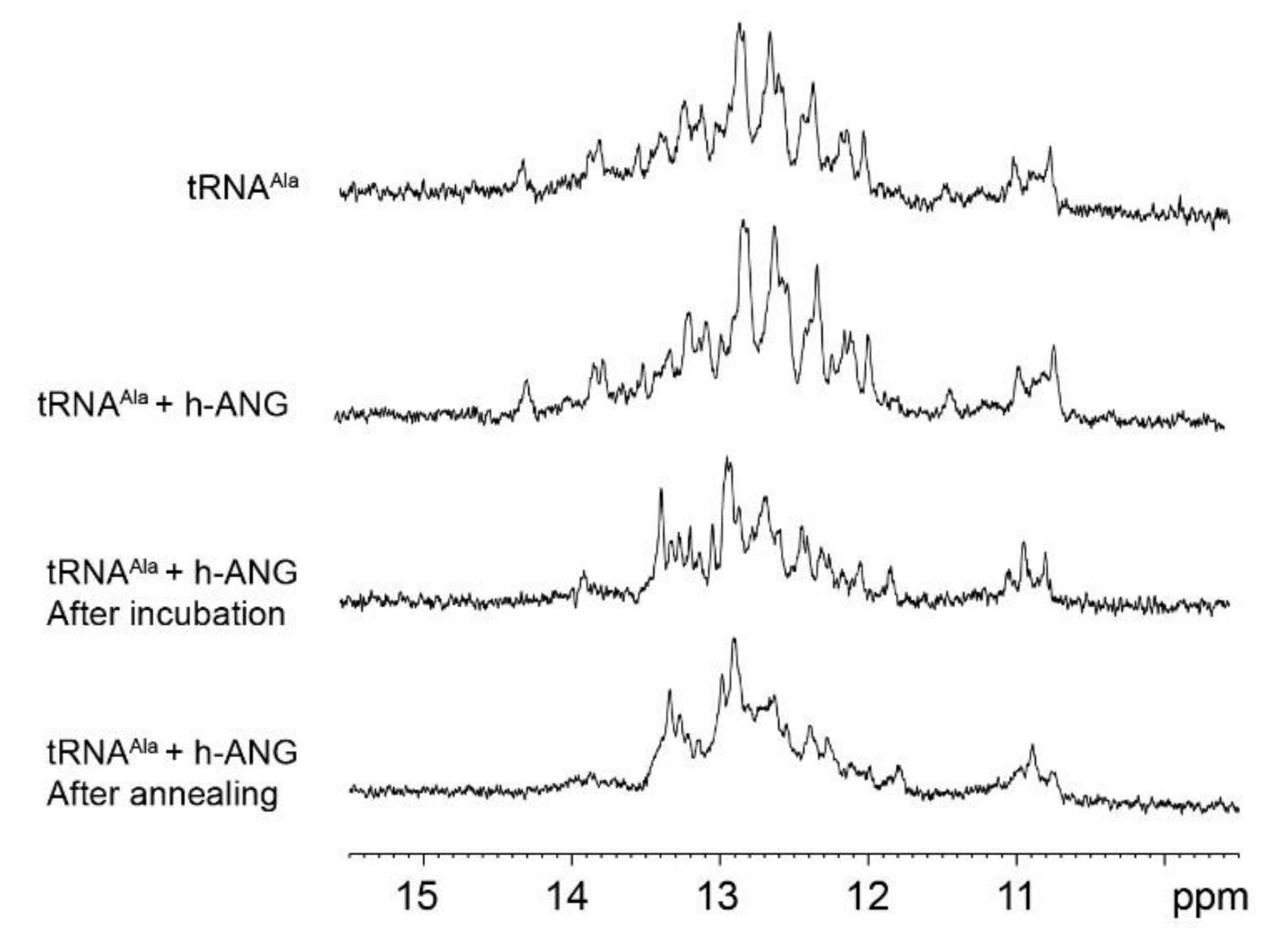
2.4. Assessment of tRNAAla Structure Following Cleavage by h-ANG
3. Discussion
3.1. Secondary Structure of h-ANG under Near-Physiological Conditions of pH and Temperature
3.2. h-ANG Adopts a Highly Rigid Structure in Solution
3.3. On the Conformation of tRNAAla
3.4. Conclusions and Implications for Protein Oligomerization
4. Materials and Methods
4.1. ANG Mutants Cloning
- H13A, F: 5′-CATTTCCTGACCCAGGCCTATGACGCTAAAC-3′,
- R: 5′-GTTTAGCGTCATAGGCCTGGGTCAGGAAATG-3′;
- C39W, F:5′ CCGTGGGTTAACTAGCCCGTGGAAAGATATC 3′,
- R: 5′ GATATCTTTCCACGGGCTAGTTAACCCACGG 3′;
- R121C, F: 5′-GTCCATCTAGATCAGTCTATCTTCTGCAGGCCT-3′,
- R: 5′-AGGCCTGCAGAAGATAGACTGATCTAGATGGAC-3′.
4.2. Expression and Purification of WT ANG and Mutants
4.3. Synthesis and Purification of tRNAAla
4.4. NMR Experiments
4.5. NMR Experiments: Ps/ns Timescale Dynamics through the {1H}-15N NOE
4.6. NMR Experiments: μs/ms Timescale Dynamics via Longitudinal (R1) and Transverse (R2) Relaxation Rates
4.7. NMR Experiments: H/D Exchange
4.8. NMR Experiments: Assignment of h-ANG Variant NMR Spectra
4.9. Characterization of h-ANG Cleavage on tRNAAla Conformation
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mulder, D.W.; Kurland, L.T.; Offord, K.P.; Beard, C.M. Familial Adult Motor Neuron Disease: Amyotrophic Lateral Sclerosis. Neurology 1986, 36, 511–517. [Google Scholar] [CrossRef] [PubMed]
- Lill, C.M.; Abel, O.; Bertram, L.; Al-Chalabi, A. Keeping up with Genetic Discoveries in Amyotrophic Lateral Sclerosis: The ALSoD and ALSGene Databases. Amyotroph. Lateral Scler. 2011, 12, 238–249. [Google Scholar] [CrossRef] [PubMed]
- Abel, O.; Powell, J.F.; Andersen, P.M.; Al-Chalabi, A. ALSoD: A User-Friendly Online Bioinformatics Tool for Amyotrophic Lateral Sclerosis Genetics. Hum. Mutat. 2012, 33, 1345–1351. [Google Scholar] [CrossRef] [PubMed]
- Hardiman, O.; van den Berg, L.H.; Kiernan, M.C. Clinical Diagnosis and Management of Amyotrophic Lateral Sclerosis. Nat. Rev. Neurol. 2011, 7, 639–649. [Google Scholar] [CrossRef] [PubMed]
- Ferraiuolo, L.; Kirby, J.; Grierson, A.J.; Sendtner, M.; Shaw, P.J. Molecular Pathways of Motor Neuron Injury in Amyotrophic Lateral Sclerosis. Nat. Rev. Neurol. 2011, 7, 616–630. [Google Scholar] [CrossRef]
- Andersen, P.M.; Al-Chalabi, A. Clinical Genetics of Amyotrophic Lateral Sclerosis: What Do We Really Know? Nat. Rev. Neurol. 2011, 7, 603–615. [Google Scholar] [CrossRef]
- Perry, J.J.P.; Shin, D.S.; Tainer, J.A. Amyotrophic Lateral Sclerosis. Adv. Exp. Med. Biol. 2010, 685, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Le Gall, L.; Anakor, E.; Connolly, O.; Vijayakumar, U.G.; Duddy, W.J.; Duguez, S. Molecular and Cellular Mechanisms Affected in ALS. J. Pers. Med. 2020, 10, 101. [Google Scholar] [CrossRef]
- Shorter, J. Phase Separation of RNA-Binding Proteins in Physiology and Disease: An Introduction to the JBC Reviews Thematic Series. J. Biol. Chem. 2019, 294, 7113–7114. [Google Scholar] [CrossRef] [Green Version]
- Colombrita, C.; Zennaro, E.; Fallini, C.; Weber, M.; Sommacal, A.; Buratti, E.; Silani, V.; Ratti, A. TDP-43 Is Recruited to Stress Granules in Conditions of Oxidative Insult. J. Neurochem. 2009, 111, 1051–1061. [Google Scholar] [CrossRef]
- Greenway, M.J.; Andersen, P.M.; Russ, C.; Ennis, S.; Cashman, S.; Donaghy, C.; Patterson, V.; Swingler, R.; Kieran, D.; Prehn, J.; et al. ANG Mutations Segregate with Familial and “sporadic” Amyotrophic Lateral Sclerosis. Nat. Genet. 2006, 38, 411–413. [Google Scholar] [CrossRef] [PubMed]
- Sorrentino, S.; Libonati, M. Human Pancreatic-Type and Nonpancreatic-Type Ribonucleases: A Direct Side-by-Side Comparison of Their Catalytic Properties. Arch. Biochem. Biophys. 1994, 312, 340–348. [Google Scholar] [CrossRef] [PubMed]
- Strydom, D.J.; Fett, J.W.; Lobb, R.R.; Alderman, E.M.; Bethune, J.L.; Riordan, J.F.; Vallee, B.L. Amino Acid Sequence of Human Tumor Derived Angiogenin. Biochemistry 1985, 24, 5486–5494. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, R.; Strydom, D.J.; Olson, K.A.; Vallee, B.L. Isolation of Angiogenin from Normal Human Plasma. Biochemistry 1987, 26, 5141–5146. [Google Scholar] [CrossRef] [PubMed]
- Spong, C.Y.; Ghidini, A.; Sherer, D.M.; Pezzullo, J.C.; Ossandon, M.; Eglinton, G.S. Angiogenin: A Marker for Preterm Delivery in Midtrimester Amniotic Fluid. Am. J. Obstet. Gynecol. 1997, 176, 415–418. [Google Scholar] [CrossRef]
- Fett, J.W.; Strydom, D.J.; Lobb, R.R.; Alderman, E.M.; Bethune, J.L.; Riordan, J.F.; Vallee, B.L. Isolation and Characterization of Angiogenin, an Angiogenic Protein from Human Carcinoma Cells. Biochemistry 1985, 24, 5480–5486. [Google Scholar] [CrossRef]
- Skog, J.; Würdinger, T.; van Rijn, S.; Meijer, D.H.; Gainche, L.; Sena-Esteves, M.; Curry, W.T.; Carter, B.S.; Krichevsky, A.M.; Breakefield, X.O. Glioblastoma Microvesicles Transport RNA and Proteins That Promote Tumour Growth and Provide Diagnostic Biomarkers. Nat. Cell Biol. 2008, 10, 1470–1476. [Google Scholar] [CrossRef]
- Tsuji, T.; Sun, Y.; Kishimoto, K.; Olson, K.A.; Liu, S.; Hirukawa, S.; Hu, G.-F. Angiogenin Is Translocated to the Nucleus of HeLa Cells and Is Involved in Ribosomal RNA Transcription and Cell Proliferation. Cancer Res. 2005, 65, 1352–1360. [Google Scholar] [CrossRef] [Green Version]
- Hu, G.F.; Xu, C.J.; Riordan, J.F. Human Angiogenin Is Rapidly Translocated to the Nucleus of Human Umbilical Vein Endothelial Cells and Binds to DNA. J. Cell. Biochem. 2000, 76, 452–462. [Google Scholar] [CrossRef]
- Yamasaki, S.; Ivanov, P.; Hu, G.-F.; Anderson, P. Angiogenin Cleaves TRNA and Promotes Stress-Induced Translational Repression. J. Cell Biol. 2009, 185, 35–42. [Google Scholar] [CrossRef] [Green Version]
- Russo, N.; Shapiro, R.; Acharya, K.R.; Riordan, J.F.; Vallee, B.L. Role of Glutamine-117 in the Ribonucleolytic Activity of Human Angiogenin. Proc. Natl. Acad. Sci. USA 1994, 91, 2920–2924. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, N.; Nobile, V.; Di Donato, A.; Riordan, J.F.; Vallee, B.L. The C-Terminal Region of Human Angiogenin Has a Dual Role in Enzymatic Activity. Proc. Natl. Acad. Sci. USA 1996, 93, 3243–3247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, H.; Feng, J.; Liu, Q.; Sun, F.; Tie, Y.; Zhu, J.; Xing, R.; Sun, Z.; Zheng, X. Stress Induces TRNA Cleavage by Angiogenin in Mammalian Cells. FEBS Lett. 2009, 583, 437–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ivanov, P.; Emara, M.M.; Villen, J.; Gygi, S.P.; Anderson, P. Angiogenin-Induced TRNA Fragments Inhibit Translation Initiation. Mol. Cell 2011, 43, 613–623. [Google Scholar] [CrossRef] [Green Version]
- Ivanov, P.; O’Day, E.; Emara, M.M.; Wagner, G.; Lieberman, J.; Anderson, P. G-Quadruplex Structures Contribute to the Neuroprotective Effects of Angiogenin-Induced TRNA Fragments. Proc. Natl. Acad. Sci. USA 2014, 111, 18201–18206. [Google Scholar] [CrossRef] [Green Version]
- Fominaya, J.M.; Hofsteenge, J. Inactivation of Ribonuclease Inhibitor by Thiol-Disulfide Exchange. J. Biol. Chem. 1992, 267, 24655–24660. [Google Scholar] [CrossRef]
- Saikia, M.; Krokowski, D.; Guan, B.-J.; Ivanov, P.; Parisien, M.; Hu, G.; Anderson, P.; Pan, T.; Hatzoglou, M. Genome-Wide Identification and Quantitative Analysis of Cleaved TRNA Fragments Induced by Cellular Stress. J. Biol. Chem. 2012, 287, 42708–42725. [Google Scholar] [CrossRef] [Green Version]
- Thiyagarajan, N.; Ferguson, R.; Subramanian, V.; Acharya, K.R. Structural and Molecular Insights into the Mechanism of Action of Human Angiogenin-ALS Variants in Neurons. Nat. Commun. 2012, 3, 1121. [Google Scholar] [CrossRef]
- Bradshaw, W.J.; Rehman, S.; Pham, T.T.K.; Thiyagarajan, N.; Lee, R.L.; Subramanian, V.; Acharya, K.R. Structural Insights into Human Angiogenin Variants Implicated in Parkinson’s Disease and Amyotrophic Lateral Sclerosis. Sci. Rep. 2017, 7, srep41996. [Google Scholar] [CrossRef]
- Lequin, O.; Thüring, H.; Robin, M.; Lallemand, J.Y. Three-Dimensional Solution Structure of Human Angiogenin Determined by 1H,15N-NMR Spectroscopy--Characterization of Histidine Protonation States and PKa Values. Eur. J. Biochem. 1997, 250, 712–726. [Google Scholar] [CrossRef]
- Tsika, A.C.; Chatzileontiadou, D.S.M.; Leonidas, D.D.; Spyroulias, G.A. NMR Study of Met-1 Human Angiogenin: (1)H, (13)C, (15)N Backbone and Side-Chain Resonance Assignment. Biomol. NMR Assign. 2016, 10, 379–383. [Google Scholar] [CrossRef] [PubMed]
- Gellera, C.; Colombrita, C.; Ticozzi, N.; Castellotti, B.; Bragato, C.; Ratti, A.; Taroni, F.; Silani, V. Identification of New ANG Gene Mutations in a Large Cohort of Italian Patients with Amyotrophic Lateral Sclerosis. Neurogenetics 2008, 9, 33–40. [Google Scholar] [CrossRef] [PubMed]
- van Es, M.A.; Schelhaas, H.J.; van Vught, P.W.J.; Ticozzi, N.; Andersen, P.M.; Groen, E.J.N.; Schulte, C.; Blauw, H.M.; Koppers, M.; Diekstra, F.P.; et al. Angiogenin Variants in Parkinson Disease and Amyotrophic Lateral Sclerosis. Ann. Neurol. 2011, 70, 964–973. [Google Scholar] [CrossRef] [PubMed]
- Shapiro, R.; Vallee, B.L. Site-Directed Mutagenesis of Histidine-13 and Histidine-114 of Human Angiogenin. Alanine Derivatives Inhibit Angiogenin-Induced Angiogenesis. Biochemistry 1989, 28, 7401–7408. [Google Scholar] [CrossRef]
- Bennett, M.J.; Schlunegger, M.P.; Eisenberg, D. 3D Domain Swapping: A Mechanism for Oligomer Assembly. Protein Sci. 1995, 4, 2455–2468. [Google Scholar] [CrossRef] [Green Version]
- Libonati, M.; Gotte, G. Oligomerization of Bovine Ribonuclease A: Structural and Functional Features of Its Multimers. Biochem. J. 2004, 380 Pt 2, 311–327. [Google Scholar] [CrossRef] [Green Version]
- Gotte, G.; Laurents, D.V.; Libonati, M. Three-Dimensional Domain-Swapped Oligomers of Ribonuclease A: Identification of a Fifth Tetramer, Pentamers and Hexamers, and Detection of Trace Heptameric, Octameric and Nonameric Species. Biochim. Biophys. Acta 2006, 1764, 44–54. [Google Scholar] [CrossRef]
- Gotte, G.; Laurents, D.V.; Merlino, A.; Picone, D.; Spadaccini, R. Structural and Functional Relationships of Natural and Artificial Dimeric Bovine Ribonucleases: New Scaffolds for Potential Antitumor Drugs. FEBS Lett. 2013, 587, 3601–3608. [Google Scholar] [CrossRef] [Green Version]
- Wishart, D.S.; Sykes, B.D. The 13C Chemical-Shift Index: A Simple Method for the Identification of Protein Secondary Structure Using 13C Chemical-Shift Data. J. Biomol. NMR 1994, 4, 171–180. [Google Scholar] [CrossRef]
- Leonidas, D.D.; Chavali, G.B.; Jardine, A.M.; Li, S.; Shapiro, R.; Acharya, K.R. Binding of Phosphate and Pyrophosphate Ions at the Active Site of Human Angiogenin as Revealed by X-Ray Crystallography. Protein Sci. 2001, 10, 1669–1676. [Google Scholar] [CrossRef]
- Acharya, K.R.; Shapiro, R.; Allen, S.C.; Riordan, J.F.; Vallee, B.L. Crystal Structure of Human Angiogenin Reveals the Structural Basis for Its Functional Divergence from Ribonuclease. Proc. Natl. Acad. Sci. USA 1994, 91, 2915–2919. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorlíčková, M.; Kejnovská, I.; Bednářová, K.; Renčiuk, D.; Kypr, J. Circular Dichroism Spectroscopy of DNA: From Duplexes to Quadruplexes. Chirality 2012, 24, 691–698. [Google Scholar] [CrossRef] [PubMed]
- Del Villar-Guerra, R.; Trent, J.O.; Chaires, J.B. G-Quadruplex Secondary Structure Obtained from Circular Dichroism Spectroscopy. Angew. Chem. 2018, 57, 7171–7175. [Google Scholar] [CrossRef] [PubMed]
- Lane, A.N. The Stability of Intramolecular DNA G-Quadruplexes Compared with Other Macromolecules. Biochimie 2012, 94, 277–286. [Google Scholar] [CrossRef]
- Garavís, M.; López-Méndez, B.; Somoza, A.; Oyarzabal, J.; Dalvit, C.; Villasante, A.; Campos-Olivas, R.; González, C. Discovery of Selective Ligands for Telomeric RNA G-Quadruplexes (TERRA) through 19F-NMR Based Fragment Screening. ACS Chem. Biol. 2014, 9, 1559–1566. [Google Scholar] [CrossRef]
- Patel, D.J. Synthetic RNA and DNA Duplexes. Premelting, Melting and Postmelting Transitions of Alternating Inosine-Cytosine Polynucleotides in Solution. Eur. J. Biochem. 1978, 83, 453–464. [Google Scholar] [CrossRef]
- Schweisguth, D.C.; Moore, P.B. On the Conformation of the Anticodon Loops of Initiator and Elongator Methionine TRNAs. J. Mol. Biol. 1997, 267, 505–519. [Google Scholar] [CrossRef] [Green Version]
- Acharya, K.R.; Shapiro, R.; Riordan, J.F.; Vallee, B.L. Crystal Structure of Bovine Angiogenin at 1.5-A Resolution. Proc. Natl. Acad. Sci. USA 1995, 92, 2949–2953. [Google Scholar] [CrossRef] [Green Version]
- Kövér, K.E.; Bruix, M.; Santoro, J.; Batta, G.; Laurents, D.V.; Rico, M. The Solution Structure and Dynamics of Human Pancreatic Ribonuclease Determined by NMR Spectroscopy Provide Insight into Its Remarkable Biological Activities and Inhibition. J. Mol. Biol. 2008, 379, 953–965. [Google Scholar] [CrossRef]
- Wlodawer, A.; Sjölin, L. Structure of Ribonuclease A: Results of Joint Neutron and X-Ray Refinement at 2.0 a Resolution. Biochemistry 1983, 22, 2720–2728. [Google Scholar] [CrossRef]
- Neira, J.L.; Sevilla, P.; Menéndez, M.; Bruix, M.; Rico, M. Hydrogen Exchange in Ribonuclease A and Ribonuclease S: Evidence for Residual Structure in the Unfolded State under Native Conditions. J. Mol. Biol. 1999, 285, 627–643. [Google Scholar] [CrossRef] [PubMed]
- Laurents, D.V.; Bruix, M.; Jiménez, M.A.; Santoro, J.; Boix, E.; Moussaoui, M.; Nogués, M.V.; Rico, M. The (1)H, (13)C, (15)N Resonance Assignment, Solution Structure, and Residue Level Stability of Eosinophil Cationic Protein/RNase 3 Determined by NMR Spectroscopy. Biopolymers 2009, 91, 1018–1028. [Google Scholar] [CrossRef] [PubMed]
- Fortian, A.; Castaño, D.; Gonzalez, E.; Laín, A.; Falcon-Perez, J.M.; Millet, O. Structural, Thermodynamic, and Mechanistical Studies in Uroporphyrinogen III Synthase: Molecular Basis of Congenital Erythropoietic Porphyria. Adv. Protein Chem. Struct. Biol. 2011, 83, 43–74. [Google Scholar] [CrossRef] [PubMed]
- Pakula, A.A.; Sauer, R.T. Reverse Hydrophobic Effects Relieved by Amino-Acid Substitutions at a Protein Surface. Nature 1990, 344, 363–364. [Google Scholar] [CrossRef] [PubMed]
- Ip, P.; Mulligan, V.K.; Chakrabartty, A. ALS-Causing SOD1 Mutations Promote Production of Copper-Deficient Misfolded Species. J. Mol. Biol. 2011, 409, 839–852. [Google Scholar] [CrossRef]
- Díaz-Casado, L.; Serrano-Chacón, I.; Montalvillo-Jiménez, L.; Corzana, F.; Bastida, A.; González-Santana, A.; González, C.; Asensio, J.L.; Santana, A.G. De Novo Design of Selective Quadruplex-Duplex Junction Ligands and Structural Characterisation of Their Binding Mode: Targeting the G4 Hot-Spot. Chem. Weinh. Bergstr. Ger. 2020. [Google Scholar] [CrossRef]
- Mompeán, M.; Baralle, M.; Buratti, E.; Laurents, D.V. An Amyloid-Like Pathological Conformation of TDP-43 Is Stabilized by Hypercooperative Hydrogen Bonds. Front. Mol. Neurosci. 2016, 9, 125. [Google Scholar] [CrossRef]
- Patel, A.; Lee, H.O.; Jawerth, L.; Maharana, S.; Jahnel, M.; Hein, M.Y.; Stoynov, S.; Mahamid, J.; Saha, S.; Franzmann, T.M.; et al. A Liquid-to-Solid Phase Transition of the ALS Protein FUS Accelerated by Disease Mutation. Cell 2015, 162, 1066–1077. [Google Scholar] [CrossRef] [Green Version]
- Buratti, E.; Baralle, F.E. TDP-43: Gumming up Neurons through Protein-Protein and Protein-RNA Interactions. Trends Biochem. Sci. 2012, 37, 237–247. [Google Scholar] [CrossRef]
- Kim, G.; Gautier, O.; Tassoni-Tsuchida, E.; Ma, X.R.; Gitler, A.D. ALS Genetics: Gains, Losses, and Implications for Future Therapies. Neuron 2020, 108, 822–842. [Google Scholar] [CrossRef]
- Papageorgiou, A.C.; Shapiro, R.; Acharya, K.R. Molecular Recognition of Human Angiogenin by Placental Ribonuclease Inhibitor--an X-ray Crystallographic Study at 2.0 aResolution. EMBO J. 1997, 16, 5162–5177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chavali, G.B.; Papageorgiou, A.C.; Olson, K.A.; Fett, J.W.; Hu, G.F.; Shapiro, R.; Acharya, K.R. The Crystal Structure of Human Angiogenin in Complex with an Antitumor Neutralizing Antibody. Struct. Lond. Engl. 1993 2003, 11, 875–885. [Google Scholar] [CrossRef]
- Notomista, E.; Cafaro, V.; Fusiello, R.; Bracale, A.; D’Alessio, G.; Di Donato, A. Effective Expression and Purification of Recombinant Onconase, an Antitumor Protein. FEBS Lett. 1999, 463, 211–215. [Google Scholar] [CrossRef] [Green Version]
- Lee, W.; Tonelli, M.; Markley, J.L. NMRFAM-SPARKY: Enhanced Software for Biomolecular NMR Spectroscopy. Bioinf. Oxf. Engl. 2015, 31, 1325–1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kjaergaard, M.; Brander, S.; Poulsen, F.M. Random Coil Chemical Shift for Intrinsically Disordered Proteins: Effects of Temperature and PH. J. Biomol. NMR 2011, 49, 139–149. [Google Scholar] [CrossRef]
- Vuister, G.W.; Delaglio, F.; Bax, A. The Use of 1JC Alpha H Alpha Coupling Constants as a Probe for Protein Backbone Conformation. J. Biomol. NMR 1993, 3, 67–80. [Google Scholar] [CrossRef]
- Farrow, N.A.; Muhandiram, R.; Singer, A.U.; Pascal, S.M.; Kay, C.M.; Gish, G.; Shoelson, S.E.; Pawson, T.; Forman-Kay, J.D.; Kay, L.E. Backbone Dynamics of a Free and Phosphopeptide-Complexed Src Homology 2 Domain Studied by 15N NMR Relaxation. Biochemistry 1994, 33, 5984–6003. [Google Scholar] [CrossRef]
- Bai, Y.; Milne, J.S.; Mayne, L.; Englander, S.W. Primary Structure Effects on Peptide Group Hydrogen Exchange. Proteins 1993, 17, 75–86. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-Z. Protein and Peptide Structure and Interactions Studied by Hydrogen Exchanger and NMR. Ph.D. Thesis, University of Pennsylvannia, Philadelphia, PA, USA, 1995. [Google Scholar]
- Pantoja-Uceda, D.; Santoro, J. Amino Acid Type Identification in NMR Spectra of Proteins via Beta- and Gamma-Carbon Edited Experiments. J. Magn. Reson. 2008, 195, 187–195. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fagagnini, A.; Garavís, M.; Gómez-Pinto, I.; Fasoli, S.; Gotte, G.; Laurents, D.V. NMR Characterization of Angiogenin Variants and tRNAAla Products Impacting Aberrant Protein Oligomerization. Int. J. Mol. Sci. 2021, 22, 1439. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031439
Fagagnini A, Garavís M, Gómez-Pinto I, Fasoli S, Gotte G, Laurents DV. NMR Characterization of Angiogenin Variants and tRNAAla Products Impacting Aberrant Protein Oligomerization. International Journal of Molecular Sciences. 2021; 22(3):1439. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031439
Chicago/Turabian StyleFagagnini, Andrea, Miguel Garavís, Irene Gómez-Pinto, Sabrina Fasoli, Giovanni Gotte, and Douglas V. Laurents. 2021. "NMR Characterization of Angiogenin Variants and tRNAAla Products Impacting Aberrant Protein Oligomerization" International Journal of Molecular Sciences 22, no. 3: 1439. https://0-doi-org.brum.beds.ac.uk/10.3390/ijms22031439